

Abstract
Objective
Adolescent alcohol use (AAU) is associated with brain anomalies, but less is known about long-term neurocognitive effects. Despite theoretical models linking AAU to diminished cognitive control, empirical work testing this relationship with specific cognitive control neural correlates (e.g., prefrontal theta-band EEG dynamics) remains scarce. A longitudinal twin design was used to test the hypothesis that greater AAU is associated with reduced conflict-related EEG theta-band dynamics in adulthood, and to examine the genetic/environmental etiology of this association.
Methods
In a large (N = 718) population-based prospective twin sample, AAU was assessed at ages 11/14/17. Twins completed a flanker task at age 29 to elicit EEG theta-band medial frontal cortex (MFC) power and medial–dorsal prefrontal cortex (MFC-dPFC) connectivity. Two complementary analytic methods (cotwin control analysis; biometric modeling) were used to disentangle the genetic/shared environmental risk towards AAU from possible alcohol exposure effects on theta dynamics.
Results
AAU was negatively associated with adult cognitive control-related theta-band MFC power and MFC-dPFC functional connectivity. Genetic influences primarily underlie these associations.
Conclusions
Findings provide strong evidence that genetic factors underlie the comorbidity between AAU and diminished cognitive control-related theta dynamics in adulthood.
Significance
Conflict-related theta-band dynamics appear to be candidate brain-based endophenotypes/mechanisms for AAU.
Keywords: Adolescent Alcohol Use, Cognitive Control, Cotwin Control, Endophenotype, Functional Connectivity, Theta
1. Introduction
A large body of work suggests that AAU is associated with a variety of neurocognitive anomalies, including structural/functional brain abnormalities and poor neuropsychological test performance (reviewed in Jacobus and Tapert, 2013; Welch et al., 2013). Specifically, heavy drinking adolescents exhibit reduced grey and white matter prefrontal cortex volume (Bellis et al., 2005; Medina et al., 2008; Malone et al., 2014; Wilson et al., 2015) and suboptimal performance on laboratory tasks of executive functioning (Nigg et al., 2006; Squeglia et al., 2009; Malone et al., 2014) compared to adolescents with limited or no history of alcohol exposure. Because most of this research was conducted using cross-sectional observational research designs, less is known about any long-term neurocognitive effects of AAU, nor is it known to what degree neurocognitive dysfunction is a manifestation of the genetic propensity to abuse alcohol versus a consequence of misuse. In addition, recent theoretical models of alcohol and substance abuse implicate dysregulation of cognitive control processes, mediated by the prefrontal cortex as an important risk factor for early alcohol engagement and misuse (Iacono et al., 2008; Zucker et al., 2011). During normative adolescent development, the early maturation of limbic and striatal systems associated with reward-seeking and appetitive motivation occurs concomitantly with the gradual development of control-related prefrontal cortex areas (Casey et al., 2008; Luciana and Collins, 2012). Protracted prefrontal cortex maturation may be exacerbated in adolescents with a familial liability towards behavioral disinhibition/undercontrol (Iacono et al., 2008; Zucker et al., 2011), which predisposes them towards engaging in immediately gratifying, yet harmful, behaviors (e.g., excessive alcohol use) that may further potentiate prefrontal dysfunction (e.g., deleterious causal effects of alcohol exposure; Goldstein and Volkow, 2002). Despite these influential models, empirical work testing the association between AAU and specific neural correlates of cognitive control remains scarce.
Situations requiring cognitive control, such as when competing responses are activated but only one should be selected (response conflict), have been strongly associated with theta-band (3–8 Hz) electrophysiological signatures that are thought to underlie conflict detection and control-related processes (reviewed in (Cavanagh and Frank, 2014; Clayton et al., 2015). Theta-band power enhancement over the medial frontal cortex (MFC) has been consistently observed following demands of response conflict/interference (Cohen et al., 2008; Nigbur et al., 2011; Nigbur et al., 2012; Cohen and Donner, 2013), and current models of cognitive control implicate the MFC in monitoring the environment for situations of conflict/uncertainty (Ridderinkhof et al., 2004a; Ridderinkhof et al., 2004b; Ullsperger et al., 2014). Upon conflict detection, the MFC is thought to signal the need for increased control to regions of the dorsal medial/lateral prefrontal cortex (dPFC), which further implements top-down control/behavioral adaptation processes to resolve response conflict (Ridderinkhof et al., 2004a; Ridderinkhof et al., 2004b; Cohen and Cavanagh, 2011; Cavanagh and Frank, 2014; Cohen, 2014a; Clayton et al., 2015). Empirical and theoretical work suggests that this interregional communication between the MFC and dPFC is biophysically realized through coordinated theta-band rhythmic activity (phase synchronized oscillations; Fries, 2005), which forms a dynamic functional network for information transfer among distant cortical areas (Cavanagh and Frank, 2014; Cohen, 2014a; Clayton et al., 2015).
Given the importance of theta-band MFC power and MFC-dPFC functional connectivity during response conflict/cognitive control, and research suggesting a link between AAU and reduce grey and white matter prefrontal cortex volume (Jacobus and Tapert, 2013; Malone et al., 2014; Wilson et al., 2015), reduced frontal theta dynamics constitutes a plausible neurophysiological mechanism linking AAU and suboptimal cognitive control.
The present investigation was designed to examine whether AAU has any long-term association with prefrontal cortex neurocognitive functioning and the degree to which genetic risk and the possible deleterious consequences of use might account for the observed association. Several lines of research support as relevant causal factors both the potential neurotoxic effects of early alcohol exposure (Jacobus and Tapert, 2013) and the heritability of alcohol-related brain dysfunction (Iacono et al., 2008). Because of this, quasi-experimental research designs/methods, such as comparing members of twin pairs who differ in their degree of early alcohol exposure, are needed to permit stronger etiological inferences (Vaidyanathan et al., 2015).
One such design is the cotwin control (CTC) method, which capitalizes on the genetic and rearing environment similarity of twins discordant for their level of environmental exposure to some agent (e.g., alcohol; McGue et al., 2010). In this design, since monozygotic (MZ) twins share all genetic and shared environmental influence, comparisons between members of an MZ twin pair account for all familial genetic/shared environmental influences, whether measured or not, that may confound any potential nonshared environmental causal effect of AAU on some outcome (e.g., EEG dynamics). As dizygotic (DZ) twins share half of their genetic material, comparisons between members of a DZ twin pair account partially for genetic and fully for shared environmental influence. The CTC method provides a powerful test of the causal effects of alcohol exposure on the brain, since the neural dynamics of the lesser-drinking twin are a close approximation of the expected dynamics of the heavier-drinking twin had he/she drank less. When testing the causal effects of AAU on neurocognitive functioning, if early alcohol exposure has a deleterious causal effect on cognitive control-related theta-band dynamics, the twin who drank more during adolescence should display less activity than the cotwin who consumed less alcohol. In contrast, if a familial vulnerability towards both AAU and weak theta-band dynamics underlies the association, the brain dynamics of both the heavier- and lesser-drinking twins should not differ substantially (McGue et al., 2010).
A second complementary method to examine etiology is biometric modeling, which estimates the amount of genetic and environmental influence shared across both AAU and theta-band dynamics that explains their observed phenotypic association. In this method, the genetic (or environmental) factors underlying AAU are allowed to correlate with the same genetic (or environmental) factors underlying theta activity. Evidence of a causal effect of AAU on reduced theta dynamics would be consistent with a significant overlap between the same nonshared environmental influences on both AAU and theta activity. In contrast, evidence for a genetic (or shared environmental) influence underlying the AAU-theta association would be consistent with a significant overlap between the same genetic (or shared environmental) factors shared across AAU and theta dynamics.
The current study was designed to evaluate whether normative levels of adolescent alcohol exposure are associated with reduced cognitive control-related theta dynamics in adulthood, and if so, whether the association is consistent with the potentially causal effects of AAU or a premorbid familial risk characteristic. A large prospective population-based sample of same-sex twins was assessed throughout adolescence and into adulthood, with comprehensive assessments of alcohol use (e.g., number of intoxications; maximum consumption; frequency/quantity of drinking) obtained at multiple points spanning adolescence (ages 11/14/17), and EEG correlates of cognitive control processes recorded at age 29. This prospective, genetically-informed twin design is thus well suited to test etiological hypotheses regarding the genetic and/or environmental influences underlying the link between adolescent alcohol exposure and adult cognitive control-related EEG components.
