

Abstract
There is a persistent and growing clinical need for readily-available substitutes for heart valves and small-diameter blood vessels. In situ tissue engineering is emerging as a disruptive new technology, providing ready-to-use biodegradable, cell-free constructs which are designed to induce regeneration upon implantation, directly in the functional site. The induced regenerative process hinges around the host response to the implanted biomaterial and the interplay between immune cells, stem/progenitor cell and tissue cells in the microenvironment provided by the scaffold in the hemodynamic environment. Recapitulating the complex tissue microstructure and function of cardiovascular tissues is a highly challenging target. Therein the scaffold plays an instructive role, providing the microenvironment that attracts and harbors host cells, modulating the inflammatory response, and acting as a temporal roadmap for new tissue to be formed. Moreover, the biomechanical loads imposed by the hemodynamic environment play a pivotal role. Here, we provide a multidisciplinary view on in situ cardiovascular tissue engineering using synthetic scaffolds; starting from the state-of-the art, the principles of the biomaterial-driven host response and wound healing and the cellular players involved, toward the impact of the biomechanical, physical, and biochemical microenvironmental cues that are given by the scaffold design. To conclude, we pinpoint and further address the main current challenges for in situ cardiovascular regeneration, namely the achievement of tissue homeostasis, the development of predictive models for long-term performances of the implanted grafts, and the necessity for stratification for successful clinical translation.
Introduction
Cardiovascular tissues, such as heart valves and blood vessels, are sophisticated dynamic tissues that can grow and adapt their structure according to the hemodynamic environment in which they function. It is this characteristic quality that makes it notoriously challenging to replace these tissues with artificial substitutes in case of end-stage disease or damage. For small-diameter arteries (e.g., peripheral arteries, arteriovenous shunts) and the semilunar heart valves in particular, the development of living, adaptive replacement tissues could greatly improve the underachieving current artificial replacements. Novel cardiovascular tissue engineering (TE) strategies are increasingly moving from an in vitro to an in situ approach. In situ TE is defined as biomaterial-induced endogenous regeneration directly in the tissue’s functional site, or in situ, starting from readily-available, resorbable grafts that gradually transform into an autologous, homeostatic replacement tissue with the ability to repair, remodel, and grow. Grafts for in situ TE can be of biological or synthetic nature and either acellular or on-the-fly preseeded. However, prerequisite for this approach is that the graft is readily-available for implantation and that the graft allows for colonization and remodeling by host cells in order to achieve an adaptive autologous tissue over time. Whereas the traditional TE dogma comprises labor-intensive and lengthy in vitro culture and conditioning phases, in situ TE has been proposed as a more cost-effective and on-demand approach, using relatively simple and shelf-ready grafts.1–3
The approach of in situ TE is built on the notion that the natural inflammatory response can be harnessed to induce endogenous tissue regeneration (Fig. 1). The resorbable immunomodulatory scaffold provides a temporary microenvironment, which functions as an instructive road map for endogenous cells to infiltrate and create new, living, and functional tissue. It is hypothesized that, upon implantation, the scaffold provides support for mature tissue formation and adequate mechanical properties to withstand the hemodynamic loads. Over time, the scaffold should slowly resorb, ultimately resulting in a purely biological structure which has the ability to repair, remodel, and grow. Proof-of-concept has been demonstrated by milestone studies describing the endogenous regeneration of small-diameter blood vessels and heart valves using acellular synthetic scaffolds or de novo engineered decellularized extracellular matrix (ECM) (see Table 1). Despite these encouraging reports, we are only just beginning to grasp a more fundamental understanding of the biomaterial-driven regeneration in the complex hemodynamic environment, and many key questions remain to be answered:
How can a scaffold recapitulate the complex layered architecture of cardiovascular tissues?
What is the influence of biomechanical stimuli on the organization of new tissue?
Is it necessary to incorporate exogenous bioactivities to enhance the robustness of the in situ TE approach?
How fast and by which mechanism should a scaffold degrade?
Is this patient-dependent and should this be personalized?
How can we predict inflammatory-driven functional regeneration?
Fig. 1.

Overview of the different stages of in situ tissue regeneration, going from a synthetic, biodegradable bare construct toward a viable substitute (a). Although many aspects underlying in situ regeneration remain unknown, it is hypothesized that the stages mirror the natural phases of the wound healing response (b), starting with the inflammatory phase, characterized by the infiltration of immune cells and the formation of a preliminary matrix. The subsequent proliferative phase is characterized by a secondary influx of immune and tissue producing cells, extracellular matrix (ECM) deposition, angiogenesis and (re-)endothelialization of the construct. Tissue homeostasis is restored after a remodeling phase of the newly formed ECM and the resolution of inflammation. Photographs of heart valves adapted from;33 photographs of vascular grafts courtesy of Renée Duijvelshoff
Table 1.
