

Abstract
The development of neuron cells in vagal nerve nuclei in medulla oblongata was studied in vitro in live newborns and stillborns from different cases. Morphological changes were studied in respiratory nuclei of dorsal motor centre (DMNV) and nucleus tractus solitarius (NTS) in medulla oblongata. The material from medulla oblongata was fixated in 10μ buffered formalin solution. Fixated material was cut in series of 10μ thickness, with starting point from obex in ± 4 mm thickness. Special histochemical and histoenzymatic methods for central nervous system were used: cresyl echt violet coloring, tolyidin blue, Sevier-Munger modification and Grimelius coloring. In immature newborns (abortions and immature) in dorsal motor nucleus of the vagus (DMNV) population stages S1, S2, S3 are dominant. In neuron population in vagal sensory nuclei (NTS) stages S1, S2 are dominant. In more advanced stages of development of newborns (premature), in DMNV stages S3 and S4 are seen and in NTS stages S2 and S3 are dominant. In mature phase of newborns (maturity) in vagal nucleus DMNV stages S5 and S6 are dominant, while in sensory nucleus NTS stages S4 and S5 are dominant. These data suggest that neuron population in dorsal motor nucleus of the vagus (DMNV) are more advanced in neuronal maturity in comparison with sensory neuron population of vagal sensory nucleus NTS. This occurrence shows that phylogenetic development of motor complex is more advanced than the sensory one, which is expected to take new information’s from the extra uterine life after birth (extra uterine vagal phenotype)
Keywords: medulla oblongata, DMNV, NTS
INTRODUCTION
Synapse forming is the most important process during neurogenesis. Synaptic contacts are a product of interaction between endogenous factors (genetic) and exogenous factors (time and space). Synapses are formed in the beginning of the third month of pregnancy. The most intensive period of synaptogenesis is between the 13th and 24th week, that is why the period is considered critical in development (organisation) of cortex and basal ganglia, because the cortex is very vulnerable (1, 2, 3, 4). In the beginning of the 4th month of foetal life many nerve fibres begin to take a white appearance due to myelin depositing, which is formed from repeated coiling of Schwan cells membrane around the axon. Myelin sheath which surrounds nerve fibres in spinal-cord has a very different origin, as it is formed from oligodentroglial cells. Although myelinisation of nerve fibres in spinal-cord starts approximately in the 4th month of intrauterine life, some of nerve fibres, which descend from higher brain centres in spinal-cord don’t myelinise until the first year after birth. Nervous system tracts myelinise approximately when they start to function (2). Analysing the development of talamus nuclei in human foetus, in vitro, using Nissl coloring method and based on basophyllic level, nuclei methachromasia, cytoplasm coloring, nucleocytoplasmatic index and the occurrence of Nissl substance, nerve cell differentiation can be divided in 6 stages (5, 6).
Stage S-1. Germinative cell level is not differentiated. In this early stage cell nuclei are closed, round, with granular material, dense chromatin, basophilic cytoplasm which can’t be seen.
Stage S-2. The stage of chromatin material segregation. Larger nuclei, with brighter coloring, chromatin material are accumulated in 4-5 basophilic granules. Cytoplasm still can’t be seen.
Stage S-3. A stage of new neuroblasts with uncoloured cytoplasm. The size of nuclei grows, nucleolus is visible in form of metachromatic granules, and basophilic chromatin disappears, while its residue goes in periphery. Cytoplasm still can’t be seen.
Stage S-4. Neuroblasts stage with semilunar cytoplasm. A narrow coloured semilunar cytoplasm appears which is visible in half of the nucleus and later in all of it.
Stage S-5. The stage of new neuron. The size of the nucleus increases and especially the size of cytoplasm. In cytoplasm, Nissl substance starts to appear as small disperse granules.
Stage S-6. The stage of partially matured neuron. Granules of Nissl substance appear to be larger and stand out as protuberance; axon rise can be seen (7).
Using optical morphological criteria for nerve cells, such as: nucleus metachromasia, cytoplasm colouring, nucleocytoplasmatic index with histochemical and histoenzymatic methods, the ontogenesis of neuronal cells in live newborns and dead newborns from different causes, in different weeks of gestation was studied in vitro in this research. The stages of nerve cell development were differentiated in these newborns (S1, S2, S3, S4, S5, and S6).
MATERIAL AND METHODS
In this research, 23 cases were included. The material was obtained from autopsy of newborns, live-born and stillborns from different cases, in different weeks of gestation. The grouping of cases was done in this order (Table 1):
TABLE 1.
The stages of development of nerve cells in nuclei population of DMNV and NTS in medulla oblongata in dependence with weight.