We hypothesized that greater AAU would be associated with reduced adult conflict-related theta-band MFC power and MFC-dPFC connectivity, which if true, would support a link between adolescent drinking and diminished cognitive control-related processes in adulthood. Significant drinking effects were followed up using two complementary methods to test etiological hypotheses. First, a cotwin control analysis of within-twin-pair differences in AAU was conducted to test the potential causal effects of AAU on adult theta dynamics. Second, biometric modeling was used to estimate the specific nature of any potential familial (genetic/shared environmental) or nonshared environmental influence underlying the AAU-EEG relationship. Given research supporting both the heritability of alcohol-related brain dysfunction (Iacono et al., 2008) and the potential causal effects of adolescent drinking on the brain (Jacobus and Tapert, 2013), we had two hypotheses regarding etiological influences:
If adolescent drinking has a causal effect on theta-band dynamics in adulthood, then a) within a twin pair, the heavier drinking twin should exhibit decreased theta activity than the lesser drinking twin (reflected by a significant within-pair CTC effect), and b) biometric modeling should be consistent with a significant nonshared environmental correlation between AAU and theta.
If familial factors underlie the association between adolescent drinking and reduced adult theta dynamics, then a) within a twin pair, theta activity should not differ between the heavier and lesser drinking twin (reflected by a non-significant within-pair CTC effect), and b) biometric modeling should be consistent with a familial influence underlying the AAU/theta covariation, with the relative magnitude of genetic/shared environmental correlations casting light on the specific nature of the familial influence.
2. Methods
2.1 Participants
Participants were MZ and same-sex DZ twins drawn from the community-based Minnesota Twin Family Study, who were initially assessed at age 11 and then followed up approximately every three or four years (for details, see Iacono and McGue, 2002; Iacono et al., 2006). Twins who had adolescent alcohol use data at the target assessment ages of 11 (age: M [SD] = 11.7 [0.4]), 14 (14.7 [0.5]), and 17 (18.1 [0.7]), and flanker EEG data at age 29 (29.1 [0.5]) served as participants for the present study. The sample consisted of 718 twins (395 females), with 459 MZ (196 complete pairs) and 259 DZ twins (105 complete pairs).
2.2 Adolescent alcohol use at ages 11, 14, and 17
To quantify adolescent alcohol use (AAU), a composite drinking index was calculated by summing responses from four self-report drinking items: (1) total number of times intoxicated from alcohol (0 = never to 6 = 150 times or more), (2) frequency of drinking in the past 12 months (0 = never to 5 = two or more times every day), (3) typical number of drinks consumed in one session in the past 12 months (0 = none to 6 = 30 or more), and (4) the maximum number of drinks consumed in a 24-hr period since last assessment (0 = none to 6 = 30 or more). At ages 11 and 14, the participants reported on their history of alcohol, nicotine, and other substance use using a computerized substance use (CSU) inventory that was conducted in a private room. At age 17, alcohol and other substance use history was obtained with the Substance Abuse Module (SAM) of the Composite International Diagnostic Interview (Robins et al., 1987), which was administered by trained interviewers. The composite drinking index was calculated separately for each adolescent assessment (11, 14, 17), and then the sum of the three drinking index scores was calculated (possible range: 0–69; see Table S1 in Supplement for descriptive statistics) and used as the measure of AAU in the current report.
Each of these four measures tap different aspects of alcohol exposure and exhibited high within-age item-level correlations (Table S2 in Supplement), and combining them into a single composite measure both produces a measure of AAU that arguably has greater construct validity than any single item while decreasing the risk of observing false-positive findings by conducting item-wise tests. Prior research offers strong support for the construct and psychometric validity of this composite index, including high internal consistency and expected parent-offspring correlations when assessed throughout adolescence (McGue et al., 2014). In addition, higher index scores have been associated with reduced prefrontal grey matter volume and neurocognitive performance in adolescents (Malone et al., 2014; Wilson et al., 2015), and diminished response inhibition-related theta-band MFC power and MFC-dPFC connectivity in young adults (Harper et al., 2016).
Furthermore, we examined the degree to which this composite index of AAU is related to pathological alcohol use during adolescence. At the age 17 assessment, lifetime presence of Diagnostic and Statistical Manual of Mental Disorders, 4th edition (American Psychiatric Association, 1994), alcohol abuse or dependence was established using a best-estimate approach, which uses both adolescent self-reported symptoms as well as parental report on the child (see Iacono et al., 1999, for interviewing and diagnostic details). By age 17, 4.3% (n = 31) participants met criteria for alcohol dependence, and 11.8% (n = 85) met criteria for abuse. AAU scores were significantly higher in those individuals diagnosed with an alcohol use disorder (AUD; collapsed across dependence/abuse) by the age 17 assessment (M [SD] = 16.65 [6.15], n = 116) than those without (M [SD] = 5.16 [5.12], n = 602; F(1,696) = 307.34, p < 0.001), and AAU scores were highly correlated with total AUD symptom counts (r = 0.61). These results indicate that a portion of this sample were problematic drinkers by age 17 (at a rate comparable to those reported in similar samples; Elkins et al., 2006), and suggest that the continuously distributed composite AAU index is related to psychopathological measures of alcohol misuse.
2.3 Flanker task and EEG recording at age 29
Behavioral and EEG data were collected at age 29 during a modified version of the Eriksen flanker task (Eriksen and Eriksen, 1974). The task consisted of three blocks of 150 pseudorandomized trials (100 congruent / 50 incongruent). During each trial, a target letter (S or H) was flanked by distractor letters that were either congruent or incongruent with the center target, which created a set of four stimulus array that occurred with the following frequencies: SSSSS (33.3%; congruent), HHHHH (33.3%; congruent), SSHSS (16.7%; incongruent), and HHSHH (16.75%; incongruent). Letters were presented centrally in white text on a black background. Participants were asked to respond quickly and accurately to the target letter H with one hand and the target letter S with the opposite hand. The target-stimulus hand mapping alternated across blocks (Blocks 1 and 3: S = right button press, H = left button press; Block 2: S = left button press, H = right button press). Short breaks and feedback regarding accuracy were given after each block. The stimulus duration was 100 ms, response window was 1150 ms, and intertrial interval (fixation point) varied between 900 and 1100 ms. Participants completed a set of practice trials before EEG recording.
EEG signals were recorded from 61 scalp electrodes (10/10 placement) at 1024 Hz with an analog DC to 205 Hz bandpass filter using the BioSemi ActiveTwo system (Biosemi, Amsterdam, Netherlands). Vertical and horizontal electrooculogram (EOG) signals were recorded from four electrodes placed above and below the right eye and bilaterally on the temples, respectively, and electrodes were placed on both earlobes to serve as an offline average reference.
2.4 EEG signal processing
Signals were processed offline using MATLAB (version 7.1, Mathworks, Inc.) and EEGLAB software (Delorme and Makeig, 2004). First, continuous signals were down sampled to 256 Hz, highpass filtered at 0.1 Hz (firfilt EEGLAB plugin; Kaiser window, order of 1286), and re-referenced to the averaged earlobe signals. An automated pipeline was used to identify and remove instances of artifacts and inter-electrode electrolyte bridging (Tenke and Kayser, 2001). Descriptive statistics (e.g., absolute temporal variance) were calculated for each electrode and 1s time-range in the continuous data, and data that exceeded four normalized median absolute deviations relative to the median (Rousseeuw and Croux, 1993) in 25% or 75% of a 1s time-range or electrode, respectively, were deleted. To correct for ocular artifacts, EEG signals were decomposed into independent components (ICs) with the Infomax algorithm (Bell and Sejnowski, 1995). The spatial and temporal characteristics of each IC were correlated with the time course of a criterion channel (bipolar vertical or horizontal EOG) and the prototypical topography (inverse weight) of a blink or saccade IC. Components were subtracted from the data if the squared correlation coefficient exceeded an empirically derived threshold derived by an expectation maximization algorithm (Mognon et al., 2011).
Next, epochs of ±2 s aligned to stimulus onset were taken, and screened for artifacts (as detailed above). Deleted electrodes were interpolated via a spherical spline method (Perrin et al., 1989) within epochs containing ≥75% of the original data/electrodes; otherwise, the trial/epoch was discarded. Error trials were excluded to avoid mixing processes related to conflict processing with error-related processes, and all trials were deleted from blocks in which accuracy was ≤50% (3.5% across all subjects/blocks). As recommended by published guidelines for preprocessing EEG data (Keil et al., 2014), epochs were baseline corrected by subtracting the mean prestimulus activity from −200 to −1 ms to remove any slow voltage drift. Finally, trial-level EEG signals were scalp Laplacian (current source density) transformed (Lagrange order = 50; m = 4; λ = 10−5) using a spherical spline surface (Kayser and Tenke, 2006). The Laplacian is a reference-free spatial filter recommended for connectivity analyses (Cohen, 2015a), which attenuates spatially-broad volume-conducted activity (e.g., single source projected to multiple electrodes) that can positively bias functional connectivity estimates (Winter et al., 2007).
