

Abstract
Background
Direct-acting antivirals (DAAs) against hepatitis C virus (HCV) are potent and highly efficacious. However, resistance-associated substitutions (RASs) relevant to DAAs can impair treatment effectiveness even at baseline. Moreover, the prevalence of baseline RASs in HCV genotype 1b-infected patients in western China is still unclear.
Materials and methods
Direct sequencing of the HCV NS3, NS5A, and NS5B regions was performed in baseline serum samples of 70 DAAs treatment-naïve HCV 1b-infected patients in western China. The sequences were analyzed with MEGA version 5.05 software. Evolutionary patterns of RASs and amino-acid covariance patterns in the NS3, NS5A, and NS5B genes were analyzed by MEGA and Cytoscape (version 3.2.1), respectively.
Results
The presence of at least one RAS in the NS3 region (C16S, T54S, Q80R/L, A87T, R117H, S122G, V132I, V170I) was observed in 85.48% (53 of 62) of patients, RASs in the NS5A region (L28M, R30Q, Q54H, P58S/T, Q62H/R, Y93H) were observed in 42.42% (28 of 66) of patients, and RASs in the NS5B region (N142S, A300T, C316N, A338V, S365A, L392I, M414L, I424V, A442T, V499A, S556G) were observed in 100% (44 of 44) of patients. Evolutionary patterns of RASs and amino-acid covariance patterns for the NS3, NS5A, and NS5B genes are reported.
Conclusion
The prevalence of RASs relevant to DAAs detected in the NS3, NS5A, and NS5B regions of HCV 1b from DAA treatment-naïve patients is high. Therefore, more attention should be paid to RASs associated with DAAs in the upcoming DAA-treatment era in China.
Keywords: hepatitis C virus, unstructured proteins, resistance-associated substitutions, direct-acting antiviral agents
Introduction
Hepatitis C virus (HCV) infection has greatly increased the burden of global disease.1 The recent estimates of global disease burden show that the seroprevalence of HCV is 2.5%, equating to >177.5 million infections.2 HCV is associated with liver cirrhosis, hepatocellular cancer, and liver failure, which has become an important cause of HCV-related deaths, resulting in 350,000 cases of patient death per year worldwide.3,4 As HCV is a RNA virus, its genome is highly variable, and the sequences are classified into seven recognized genotypes (GTs) and a large number of subtypes.5 HCV sequences belonging to different GTs show a 30%–35% difference at the nucleotide level.5 GT1 is estimated to account for more HCV cases than any other GTs, at 83.4 million cases (46.2%), and more than a third of GT1 infections occur in East Asia.6 Additionally, 56.8% of HCV infections in China are of subtype 1b.7
Pegylated-interferon and ribavirin have been the standard of care for HCV-infected patients. However, severe side effects of this treatment lead to about 50% of patients being unable to tolerate therapy, and the sustained virological response rate after this treatment is only approximately 50% for HCV GT1 infections.8 Remarkably, direct-acting antivirals (DAAs) with the ability directly to inhibit the HCV unstructured proteins NS3/4A protease, NS5A protein, and NS5B polymerase, which are important for HCV replication in hepatocytes, have resulted in very high sustained virological response rates of more than 90% in Phase II and Phase III studies after short-term treatment.9 However, due to the low fidelity of HCV polymerase, high HCV-replication rate, and strong selective pressures on the virus, multiple variants differing by up to a few percent exist even within individual isolates, ie, HCV replicates as a quasispecies.10,11 Some quasispecies may carry the baseline resistance-associated substitutions (RASs), which convey varying degrees of resistance to DAAs. Indeed, RASs have been observed in both in vitro and clinical trials.12 In particular, the clinically relevant baseline RASs that reduce susceptibility to DAAs and play a role in treatment failure in postmarketing studies or clinical practice play a potent role in treatment failure.13,14 DAAs have not been approved in China; however, this approval is imminent. Therefore, it is necessary to have an understanding of the prevalence of RASs in treatment-naïve HCV GT 1b-infected patients in western China.
Materials and methods
Patients
Serum samples were obtained from 70 patients who had been chronically infected with HCV 1b and never been treated for HCV with any DAAs. Patients were recruited between 2015 and 2016 from the Second Affiliated Hospital of Chongqing Medical University and were diagnosed according to the 2016 European Association for the Study of the Liver recommendations on treatment of HCV.15 HCV GT and subtype were determined by reverse-transcription polymerase chain reaction (PCR) and sequencing of the HCV core region. Patients enrolled in the study were not coinfected with HBV or HIV, and all were from the Chinese Han population. The study was approved by the ethics committee of the Second Affiliated Hospital of Chongqing Medical University, and written informed consent was obtained from each study participant.
Amplification and sequencing of HCV NS3, NS5A, and NS5B genes
Viral RNA was extracted from 140 μL serum samples using the MiniBest viral RNA extraction kit (Takara, Kusatsu, Japan). cDNA was synthesized from HCV RNA using a PrimeScript RT reagent kit with genomic DNA eraser (Takara) following the manufacturer’s instructions. Cycling conditions were incubation at 42°C for 2 minutes to remove genomic DNA, followed by 37°C for 15 minutes to synthesize cDNA. Full-length HCV NS3, NS5A, and NS5B genes were amplified using PrimeStar HS (Takara) in a nested PCR. Primers are shown in Table S1. PCR products in the first PCR round were obtained using a predenaturation step at 95°C for 5 minutes, followed by 35 cycles of denaturation at 98°C for 10 seconds, annealing at 62°C for 20 seconds, and extension at 72°C for 1 minutes and 20 seconds for the NS3 and NS5A genes and 35 cycles of denaturation at 98°C for 10 seconds, annealing at 56°C for 20 seconds, and extension at 72°C for 2 minutes for the NS5B gene. For all genes, these steps were followed by a final 10-minute extension step at 72°C. PCR volume contained 25 μL of 2× reaction premix, 1 μL each primer, 5 μL cDNA template, and nuclease-free H2O for a final volume of 50 μL. Three microliters from the first round PCR product was used as a template in the second round of PCR, which was done with a similar reaction volume and conditions as the first round, but with an annealing-temperature increase to 64°C for the NS3 and NS5A genes and to 60°C for the NS5B gene.
Direct sequencing of nested PCR products was performed using an automatic sequencer (ABI Prism 3100; Thermo Fisher Scientific, Waltham, MA, USA). Sequencing primers were the second-round PCR primers for each gene. Correction of the base sequences was performed with reference to direct-sequencing chromatograms. Representative chromatograms from the direct sequencing are shown in Figure S1.
Sequence analysis and phylogeny
Clinically relevant RASs to DAAs were summarized from the literature and are presented in Table S2. The NS3, NS5A, and NS5B nucleotide sequences were aligned with the HCV 1b reference sequence (GenBank accession AJ238799) and analyzed using BioEdit 7.0.9 software. A variant type was indicated by replacement of the consensus amino acid with a novel one, eg, Y93H and Y93N in the NS5A region were described as two variant types.
A phylogenetic tree of each gene was created using the maximum-likelihood statistical method with the Jones–Taylor–Thornton two-parameter model in MEGA version 5.05.16 A γ-parameter of 0.5 was used to model differences in substitution rates among base sites. The reliability of the cluster descending from that node was assessed by a bootstrap analysis with 1,000 replicates.
Structural bioinformatic modeling and amino-acid covariance analysis
Protein-homology modeling was performed with Swiss-Model (https://swissmodel.expasy.org), with an identity of more than 90% used to model the X-ray crystal structures of HCV NS3 protease, NS5A, protein and NS5B polymerase for GT 1b. Structural editing and analysis were done using PyMol (Schrödinger, New York, NY, USA). The three structures were loaded into PyMol to analyze the cartoon structures, the stick structures of RASs, and the drug-binding pocket.
Sequences consisting of the NS3, NS5A, and NS5B genes were aligned and translated into amino acid sequences using BioEdit 7.09. Then, all possible amino-acid covariant pairs were determined by employing the observed-minus-expected-squared (OMES) algorithm, which can exclude invariant positions and spurious correlations generated by random differences, as described in other work.17 To identify the covarying pairs, we calculated an OMES score for every possible pair. An OMES score of 0.5 was used as the cutoff for selecting covariant amino-acid pairs for analyses.18 Finally, amino-acid covariant networks were visualized by Cytoscape (version 3.2.1).19
Statistical analysis
Concentration trends of data were expressed as medians (minimum–maximum). Categorical variables were summarized as numbers. Categorical variables were compared between groups using the χ2 test. Continuous data were compared between groups using Student’s t-test. P<0.05 was considered statistically significant. Statistical analyses were performed using SPSS 17 software (SPSS, Chicago, IL, USA).