Selection of recent in vivo studies applying in situ tissue engineered vascular grafts
| Primary research target | Material | Design | Model | Main findings | Ref. |
|---|---|---|---|---|---|
| Mechanisms of in situ regeneration | PGA + P(CL-LA) | Large pore (45 µm) non-woven mesh + BMC in fibrin | Mouse (IVC) | In situ regeneration is an inflammation-driven process. Preseeded BMC have paracrine function via MCP-1 | 19 |
| PGA or PLA + P(CL-LA) | Large pore non-woven mesh ± BMC in fibrin | Mouse (IVC or IA) | Temporal characterization of ECM formation, remodeling and resulting biomechanical properties in venous and arterial circulation | 159–161, 189 | |
| PCL + fibrin ± MCP-1 | Electrospun grafts shielded with GoreTex sheaths | Rat (AA) | Circulatory cells have regenerative capacity, which can be boosted by MCP-1 | 36 | |
| Degradation | PCL/Chitosan | Electrospun nanofibrous grafts | Mouse (IA), Sheep (CA) | Fast degradation of chitosan as “sacrifical material” helps regeneration | 190 |
| PLA-PGA or PLA + P(CL-LA) | Microfibrous graft + nanofibrous outer layer | Mouse (IA) | Fast degradation prevents calcification in contrast to slow degrading grafts | 162 | |
| PCL | Micro/nano-fibrous grafts | Rat (AA) | Insufficient regeneration and calcification at long-term (18 months) follow-up | 131 | |
| PGS/heparin + PCL sheath | Porous PGS/heparin core with dense non-woven PCL sheath | Rat (AA) | Timely degradation and mechanotransduction important for regeneration | 22 | |
| Micro-structure | PLA-PLCL or PLA | Large-pore (30 µm) vs. small-pore (5 µm) grafts | Mouse (IA) | Large-pore grafts prevent calcification in comparison to small-pore grafts | 191 |
| PLCL + PLA | Large-pore (29 µm) vs. small-pore (13 µm) PLCL core with PLA reinforcement | Mouse (IA) | No effect of pore size in terms of cell infiltration, macrophage polarization and tissue formation. No aneurysms | 192 | |
| PCL/PEO | Three-layered graft with a large-pore (26 µm) inner layer with dense outer and thin dense inner layers | Rabbit (CA) | Porous middle layer leads to better cell infiltration, faster degradation and improved regeneration, proposedly due to increased MCP-1 and VEGF | 193 | |
| PCL | Large-pore (30 µm) microfbrous vs. small-pore (2 µm) nanofibrous grafts | Rat (AA) | Large pores promote M2 macrophage polarization, enhanced cell infiltration and regeneration | 194 | |
| PCL | Electrospun nano-/microfibrous bilayered grafts | Rat (AA) | Layered grafts are a tool to control cell infiltration and regeneration | 116 | |
| PCL | Inner layer of circumferentially aligned microfibers + random nanofibrous outer layer | Rat (AA) | Contact guidance by aligned fibers stimulates native-like SMC organization | 195 | |
| Mechanical properties | TPU | Electrospun, microfibrous graft | Rat (AA) | Thin-walled graft with adequate biomechanical properties | 196 |
| PCL/elastin | Multi-layered electrospun graft | Rabbit (CA) | Native-like mechanical properties using recombinant elastin | 104 | |
| PGS + PCL sheath | Porous PGS reinforced by PCL sheath with varying densities | Rat (AA) | Variation of the outer sheath density directly influences regeneration (SMC organization and M2 polarization) | 197 | |
| PGA + P(CL-LA) | Non-woven porous mesh | Mouse (IVC) + in silico model | Biomechanical diversity between 12 and 24 months of implantation due to variations in the ratio collagen type 1/3 | 177 | |
| Proof-of-principle | PLA or PGA + P(CL-LA) | Porous graft reinforced with PLA or PGA mesh | Human (TCPC) | First clinical trials. Graft stenosis identified as primary mode of failure | 16–18 |
| Supramolec-ular polyester | Electrospun microfibrous grafts | Human (TCPC) | Ongoing clinical trial. Feasibility and safety up to 12 months in pediatric patients. | 21 | |
| PGS + PCL sheath | Porous electrospun PGS core with PCL reinforcement | Mouse (IA) | Long-term (1 year) functionality in arterial circulation in mice, luminal enlargement | 198 | |