2.5 EEG time-frequency analysis
Single-trial signals were transformed into time-frequency representations via wavelet convolution (Cohen, 2014b) by multiplying the EEG power spectrum (calculated via fast Fourier transform [FFT]) by the power spectrum of complex Morlet wavelets [ ], where f is frequency (ranging from 2 to 40 Hz in 25 logarithmic steps), t is time, and σ defines the width of each frequency band, which was set according to c/(2πf), where c is the number of wavelet cycles (increasing from 3.5 to 8 in 25 logarithmic steps to obtain comparable time/frequency precision), and then taking the inverse FFT. The complex signals were down sampled in time to 64 time bins/second. Since unequal trial numbers between conditions can bias power and functional connectivity estimates, for each subject, a random sample (without replacement) of congruent trials was selected to match the number of incongruent trials (M [SD] = 132.90 [34.35]) before calculating power and interelectrode connectivity. This was performed 25 times, and the results were averaged.
The decision to focus on time-frequency EEG activity instead of more traditional time-domain event-related potential (ERP) components (e.g., N2 or P3) in the current report was based on several factors. Time-frequency components, such as theta power, have been demonstrated to more closely index response conflict and cognitive control-related processes, and possess greater statistical power, than conventional ERP measures (Nigbur et al., 2012; Cohen and Donner, 2013). Furthermore, as previous work suggests that the response conflict effect is primarily reflected in activity that is time-locked, but not phase-locked, to stimulus onset (Cohen and Donner, 2013), which is, by definition, activity that is lost using ERP methods (Makeig et al., 2004), time-frequency analysis was required to optimally characterize the electrophysiological and neurocognitive dynamics during cognitive control.
2.5.1 Time-frequency power
From the resulting complex signal Zt, an estimate of the frequency-specific power at each time point was calculated as [real(Zt)2 + imag(Zt)2]. The trial-averaged power was decibel transformed (dB Powertf = 10 × log10[Powertf/Baseline Powerf]), where, for each channel, frequency, and condition, the average prestimulus power from −450 to −250 ms served as the baseline power.
The baseline period for time-frequency analysis differed from that used during preprocessing for the time-domain signals (section 2.4) to avoid potentially mixing post-stimulus activity with the baseline interval following wavelet convolution. Specifically, the temporal smoothing inherent in wavelets may cause some early post-stimulus activity to mix with the early pre-stimulus period (Cohen, 2014b), and because of this, it is important to choose a baseline that ends no closer to the event onset than one-half the wavelet width (σt) at the frequency of interest (Herrmann, Grigutsch, & Busch, 2005; Roach and Mathalon, 2008). At the lower-bound of the theta region of interest (see section 2.6) σt equaled 194 ms, and so a baseline interval of −450 to −250 ms was chosen for the time-frequency power and connectivity analyses.
2.5.2 Functional connectivity
Interelectrode functional connectivity was calculated with the weighted phase-lag index (wPLI; for a discussion and mathematical definition, see Vinck et al., 2011), which is defined as:
where is the cross-spectrum of the complex signals Z between electrodes 1 and 2 (obtained from the wavelet analysis), is the complex conjugate of, E{·} is the expected value operator, sgn is the sign function, |·| is the magnitude, and is the imaginary component of the cross-spectrum. In other words, the wPLI is the absolute value of the average sign of phase differences over trials between two electrodes, weighted by the average distance of the phase differences from the real axis, and values can range from 0 (no connectivity, or 0°/180° phase lag differences due to volume conduction) to 1 (perfect non-0°/180° phase-lagged functional connectivity). Because single source volume conducted activity creates a cross-spectrum that has no phase lag (0°/180°) and can spuriously inflate connectivity estimates, the wPLI partials out no and random phase lag differences and is thus a measure of connectivity largely insensitive to volume conduction artifacts compared to other measures (Vinck et al., 2011; Cohen, 2015b). To remove tonic interelectrode connectivity not modulated by task demands, the wPLI values were baseline corrected for each electrode/frequency/condition by subtracting the mean prestimulus wPLI between −450 to −250 ms (wPLItf = wPLItf − Baseline wPLIf).
2.6 Time-frequency component selection
As depicted in Figure 1A, theta power values were calculated as the average power over a region of interest (ROI) spanning 3–8 Hz and 250–578 ms post-stimulus separately for each condition and subject, defined by the grand average time-frequency power representation. As expected, the topographic map of theta power demonstrated a focal increase over electrodes FCz and Cz, and the pooled power across these electrodes was chosen for statistical analysis of medial frontal cortex (MFC) theta power.
Figure 1.
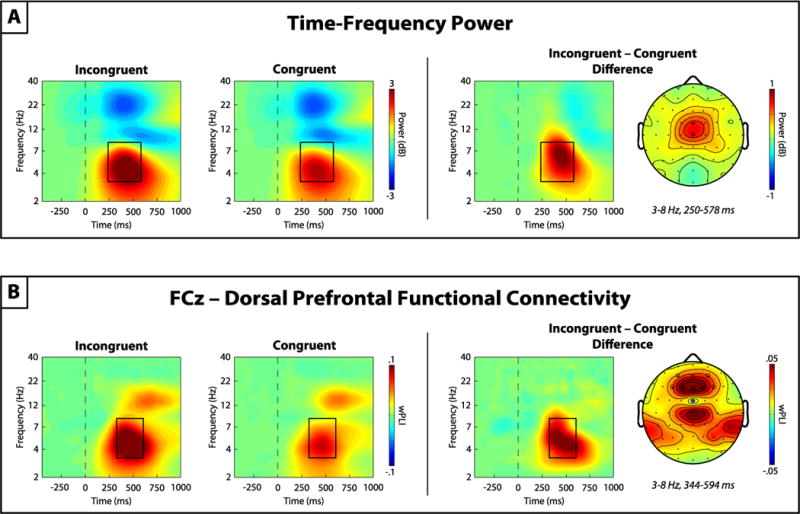
Time-frequency theta-band (3–8 Hz) EEG dynamics. (A) Left: The grand average time-frequency plots of stimulus-locked (time = 0, dashed line) medial frontal power at channels FCz/Cz for incongruent and congruent conditions. Note the strong increase in theta-band power following incongruent stimuli. Right: The response conflict effect (difference between incongruent and congruent trials) on time-frequency power. The time-frequency plot of the response conflict effect for power pooled across FCz/Cz (white electrodes) and the associated topographic distribution of theta-band power show robust theta power enhancement over the medial frontal cortex for incongruent trials. (B) Left: Same as (A), but for FCz–seeded (purple electrode) functional connectivity as measured by the weighted phase lag index (wPLI). These plots illustrate enhanced connectivity between a medial frontal cortex channel (FCz) and a cluster of dorsal medial and dorsolateral prefrontal channels (dPFC; white electrodes), which is augmented during incongruent trials. The black outline boxes denote the region of interest used for statistical analyses.
Guided by prior work (Cohen and Cavanagh, 2011; Harper et al., 2016; Nigbur et al., 2012) and a priori hypotheses regarding theta-band functional connectivity between MFC and dorsal prefrontal cortex (dPFC) electrodes during response conflict/cognitive control (Clayton et al., 2015), pairwise wPLI values were calculated between MFC seed electrode FCz and all other channels. As seen in Figure 1B, the grand averaged plots of connectivity between FCz and a cluster of dPFC electrodes (Fz/F1-4/AFz/AF3-4) revealed a focal connectivity enhancement across a 3–8 Hz and 344–594 ms post-stimulus window. The topographic map of the mean wPLI values across this ROI demonstrated robust connectivity between the MFC seed electrode FCz and electrodes over the dPFC. The mean wPLI across this ROI was calculated and pooled across dPFC electrodes for each subject and condition. For convenience, this measure is referred to as MFC-dPFC connectivity below. While MFC-based connectivity was observed with other areas, particularly midcentral/lateral parietal regions, the decision to focus on dPFC connectivity was guided by a priori hypothesis regarding MFC-dPFC connectivity during cognitive control (Cavanagh and Frank, 2014; Cohen, 2014a; Clayton et al., 2015).