Results
Characteristics of patients
For all 70 patients, the NS3, NS5A, and NS5B genes were successfully amplified and sequenced in 62, 66, and 44 patients, respectively. In total, 66 patients were included for further analysis in this study. The clinical and virological characteristics of these patients are shown in Table 1.
Table 1.
Baseline characteristics of patients with HCV 1b infection in western China
| Characteristics | Values |
|---|---|
| Age (years), median (range) | 47 (18–78) |
| Male (female) | 36 (30) |
| Alb (g/L), median (range) | 44.4 (16.8–54.9) |
| ALT (IU/L), median (range) | 62 (5–230) |
| AST (IU/L), median (range) | 49.5 (14–586) |
| γ-GGT (IU/L), median (range) | 53 (9–1,955) |
| Total bilirubin (mg/dL), median (range) | 0.72 (0.37–3.52) |
| Direct bilirubin (mg/dL), median (range) | 0.34 (0.16–2.68) |
| Plt count (×109/L), median (range) | 174 (56–315) |
| AST/Plt, median (range) | 0.31 (0.07–7.23) |
| Hemoglobin (g/dL), median (range) | 149 (77–178) |
| HCV RNA (log10 IU/mL), median (range) | 6.39 (3.00–7.73) |
Abbreviations: HCV, hepatitis C virus; Alb, albumin; Plt, platelet.
Prevalence of RASs relevant to NS3 protease inhibitors
In total, RASs conferring resistance to NS3 protease inhibitors (PIs) were detected in 53 of 62 (85.48%) sequences, as shown in Figure 1A. Among these, S122G (resistance to simeprevir/asunaprevir) was the most common, observed in 85.48% of sequences, followed by V170I-associated resistance to telaprevir (25.81%) and T54S-associated resistance to telaprevir and boceprevir (14.52%). The other RASs were observed at frequencies lower than 10%, such as Q80R/L (resistance to simeprevir, faldaprevir, and asunaprevir), V132I and R117H (resistance to boceprevir), C16S (resistance to telaprevir), and A87T (resistance to telaprevir/boceprevir). No RASs showing high-level resistance to DAAs were detected. It is noteworthy that R155, V156, and D168, which are important variants resistant to PIs, were not detected in this study. In addition, some substitutions like Y56F, R123K, and S174A were also detected in this study.
Figure 1.

Prevalence of clinically relevant RASs in various regions against different DAA classes.
Notes: Frequency of clinically relevant RASs in the NS3 region (A), NS5A region (B), and NS5B region (C). Total frequency of clinically relevant RASs against different DAA classes (D).
Abbreviations: RASs, resistance-associated substitutions; DAA, direct-acting antivirals.
Moreover, among the 62 patients, 25 (40.32%) had a mixture of variants that contained multiple NS3-resistance variants. In detail, 18 (29.03%) patients carried two RAS mixtures, such as V170I + S122G or T54S + S122G, and five (8.06%) patients carried three RAS mixtures, such as Q80R + T54S + S122G or Q80R + S122G + V170I. Importantly, two mixtures of four RASs (Q80R + T54S + V170I + S122G and C16S + R117H + V170I + S122G) were also each detected in one patient.
Prevalence of RASs relevant to NS5A inhibitors
In total, these RASs were detected in 36 of 66 (54.55%) sequences, as shown in Figure 1B. In detail, the most commonly observed RAS was Q54H (resistance to daclatasvir, 42.42%). The next-most common RASs, P58S/T (resistance to daclatasvir) and Q62H/R (resistance to daclatasvir), were detected in 6.06% and 9.09% of sequences, respectively. Lastly, L28M (resistance to daclatasvir/ombitasvir) and R30Q (resistance to daclatasvir) were identified in only one patient each. Notably, the key RAS Y93H/N, which is associated with resistance to daclatasvir/ledipasvir/ombitasvir, was detected in 7.58% of sequences and associated with a high level of resistance. But another key RAS, L31V/M, associated with resistance to daclatasvir/ledipasvir, was not observed in this study. In addition, five different mixtures of RASs were observed (Q54H + P58S, Q54H + Q62H, Q62H + Y93H, Q54H + Y93H, and R30Q + Q54H + P58S + Q62R) in six patients.
Prevalence of RASs relevant to NS5B polymerase inhibitors
The prevalence of RASs associated with resistance to NS5B nucleotide analogue inhibitors (NIs) and non-NIs (NNIs) was also differentially distributed, as shown in Figure 1C. Among RASs relevant to NIs, key RASs for sofosbuvir and mericitabine correlate with variants at positions S282 and L159. In this study, no sequence containing L159F and S282T/R was found. However, the prevalence values of N142S (resistance to sofosbuvir) and A300T (resistance to R7128) were 4.55% and 6.82%, respectively. Compared to RASs relevant to NIs, RASs associated with resistance to NNIs were more commonly observed. Remarkably, C316N (resistance to dasabuvir) was found in all sequences in this study. In addition, A338V (resistance to beclabuvir) and I424V (resistance to beclabuvir) occurred in 47.73% and 22.73% of sequences, respectively. S365A (resistance to nesbuvir), L392I (resistance to beclabuvir), and A442T (resistance to beclabuvir) represented a minority of sequences, with frequencies of 2.27%, 6.82%, and 2.27%, respectively. Notably, the key RASs M414L (resistance to dasabuvir) and S556G (resistance to dasabuvir) were present in 4.55% and 2.27% of sequences, respectively. The RAS V499A (resistance to beclabuvir) was presented in 6.82% of sequences. There were also some substitutions detected, such as S368F, A421E, and C451Y. In addition, 37 of 44 (84.09%) sequences contained more than one RAS: 26 (59.09%) patients had two RAS mixtures, such as C316N + A338V, C316N + I424V, and C316N + A300T. Eleven (25%) patients had mixtures of three or more RASs, such as C316N + S556G + A338V and C316N + S556N + V499A.
Prevalence of multiple RASs relevant to different DAA classes
Overall, the frequency of multiple RASs associated with resistance to different DAA classes was high, and varied for different RAS combinations (52.46%–97.62%, Figure 1D). The highest prevalence of multiple RASs was in the NS3 + NS5B regions, followed by the NS5A + NS5B, NS3 + NS5B, and NS3 + NS5A + NS5B regions. C316N was observed in all NS5B sequences, which may contribute to its high prevalence. We analyzed the results again after excluding C316N. Similar results at a lower frequency were observed, as presented in Figure S2.
Evolutionary patterns of RASs in NS3, NS5A, and NS5B genes
To study whether RASs occurring at baseline were correlated with ongoing random variation and/or some specific evolutionary patterns in transmission clusters, phylogenetic trees of the NS3, NS5A, and NS5B genes were created, as described in the Materials and methods section. The results showed that most baseline RASs in the NS3 and NS5B genes seemed to occur randomly, with no special evolutionary pattern occurring, as observed in Figure S3. However, a clade without RASs in the NS5A gene showed a prominent concentration trend, as shown in Figure 2. Therefore, clinical characteristic analyses were performed in patients with and without RASs in the NS5A gene to investigate the possible reason for this phenomenon. However, there were no significant differences between the two groups, as shown in Table S3.
Figure 2.
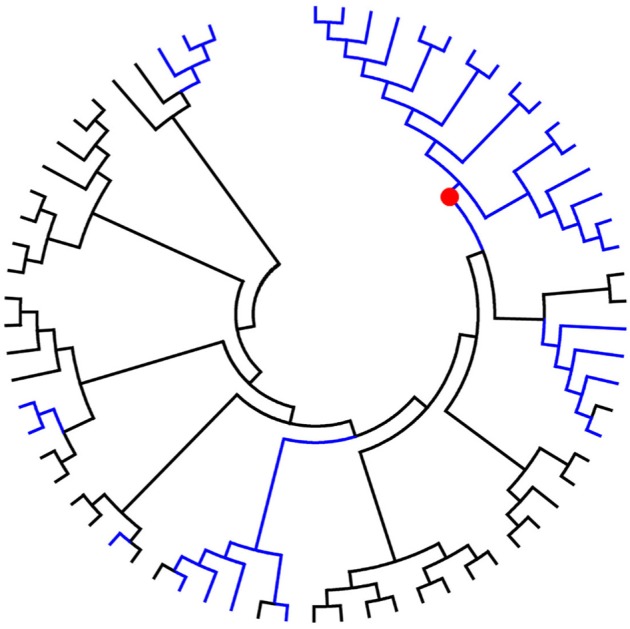
Phylogenetic tree for the hepatitis C 1b NS5A gene.