| PCL | Electrospun microfibrous grafts | Rat (AA), Pig (CA) | Feasibility of regenerative PCL grafts in arterial circulation vs. ePTFE graft | 130, 199 | |
| Hyaluronan-based graft | Coated onto rotating steel core | Rat (AA), Rat (IVC), Pig (CA) | Feasibility of regenerative hyaluronan grafts in arterial and venous circulation | 200–202 | |
| In vivo engineered autologous tissue capsule | Fibrocellular matrix created by controlling the FBR to subcutaneously implanted rods | Pig (CA) | Control of FBR to create fibrocellular vascular grafts with sufficient mechanical strength | 203, 204 | |
| Decellu-larized SIS | Heparin + VEGF functionalized | Sheep (CA) | Successful in situ regeneration in arterial circulation in large animal | 205 | |
| EC/SMC preseeded with fibrin | Sheep (CA) | Successful in situ regeneration in arterial circulation in large animal; cell pre-seeding beneficial but not required | 206 | ||
| Decellulariz-ed de novo engineered allograft | In vitro tissue-engineered vascular graft from human cells, decellularized | Baboon (AV access) | Grafts maintain functionality in a high flow environment, suitable for hemodialysis. | 207 | |
| Decellulariz-ed de novo engineered allograft | In vitro tissue-engineered vascular graft, decellularized | Growing lamb (PA) | Grafts show somatic growth and maintain functionality up to ~1 year follow-up | 20 | |
| Comorbidities | PCL ± RGD | Nanofibrous grafts | Rat (AA), healthy vs. type II diabetic | Increased complications and impaired regenerative capacity in diabetic vs. healthy rats | 184 |
| Graft functiona-lization | UPy-based elastomers | Microfibrous grafts functionalized with UPy-SDF-1α peptide or UPy-PEG + RGD | Rat (AA) | Control over specific cell recruitment and adhesion using supramolecular building blocks | 148, 208 |
| PLCL + heparin/ SP | Microfibrous grafts with covalently bound SP and/or heparin | Rat (subcu-taneous) | Recruitment of MSC and enhanced M2 polarization by heparin/SP functionalization | 209, 210 |
PGA polyglycolic acid, PCL poly(ε-caprolactone), PLA poly(L-lactic acid), MCP-1 monocyte chemoattractant protein-1, PGS poly(glycerol sebacate), SIS small intestine submucosa, PLCL poly (L-lactic co-ε-caprolactone), PEO poly(ethylene oxide), TPU thermoplastic polyurethane, RGD arginine–glycine–aspartic acid, UPy ureido-pyrimidinone, SP substance P, BMC bone marrow-derived cells, VEGF vascular endothelial growth factor, EC endothelial cells, SMC smooth muscle cells, FBR foreign body response, SDF-1α stromal cell-derived factor-1α, PEG poly(ethylene glycol), IVC inferior vena cava, IA infrarenal aorta, AA abdominal aorta, CA carotid artery, TCPC total cavo-pulmonary connection, AV arteriovenous, PA pulmonary artery, ECM extracellular matrix, ePTFE expanded polytetrafluoroethylene, MSC mesenchymal stromal cells
To address these questions, this review provides the state-of-the-art across scientific disciplines regarding the physical, biochemical and biomechanical environmental cues involved in biomaterial-driven in situ tissue regeneration; insights that are essential for the rational design of new, robust immunomodulatory scaffolds for in situ cardiovascular TE. Notwithstanding the promising results obtained with decellularized biological scaffolds, we focus our attention primarily on synthetic-based scaffolds. Synthetic scaffolds have the inherent advantage of being produced under controlled processes, and thus bypassing any reproducibility issues associated with biological scaffolds, as well as being fully tailorable. Therein we take an interdisciplinary point of view, taking lessons from regenerative biology, biomaterials science, immunology, and mechanobiology. To conclude we address the imminent challenges in the field of cardiovascular in situ TE: achieving tissue homeostasis, the development of appropriate preclinical models, and the considerations with respect to stratification for clinical translation.