Material selected for research was obtained from medulla oblongata. In medulla oblongata horizontal cuttings were made, with initial orientation from the obex in the height ± 4 mm, where the projection of dorsal vagal nuclei is done (dorsal motor nucleus of nervus vagus and nucleus of tractus solitarius). The material obtained from medulla oblongata was fixated in buffered 10 % solution of formalin. These research methods were used: histochemical methods - hematoksiline and eosin coloring, cresyl echt violet colouring for nervous and glial cells, cresyl echt violet colouring for Nissl substance, argyrophyle granules coloring (Grimelius), Servier-Munger modification for nerve endings colouring. Preparative cuttings were made in microtome and kriotom in 7 μ and 10 μ.
RESULTS
Morphologic investigation was made in serial tissue cut-outs in medulla oblongata, starting from obex with diameter of ± 4 mm. In continuous manner in horizontal cut-outs the visualisation of vagal nuclei was done (DMNV, NTS). The stages of development of nerve cells in the population of nervus vagus nuclei are shown in the following figures (Figure 1-8):
FIGURE 1.
Horizontal serial cut in medulla oblongata, starting from obex. Population of neurons in nucleus DMNV and NTS in newborns; (H&E; 100 x).
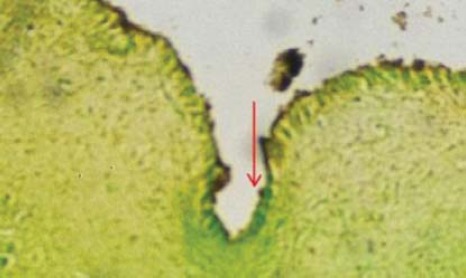
FIGURE 2.

Horizontal serial exit in obex level (arrow) in abortus. Migratory alignment of new neuroblasts is seen; (H&E; 400 x).
FIGURE 3.

Population of neurons in stages SI andS2 in nucleus DMNVin newborns; (Cresyl violet 400 x).
FIGURE 4.

Population of neurons in nucleus DMNV stages SI, S2, S3 of development in newborns; (PTAH; 400 x).
FIGURE 5.

Stage S4 of neuroblasts in vagal nudei in newborns; (LuxolfestbluMBS;400x).
FIGURE 6.

Stages S5 and S6 of cell development in matures; (PTAH;400x).
FIGURE 7.

Pronounced cholinoreactivity in neuron populations in nucleus of NTS in medulla oblongata, in matures; (Indoxyl HAT and ACHE; 400 x).
FIGURE 8.