Notes: Black lines represent unresistant lineages; blue lines represent lineages carrying the resistance mutation; red dots represent the emergence of concentration-trend clades of unresistant isolates.
Structural bioinformatic modeling and amino-acid covariance patterns of NS3, NS5A, and NS5B genes
In order to investigate the structures of the different viral unstructured proteins and reveal the mutations and drug-binding pockets, protein-structure bioinformatic modeling was performed on the HCV NS3, NS5A, and NS5B proteins. The HCV NS3 protease crystal structure and the stick structures of such RASs as R117I, Q80R, and S122G are shown in Figure 3A. The NS5A protein-crystal structure and the stick structures of RASs Q54H, Q62H, Y93H, and P58S are presented in Figure 3B. The NS5B polymerase-crystal structure and stick structures of RASs A338V, A300T, N142S, C316N, and M414L are shown in Figure 3C. Drug-binding pockets in different viral proteins are also shown in Figure 3.
Figure 3.

Structures and drug-binding pockets in different HCV unstructured proteins.
Notes: Crystal structures and stick views of RASs in the drug-binding pockets of the HCV NS3 protease (A), NS5A protein (B), and NS5B polymerase (C). Color of crystal structures determined by secondary structure of proteins, with α-helices colored red, β-sheets colored yellow, random coils and other structures colored green for NS3 and NS5B, α-helices colored yellow, and β-sheets colored red in NS5A. The stick structures of RASs are identified by color and amino acid sites.
Abbreviations: HCV, hepatitis C virus; RASs, resistance-associated substitutions.
Next, to understand the covariance relationship among the different regions, a covariance-network analysis was undertaken. As shown in Figure 4A, there were a considerable number of amino-acid covariance positions (nodes) and interacting lines (edges), constituting a huge covariance network. These nodes included both RASs and non-RASs. Most of them were found in the NS5B region, such as R120, M414, and V322. Further analysis focused on the RAS-covariance relationships only, and a similar but smaller network was observed, as shown in Figure 4B. RASs at positions R117 in the NS3 region, R30 in the NS5A region, and N142, L392, M414, I424, and V499 in the NS5B region interacted with one another.
Figure 4.

Amino-acid covariance networks for NS3, NS5A, and NS5B sequences in HCV 1b.
Notes: Amino-acid covariance networks for all variant positions (A) and clinically relevant resistance-associated substitutions (RASs) (B). The size of the nodes is determined by the number of edges that they contact. Red nodes are within the NS3 region, green nodes in the NS5A region, and blue nodes in the NS5B region. White font indicates non-RASs, and black font indicates RASs. Numbered amino acid residue position is provided relative to the first amino acid of the NS3, NS5A, or NS5B region.
Discussion
In this study, clinically relevant RASs associated with resistance to DAAs were frequently observed in treatment-naïve HCV 1b-infected patients in western China. RASs in the NS5B region were the most frequently observed (the C316N variant was detected in 100% of sequences), followed by those in the NS3 region and NS5A region (85.48% and 42.42%, respectively).
In the NS3 region, S122G, which has been confirmed to provide a low level of resistance, was the most commonly observed RAS. S122G is a major RAS relevant to simeprevir and asunaprevir. The occurrence of this RAS in this study was much higher than that observed in an open-label trial in Japan (85.48% vs 36.7%).20 The higher number of patients included in their study may be a reason for this discrepancy. Another key RAS, Q80K, was observed in none of the 62 HCV 1b sequences, while 6.45% of sequences were detected to contain Q80R/L, which is commonly observed in GT 1b-infected patients in whom treatment has failed.21 In addition, the low genetic barrier to resistance and considerable cross-resistance have been implicated as being involved in RASs at positions V36, T54, R155, and A156.12 The first-generation PIs boceprevir and telaprevir are not recommend for HCV treatment,15 but in our study only the T54S variant was found, in 14.52% of sequences. Substitutions at positions R155 and D168, which have been confirmed to reduce the activity of simeprevir, asunaprevir, paritaprevir, and vanipre-vir, were not found in this study either.12,22
Current NS5A inhibitors are characterized by broad genotypic coverage; however, low barriers to resistance make RASs against NS5A inhibitors of wide concern.23 The prevalence of RASs relevant to NS5A inhibitors in this study was higher than that reported by a previous study (42.42% vs 29.6%).24 This discrepancy might be because more clinically relevant RASs were identified in this study than in prior work. RASs at positions 28, 30, 31, and 93 have been frequently observed in both laboratory and clinical trials,25 but in this study the most commonly occurring variant was Q54H, which confers low-level resistance to daclatasvir.26 Furthermore, L31M and Y93H are regarded as key resistance variants, and the simultaneous presence of these two substitutions at baseline will significantly affect the outcome of NS5A-based antiviral therapy.27 The prevalence of Y93H, which confers high-level resistance to DAAs, in this study was 7.58%, which is similar to findings from other reports (3.8%–14.1%).27,28 Nevertheless, none of the sequences contained L31M, which confers a low–medium level of resistance to daclatasvir and ledipasvir, and this finding is different from previously reported frequencies (2.1%–6.3%).28 This discrepancy may be a result of differences in sensitivity between population sequencing and ultradeep sequencing for detection of viral populations. Although no Y93H + L31M variants were found in this study, other concomitant variants, such as R30Q and P58S, should be given more attention. These two variants are regarded as secondary-resistance variants, and their presence will enhance the resistance conferred by primary variants.29 In addition, the variant combination of L28M and L31F has been shown to have a higher level of resistance to ombitasvir than either of the variants separately did.30 In this study, L28M was detected without the presence of L31F.
NS5B polymerase inhibitors, including the NNI dasabuvir and the NI sofosbuvir, are potent agents with a high resistance barrier.31 Additionally, no cross-resistance has been observed with currently approved NS5B inhibitors, with one exception, C316N, which is associated with low-level resistance to dasabuvir and sofosbuvir.32,33 Baseline polymorphisms at position 316 vary by geographical location and subtype. C316 is highly conserved in GT 1a (99.89%), but polymorphic in GT 1b (81.83%), based on frequencies.32 In addition, HCV 1b isolates harboring C316N at baseline are more frequently observed in Asia compared to Europe and the US (91.6% vs 32% vs 5%, respectively).34 Remarkably, the polymorphic C316N was detected in 100% of the 44 sequences in this study, in agreement with the results of another study in China.35 This indicated that C316N might confer enhanced fitness for chronic HCV 1b patients in China and that more attention should be paid to this substitution in Chinese HCV 1b patients. The other key RASs that may lead to failure of sofosbuvir treatment are L159F, S282T, L320F, V321A, and L159F + V321A, which are observed in 2.2%–4.4% of subjects.32 However, none of these RASs was detected in this study, and it may be that mutations in these positions will greatly influence viral fitness. Moreover, M414L and S556G, which can lead to the treatment failure of dasabuvir, were commonly observed in this study, similarly to the results of previous studies.12
In this study, we found that a clade without RASs in the NS5A gene showed a more prominent concentration trend than others. However, we did not find any factors that were responsible for this observation. A larger-population study may be needed to clarify this phenomenon. In addition, the covariance relationships between RAS and non-RAS nodes or between RASs indicated a complex interacting relationship in the HCV genome at baseline. The RASs in the NS3 region, the NA5A region, and the NS5B region affected one another, which may indicate that an RAS found in one of the three regions affects RASs in the other two regions. Therefore, this finding indicates that more attention should be paid to multisegment combined drug resistance when using DAAs in future.
To our knowledge, this study is the first to report the prevalence of RASs in western Chinese chronic HCV 1b patients. Moreover, the latest and most comprehensive panel of clinically relevant RASs was included to analyze all DAA-resistance regions (the NS3, NS5A, and NS5B regions). However, there are several limitations to this study. First, the number of patients was limited, which may have led to relatively high prevalence. Second, some NS5B genes were unsuccessfully amplified, which reduced the sample size of this study. Third, direct sequencing, not ultradeep pyrosequencing, was used in this study, which hindered our ability to investigate the prevalence of minority variants (<20%), though RASs detected in this study represent the main variants.