Cardiovascular TE—progressing from in vitro to in situ
Recapitulating the sophisticated biomechanics-dictated cardiovascular microstructures
For both heart valves and blood vessels, the preferred replacement option is a living, adaptive graft. Although the various cardiovascular target tissues (semilunar heart valves, small-diameter arteries, arteriovenous shunts) all come with their own specific requirements, exposure to the hemodynamic environment is the common denominator that poses a major challenge for tissue engineered constructs. Contact with blood implies the need for an endothelium or a surrogate non-thrombogenic layer to prevent thrombus formation, a common complication in synthetic substitutes. Moreover, the hemodynamic loads (e.g., cyclic strains, shear stresses) define the tissue microstructure, leading to a highly-organized and layered structure. For heart valves in particular, it is well-recognized that the valve’s three-layered microstructure (e.g., fibrosa, spongiosa, and ventricularis) is essential to maintain life-long mechanical function.4, 5 Many strategies have been followed to engineer such tissues in vitro, as excellently reviewed in more detail elsewhere.6–8 However, reproducing the sophisticated native microstructure and its inherent mechanical function has proven to be highly challenging, resulting in varying success. To avoid the complexity of in vitro culture, cardiovascular TE is increasingly progressing towards in situ strategies, relying on the endogenous regenerative capacity of the body. The approach of in situ TE for cardiovascular application is not necessarily new. Already in the 1970’s, reports by Schoen et al. and Sparks describe the exploitation of the foreign body reaction (FBR) to cylindrical implants to create vascular grafts composed of fibrous capsule tissue.9, 10 Although the regenerative process in this strategy does not occur directly in situ, these studies demonstrate the potential of FBR-driven endogenous tissue regeneration. By now, this notion has evolved into various strategies to induce endogenous regeneration directly in situ, and the proofs-of-concepts for both vascular and valvular replacements have been achieved, as described in the following.
In situ tissue engineering of vascular grafts
Among the first reports of vascular in situ tissue engineering is a series of studies by Van der Lei and colleagues, who attempted the use of a biodegradable polyurethane-based scaffold for direct in situ arterial regeneration.11, 12 These grafts often developed aneurysms due to lack of mechanical strength of the scaffolds and the authors postulate that pre-seeding with either pre-cultured endothelial cells (EC) or smooth muscle cells (SMC) would be beneficial to accelerate the regenerative process, essentially deviating from the in situ approach as we defined it here. The necessity for pre-seeding of cells into synthetic grafts to stimulate in situ regeneration is still a source of active debate, and the underlying mechanisms remain largely unknown. Pioneering work by the groups of Shin’oka and Breuer using on-the-fly preseeding with bone marrow-derived mononuclear cells (BM-MNC) revealed that vascular neotissue arises from the ingrowth of EC and SMC from the neighboring blood vessel wall and that, contrary to the classic tissue engineering paradigm, preseeded cells do not contribute directly to the vascular neotissue, but instead rapidly disappear after implantation.13, 14 Furthermore, vascular neotissue formation was proven to be a host macrophage-mediated regenerative process, with cell seeding being not essential for vascular tissue formation.15 Ultimately, the first clinical trials using BM-MNC were shown to improve neotissue formation for replacements of large-bore venous conduits using biodegradable composite polymer scaffolds.16–18 However, seeded BM-MNCs were no longer detectable within a few days of implantation, proving that they primarily ameliorated the regenerative process by paracrine signaling to recruit host immune cells, rather than terminally differentiating into functional tissue cells.19 Despite the promising clinical outcomes described in these clinical studies, stenosis remained the most prevalent graft-related complication in this low-pressure application.18 More recently, Syedain et al. demonstrated proof of somatic growth of an in situ TE pulmonary artery replacement based on decellularized de novo engineered ECM in lambs.20 With respect to acellular synthetic grafts, ongoing clinical trials initiated by Xeltis BV demonstrate promising initial results up to 12 months follow-up, using highly porous electrospun grafts from a resorbable supramolecular polyester for total cavo-pulmonary connection in pediatric patients.21 Although longer follow-up is warranted, these results are indicative of the clinical potential of in situ TE grafts in the low pressure circulation. Application in the less forgiving high-pressure systemic circulation remains challenging, with aneurysm formation being the most common complication. As underlined in the landmark study by Wu et al., scaffold compliance and appropriate mechanotransduction are particularly important properties for the regeneration of arterial grafts.22 Many reports on in situ TE of vascular grafts have contributed to our understanding of the regenerative processes and the influence of the numerous scaffold and host parameters that play a role in this, as summarized in Table 1.
In situ tissue engineering of heart valves
With respect to heart valves, the most compelling results to date have been achieved using acellular biological scaffolds, such as decellularized porcine small-intestine submucosa,23, 24 decellularized allografts,25–27 and de novo engineered decellularized ECM.28, 29 Decellularized native grafts have been used in various clinical trials and results indicate that these are suitable base materials for in situ TE.26 Decellularized de novo engineered valves have been proposed as an alternative to negate the need for a donor valve.30 Results by Driessen-Mol et al. demonstrate the feasibility of implanting such valves using minimally invasive delivery techniques leading to extensive recellularization with host cells upon implantation as pulmonary valve replacement in sheep.29 However, shortening of leaflets and progressive insufficiency was reported in this study. The Tranquillo group reported the progressive insufficiency of decellularized in vitro engineered valves when implanted as pulmonary valves in a growing lamb model.31 In this study, the reported insufficiency was attributed to growth of the valvular root and rather than leaflet shortening. A recent study by the same group demonstrates the sustained functionality of similar valves up to 6 months in the aortic position in sheep.28 Interestingly, these valves demonstrated recellularization patterns with vimentin-positive and α-smooth muscle actin (α-SMA)-negative cells, progressing from the root towards the free edge of the valve leaflet, strongly suggesting that these cells originate from the neighboring arterial tissue. Moreover, cellularization mainly occurred on the arterial side of the valve leaflet, while cellularization on the ventricular side was sparse.