2.7 Statistical analyses
Statistical analyses were conducted in R (R Core Team, 2017). Linear mixed models (LMMs) were fit using lmer from the lme4 package (Bates et al., 2015) with Kenward-Roger adjusted denominator degrees of freedom from the lmerTest package (Kuznetsova et al., 2014). LMMs are commonly used with nested designs, in which, for example, repeated measures are nested within individuals, who are in turn nested within twin pairs, as in the present study. Random intercepts at the individual and twin-pair level accounted for within-individual and -pair correlations, respectively, with respect to all dependent measures, in all LMMs. The fixed effect of stimulus category (incongruent, congruent) on behavior and the EEG measures was evaluated with one-level LMMs.
To examine the (individual-level) association between adolescent alcohol use and each adult outcome of interest (i.e., incongruent or congruent theta-band MFC power, MFC-dPFC connectivity, error rates, reaction times), for each outcome, separate LMM regressions were fit with drinking index scores as the independent variable (fixed effect) and the adult outcome (e.g., incongruent theta-band MFC power) as the dependent variable.
For all significant associations between AAU and some outcome (e.g., adult theta power), two statistical methods (described below) were used to examine the etiological nature of the association.
Significant AAU effects were followed up with a cotwin control analysis (301 complete twin pairs, 196 MZ pairs), where LMM regressions were fit with the adult outcome (e.g., theta power) as the dependent variable and the within-pair AAU effect as the independent (fixed effect) variable (Begg and Parides, 2003), which is the deviation of each individual twin’s drink index score from his/her twin-pair drink index mean (e.g., Twin A drinking score – mean[Twin A score, Twin B score]). The within-pair effect reflects the nonshared environmental effects of AAU unconfounded by familial factors (shared genetic/environmental influences), and is therefore a more appropriate test of whether or not alcohol exposure has a causal effect on the outcome of interest (Begg and Parides, 2003; McGue et al., 2010). Within-pair comparisons of MZ and DZ pairs provide a complete shared environmental control, while MZ pairs provide complete genetic control and DZ pairs provide partial (50%) genetic control. A significant within-pair effect for theta dynamics suggests a causal effect of AAU on the EEG measure, while observing an effect at the individual level in the absence of a significant within-pair effect suggests that the individual-level effect is due primarily to familial factors shared by twins that are confounded with exposure and theta dynamics (McGue et al., 2010).
Bivariate biometric models, which treat the twin pair as the unit of analysis, were estimated using a Cholesky decomposition, to determine the respective roles of genes and environment on the association between AAU and each respective theta measure. Figure 2 provides a graphical depiction of these models, which decompose the observed phenotypic correlation between theta dynamics and AAU into that due to latent variables representing additive genetics (A), reflecting the aggregate/additive effects of alleles over multiple loci, shared environment (C), reflecting the common environmental factors that contribute to twin (familial) similarity, and nonshared environment (E), reflecting the environmental factors unique to each individual twin (i.e., not shared by members of a twin pair) that contribute to twin differences. In these models, A and C are those factors that make twins similar, and E is the factor that make twins different. When observing two phenotypes, such as adolescent alcohol use and theta, biometric modeling tests whether the genetic (A), shared environmental (C), or nonshared environmental (E) factors that influence drinking are the same factors that also influence theta power by estimating the magnitude of the correlations between A, C, and E influences on AAU and the same influences on adult theta dynamics. The magnitude of the genetic, shared environmental, and nonshared environmental correlations between AAU and theta indexes the degree to which such influences are common to both phenotypes. The nonshared environmental correlation is analogous to the within-pair effect in CTC analyses, whereas the genetic and shared environmental correlations reflect the familial factors underlying AAU-EEG associations in the absence of a significant nonshared environmental (within-pair) effect. For example, if additive genetic factors primarily explain the association between drinking and the EEG measure, one would expect to observe a significant genetic correlation (i.e., the correlation between the genetic factors associated with drinking and the genetic factors associated with drinking) in the biometric modeling. These models also estimate the proportion of the phenotypic correlation between AAU and theta dynamics that is explained by genetic and environmental factors.
Figure 2.
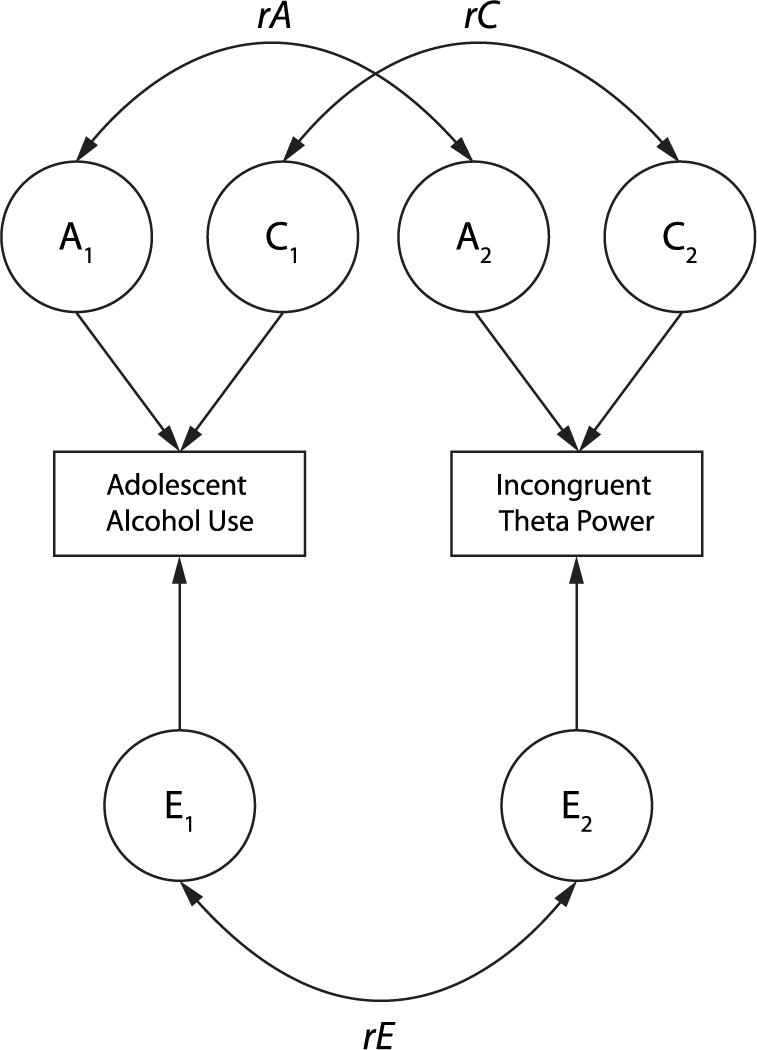
Graphical depiction of a general ACE bivariate model illustrating the variance in each individual observed phenotype (indicated in rectangular boxes) parsed into that explained by latent variables (indicated in circles) reflecting additive genetic (A), shared environmental (C), and nonshared environmental (E) effects, and the associated genetic (rA), shared environmental (rC), and nonshared environmental (rE) correlations, between adolescent alcohol use and incongruent theta-band power. The rE correlation is analogous to the within-pair exposure effect in the cotwin control analysis, while rA and rC capture the familial risk effect.
Biometric ACE models were fit to the raw data using full information maximum likelihood, which provides unbiased parameter estimates when data are missing under relatively mild assumptions, in the OpenMx package in R (Boker et al., 2011). ACE models were chosen based on previous work demonstrating that for many similar task-related frontal EEG components (Anokhin et al., 2008; Demiral et al., 2017; Malone et al., 2017), ACE models fit better than other potential twin models, such as ADE non-additive genetic dominance (D) effect models. The magnitude of the influence of each ACE latent variable on a phenotype is estimated by comparing the expected covariance matrix, derived from the known correlations between family members with respect to A (expected twin correlation: MZ = 1, DZ = 0.5), C (MZ and DZ = 1), and E (MZ and DZ = 0), to the observed covariance matrix. Initially, a base model was fit that estimated all ACE influences, and the associated genetic and environmental correlations, on AAU and theta-band dynamics. Next, to test the overall relative influence of the two sources of familial influence on the AAU-theta associations, the fit of the base model was compared to more restrictive and parsimonious nested models dropping either A or C (i.e., CE and AE models) on two metrics. First, change in model fit was assessed via the difference in the −2 log-likelihood values (−2LL), which approximates a χ2 distribution, between the base and nested models using a 3 degree of freedom likelihood ratio test. A non-significant change in −2LL is considered evidence that the nested model (e.g., AE) does not significantly worsen model fit. In addition, change in model fit was compared using the Bayesian Information Criterion (BIC), which is a combined metric of goodness of fit and model parsimony, where a difference in BIC of > 2 is considered positive evidence in support of the model with lower BIC (Raftery, 1995).