In conclusion, the prevalence of RASs clinically relevant to DAAs was high in treatment-naïve HCV 1b-infected patients in western China, especially for variants S122G, Q54H, and C316N. Further studies are needed to clarify the role of these RASs so that we can improve our strategy for managing and using DAAs in China in future.
Supplementary material
Chromatograms of Sanger sequencing.
Notes: Chromatograms of Sanger sequencing in part of NS3 segment with 80Q (A); with 80R (B). Amino acid site 80 occurring bases substitution leading to amino acid Q change to R which resistance to DAAs, thus, Q80R was described as a variant type.
The prevalence of clinically relevant resistance associated substitutions (without C316N) in various DAA classes.
Abbreviation: RASs, resistance-associated substitutions.
Phylogenetic trees for HCVGT1b NS3 and NS5B genes.
Notes: (A) Phylogenetic tree for HCV GT1b NS3 gene; (B) phylogenetic tree for HCV GT1b NS5B gene. Blue lines represent nonresistant lineages; black lines represent lineages carrying the resistance associated substitutions.
Table S1.
| Target gene | Primers | 5′–3′ sequences |
|---|---|---|
| NS3 | NS3F out | CGAGACCTTGCGGTGGCAG |
| NS3R out | CAGCCGTYTCCGCTTGGTCC | |
| NS3F in | CATCACCTGGGGGGCAGACACC | |
| NS3F in | GTCAGTTGAGTGGCACTCATCAC | |
| NS5A | NS5AF out | GTTTGGGACTGGATATGC |
| NS5AR out | CGTCACGTAGTGGAAATC | |
| NS5AF in | ACCTGGCTCCAGTCCAAG | |
| NS5AF in | CCTCCACRTACTCCTCAG | |
| NS5B | NS5BF out | CGYTGAGTCRTAYTCCTCCATGC |
| NS5BR out | GGGCRCGAGACASGCTGTGATA | |
| NS5BF in | CTCAGYGACGGGTCYTGGTC | |
| NS5BR in | GCTGTGATATATGTCTCCCC |
Abbreviation: HCV, hepatitis C virus.
Table S2.
Summary of HCV RASs to DAAs
| Region | Residue | RAS(s) | DAA(s) | Reference(s) |
|---|---|---|---|---|
| NS3 | C16 | S | Tvr | 4 |
| V36 | A/M/L/G/I/C | Tvr, Boc, Asv, Ptv, Fdv | 5–9 | |
| Q41 | R | Smv | 10, 11 | |
| F43 | S/V/I/L | Boc, Tvr, Ptv, Smv | 6, 10, 12 | |
| T54 | A, S, C, G, V | Tvr, Boc, Fdv, Asv | 5, 9, 10, 12 | |
| V55 | A | Boc, Tvr | 6, 9 | |
| F, I, K, T | Boc, Asv | 1, 13 | ||
| Y56 | H | Ptv, Asv | 5 | |
| L | Asv | 5, 10 | ||
| Q80 | K, R, H, N, G, L | Smv, Asv/Ptv | 1, 5, 6, 10 | |
| A87 | T | Tvr, Boc | 14 | |
| V107 | I | Boc | 5 | |
| R109 | K | Smv, Boc | 15 | |
| R117 | H | Boc | 14 | |
| S122 | A, I, R, T, G, N | Smv, Asv | 5, 9, 16 | |
| R123 | T | Tvr, Boc | 17 | |
| V132I | I | Tvr | 18 | |
| R155 | K/G/T/M/I/L/S/Q/P/N/W | Tvr, Boc, Smv, Vnv, Fdv, Asv, Ptv | 5, 6, 9, 10, 19, 20 | |
| A156 | S/T/F/N/V/I/G/D | Tvr, Boc, Smv, Vnv, Fdv, Asv, Gzv | 5, 6, 10, 19, 20 | |
| V158 | I, M | Boc, Tvr | 5, 6 | |
| D168 | Q/A/Y/V/E/T/N/P/I/H/G/F/S/K | Smv, Vnv, Asv, Fdv, Ptv, Gzv | 5, 6, 9, 10, 13, 21, 22 | |
| V170 | A | Boc, Tvr | 6 | |
| T | Boc, Smv | 5 | ||
| I | Tvr | 18 | ||
| S174 | F/P | Tvr | 14 | |
| M175 | L | Boc | 5 | |
| NS5A | L28 | F, M, T | Dcv, Omv | 5, 6, 18, 23, 24 |
| P29 | S, X | Dcv | 5 | |
| R30 | G, H, P, Q | Dcv | 5, 10 | |
| L31 | V/M/F/I | Dcv, Omv, Ldv | 5, 6, 9 | |
| P32 | L, S, X | Dcv, Ldv | 5, 6, 10 | |
| Q54 | H/N/L/Y | Dcv, Omv | 18, 23, 24 | |
| P58 | S/A/H/T | Dcv | 5, 24 | |
| D | Ldv | 6 | ||
| Q/E62 | D/H | Dcv | 5, 18 | |
| R/A/P/S | Dcv | 24, 25 | ||
| A92 | K/T | Dcv | 5, 6, 24 | |
| Y93 | H/N/C | Dcv, Ldv, Omv | 5, 6, 23, 24, 26 | |
| NS5B | H95 | Q/R | Dsv | 18, 27 |
| L159 | F | Sof, Mcb | 5, 28, 29 | |
| N142 | T/S | R1479, Sof | 27, 30 | |
| S282 | T/R | Sof, Mcb | 5, 6, 30 | |
| M289 | I/L/V | Sof | 3, 6 | |
| A300 | T | R7128 | 31 | |
| V321 | A | Sof | 5 | |
| C316 | H, N, Y, S | Dsv, Nbv, Sof | 5, 6, 9 | |
| L320 | F, I, S | Sof, Mcb, PSI938 | 6, 28–30 | |
| V321 | A | Sof | 5 | |
| A338 | V | Bcv | 18, 32 | |
| S365 | A, L, T | Nbv | 18, 32, 33 | |
| S368 | T | Dsv | 6 | |
| L392 | I | GS9669 | 18 | |
| N411 | S | Dsv | 6 | |
| M414 | L/T/I/V/Q | Dsv, A782759, Nbv | 5, 6, 13 | |
| A421 | V | BMS791325 | 5, 13 | |
| M423 | I, T | Nbv | 13, 34 | |
| I424 | V | Bcv | 35 | |
| M426 | T/V/I | A782759, Nbv | 3, 34 | |
| A442 | T | Bcv | 5, 18, 19 | |
| C445 | F, | GS9190 | 15, 36 | |
| E446 | K, Q | Dsv | 6, 9 | |
| Y448 | C, H | Dsv, GS9190 | 6, 37, 38 | |
| V494 | A, T, L, I | Nbv | 13, 15 | |
| P495 | A, L, S | Bcv, BI207127 | 5, 27, 36, 39 | |
| V499 | A | Bcv | 18, 40 | |
| A553 | I, T, V | Dsv | 6, 10 | |
| G554 | S, D | Dsv | 6, 10 | |
| S556 | G, N, R, C | Dsv | 5, 6, 13, 41 | |
| G558 | R | Dsv | 6 | |
| D559 | G, S, N | Dsv | 6, 9 | |
| Y561 | H | Dsv | 6 |
Abbreviations: HCV, hepatitis C virus; RASs, resistance-associated substitutions; Asv, asunaprevir; Boc, boceprevir; Ptv, paritaprevir; Smv, simeprevir; Tvr, telaprevir; Fdv, faldaprevir; Vnv, vaniprevir; Gzv, grazoprevir; Dcv, daclatasvir; Ldv, ledipasvir; Omv, ombitasvir; Sof, sofosbuvir; Dsv, dasabuvir; Bcv, beclabuvir; Mcb, mericitabine; Nbv, nesbuvir.
Table S3.