To date, experiences of in situ heart valve TE using synthetic-based scaffolds is limited. Weber et al. reported on the implantation of a fast-degrading scaffold based on polyglycolic acid, on-the-fly pre-seeded with BM-MNC, in non-human primates.32 However, the rapid degradation of the scaffold did not allow for sufficient neo-tissue formation to maintain functionality. In collaboration with our clinical partners, we demonstrated the proof-of-concept of using a synthetic scaffold for in situ heart valve TE by using a tunable supramolecular elastomer.33 These valves demonstrated sustained functionality up to 12 months when implanted in the pulmonary position of sheep, and reported preliminary results demonstrate the compatibility with minimally invasive delivery. In this study, we observed spatiotemporal cellularization patterns similar to the aforementioned results of Syedain et al.,28 indicative of gradual regeneration of the valve starting from the hinge region towards the free edge of the leaflet. Prior experience by our group has elucidated that the valve leaflets are acutely colonized with circulatory cells within 1 day of implantation (unpublished data), suggesting that these cells provide the initial cue for neighboring tissue cells to migrate into the valve. Importantly, in our study the degradation of the synthetic scaffold material was observed to be highly localized and was most pronounced in regions with extensive cellularization and neo-tissue formation.33 This suggests that the degradation is cell-driven and correlated to ECM formation, which would mean that the structural integrity of the valves is warranted at all times, although further research is needed to validate this. Using derivative technology, Xeltis BV has recently initiated the first clinical trials using an electrospun resorbable supramolecular polymer as pulmonary valve replacement in pediatric patients (XPlore-I and XPlore-II, NCT numbers: NCT02700100, NCT03022708). However, long-term functionality and growth potential of these in situ TE valves remains to be proven, and translation to the clinically most relevant aortic position is all but trivial given the harsh hemodynamic loads in the systemic circulation. Hence, it is important to get a more fundamental understanding of the endogenous regenerative processes and the cues that can be incorporated in a scaffold to modulate these.
In situ TE - summoning the natural regenerative potential
Inflammation as the driver of regeneration
The in situ TE approach relies on the regenerative capacity of the body. This natural regenerative capacity is highly species-dependent, and, in adult mammals, scarring is the default repair mechanism in response to trauma rather than functional regeneration. Upon synthetic scaffold implantation, disruption of the tissue structure and subsequent cell damage will initiate an inflammatory response by the host. This is the onset of the classical phased wound healing cascade, culminating in an FBR to the material, as expertly reviewed in detail elsewhere.34, 35 Briefly, at the very early stages of implantation, blood-biomaterial interactions lead to the adsorption of endogenous proteins from blood or interstitial fluid to the biomaterial surface. The so formed provisional matrix is rich in mitogens, chemoattractants, cytokines and growth factors which control the subsequent phases of the wound healing and FBR. Following the provisional matrix formation and within the first days after implantation, acute inflammation occurs, with an instantaneous influx of innate immune cells, predominantly neutrophils and monocytes. Chronic inflammation develops as inflammatory stimuli persist at the implant side, with macrophages controlling the microenvironment in cross-talk with lymphocytes, as well as secondary cells, such as fibroblasts, and various stem and progenitor cells, mirroring the cascade of normal wound healing (Fig. 1). Depending on the scaffold properties, the end-stage FBR is characterized either by the accumulation of foreign body giant cells, fibrous encapsulation of the scaffold and eventual graft failure, or by a diminished numbers of macrophages, marked increase in tissue resident cells, and tissue remodeling toward an organized and functional regenerated tissue.