3. Results
3.1 Behavioral results
The expected conflict effect on behavior was observed, with greater error rates and longer reaction times in the incongruent than the congruent condition. Specifically, error rates were higher for incongruent (M [SD] = 4.50% [5.58]) than for congruent stimuli (M [SD] = 3.14 [4.54]; t(717) = 13.26, p < 0.001). Mean reaction time was slower following incongruent (M [SD] = 564.39 ms [66.87]) than congruent stimuli (M [SD] = 519.38 ms [67.24]; t(717) = 63.05, p < 0.001). Adolescent alcohol use was not related to error rates or reaction time in either condition (ps ≥ .19).
3.2 Descriptive statistics and intraclass correlations
Table 1 presents descriptive statistics and twin intraclass correlations for adolescent alcohol use, MFC theta power, and MFC-dPFC theta-band connectivity. In all cases, the MZ intraclass correlations were highly significant, and larger than the DZ correlations, suggesting a genetic influence on these measures.
Table 1.
Descriptive statistics and twin intraclass correlations
| Mean (SD) |
Intraclass Correlations (95% CI) |
||
|---|---|---|---|
| MZ | DZ | ||
| AAU | 7.01 (6.78) |
0.79 (0.74, 0.83) |
0.53 (0.40, 0.63) |
| Incongruent MFC theta power | 2.50 (141) |
0.61 (0.53, 0.68) |
0.25 (0.09, 0.39) |
| Congruent MFC theta power | 1.78 (112) |
0.59 (0.50, 0.66) |
0.24 (0.08, 0.38) |
| Incongruent MFC-dPFC theta functional connectivity | 0.10 (0.09) |
0.38 (0.27, 0.48) |
0.23 (0.07, 0.37) |
| Congruent MFC-dPFC theta functional connectivity | 0.06 (0.06) |
0.30 (0.19, 0.41) |
0.12 (−0.04, 0.27) |
Notes: Abbreviations: AAU = adolescent alcohol use; MFC = medial frontal cortex (electrodes FCz/Cz); dPFC = dorsal prefrontal cortex (electrodes Fz/F1-4/AFz/AF3-4); MZ = monozygotic; DZ = dizygotic; CI = confidence interval. Values are in decibels for power, and weighted phase lag index units for theta-band connectivity (all relative to a −450 to −250 baseline). Sample sizes for intraclass correlations were 459 MZ twins (196 full pairs) and 259 DZ twins (105 full pairs).
3.3 Response conflict modulation of theta-band dynamics
As depicted in the incongruent minus congruent difference plots in Figure 1A, MFC theta power was greater during incongruent trials than congruent trials [t(717) = 29.59, p < 0.001]. Incongruent trials were also associated with greater theta-band functional connectivity between MFC electrode FCz and a cluster of electrodes over the dorsal prefrontal cortex (Figure 1B) than congruent trials [t(717) = 15.84, p < 0.001].
3.4 Adolescent alcohol use (AAU) and adult theta-band EEG dynamics
Table 2 presents the results from the separate LMMs between AAU and adult theta-band dynamics. Adolescent alcohol exposure was negatively associated with MFC power and MFC-dPFC functional connectivity during incongruent trials. Congruent MFC power and MFC-dPFC connectivity were not significantly related to AAU. These results suggest that AAU is specifically associated with diminished cognitive control/conflict-related theta-band dynamics, rather than overall power and connectivity, in adulthood.
Table 2.
Relationship between medial frontal theta-band dynamics and adolescent drinking
| MFC Power Incongruent
|
MFC Power Congruent
|
MFC-dPFC FC Incongruent
|
MFC-dPFC FC Congruent
|
|||||
|---|---|---|---|---|---|---|---|---|
| t (df) | p-value | t (df) | p-value | t (df) | p-value | t (df) | p-value | |
| AAU | −2.88 (668) |
0.004 | −1.88 (662) |
0.061 | −3.10 (608) |
0.002 | −1.71 (580) |
0.088 |
Notes: Results of the separate linear mixed models of the relationship between AAU and theta-band MFC power or MFC-dPFC functional connectivity for incongruent or congruent Flanker conditions. P-values were calculated via Kenward-Roger approximation.
Abbreviations: AAU = adolescent alcohol use; MFC = medial frontal cortex; dPFC = dorsal prefrontal cortex; FC = functional connectivity.
Two complementary analytic methods (cotwin control analysis; bivariate biometric modeling) were used to investigate the etiological basis of these two significant AAU–EEG associations.
3.5 Cotwin control analysis of potential causal AAU effects
Within the separate CTC models, the within-pair effect was not significantly associated with either incongruent MFC power [t(300) = −1.47, p = 0.142] or incongruent MFC-dPFC connectivity [t(300) = −1.04, p = 0.301]. In both cases, the within-twin pair effect did not differ between MZ and DZ twins; adding fixed-effect terms for zygosity and the zygosity by within-pair interaction did not improve model fit [Δχ2s(2) ≤ 1.54, ps ≥ 0.463]; thus, these terms were removed from the final models reported above. This pattern of CTC results is consistent with a preexisting familial risk, and not a causal effect of early drinking, underlying the association between AAU and reduced adult conflict-related theta-band dynamics.
3.6 Bivariate biometric modeling of AAU and adult theta-band dynamics
Model fitting results for the two separate bivariate models of AAU and either incongruent MFC power or incongruent MFC-dPFC connectivity are presented in Table 3. In both cases, the model fitting results indicated that dropping A from the models significantly worsened fit, as evidenced by significant likelihood ratio tests and more positive BIC values compared to the base ACE model. In contrast, dropping C from both models had a negligible effect on model fit, as indicated by non-significant likelihood ratio tests and more negative BIC values than the base ACE model, which suggests no significant contribution of shared environmental effects to AAU or adult theta-band dynamics.
Table 3.
Fit statistics for full and nested bivariate models
| Models | −2LL | df | BIC | ΔBIC | Δ−2LL | p-value |
|---|---|---|---|---|---|---|
| AAU – MFC Power Incongruent | ||||||
| Base ACE Model | 6988.17 | 1425 | 7054.53 | |||
| Drop C | 6991.02 | 1428 | 7039.29 | −15.24 | 2.85 | 0.415 |
| Drop A | 7033.67 | 1428 | 7081.93 | 27.40 | 45.50 | < 0.001 |
| AAU – MFC-dPFC FC Incongruent | ||||||
| Base ACE Model | 7435.34 | 1425 | 7501.70 | |||
| Drop C | 7438.12 | 1428 | 7486.39 | −15.31 | 2.78 | 0.427 |
| Drop A | 7469.81 | 1428 | 7518.07 | 16.37 | 34.47 | < 0.001 |
Notes: Model fit indices for the bivariate models. To test the relative contribution of genetic/shared environmental influence, relative fits for the nested models dropping C or A influence (AE or CE models, respectively) were compared to the base ACE models on two metrics: 1) change in BIC (ΔBIC), which is a combined metric of goodness of fit and model parsimony, with lower BIC values indicating better relative fit, and 2) change in the −2LL (Δ−2LL), which follows a χ2 distribution, using a 3 degrees of freedom likelihood ratio test. The best-fitting models are in bold. In all cases, dropping the shared environmental influence did not worsen fit, whereas dropping the additive genetic influence significantly degraded model fit. Abbreviations: −2LL = −2 log likelihood value; df = degrees of freedom; BIC = Bayesian information criteria (parameter-adjusted); A = additive genetic effects; C = shared environmental effects; E = nonshared environmental effects; AAU = adolescent alcohol use; MFC = medial frontal cortex; dPFC = dorsal prefrontal cortex; FC = functional connectivity.
Table 4 presents the parameter estimates and 95% confidence intervals derived from the best fitting bivariate AE models for each separate theta-band measure. AAU and conflict-related theta-band dynamics showed significant genetic heritability. There were modest but statistically significant negative phenotypic correlations between AAU and adult theta-band measures. As expected given the cotwin control results, the nonshared environmental correlations (rE) between each EEG measure and AAU were effectively zero, while the genetic correlations (rG) were modest and statistically significant. In addition, a large majority (79–96%) of the phenotypic correlation between AAU and each respective theta measure was due to genetic influences.
Table 4.