Baseline characteristics of patients with and without NS5A RASs
| Characteristics | Wild type (n=36) | Mutation type (n=16) | P |
|---|---|---|---|
| Age (years), median (range) | 50 (18–70) | 42 (35–58) | 0.192 |
| Male (female) | 21 (15) | 7 (9) | >0.05 |
| Alb (g/L), median (range) | 44.35 (36.4–54.9) | 45.25 (24.6–48) | 0.513 |
| ALT (U/L), median (range) | 61 (10–189) | 66 (20–230) | 0.505 |
| AST (U/L), median (range) | 54 (14–163) | 47 (21–199) | 0.657 |
| γ-GGT (IU/L), median (range) | 57 (15–207) | 33 (9–97) | 0.054 |
| Total bilirubin (mg/dL), median (range) | 0.85 (0.44–1.91) | 0.63 (0.37–1.25) | 0.597 |
| Direct bilirubin (mg/dL), median (range) | 0.34 (0.22–0.79) | 0.33 (0.16–0.90) | 0.746 |
| Plt count (×109/L), median (range) | 153 (57–240) | 184 (56–315) | 0.323 |
| AST/Plt, median (range) | 0.43 (0.12–1.63) | 0.26 (0.1–1.80) | 0.867 |
| Hemoglobin (g/dL), median (range) | 153 (85–171) | 142.5 (77.4–178) | 0.907 |
| HCV RNA (log10 IU/mL), median (range) | 6.35 (3.00–7.5) | 6.57 (4.37–7.23) | 0.444 |
Abbreviations: HCV, hepatitis C virus; RASs, resistance-associated substitutions; Alb, albumin; Plt, platelet.
References
- 1.Paolucci S, Fiorina L, Piralla A, et al. Naturally occurring mutations to HCV protease inhibitors in treatment-naïve patients. Virol J. 2012;9:245. doi: 10.1186/1743-422X-9-245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Dietz J, Susser S, Berkowski C, Perner D, Zeuzem S, Sarrazin C. Consideration of Viral resistance for optimization of direct antiviral therapy of hepatitis C virus genotype 1-infected patients. PloS One. 2015;10(8):e0134395. doi: 10.1371/journal.pone.0134395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Paolucci S, Fiorina L, Mariani B, et al. Naturally occurring resistance mutations to inhibitors of HCV NS5A region and NS5B polymerase in DAA treatment-naïve patients. Virol J. 2013;10:355. doi: 10.1186/1743-422X-10-355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Yang W, Zhao Y, Fabrycki J, et al. Selection of replicon variants resistant to ACH-806, a novel hepatitis C virus inhibitor with no cross-resistance to NS3 protease and NS5B polymerase inhibitors. Antimicrob Agents Chemother. 2008;52(6):2043–2052. doi: 10.1128/AAC.01548-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Lontok E, Harrington P, Howe A, et al. Hepatitis C virus drug resistance-associated substitutions: state of the art summary. Hepatology. 2015;62(5):1623–1632. doi: 10.1002/hep.27934. [DOI] [PubMed] [Google Scholar]
- 6.Sarrazin C. The importance of resistance to direct antiviral drugs in HCV infection in clinical practice. J Hepatol. 2016;64(2):486–504. doi: 10.1016/j.jhep.2015.09.011. [DOI] [PubMed] [Google Scholar]
- 7.Barbotte L, Ahmed-Belkacem A, Chevaliez S, et al. Characterization of V36C, a novel amino acid substitution conferring hepatitis C virus (HCV) resistance to telaprevir, a potent peptidomimetic inhibitor of HCV protease. Antimicrob Agents Chemother. 2010;54(6):2681–2683. doi: 10.1128/AAC.01796-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sullivan JC, De Meyer S, Bartels DJ, et al. Evolution of treatment-emergent resistant variants in telaprevir phase 3 clinical trials. Clin Infect Dis. 2013;57(2):221–229. doi: 10.1093/cid/cit226. [DOI] [PubMed] [Google Scholar]
- 9.Chen ZW, Li H, Ren H, Hu P. Global prevalence of pre-existing HCV variants resistant to direct-acting antiviral agents (DAAs): mining the GenBank HCV genome data. Sci Rep. 2016;6:20310. doi: 10.1038/srep20310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Pawlotsky JM. Hepatitis C virus resistance to direct-acting antiviral drugs in interferon-free regimens. Gastroenterology. 2016;151(1):70–86. doi: 10.1053/j.gastro.2016.04.003. [DOI] [PubMed] [Google Scholar]
- 11.Vermehren J, Susser S, Lange CM, et al. Mutations selected in the hepatitis C virus NS3 protease domain during sequential treatment with boceprevir with and without pegylated interferon alfa-2b. J Viral Hepat. 2012;19(2):120–127. doi: 10.1111/j.1365-2893.2011.01449.x. [DOI] [PubMed] [Google Scholar]
- 12.Kwong AD, Najera I, Bechtel J, et al. Sequence and phenotypic analysis for resistance monitoring in hepatitis C virus drug development: recommendations from the HCV DRAG. Gastroenterology. 2011;140(3):755–760. doi: 10.1053/j.gastro.2011.01.029. [DOI] [PubMed] [Google Scholar]
- 13.Bartels DJ, Sullivan JC, Zhang EZ, et al. Hepatitis C virus variants with decreased sensitivity to direct-acting antivirals (DAAs) were rarely observed in DAA-naïve patients prior to treatment. J Virol. 2013;87(3):1544–1553. doi: 10.1128/JVI.02294-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Besse B, Coste-Burel M, Bourgeois N, Feray C, Imbert-Marcille BM, André-Garnier E. Genotyping and resistance profile of hepatitis C (HCV) genotypes 1–6 by sequencing the NS3 protease region using a single optimized sensitive method. J Virol Methods. 2012;185(1):94–100. doi: 10.1016/j.jviromet.2012.06.011. [DOI] [PubMed] [Google Scholar]
- 15.Bartels DJ, Zhou Y, Zhang EZ, et al. Natural prevalence of hepatitis C virus variants with decreased sensitivity to NS3.4A protease inhibitors in treatment-naïve subjects. J Infect Dis. 2008;198(6):800–807. doi: 10.1086/591141. [DOI] [PubMed] [Google Scholar]
- 16.McPhee F, Friborg J, Levine S, et al. Resistance analysis of the hepatitis C virus NS3 protease inhibitor asunaprevir. Antimicrob Agents Chemother. 2012;56(7):3670–3681. doi: 10.1128/AAC.00308-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Tong X, Guo Z, Wright-Minogue J, et al. Impact of naturally occurring variants of HCV protease on the binding of different classes of protease inhibitors. Biochemistry. 2006;45(5):1353–1361. doi: 10.1021/bi051565g. [DOI] [PubMed] [Google Scholar]
- 18.Wang Y, Rao HY, Xie XW, Wei L. Direct-acting antiviral agents resistance-associated polymorphisms in Chinese treatment-naïve patients infected with genotype 1b hepatitis C virus. Chin Med J (Engl) 2015;128(19):2625–2631. doi: 10.4103/0366-6999.166038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Sarrazin C, Kieffer TL, Bartels D, et al. Dynamic hepatitis C virus genotypic and phenotypic changes in patients treated with the protease inhibitor telaprevir. Gastroenterology. 2007;132(5):1767–1777. doi: 10.1053/j.gastro.2007.02.037. [DOI] [PubMed] [Google Scholar]
- 20.Kieffer TL, Sarrazin C, Miller JS, et al. Telaprevir and pegylated interferon-alpha-2a inhibit wild-type and resistant genotype 1 hepatitis C virus replication in patients. Hepatology. 2007;46(3):631–639. doi: 10.1002/hep.21781. [DOI] [PubMed] [Google Scholar]