Migration ofcholinoreacüve neuroblasts along blood vessel in neuron population level in DMNV in newborns; (Indoxyl HAT and ACHE; 400 x).
DISCUSSION
Respiratory control is a result of complex interactions between neurons and brain stem. Three neuronal groups ensure normal breathing: dorsal respiratory group (nucleus dorsalis nervus vagus-DMNV, nucleus tractus solitarius-NTS), ventral respiratory group (Botzinger complex, nucleus retroambigualis and nucleus ambigus), pons respiratory group (KUiker-Fuse complex and nucleus parabrachialis medialis). Differentiated neurons of these groups are responsible for the activity of respiratory cycle. Coordination of these neurons in group manner generates respiratory rhythm (8). In our examined material, in immature newborns (abortus and immature) in population of dorsal motor nucleus of the vagus (DMNV), stages S1, S2, S3 are dominant. In population of neurons of vagal sensory nucleus (NTS) stages S1, S2 dominate. In more advanced stages of newborns (prematurity), in DMNV, stages S3 and S4 are seen, while in NTS stages S2 and S3 dominate. In mature phase of newborns (maturity) in population of vagal nucleus DMNV stages S5 and S6 dominate and in sensory nucleus NTS stages S4 and S5. Histoenzymatic cholinoreactivity of choline-acetyl- transferase (HAT) and acetylcholynesterase (ACHE) is positive in neuron populations of dorsal vagal motor nucleus and sensory vagal nucleus (DMNV and NTS). Argentafine and argyrophilic reaction does not manifest with colouring in these nuclei. Argyrophilic reaction is visibly positive in ventricular ependyma of fourth ventricular base. Chromaffin reaction for adrenaline, noradrenalin and dopamine does not mark DMNV and NTS nuclei (9).
In early stages of nerve cells development in motor and sensory nerve populations DMNV and NTS of early foetal ages, enzyme activity HAT and ACHE is visibly pronounced (stages S1, S2, S3). In matures stages S5 and S6 with smaller enzyme activity is seen. It is characteristic for nervous system that neurons from different parts migrate in higher levels of nervous system. Neurons migrate in two directions, in radiate and tangentially. It is thought that one of the mechanisms of migration of new neurons is along glial radial fibres. In the beginning of development nervous cells migrate 1 mm per 24 hours, while in later stages of neurogenesis for migration of cortical neurons almost two weeks are needed (10, 11, and 12). In our material we can see perivascular neuronal migration in respiratory neuron population. It is mentioned by other authors as a possible mechanism of migration (11). Neuron differentiation includes processes in which structural changes are made, biochemical maturation, development of biochemical properties, synthesis of specific transmitters, structural changes of neurons, such as: increase of nervous surface, which is achieved by growth of cell body, dendritic branching, axon growth and development of synaptic contacts (13, 14). In vitro, population of neurons in dorsal vagal motor nucleus (DMNV) is more mature than the population of sensory neurons in vagal sensory nucleus NTS. This occurrence shows that phylogenesis of motor complex is more advanced than the sensory component, which after birth is expected to receive new information from extra uterine life (extra uterine vagal phenotype). Similar data were seen by other authors (3, 13, 14).
CONCLUSION
Based on the results obtained from morphologic examination of respiratory organs in vitro in newborns in different stages of gestational development, we have reached these conclusions:
-Population of neurons in dorsal vagal motor nucleus (DMNV) is more mature than the population of sensory neurons in vagal sensory nucleus NTS. This occurrence shows that phylogenesis of motor complex is more advanced than the sensory component, which after birth is expected to receive new information from extra uterine life (extra uterine vagal phenotype).
-Chromaffin reaction for adrenaline, noradrenalin and dopamine does not mark DMNV and NTS nuclei.
-Enzyme activity HAT and ACHE is visibly pronounced (stages S1, S2, S3). In matures stages S5 and S6 with smaller enzyme activity is seen.
-Glial cells in motoric nucleus are also more present in immature newborns, with a tendency of gradual decline in more advanced ages.
-Average diameter of vagal nerve cells in dorsal vagal motor nucleus DMNV and sensory vagal nucleus NTS continually raises, starting from immature, premature and matures where neuronal developmental stage (S6) is reached, stage of a partially mature neuron.
REFERENCES
- 1.Erlichmana JS. Nicotinic receptors mediate spontaneous GABA release in the rat dorsal motor nucleus of the vagus. Neuroscience Aug. 1997;79:671–681. doi: 10.1016/s0306-4522(97)00026-2. [DOI] [PubMed] [Google Scholar]
- 2.Goodwin BP, Anderson GF, Barraco RA. Characterization of muscarinic receptors in the rat NTS. Neurosci Lett May. 1995;19:131–135. doi: 10.1016/0304-3940(95)11563-3. [DOI] [PubMed] [Google Scholar]
- 3.Miguel MP. Neuropathologic correlates of perinatal asphyxia. Int. Pediatr. 2000;15(4):221–228. [Google Scholar]
- 4.Kenney HC, Armstrong DD. Perinatal Neuropathology. In: Graham DI, Lantos PL, editors. Greenfield’s Neuropathology. London: Arnold Publushers; 1997. [Google Scholar]
- 5.Fitzgerald JT. Basic and Applied. Tindal. Vol. 122. London: 1985. Neuroanatomy; pp. 146–148. [Google Scholar]
- 6.Olszewski J, Baster D. Karger. Vol. 98. London: 1985. Cytoarchitecture of the human brain stem; pp. 122–125. [Google Scholar]
- 7.Seeley RR, Stephens DT, et al. Anatomy & Physiology. Missouri: Mosby; 1992. pp. 369–380. 522-525. [Google Scholar]
- 8.Berry PA. Spontaneus synaptic activites in rat nucleus tractus solitarius neurons in vitro evidence for excitatory processing. Brain Res. 1993;630(1-2):125–135. doi: 10.1016/0006-8993(93)90650-c. [DOI] [PubMed] [Google Scholar]
- 9.Mouton PR, et al. Stereologycal leng estimation using spherical probes. J. Microscopy. 2002;206:30–35. doi: 10.1046/j.1365-2818.2002.01006.x. [DOI] [PubMed] [Google Scholar]
- 10.Muton PR. Baltimore and London: The Johns Hopkins University Press; 2002. Principles and practices of unbiased stereology: an introduction for bioscientis. [Google Scholar]
- 11.Sawchenko PE. Central and peripheral mechanisms. Alan R. Liss, Inc; 1987. Anatomic and biochemical specificity in antral autonomic pathways In: Organization of the ANS; pp. 267–281. [Google Scholar]
- 12.Islami H, Šukalo A, Shabani R, Disha M, Kutllovci S. Examination of ontogenetic-morphologic growth of cholinergic receptor system in isolated preparation of human trachea in vitro. Med. Arh. 2006;60(1):13–17. [PubMed] [Google Scholar]
- 13.Islami H, Shabani R, Haliti N, Bexheti S, Rozafa K, Raka D, Šukalo A, Dauti H. Examination of ontogenetic-morphologic growth of adrenergic receptor system in isolated preparation of human trachea in vitro. Med. Arh. 2008 in process. [PubMed] [Google Scholar]
- 14.Islami H, Shabani R, Haliti N, Bexheti S, Rozafa K, Raka D, Šukalo A, Izairi R, Dauti H, Qehaja N. In vitro examination of degerative evolutuion of adrenergic nerve endings in pulmonary inflammatory processes in newborns. Bos J. of basic Med. Sci. 2008;3:220–225. doi: 10.17305/bjbms.2008.2921. [DOI] [PMC free article] [PubMed] [Google Scholar]