Genetic and environmental influences on the association between AAU and theta-band dynamics
| AAU | Theta-band Phenotype | Model-implied correlations | Proportion (%) of rP due to |
|||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Phenotype 1 – Phenotype 2 | r | A | E | A | E | rP | rG | rE | A | E |
| AAU – MFC Power Incongruent | −.12 | .80 (.75, .84) |
.20 (.16, .25) |
.60 (.51, .67) |
.40 (.33, .49) |
−.12 (−.20, −.04) |
−.16 (−.28, −.04) |
−.02 (−.15, .12) |
96 | 4 |
| AAU – MFC-dPFC FC Incongruent | −.13 | .80 (.75, .84) |
.20 (.16, .25) |
.39 (.28, .50) |
.61 (.50, .72) |
−.12 (−.20, −.05) |
−.1 7 (−.33, −.02) |
−.08 (−.21, .06) |
79 | 21 |
Notes: Parameter estimates from the separate bivariate AE models between AAU and each theta-band EEG measure. Abbreviations: AAU = adolescent alcohol use; MFC = medial frontal cortex; dPFC = dorsal prefrontal cortex; FC = functional connectivity; r = observed phenotypic correlation; A = additive genetic effects with 95% confidence intervals (CIs); E = nonshared environmental effects; rP = model-implied phenotypic correlation; rG = genetic correlation; rE = nonshared environmental correlation.
Overall, the results of the bivariate biometric modeling are consistent with those obtained from the cotwin control analysis, which together suggest that heritable genetic factors, and not a nonshared environmental alcohol exposure effect, underlie the association between AAU and diminished adult conflict-related theta-band dynamics.
4. Discussion
The present study evaluated the association between adolescent alcohol use and adult EEG correlates of cognitive control processes in a large population-based twin sample prospectively assessed at ages 11, 14, 17, and 29. Consistent with past work reporting an association between AAU and various brain anomalies, we found that greater adolescent alcohol exposure was associated with reduced theta-band MFC power and MFC-dPFC functional connectivity during demands of response conflict/cognitive control processes at age 29. A novel contribution of the current study derived from our use of a longitudinal twin design to examine the genetic and environmental influence on the prospective association between AAU and adult theta. Findings from two complementary tests of etiology (cotwin control analysis of within-twin-pair differences in drinking; bivariate biometric modeling) jointly suggested that the relationship between diminished adult theta-band dynamics and AAU was best explained by heritable genetic influences. The current report provides, to the best of our knowledge, the first evidence that adolescent drinking is related to reduced adult theta-band processes during demands of cognitive control, and that a premorbid genetic risk towards early alcohol use, and not the direct causal effect of alcohol exposure, likely underlies the relationship between adolescent drinking and diminished adult theta dynamics. As such, deviations in theta dynamics appear to possess key characteristics required of an endophenotype for AAU (Iacono et al., 2016). Should genetic variants associated with theta dynamics be identified, our findings have the potential to provide insights into how these variants are related to brain function associated with AAU liability.
Heavier adolescent drinking was negatively associated with reduced adult theta-band power over the MFC and functional connectivity between the MFC and dPFC regions during demands of response conflict (incongruent trials). This finding supports the hypothesis that AAU is related to diminished prefrontal cortex processes related to cognitive control, and is consistent with previous work detailing prefrontal cortex dysfunction in early drinkers (Jacobus and Tapert, 2013; Welch et al., 2013). In the context of recent models of cognitive control (Cavanagh and Frank, 2014; Clayton et al., 2015), the consequence of diminished theta-band dynamics may be a reduced ability to ignore irrelevant distracting information to suppress an inappropriate competing response (MFC power), and decreased execution of behavioral adaptation/cognitive control processes to resolve conflict (MFC-dPFC connectivity).
Converging evidence from the cotwin control analysis of within-pair differences in adolescent alcohol exposure and bivariate biometric modeling suggested that the association between AAU and theta dynamics was consistent with a premorbid genetic risk towards both early drinking and reduced theta in adulthood. Results of the cotwin control analysis suggested that within-pair differences in AAU were not related to conflict-related MFC power or MFC-dPFC connectivity; in other words, within a twin pair, the heavier drinking twin exhibited MFC power and MFC-dPFC connectivity comparable to his/her lesser drinking cotwin. The CTC results argue against a causal effect of adolescent alcohol exposure on adult theta-band dynamics, as a deleterious causal effect should produce a difference between heavier- and lesser-exposed twins (e.g., decreased theta in the greater exposed twin), and these findings are instead consistent with a familial confounding underlying the individual-level association (McGue et al., 2010). In addition, the bivariate biometric modeling provided corroborating support for a familial association between AAU and theta-band dynamics. The modeling suggested that AAU and theta dynamics were heritable, and, though the genetic correlations between AAU and each theta measure were modest in size, shared genetic factors accounted for a majority of the phenotypic association between AAU and reduced adult theta. Taken together, the results of these two methods of testing etiological hypotheses provide strong evidence that the association between adolescent alcohol use and reduced theta-band correlates of cognitive control/conflict in adulthood is better explained by genetic influences common to both phenotypes, rather than a direct effect of alcohol exposure on the brain.
The finding that conflict-related theta-band EEG activity and AAU share a common premorbid genetic liability is consistent with recent theories of alcohol and substance use disorder development, which broadly propose that a core risk pathway to substance misuse is through poor cognitive control processes and behavioral disinhibition. In these models, a genetic vulnerability towards early-onset substance misuse is partly expressed through a preexisting dysregulation of cognitive control-related brain mechanisms, which may then lead to difficulty inhibiting inappropriate responses/impulses and/or a bias towards immediate over long-term rewards (Iacono et al., 2008; Dick et al., 2010; Zucker et al., 2011). Given the established role of MFC theta power and MFC-dPFC connectivity during demands of cognitive control (Cavanagh and Frank, 2014), it is plausible that these EEG dynamics may be candidate brain-based mechanisms or expressions of this genetic vulnerability towards alcohol/substance misuse. Taken together, these findings reflect a step towards understanding the neurophysiological correlates/potential mechanisms of alcohol-related cognitive control dysfunction and disinhibition (Iacono et al., 2008; Dick et al., 2010; Zucker et al., 2011), and strongly suggest that diminished cognitive control-related theta dynamics are part of a constellation of heritable characteristics associated with adolescent drinking.
Overt behavioral performance did not relate to AAU. This may suggest the presence of some compensatory mechanism (e.g., sensorimotor beta; attention-related alpha) that prevents behavioral slips even when prefrontal response conflict processes are lacking, that other cognitive control-related processes important to optimal task performance are relatively intact in the presence of decreased conflict-related theta, or that EEG correlates of response conflict/cognitive control may be more closely associated with adolescent drinking than performance-based measures. In addition, despite the finding that theta dynamics were significantly related to drinking only during response conflict trials, given the lack of association between AAU and behavioral performance, it is possible that individual differences in other more global cognitive states that may affect EEG characteristics, such as wakefulness or general vigilance (Rangaswamy and Porjesz, 2014; Clayton et al., 2015), may have contributed to the observed effects. Further work is needed to tease apart the relationship between adolescent drinking and the various subdomains (e.g., sustained attention, conflict processing) of cognitive control.
There are of course some limitations to this study. As this report focused on normative levels of adolescent drinking in an epidemiologically-derived sample, the results may not easily generalize to clinical populations or adolescents with severe alcohol exposure. However, it should be noted that 16% of our sample (n = 116) did meet clinical criteria for an alcohol use disorder by age 17, suggesting that problematic adolescent drinking was represented in our sample. Similarly, these results may not generalize to cases of very early exposure, as very few twins reported any alcohol use prior to age 11. Additionally, given our focus on adolescent drinking, we did not explore the effects of adult alcohol use on brain dynamics. As the genetic and environmental influences on alcohol use behaviors changes with age (Vrieze et al., 2012), future work should explore whether the etiological influence underlying the alcohol-theta association changes across the lifespan. Given that AAU did not relate significantly to behavioral performance, more work is needed to determine the clinical significance of reduced midfrontal theta. Along this line, future studies should investigate whether diminished conflict-related theta is present in other psychopathologies characterized by frontal lobe dysfunction (e.g., illicit drug addiction, attention-deficit/hyperactivity disorder, antisocial personality disorder), and test whether reduced theta can be used to predict future problematic alcohol use. Finally, despite having a sample that exceeded 700 participants, it is possible that an environmental influence could be detected in an even larger sample, and thus a potential environmental influence cannot be unequivocally ruled out.
These results provide strong evidence suggesting that a heritable genetic vulnerability underlies the comorbidity between adolescent drinking and diminished adult conflict-related prefrontal theta-band dynamics, and offer empirical support for theoretical models implicating poor cognitive control processes as an expression of the genetic liability towards behavioral disinhibition and early-onset substance misuse.