- 21.Bae A, Sun SC, Qi X, et al. Susceptibility of treatment-naïve hepatitis C virus (HCV) clinical isolates to HCV protease inhibitors. Antimicrob Agents Chemother. 2010;54(12):5288–5297. doi: 10.1128/AAC.00777-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Susser S, Vermehren J, Forestier N, et al. Analysis of long-term persistence of resistance mutations within the hepatitis C virus NS3 protease after treatment with telaprevir or boceprevir. J Clin Virol. 2011;52(4):321–327. doi: 10.1016/j.jcv.2011.08.015. [DOI] [PubMed] [Google Scholar]
- 23.Fridell RA, Wang C, Sun JH, et al. Genotypic and phenotypic analysis of variants resistant to hepatitis C virus nonstructural protein 5A replication complex inhibitor BMS-790052 in humans: in vitro and in vivo correlations. Hepatology. 2011;54(6):1924–1935. doi: 10.1002/hep.24594. [DOI] [PubMed] [Google Scholar]
- 24.Karino Y, Toyota J, Ikeda K, et al. Characterization of virologic escape in hepatitis C virus genotype 1b patients treated with the direct-acting antivirals daclatasvir and asunaprevir. J Hepatol. 2013;58(4):646–654. doi: 10.1016/j.jhep.2012.11.012. [DOI] [PubMed] [Google Scholar]
- 25.Sun JH, O’Boyle DR, Zhang Y, et al. Impact of a baseline polymorphism on the emergence of resistance to the hepatitis C virus nonstructural protein 5A replication complex inhibitor, BMS-790052. Hepatology. 2012;55(6):1692–1699. doi: 10.1002/hep.25581. [DOI] [PubMed] [Google Scholar]
- 26.Wang C, Huang H, Valera L, et al. Hepatitis C virus RNA elimination and development of resistance in replicon cells treated with BMS-790052. Antimicrob Agents Chemother. 2012;56(3):1350–1358. doi: 10.1128/AAC.05977-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Le Pogam S, Seshaadri A, Kosaka A, et al. Existence of hepatitis C virus NS5B variants naturally resistant to non-nucleoside, but not to nucleoside, polymerase inhibitors among untreated patients. J Antimicrob Chemother. 2008;61(6):1205–1216. doi: 10.1093/jac/dkn085. [DOI] [PubMed] [Google Scholar]
- 28.Tong X, Le Pogam S, Li L, et al. In vivo emergence of a novel mutant L159F/L320F in the NS5B polymerase confers low-level resistance to the HCV polymerase inhibitors mericitabine and sofosbuvir. J Infect Dis. 2014;209(5):668–675. doi: 10.1093/infdis/jit562. [DOI] [PubMed] [Google Scholar]
- 29.Gane EJ, Stedman CA, Hyland RH, et al. Efficacy of nucleotide polymerase inhibitor sofosbuvir plus the NS5A inhibitor ledipasvir or the NS5B non-nucleoside inhibitor GS-9669 against HCV genotype 1 infection. Gastroenterology. 2014;146(3):736–743.e1. doi: 10.1053/j.gastro.2013.11.007. [DOI] [PubMed] [Google Scholar]
- 30.Sarrazin C, Dvory-Sobol H, Svarovskaia ES, et al. Prevalence of resistance-associated substitutions in HCV NS5A, NS5B, or NS3 and outcomes of treatment with ledipasvir and sofosbuvir. Gastroenterology. 2016;151(3):501–512.e1. doi: 10.1053/j.gastro.2016.06.002. [DOI] [PubMed] [Google Scholar]
- 31.Ali S, Leveque V, Le Pogam S, et al. Selected replicon variants with low-level in vitro resistance to the hepatitis C virus NS5B polymerase inhibitor PSI-6130 lack cross-resistance with R1479. Antimicrob Agents Chemother. 2008;52(12):4356–4369. doi: 10.1128/AAC.00444-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Patino-Galindo JA, Salvatierra K, Gonzalez-Candelas F, Lopez-Labrador FX. Comprehensive screening for naturally occurring hepatitis C virus resistance to direct-acting antivirals in the NS3, NS5A, and NS5B genes in worldwide isolates of viral genotypes 1 to 6. Antimicrob Agents Chemother. 2016;60(4):2402–2416. doi: 10.1128/AAC.02776-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.McCown MF, Rajyaguru S, Le Pogam S, et al. The hepatitis C virus replicon presents a higher barrier to resistance to nucleoside analogs than to nonnucleoside polymerase or protease inhibitors. Antimicrob Agents Chemother. 2008;52(5):1604–1612. doi: 10.1128/AAC.01317-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Troke PJ, Lewis M, Simpson P, et al. Characterization of resistance to the nonnucleoside NS5B inhibitor filibuvir in hepatitis C virus-infected patients. Antimicrob Agents Chemother. 2012;56(3):1331–1341. doi: 10.1128/AAC.05611-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Alves R, Queiroz AT, Pessoa MG, et al. The presence of resistance mutations to protease and polymerase inhibitors in hepatitis C virus sequences from the Los Alamos databank. J Viral Hepat. 2013;20(6):414–421. doi: 10.1111/jvh.12051. [DOI] [PubMed] [Google Scholar]
- 36.Zeuzem S, Asselah T, Angus P, et al. Efficacy of the protease inhibitor BI 201335, polymerase inhibitor BI 207127, and ribavirin in patients with chronic HCV infection. Gastroenterology. 2011;141(6):2047–2055. doi: 10.1053/j.gastro.2011.08.051. quiz e2014. [DOI] [PubMed] [Google Scholar]
- 37.Howe AY, Cheng H, Thompson I, et al. Molecular mechanism of a thumb domain hepatitis C virus nonnucleoside RNA-dependent RNA polymerase inhibitor. Antimicrob Agents Chemother. 2006;50(12):4103–4113. doi: 10.1128/AAC.00365-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Shih IH, Vliegen I, Peng B, et al. Mechanistic characterization of GS-9190 (tegobuvir), a novel nonnucleoside inhibitor of hepatitis C virus NS5B polymerase. Antimicrob Agents Chemother. 2011;55(9):4196–4203. doi: 10.1128/AAC.00307-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Sims KD, Lemm J, Eley T, et al. Randomized, placebo-controlled, single-ascending-dose study of BMS-791325, a hepatitis C virus (HCV) NS5B polymerase inhibitor, in HCV genotype 1 infection. Antimicrob Agents Chemother. 2014;58(6):3496–3503. doi: 10.1128/AAC.02579-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Trevino A, de Mendoza C, Parra P, et al. Natural polymorphisms associated with resistance to new antivirals against HCV in newly diagnosed HIV-HCV-coinfected patients. Antivir Ther. 2011;16(3):413–416. doi: 10.3851/IMP1760. [DOI] [PubMed] [Google Scholar]
- 41.Sun SC, Bae A, Qi X, et al. Natural variation in drug susceptibility to HCV polymerase inhibitors in treatment-naïve HCV patient isolates. J Vir Hepat. 2011;18(12):861–870. doi: 10.1111/j.1365-2893.2010.01396.x. [DOI] [PubMed] [Google Scholar]
Acknowledgments
This study was supported in part by grants from the National Natural Science Foundation of China (30930082, 81171561, 30972584, and 81772171) and the National Science and Technology Major Project of China (2008ZX10002-006, 2012ZX10002007001, 2017ZX10202203-007, and 2017ZX10202203-008).