The presence of inflammation can be considered ambivalent; on one hand essential for wound healing, while on the other hand detrimental if any of the phases is disturbed, leading to chronic inflammation, excessive scar tissue formation, and eventual graft failure. Indeed, inflammation is believed to play an all-determining role in biomaterial-driven tissue regeneration. Roh et al. postulated that the in situ regeneration of the inferior vena cava in mice occurred via an inflammation-mediated cascade.19 A recent study by our group applying a shielded porous scaffolds in the abdominal aorta of rats revealed a similar phased regenerative cascade, and it was demonstrated that by modulating the initial inflammatory response in the first days after implantation, late-term tissue development was remarkably enhanced 3 months downstream.36 Using an immunodeficient mouse model, Hibino et al. proposed that platelet activation and the involvement of Natural Killer cells are critical factors for adverse graft remodeling, leading to vascular graft stenosis.37 These results are illustrative of the causality between the initial inflammatory response to an implant and downstream tissue outcome, and show the potential of modulating the long-term regenerative process via the initial scaffold properties.
The plastic macrophage as a target for immunomodulation
Macrophages have been identified as the commanding cellular mediators in the regenerative process. In several animal studies, macrophage depletion resulted in a significant delay of re-epithelialization, decreased collagen deposition, and impaired healing.38 Remarkably, in regenerating species such as salamanders, systemic depletion of macrophages led to failure of growth after limb amputation, while by replenishing the endogenous macrophage population the full regenerative capacity was restored.39 Similarly, in injured arteries of rabbits and rats40 and in tissue-engineered vascular grafts of mouse models,15 systemic macrophage depletion blocked the production of EC, SMC and, ultimately, vascular neotissue.
The functional role of the macrophage is determined by its phenotypic and functional plasticity.41, 42 Rather than being purely “big eaters”, inflammatory cells involved in phagocytosis and pathogen clearance, the current consensus is that macrophages are highly plastic cells that adapt their function depending on the tissue microenvironment. As such they can reversely polarize into functionally distinctly different phenotypes, being the so-called pro-inflammatory “M1” phenotype or the pro-wound healing “M2” phenotype. Within the tissue microenvironment, the complex integration of tissue-specific signals, microbial factors, and soluble mediators determines phenotypic changes, and differential activation of these cells. It is now widely accepted that under the crucial influence of soluble mediators secreted by type 1 or type 2 T helper cells (TH1 and TH2, respectively), macrophages polarize toward the M1 or M2 phenotypes, respectively. More recently a general scheme for macrophage polarization was proposed, based on three different homeostatic activities—host defense, wound healing and immune regulation. It was proposed that M1 and M2a,b,c phenotypes are extremes of a much broader spectrum of functional states43 with overlapping M1–M2 characteristics.44 Depending on their polarization state, macrophages mediate the formation and remodeling of new tissue by secreting essential growth factors and cytokines that either promote or inhibit functional tissue formation (e.g., transforming growth factor-β, TGF-β; tumor necrosis factor-α, TNF-α; matrix metalloproteinases; platelet-derived growth factor, PDGF).42, 45 Besides this regulatory role, it was recently acknowledged that macrophages also directly contribute to tissue formation via producing ECM components themselves, including fibronectin, tropoelastin, various types of collagen and glycosaminoglycans (GAGs) and proteoglycans, such as versican.46–48 Moreover, indications for transdifferentiation of macrophages toward the mesenchymal lineage and vice versa can be found in different species and pathologies, but that remains a subject of ample debate.49–51 Most important for in situ TE is the notion that the balance of M2/M1 macrophages infiltrating an implanted biomaterial is predictive for long-term tissue outcome.52 Consequently, modulation of the macrophage phenotype has become a valuable target to steer the inflammatory response toward functional regeneration.
Multipotent cells in cross-talk with the inflammatory environment
Supporting the macrophage during the wound healing process, a secondary influx of tissue producing cells, including mature fibroblasts and EC as well as various (circulating) stem/progenitor cells, of both mesenchymal and hematopoietic origin, governs ECM production and remodeling (Fig. 2).49, 53, 54 For cardiovascular in situ TE, the origin of mature tissue cells is unknown so far. Although ingrowth of cells from neighboring tissues has been proposed as the predominant route of cellularization in vascular grafts,14 its significance for the human situation has been contested.55 Given that human cardiovascular tissues have a low natural regenerative potential and a low cellular turnover, regeneration of such tissues is likely to be dependent on the recruitment of cells from high-turnover sites, such as the blood and the bone marrow. Bone marrow-derived progenitors are known to contribute to the valvular interstitial cell population in healthy adult heart valves and homing of progenitors is a normal homeostatic process.56–58 The peripheral blood contains various progenitor cell populations, such as endothelial progenitor cells (EPC) and smooth muscle progenitor cells, such as fibrocytes. Although the participation of circulating CD34+ progenitors in regeneration is topic of active debate, blood stream-derived cellularization of vascular grafts was proven in dogs, with patchy endothelial coverage with underlying α-SMA+ SMCs.59, 60 More recently, we demonstrated the regenerative potential of circulatory cells in a rat model of arterial regeneration with a suggested role for CD34+ progenitors.36 Table 2 summarizes the relevant cell types for in situ TE and their proposed role in the regenerative process.