Supplementary Material
Highlights.
Adolescent alcohol use (AAU) predicted reduced adult conflict-related frontal theta EEG dynamics.
Genetic factors primarily accounted for the association between AAU and adult theta dynamics.
These novel results advance prefrontal theta dynamics as an endophenotype for adolescent drinking.
Acknowledgments
This work was supported by the National Institute of Health grants DA005147 and AA009367. J.H. was supported by the National Science Foundation Graduate Research Fellowship under Grant No. 00039202.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Conflict of Interest
None of the authors have potential conflict of interest to be disclosed.
References
- American Psychiatric Association. Diagnostic and statistical manual of mental disorders. 4th. Washington, DC: Author; 1994. [Google Scholar]
- Anokhin AP, Golosheykin S, Heath AC. Heritability of frontal brain function related to action monitoring. Psychophysiology. 2008;45:524–34. doi: 10.1111/j.1469-8986.2008.00664.x. [DOI] [PubMed] [Google Scholar]
- Bates D, Maechler M, Bolker B, Walker S. Fitting linear mixed-effects models using lme4. J Stat Softw. 2015;67:1–48. [Google Scholar]
- Begg MD, Parides MK. Separation of individual-level and cluster-level covariate effects in regression analysis of correlated data. Stat Med. 2003;22:2591–602. doi: 10.1002/sim.1524. [DOI] [PubMed] [Google Scholar]
- Bell AJ, Sejnowski TJ. An information maximization approach to blind separation and blind deconvolution. Neural Comput. 1995;7:1129–59. doi: 10.1162/neco.1995.7.6.1129. [DOI] [PubMed] [Google Scholar]
- Bellis MD, Narasimhan A, Thatcher DL, Keshavan MS, Soloff P, Clark DB. Prefrontal cortex, thalamus, and cerebellar volumes in adolescents and young adults with adolescent‐ onset alcohol use disorders and comorbid mental disorders. Alcohol Clin Exp Res. 2005;29:1590–600. doi: 10.1097/01.alc.0000179368.87886.76. [DOI] [PubMed] [Google Scholar]
- Boker S, Neale M, Maes H, Wilde M, Spiegel M, Brick T, et al. OpenMx: An Open Source Extended Structural Equation Modeling Framework. Psychometrika. 2011;76:306–17. doi: 10.1007/s11336-010-9200-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casey BJ, Getz S, Galvan A. The adolescent brain. Dev Rev. 2008;28:62–77. doi: 10.1016/j.dr.2007.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cavanagh JF, Frank MJ. Frontal theta as a mechanism for cognitive control. Trends Cogn Sci. 2014;18:414–21. doi: 10.1016/j.tics.2014.04.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clayton MS, Yeung N, Kadosh RC. The roles of cortical oscillations in sustained attention. Trends Cogn Sci. 2015;19:188–95. doi: 10.1016/j.tics.2015.02.004. [DOI] [PubMed] [Google Scholar]
- Cohen MX. A neural microcircuit for cognitive conflict detection and signaling. Trends Cogn Sci. 2014a;37:480–90. doi: 10.1016/j.tins.2014.06.004. [DOI] [PubMed] [Google Scholar]
- Cohen MX. Analyzing Neural Time Series Data: Theory and Practice. Cambridge, MA: MIT Press; 2014b. [Google Scholar]
- Cohen MX. Comparison of different spatial transformations applied to EEG data: A case study of error processing. Int J Psychophysiol. 2015a;97:245–57. doi: 10.1016/j.ijpsycho.2014.09.013. [DOI] [PubMed] [Google Scholar]
- Cohen MX. Effects of time lag and frequency matching on phase-based connectivity. J Neurosci Methods. 2015b;250:137–46. doi: 10.1016/j.jneumeth.2014.09.005. [DOI] [PubMed] [Google Scholar]
- Cohen MX, Cavanagh JF. Single-trial regression elucidates the role of prefrontal theta oscillations in response conflict. Front Psychol. 2011;2:30. doi: 10.3389/fpsyg.2011.00030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohen MX, Donner TH. Midfrontal conflict-related theta-band power reflects neural oscillations that predict behavior. J Neurophysiol. 2013;110:2752–63. doi: 10.1152/jn.00479.2013. [DOI] [PubMed] [Google Scholar]
- Cohen MX, Ridderinkhof KR, Haupt S, Elger CE, Fell J. Medial frontal cortex and response conflict: Evidence from human intracranial EEG and medial frontal cortex lesion. Brain Res. 2008;1238:127–42. doi: 10.1016/j.brainres.2008.07.114. [DOI] [PubMed] [Google Scholar]
- Delorme A, Makeig S. EEGLAB: an open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J Neurosci Methods. 2004;134:9–21. doi: 10.1016/j.jneumeth.2003.10.009. [DOI] [PubMed] [Google Scholar]
- Demiral SB, Golosheykin S, Anokhin AP. Genetic influences on functional connectivity associated with feedback processing and prediction error: Phase coupling of theta-band oscillations in twins. Int J Psychophysiol. 2017;115:133–41. doi: 10.1016/j.ijpsycho.2016.12.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dick DM, Smith G, Olausson P, Mitchell SH, Leeman RF, O’Malley SS, et al. Understanding the construct of impulsivity and its relationship to alcohol use disorders. Addict Biol. 2010;15:217–26. doi: 10.1111/j.1369-1600.2009.00190.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elkins IJ, King SM, McGue M, Iacono WG. Personality traits and the development of nicotine, alcohol, and illicit drug disorders: Prospective links from adolescence to young adulthood. J Abnorm Psychol. 2006;115:26–39. doi: 10.1037/0021-843X.115.1.26. [DOI] [PubMed] [Google Scholar]
- Eriksen BA, Eriksen CW. Effects of noise letters upon identification of a target letter in a nonsearch task. Percept Psychophys. 1974;16:143–9. [Google Scholar]
- Fries P. A mechanism for cognitive dynamics: neuronal communication through neuronal coherence. Trends Cogn Sci. 2005;9:474–80. doi: 10.1016/j.tics.2005.08.011. [DOI] [PubMed] [Google Scholar]
- Goldstein RZ, Volkow ND. Drug addiction and its underlying neurobiological basis: Neuroimaging evidence for the involvement of the frontal cortex. Am J Psychiatry. 2002;159:1642–52. doi: 10.1176/appi.ajp.159.10.1642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harper J, Malone SM, Iacono WG. Impact of alcohol use on EEG dynamics of response inhibition: a cotwin control analysis. Addict Biol. 2016 doi: 10.1111/adb.12481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herrmann CS, Grigutsch M, Busch NA. EEG Oscillations and Wavelet Analysis. In: Handy TC, editor. Event-related potentials: A methods handbook. Cambridge, MA: MIT Press; 2005. pp. 229–259. [Google Scholar]
- Iacono WG, Carlson SR, Taylor J, Elkins IJ, McGue M. Behavioral disinhibition and the development of substance-case disorders: Findings from the Minnesota Twin Family Study. Dev Psychopathol. 1999;11:869–900. doi: 10.1017/s0954579499002369. [DOI] [PubMed] [Google Scholar]
- Iacono WG, Malone SM, McGue M. Behavioral disinhibition and the development of early-onset addiction: common and specific influences. Annu Rev Clin Psychol. 2008;4:325–48. doi: 10.1146/annurev.clinpsy.4.022007.141157. [DOI] [PubMed] [Google Scholar]
- Iacono WG, Malone SM, Vrieze SI. Endophenotype best practices. Int J Psychophysiol. 2016;111:115–44. doi: 10.1016/j.ijpsycho.2016.07.516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iacono WG, McGue M. Minnesota Twin Family Study. Twin Res. 2002;5:482–7. doi: 10.1375/136905202320906327. [DOI] [PubMed] [Google Scholar]
- Iacono WG, McGue M, Krueger RF. Minnesota Center for Twin and Family Research. Twin Res Hum Genet. 2006;9:978–84. doi: 10.1375/183242706779462642. [DOI] [PubMed] [Google Scholar]
- Jacobus J, Tapert SF. Neurotoxic Effects of Alcohol in Adolescence. Annu Rev Clin Psychol. 2013;9:703–21. doi: 10.1146/annurev-clinpsy-050212-185610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kayser J, Tenke CE. Principal components analysis of Laplacian waveforms as a generic method for identifying ERP generator patterns: I. Evaluation with auditory oddball tasks. Clin Neurophysiol. 2006;117:348–68. doi: 10.1016/j.clinph.2005.08.034. [DOI] [PubMed] [Google Scholar]
- Keil A, Debener S, Gratton G, Junghöfer M, Kappenman ES, Luck SJ, et al. Committee report: Publication guidelines and recommendations for studies using electroencephalography and magnetoencephalography. Psychophysiology. 2014;51:1–21. doi: 10.1111/psyp.12147. [DOI] [PubMed] [Google Scholar]
- Kuznetsova A, Brockhoff P, Christensen R. lmerTest: tests in linear mixed effects models. R package version 2.0-32. 2014 URL https://CRAN.R-project.org/package=lmerTest.