Footnotes
Author contributions
PH and HR conceived the study, ZL and ZC conducted the experiments, ZL collected the data, and ZL, ZC, and HL analyzed the data. All authors contributed toward data analysis, drafting and revising the paper and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
- 1.Thrift AP, El-Serag HB, Kanwal F. Global epidemiology and burden of HCV infection and HCV-related disease. Nat Rev Gastroenterol Hepatol. 2017;14(2):122–132. doi: 10.1038/nrgastro.2016.176. [DOI] [PubMed] [Google Scholar]
- 2.Petruzziello A, Marigliano S, Loquercio G, Cozzolino A, Cacciapuoti C. Global epidemiology of hepatitis C virus infection: an up-date of the distribution and circulation of hepatitis C virus genotypes. World J Gastroenterol. 2016;22(34):7824–7840. doi: 10.3748/wjg.v22.i34.7824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lauer GM, Walker BD. Hepatitis C virus infection. N Engl J Med. 2001;345(1):41–52. doi: 10.1056/NEJM200107053450107. [DOI] [PubMed] [Google Scholar]
- 4.World Health Organization . Guidelines for the Screening, Care, and Treatment of Persons with Chronic Hepatitis C Infection: Updated Version. Geneva: WHO; 2016. [PubMed] [Google Scholar]
- 5.Smith DB, Bukh J, Kuiken C, et al. Expanded classification of hepatitis C virus into 7 genotypes and 67 subtypes: updated criteria and genotype assignment web resource. Hepatology. 2014;59(1):318–327. doi: 10.1002/hep.26744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Messina JP, Humphreys I, Flaxman A, et al. Global distribution and prevalence of hepatitis C virus genotypes. Hepatology. 2015;61(1):77–87. doi: 10.1002/hep.27259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Rao H, Wei L, Lopez-Talavera JC, et al. Distribution and clinical correlates of viral and host genotypes in Chinese patients with chronic hepatitis C virus infection. J Gastroenterol Hepatol. 2014;29(3):545–553. doi: 10.1111/jgh.12398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Niederau C, Hüppe D, Zehnter E, et al. Chronic hepatitis C: treat or wait? Medical decision making in clinical practice. World J Gastroenterol. 2012;18(12):1339–1347. doi: 10.3748/wjg.v18.i12.1339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lawitz E, Lalezari JP, Hassanein T, et al. Sofosbuvir in combination with peginterferon alfa-2a and ribavirin for non-cirrhotic, treatment-naïve patients with genotypes 1, 2, and 3 hepatitis C infection: a randomised, double-blind, phase 2 trial. Lancet Infect Dis. 2013;13(5):401–408. doi: 10.1016/S1473-3099(13)70033-1. [DOI] [PubMed] [Google Scholar]
- 10.Ogata N, Alter HJ, Miller RH, Purcell RH. Nucleotide sequence and mutation rate of the H strain of hepatitis C virus. Proc Natl Acad Sci U S A. 1991;88(8):3392–3396. doi: 10.1073/pnas.88.8.3392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Coppola N, Minichini C, Starace M, Sagnelli C, Sagnelli E. Clinical impact of the hepatitis C virus mutations in the era of directly acting antivirals. J Med Virol. 2016;88(10):1659–1671. doi: 10.1002/jmv.24527. [DOI] [PubMed] [Google Scholar]
- 12.Lontok E, Harrington P, Howe A, et al. Hepatitis C virus drug resistance-associated substitutions: state of the art summary. Hepatology. 2015;62(5):1623–1632. doi: 10.1002/hep.27934. [DOI] [PubMed] [Google Scholar]
- 13.Nitta S, Asahina Y, Matsuda M, et al. Effects of resistance-associated NS5A mutations in hepatitis C virus on viral production and susceptibility to antiviral reagents. Sci Rep. 2016;6:34652. doi: 10.1038/srep34652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Iio E, Shimada N, Abe H, et al. Efficacy of daclatasvir/asunaprevir according to resistance-associated variants in chronic hepatitis C with genotype 1. J Gastroenterol. 2017;52(1):94–103. doi: 10.1007/s00535-016-1225-x. [DOI] [PubMed] [Google Scholar]
- 15.European Association for the Study of the Liver EASL recommendations on treatment of hepatitis C 2016. J Hepatol. 2017;66(1):153–194. doi: 10.1016/j.jhep.2016.09.001. [DOI] [PubMed] [Google Scholar]
- 16.Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol. 2011;28(10):2731–2739. doi: 10.1093/molbev/msr121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Donlin MJ, Szeto B, Gohara DW, Aurora R, Tavis JE. Genome-wide networks of amino acid covariances are common among viruses. J Virol. 2012;86(6):3050–3063. doi: 10.1128/JVI.06857-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Aurora R, Donlin MJ, Cannon NA, Tavis JE. Genome-wide hepatitis C virus amino acid covariance networks can predict response to antiviral therapy in humans. J Clin Invest. 2009;119(1):225–236. doi: 10.1172/JCI37085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Cline MS, Smoot M, Cerami E, et al. Integration of biological networks and gene expression data using Cytoscape. Nat Protoc. 2007;2(10):2366–2382. doi: 10.1038/nprot.2007.324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Izumi N, Hayashi N, Kumada H, et al. Once-daily simeprevir with peginterferon and ribavirin for treatment-experienced HCV genotype 1-infected patients in Japan: the CONCERTO-2 and CONCERTO-3 studies. J Gastroenterol. 2014;49(5):941–953. doi: 10.1007/s00535-014-0949-8. [DOI] [PubMed] [Google Scholar]
- 21.Lenz O, Verbinnen T, Fevery B, et al. Virology analyses of HCV isolates from genotype 1-infected patients treated with simeprevir plus peginterferon/ribavirin in Phase IIB/III studies. J Hepatol. 2015;62(5):1008–1014. doi: 10.1016/j.jhep.2014.11.032. [DOI] [PubMed] [Google Scholar]
- 22.Bartels DJ, Zhou Y, Zhang EZ, et al. Natural prevalence of hepatitis C virus variants with decreased sensitivity to NS3.4A protease inhibitors in treatment-naïve subjects. J Infect Dis. 2008;198(6):800–807. doi: 10.1086/591141. [DOI] [PubMed] [Google Scholar]
- 23.Gao M. Antiviral activity and resistance of HCV NS5A replication complex inhibitors. Curr Opin Virol. 2013;3(5):514–520. doi: 10.1016/j.coviro.2013.06.014. [DOI] [PubMed] [Google Scholar]
- 24.Chen ZW, Li H, Ren H, Hu P. Global prevalence of pre-existing HCV variants resistant to direct-acting antiviral agents (DAAs): mining the GenBank HCV genome data. Sci Rep. 2016;6:20310. doi: 10.1038/srep20310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Sarrazin C. The importance of resistance to direct antiviral drugs in HCV infection in clinical practice. J Hepatol. 2016;64(2):486–504. doi: 10.1016/j.jhep.2015.09.011. [DOI] [PubMed] [Google Scholar]
- 26.Nakamoto S, Kanda T, Wu S, Shirasawa H, Yokosuka O. Hepatitis C virus NS5A inhibitors and drug resistance mutations. World J Gastroenterol. 2014;20(11):2902–2912. doi: 10.3748/wjg.v20.i11.2902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hézode C, Hirschfield GM, Ghesquiere W, et al. Daclatasvir plus peginterferon alfa and ribavirin for treatment-naïve chronic hepatitis C genotype 1 or 4 infection: a randomised study. Gut. 2015;64(6):948–956. doi: 10.1136/gutjnl-2014-307498. [DOI] [PubMed] [Google Scholar]
- 28.Bartels DJ, Sullivan JC, Zhang EZ, et al. Hepatitis C virus variants with decreased sensitivity to direct-acting antivirals (DAAs) were rarely observed in DAA-naïve patients prior to treatment. J Virol. 2013;87(3):1544–1553. doi: 10.1128/JVI.02294-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Fridell RA, Qiu D, Wang C, Valera L, Gao M. Resistance analysis of the hepatitis C virus NS5A inhibitor BMS-790052 in an in vitro replicon system. Antimicrob Agents Chemother. 2010;54(9):3641–3650. doi: 10.1128/AAC.00556-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Bagaglio S, Andolina A, Merli M, Uberti-Foppa C, Morsica G. Frequency of natural resistance within NS5A replication complex domain in hepatitis C genotypes 1a, 1b: possible implication of subtype-specific resistance selection in multiple direct acting antivirals drugs combination treatment. Viruses. 2016;8(4):91. doi: 10.3390/v8040091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Schinazi R, Halfon P, Marcellin P, Asselah T. HCV direct-acting antiviral agents: the best interferon-free combinations. Liver Int. 2014;34(Suppl 1):69–78. doi: 10.1111/liv.12423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Donaldson EF, Harrington PR, O’Rear JJ, Naeger LK. Clinical evidence and bioinformatics characterization of potential hepatitis C virus resistance pathways for sofosbuvir. Hepatology. 2015;61(1):56–65. doi: 10.1002/hep.27375. [DOI] [PubMed] [Google Scholar]
- 33.Hang JQ, Yang Y, Harris SF, et al. Slow binding inhibition and mechanism of resistance of non-nucleoside polymerase inhibitors of hepatitis C virus. J Biol Chem. 2009;284(23):15517–15529. doi: 10.1074/jbc.M808889200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Krishnan P, Tripathi R, Schnell G, et al. Resistance analysis of baseline and treatment-emergent variants in hepatitis C virus genotype 1 in the AVIATOR study with paritaprevir-ritonavir, ombitasvir, and dasabuvir. Antimicrob Agents Chemother. 2015;59(9):5445–5454. doi: 10.1128/AAC.00998-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Li Z, Zhang Y, Liu Y, et al. Naturally occurring drug resistance associated variants to hepatitis C virus direct-acting antiviral agents in treatment-naïve HCV genotype 1b-infected patients in China. Medicine (Baltimore) 2017;96(19):e6830. doi: 10.1097/MD.0000000000006830. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Chromatograms of Sanger sequencing.
Notes: Chromatograms of Sanger sequencing in part of NS3 segment with 80Q (A); with 80R (B). Amino acid site 80 occurring bases substitution leading to amino acid Q change to R which resistance to DAAs, thus, Q80R was described as a variant type.
The prevalence of clinically relevant resistance associated substitutions (without C316N) in various DAA classes.
Abbreviation: RASs, resistance-associated substitutions.
Phylogenetic trees for HCVGT1b NS3 and NS5B genes.
Notes: (A) Phylogenetic tree for HCV GT1b NS3 gene; (B) phylogenetic tree for HCV GT1b NS5B gene. Blue lines represent nonresistant lineages; black lines represent lineages carrying the resistance associated substitutions.