Fig. 2.
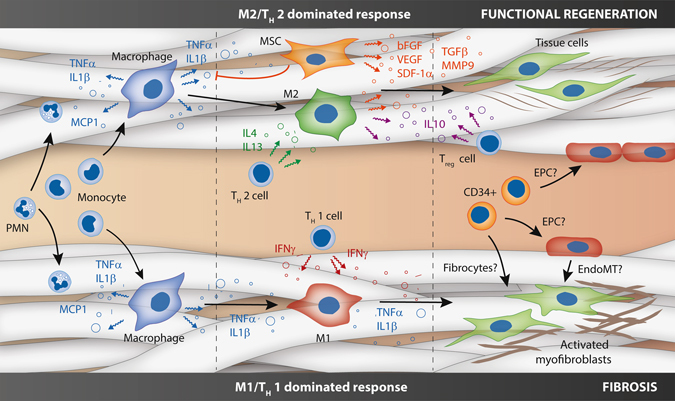
Cartoon of the various (simplified) cell-cell interactions in in situ tissue engineering, as hypothesized based on the state-of-the-art. After the instantaneous response of protein adhesion and platelet activation (not depicted), circulating polymorphonuclear cells and monocytes are recruited to an implanted scaffold in response to various chemokines (e.g., monocyte chemoattractant protein (MCP)-1). Upon activation, the monocytes give rise to macrophages in the scaffold, which are a source of pro-inflammatory factors (e.g., tumor necrosis factor (TNF)-α, Interleukin (IL)‐1β). Depending on the scaffold properties, this is followed by an M1/TH 1 cell dominated response pro-inflammatory response (bottom) of an M2/TH 2 cell dominated pro-regenerative response (top). The former is characterized by the prolonged presence of M1 macrophages, instigated by TH1 cell-secreted pro-inflammatory cytokines, such as interferon-γ. Recruited fibroblasts typically acquire an activated phenotype, producing non-functional cross-linked fibrous scar tissue. In contrast, the pro-regenerative process is dominated by M2 macrophages under influence of TH2 cell secreted cytokines (e.g., interleukin (IL)-4 and -13). Mesenchymal stromal cells play an important immunomodulatory role by inhibiting pro-inflammatory factors, such as TNF-α, as well as secreting numerous trophic factors (e.g., basic fibroblast growth factor, bFGF; vascular endothelial growth factor, VEGF; stromal cell-derived factor-1α, SDF-1α; transforming growth factor β, TGF-β; matrix metalloproteinase 9, MMP-9). This biochemical milieu attracts tissue cells and modulates the formation and remodeling of well-organized functional neotissue. Upon scaffold degradation, T reg cells inhibit the inflammatory process by secretion of, e.g., IL-10. Homing of circulatory CD34+ progenitor cells, such as fibrocytes and endothelial progenitor cells, as well as endothelial-to-mesenchymal transformation may contribute to cellularization and pathophysiological neotissue formation, although these processes are topic of active debate
Table 2.
Overview of cell types involved in in situ TE and their proposed functions
| Cell type | Function | Via | |
|---|---|---|---|
| Granulocytes | Neutrophils | Immunomodulation | Release of immunomodulatory factors |
| Eosinophils | Biomaterial degradation | Phagocytosis and enzyme/radical excretion | |
| Basophils | |||
| Monocytes | Classical (CD14 +/ CD16−/CCR2 + ) | Immunomodulation | Antigen presentation (e.g., T-cell activation) |
| intermediate (CD14 + /CD16 + /CCR2++) | Production of immunomodulatory cytokines | ||
| non-classical (CD14dim/CD16+/CCR2−) | Differentiation into macrophages or dendritic cells | ||
| Macrophages | Host-defense | Immunomodulation | Antigen presentation (e.g. T-cell activation) |
| Wound healing | Biomaterial degradation | Immunomodulatory cytokine production | |
| Immune regulation | Tissue formation and remodeling | Phagocytosis and enzyme/radical secretion | |
| FBGCs formation | |||
| Production of trophic factors (e.g., TGF-β, VEGF) | |||
| Secretion of ECM components | |||
| Potential transdifferentiation into myofibroblast-like cells | |||
| Dendritic cells | Immunogenic | Immunomodulation | Efficient antigen presentation |
| Tolerogenic | Immunomodulatory cytokine secretion | ||
| Mast cells | Immunomodulation | Production of pro-inflammatory cytokines (e.g., IL-1β, TNF-α, MCP-1) | |
| Tissue formation and remodeling | Secretion of proteases and anti-inflammatory cytokines (e.g., IL-10, IL-4) | ||
| Lymphocytes | T-cells | Immunomodulation | Activation of macrophages to kill antigen bearing cells |
| B-cells | Activation of B-cells to produce antibodies | ||
| NK cells | Promoting macrophage-biomaterial adhesion | ||
| Stimulation of FBGC formation | |||
| Tissue formation and remodeling | Influencing macrophage polarization and cytokine production | ||
| Mesenchymal stem cells (CD73+/CD90+/CD105+/CD14-/ CD34-/CD45-) | Immunomodulation | Production of mediators to diminish immune cell activation (e.g., IL-10) | |