- Luciana M, Collins PF. Incentive Motivation, Cognitive Control, and the Adolescent Brain: Is It Time for a Paradigm Shift? Child Dev Perspect. 2012;6:392–9. doi: 10.1111/j.1750-8606.2012.00252.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Makeig S, Debener S, Onton J, Delorme A. Mining event-related brain dynamics. Trends Cogn Sci. 2004;5:204–10. doi: 10.1016/j.tics.2004.03.008. [DOI] [PubMed] [Google Scholar]
- Malone SM, Luciana M, Wilson S, Sparks JC, Hunt RH, Thomas KM, et al. Adolescent Drinking and Motivated Decision-Making: A Cotwin-Control Investigation with Monozygotic Twins. Behav Genet. 2014;44:407–18. doi: 10.1007/s10519-014-9651-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malone SM, McGue M, Iacono WG. What can time-frequency and phase coherence measures tell us about the genetic basis of P3 amplitude? Int J Psychophysiol. 2017;115:40–56. doi: 10.1016/j.ijpsycho.2016.11.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGue M, Malone S, Keyes M, Iacono WG. Parent-Offspring Similarity for Drinking: A Longitudinal Adoption Study. Behav Genet. 2014;44:620–8. doi: 10.1007/s10519-014-9672-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGue M, Osler M, Christensen K. Causal Inference and Observational Research: The Utility of Twins. Perspect Psychol Sci. 2010;5:546–56. doi: 10.1177/1745691610383511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Medina KL, McQueeny T, Nagel BJ, Hanson KL, Schweinsburg AD, Tapert SF. Prefrontal cortex volumes in adolescents with alcohol use disorders: Unique gender effects. Alcohol Clin Exp Res. 2008;32:386–94. doi: 10.1111/j.1530-0277.2007.00602.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mognon A, Jovicich J, Bruzzone L, Buiatti M. ADJUST: An automatic EEG artifact detector based on the joint use of spatial and temporal features. Psychophysiology. 2011;48:229–40. doi: 10.1111/j.1469-8986.2010.01061.x. [DOI] [PubMed] [Google Scholar]
- Nigbur R, Cohen MX, Ridderinkhof KR, Stuermer B. Theta Dynamics Reveal Domain-specific Control over Stimulus and Response Conflict. J Cogn Neurosci. 2012;24:1264–74. doi: 10.1162/jocn_a_00128. [DOI] [PubMed] [Google Scholar]
- Nigbur R, Ivanova G, Stuermer B. Theta power as a marker for cognitive interference. Clin Neurophysiol. 2011;122:2185–94. doi: 10.1016/j.clinph.2011.03.030. [DOI] [PubMed] [Google Scholar]
- Nigg JT, Wong MM, Martel MM, Jester JM, Puttler LI, Glass JM, et al. Poor response inhibition as a predictor of problem drinking and illicit drug use in adolescents at risk for alcoholism and other substance use disorders. J Am Acad Child Adolesc Psychiatry. 2006;45:468–75. doi: 10.1097/01.chi.0000199028.76452.a9. [DOI] [PubMed] [Google Scholar]
- Perrin F, Pernier J, Bertrand O, Echallier JF. Spherical splines for scalp potential and current density mapping. Electroencephalogr Clin Neurophysiol. 1989;72:184–7. doi: 10.1016/0013-4694(89)90180-6. [DOI] [PubMed] [Google Scholar]
- R Core Team. R: A language and environment for statistical computing. Vienna, Austria: 2017. https://www.r-project.org/ [Google Scholar]
- Raftery AE. Bayesian model selection in social research. Sociol Methodol. 1995;25:111–63. [Google Scholar]
- Rangaswamy M, Porjesz B. Understanding alcohol use disorders with neuroelectrophysiology. In: Sullivan EV, Pfefferbaum A, editors. Handb Clin Neurol. New York, NY: Elsevier; 2014. pp. 383–414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ridderinkhof KR, Ullsperger M, Crone EA, Nieuwenhuis S. The role of the medial frontal cortex in cognitive control. Science. 2004a;306:443–7. doi: 10.1126/science.1100301. [DOI] [PubMed] [Google Scholar]
- Ridderinkhof KR, van den Wildenberg WPM, Segalowitz SJ, Carter CS. Neurocognitive mechanisms of cognitive control: The role of prefrontal cortex in action selection, response inhibition, performance monitoring, and reward-based learning. Brain Cogn. 2004b;56:129–40. doi: 10.1016/j.bandc.2004.09.016. [DOI] [PubMed] [Google Scholar]
- Roach BJ, Mathalon DH. Event-related EEG time-frequency analysis: An overview of measures and an analysis of early gamma band phase locking in schizophrenia. Schizophr Bull. 2008;111:1719–32. doi: 10.1093/schbul/sbn093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robins L, Babor T, Cottler L. Composite international diagnostic interview: expanded substance abuse module. St Louis: Authors; 1987. [Google Scholar]
- Rousseeuw PJ, Croux C. Alternatives to the median absolute deviation. J Am Stat Assoc. 1993;88:1273–83. [Google Scholar]
- Squeglia LM, Spadoni AD, Infante MA, Myers MG, Tapert SF. Initiating Moderate to Heavy Alcohol Use Predicts Changes in Neuropsychological Functioning for Adolescent Girls and Boys. Psychol Addict Behav. 2009;23:715–22. doi: 10.1037/a0016516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tenke CE, Kayser J. A convenient method for detecting electrolyte bridges in multichannel electroencephalogram and event-related potential recordings. Clin Neurophysiol. 2001;112:545–50. doi: 10.1016/s1388-2457(00)00553-8. [DOI] [PubMed] [Google Scholar]
- Ullsperger M, Danielmeier C, Jocham G. Neurophysiology of Performance Monitoring and Adaptive Behavior. Physiol Rev. 2014;94:35–79. doi: 10.1152/physrev.00041.2012. [DOI] [PubMed] [Google Scholar]
- Vaidyanathan U, Vrieze SI, Iacono WG. The Power of Theory, Research Design, and Transdisciplinary Integration in Moving Psychopathology Forward. Psychol Inq. 2015;26:209–30. doi: 10.1080/1047840X.2015.1015367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vinck M, Oostenveld R, van Wingerden M, Battaglia F, Pennartz CMA. An improved index of phase-synchronization for electrophysiological data in the presence of volume-conduction, noise and sample-size bias. Neuroimage. 2011;55:1548–65. doi: 10.1016/j.neuroimage.2011.01.055. [DOI] [PubMed] [Google Scholar]
- Vrieze SI, Hicks BM, Iacono WG, McGue M. Decline in Genetic Influence on the Co-Occurrence of Alcohol, Marijuana, and Nicotine Dependence Symptoms From Age 14 to 29. Am J Psychiatry. 2012;169:1073–81. doi: 10.1176/appi.ajp.2012.11081268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Welch KA, Carson A, Lawrie SM. Brain Structure in Adolescents and Young Adults with Alcohol Problems: Systematic Review of Imaging Studies. Alcohol Alcohol. 2013;48:433–44. doi: 10.1093/alcalc/agt037. [DOI] [PubMed] [Google Scholar]
- Wilson S, Malone SM, Thomas KM, Iacono WG. Adolescent drinking and brain morphometry: A co-twin control analysis. Dev Cogn Neurosci. 2015;16:130–8. doi: 10.1016/j.dcn.2015.07.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winter WR, Nunez PL, Ding J, Srinivasan R. Comparison of the effect of volume conduction on EEG coherence with the effect of field spread on MEG coherence. Stat Med. 2007;26:3946–57. doi: 10.1002/sim.2978. [DOI] [PubMed] [Google Scholar]
- Zucker RA, Heitzeg MM, Nigg JT. Parsing the Undercontrol-Disinhibition Pathway to Substance Use Disorders: A Multilevel Developmental Problem. Child Dev Perspect. 2011;5:248–55. doi: 10.1111/j.1750-8606.2011.00172.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.