Table S1.
| Target gene | Primers | 5′–3′ sequences |
|---|---|---|
| NS3 | NS3F out | CGAGACCTTGCGGTGGCAG |
| NS3R out | CAGCCGTYTCCGCTTGGTCC | |
| NS3F in | CATCACCTGGGGGGCAGACACC | |
| NS3F in | GTCAGTTGAGTGGCACTCATCAC | |
| NS5A | NS5AF out | GTTTGGGACTGGATATGC |
| NS5AR out | CGTCACGTAGTGGAAATC | |
| NS5AF in | ACCTGGCTCCAGTCCAAG | |
| NS5AF in | CCTCCACRTACTCCTCAG | |
| NS5B | NS5BF out | CGYTGAGTCRTAYTCCTCCATGC |
| NS5BR out | GGGCRCGAGACASGCTGTGATA | |
| NS5BF in | CTCAGYGACGGGTCYTGGTC | |
| NS5BR in | GCTGTGATATATGTCTCCCC |
Abbreviation: HCV, hepatitis C virus.
Table S2.
Summary of HCV RASs to DAAs
| Region | Residue | RAS(s) | DAA(s) | Reference(s) |
|---|---|---|---|---|
| NS3 | C16 | S | Tvr | 4 |
| V36 | A/M/L/G/I/C | Tvr, Boc, Asv, Ptv, Fdv | 5–9 | |
| Q41 | R | Smv | 10, 11 | |
| F43 | S/V/I/L | Boc, Tvr, Ptv, Smv | 6, 10, 12 | |
| T54 | A, S, C, G, V | Tvr, Boc, Fdv, Asv | 5, 9, 10, 12 | |
| V55 | A | Boc, Tvr | 6, 9 | |
| F, I, K, T | Boc, Asv | 1, 13 | ||
| Y56 | H | Ptv, Asv | 5 | |
| L | Asv | 5, 10 | ||
| Q80 | K, R, H, N, G, L | Smv, Asv/Ptv | 1, 5, 6, 10 | |
| A87 | T | Tvr, Boc | 14 | |
| V107 | I | Boc | 5 | |
| R109 | K | Smv, Boc | 15 | |
| R117 | H | Boc | 14 | |
| S122 | A, I, R, T, G, N | Smv, Asv | 5, 9, 16 | |
| R123 | T | Tvr, Boc | 17 | |
| V132I | I | Tvr | 18 | |
| R155 | K/G/T/M/I/L/S/Q/P/N/W | Tvr, Boc, Smv, Vnv, Fdv, Asv, Ptv | 5, 6, 9, 10, 19, 20 | |
| A156 | S/T/F/N/V/I/G/D | Tvr, Boc, Smv, Vnv, Fdv, Asv, Gzv | 5, 6, 10, 19, 20 | |
| V158 | I, M | Boc, Tvr | 5, 6 | |
| D168 | Q/A/Y/V/E/T/N/P/I/H/G/F/S/K | Smv, Vnv, Asv, Fdv, Ptv, Gzv | 5, 6, 9, 10, 13, 21, 22 | |
| V170 | A | Boc, Tvr | 6 | |
| T | Boc, Smv | 5 | ||
| I | Tvr | 18 | ||
| S174 | F/P | Tvr | 14 | |
| M175 | L | Boc | 5 | |
| NS5A | L28 | F, M, T | Dcv, Omv | 5, 6, 18, 23, 24 |
| P29 | S, X | Dcv | 5 | |
| R30 | G, H, P, Q | Dcv | 5, 10 | |
| L31 | V/M/F/I | Dcv, Omv, Ldv | 5, 6, 9 | |
| P32 | L, S, X | Dcv, Ldv | 5, 6, 10 | |
| Q54 | H/N/L/Y | Dcv, Omv | 18, 23, 24 | |
| P58 | S/A/H/T | Dcv | 5, 24 | |
| D | Ldv | 6 | ||
| Q/E62 | D/H | Dcv | 5, 18 | |
| R/A/P/S | Dcv | 24, 25 | ||
| A92 | K/T | Dcv | 5, 6, 24 | |
| Y93 | H/N/C | Dcv, Ldv, Omv | 5, 6, 23, 24, 26 | |
| NS5B | H95 | Q/R | Dsv | 18, 27 |
| L159 | F | Sof, Mcb | 5, 28, 29 | |
| N142 | T/S | R1479, Sof | 27, 30 | |
| S282 | T/R | Sof, Mcb | 5, 6, 30 | |
| M289 | I/L/V | Sof | 3, 6 | |
| A300 | T | R7128 | 31 | |
| V321 | A | Sof | 5 | |
| C316 | H, N, Y, S | Dsv, Nbv, Sof | 5, 6, 9 | |
| L320 | F, I, S | Sof, Mcb, PSI938 | 6, 28–30 | |
| V321 | A | Sof | 5 | |
| A338 | V | Bcv | 18, 32 | |
| S365 | A, L, T | Nbv | 18, 32, 33 | |
| S368 | T | Dsv | 6 | |
| L392 | I | GS9669 | 18 | |
| N411 | S | Dsv | 6 | |
| M414 | L/T/I/V/Q | Dsv, A782759, Nbv | 5, 6, 13 | |
| A421 | V | BMS791325 | 5, 13 | |
| M423 | I, T | Nbv | 13, 34 | |
| I424 | V | Bcv | 35 | |
| M426 | T/V/I | A782759, Nbv | 3, 34 | |
| A442 | T | Bcv | 5, 18, 19 | |
| C445 | F, | GS9190 | 15, 36 | |
| E446 | K, Q | Dsv | 6, 9 | |
| Y448 | C, H | Dsv, GS9190 | 6, 37, 38 | |
| V494 | A, T, L, I | Nbv | 13, 15 | |
| P495 | A, L, S | Bcv, BI207127 | 5, 27, 36, 39 | |
| V499 | A | Bcv | 18, 40 | |
| A553 | I, T, V | Dsv | 6, 10 | |
| G554 | S, D | Dsv | 6, 10 | |
| S556 | G, N, R, C | Dsv | 5, 6, 13, 41 | |
| G558 | R | Dsv | 6 | |
| D559 | G, S, N | Dsv | 6, 9 | |
| Y561 | H | Dsv | 6 |
Abbreviations: HCV, hepatitis C virus; RASs, resistance-associated substitutions; Asv, asunaprevir; Boc, boceprevir; Ptv, paritaprevir; Smv, simeprevir; Tvr, telaprevir; Fdv, faldaprevir; Vnv, vaniprevir; Gzv, grazoprevir; Dcv, daclatasvir; Ldv, ledipasvir; Omv, ombitasvir; Sof, sofosbuvir; Dsv, dasabuvir; Bcv, beclabuvir; Mcb, mericitabine; Nbv, nesbuvir.
Table S3.
Baseline characteristics of patients with and without NS5A RASs
| Characteristics | Wild type (n=36) | Mutation type (n=16) | P |
|---|---|---|---|
| Age (years), median (range) | 50 (18–70) | 42 (35–58) | 0.192 |
| Male (female) | 21 (15) | 7 (9) | >0.05 |
| Alb (g/L), median (range) | 44.35 (36.4–54.9) | 45.25 (24.6–48) | 0.513 |
| ALT (U/L), median (range) | 61 (10–189) | 66 (20–230) | 0.505 |
| AST (U/L), median (range) | 54 (14–163) | 47 (21–199) | 0.657 |
| γ-GGT (IU/L), median (range) | 57 (15–207) | 33 (9–97) | 0.054 |
| Total bilirubin (mg/dL), median (range) | 0.85 (0.44–1.91) | 0.63 (0.37–1.25) | 0.597 |
| Direct bilirubin (mg/dL), median (range) | 0.34 (0.22–0.79) | 0.33 (0.16–0.90) | 0.746 |
| Plt count (×109/L), median (range) | 153 (57–240) | 184 (56–315) | 0.323 |
| AST/Plt, median (range) | 0.43 (0.12–1.63) | 0.26 (0.1–1.80) | 0.867 |
| Hemoglobin (g/dL), median (range) | 153 (85–171) | 142.5 (77.4–178) | 0.907 |
| HCV RNA (log10 IU/mL), median (range) | 6.35 (3.00–7.5) | 6.57 (4.37–7.23) | 0.444 |
Abbreviations: HCV, hepatitis C virus; RASs, resistance-associated substitutions; Alb, albumin; Plt, platelet.