| Tissue formation and remodeling | Production of trophic factors (e.g., bFGF, SDF-1α) | ||
| Differentiation into tissue cells | |||
| Endothelial progenitor cells (CD34+/VEGFR+/CD133+) | EOCs (myeloid origin) | Tissue formation and remodeling | Production of stimulatory factors for re-endothelialisation and vascularization (e.g., VEGF) |
| ECFCs (non-myeloid) | Formation of endothelial cells | ||
| Potential differentiation into myofibroblast-like cell (e.g., EndoMT) | |||
| Smooth muscle progenitor cells | CD14+/CD105 + cells | Immunomodulation | Production of immunomodulatory cytokines and growth factors |
| Fibrocytes (CD34+/CD45+/Col I+) | Tissue formation and remodeling | Production of ECM proteins | |
| Secretion of remodeling factors (e.g., MMP’s) | |||
FBGC foreign body giant cell, TGF-β transforming growth factor, VEGF vascular endothelial growth factor, ECM extracellular matrix, IL interleukin, TNF-α tumor necrosis factor alpha, MCP-1 monocyte chemoattractant protein, NK cell natural killer cell, bFGF basic fibroblast growth factor, SDF-1α stromal cell-derived factor 1 alpha, EOCs early-outgrowth cells, ECFCs endothelial colony-forming cells, EndoMT endothelial-to-mesenchymal transition, MMP matrix metalloproteinase, Col I collagen type I
Irrespective of cell origin, the interactions between cells in the local scaffold microenvironment are pivotal in the regenerative process (Fig. 2). Local inflammatory reactions contribute to successful regeneration, setting the “soil” for colonizing stem cells, either endogenously recruited or seeded, at the site of injury, as recently reviewed by Forbes and Rosenthal.61 In this line, Ballotta et al. described the synergistic expression of trophic factors (e.g., basic fibroblast growth factor, bFGF; stromal cell-derived factor (SDF)-1α) by human mesenchymal stromal cells (MSCs) when activated by peripheral blood mononuclear cells in a 3D electrospun scaffold in hemodynamic conditions.62 Moreover, MSCs have been described to exert strong immunomodulatory functions by inhibiting the secretion of inflammatory factors, such as TNF-α and interferon-γ by TH1 cells, as well as increasing the secretion of pro-regenerative factors, such as IL-4 and IL-10, by TH2 and Treg cells.63 Various studies employing in vitro co-cultures have unraveled paracrine signaling mechanisms between macrophages and fibroblasts or SMCs, both in 2D and in 3D scaffolds.64–68 Interestingly, Song et al. demonstrated that the macrophage phenotype has an effect on the fibrous matrix production of human fibroblasts via the secretion of either pro-fibrogenic (e.g., TGF-β1, PDGF) or anti-fibrogenic and fibrolytic factors (e.g., TNF-α, MMP-7).69 Using 3D in vitro co-culture, McBane et al. proposed that monocyte-derived macrophages induce a shift in SMC phenotype form a more synthetic, migratory phenotype to a resting, contractile phenotype.70
To conclude, an implanted scaffold serves as an artificial microenvironment that can boost the regenerative capacity of tissue sites with a naturally low regenerative potential, by modulating (1) the local inflammatory environment and (2) the recruitment of stem and progenitor cells to the scene.
Modulating the regenerative response in the hemodynamic environment
Rational scaffold design
Recent reviews have elaborately described the fundamental potential of modulating the immune response, and more specifically macrophage polarization, using biomaterial properties.71, 72 To apply these principles to in situ cardiovascular TE, we should consider the heterogeneity of the infiltrating populations and the interplay between these cells in the mechanically and biochemically dynamic environment. Consequently, in-depth insights are required on the single and combined temporal effects of environmental cues on cell behavior. Rational scaffold design requires interdependent, multi-scale considerations, including micro-mechanical and macromechanical properties, architecture, degradation rate, and bioactivity (Fig. 3).2
Fig. 3.

Schematic representation of the design strategies that can be employed to tailor resident cell behavior inside the graft. The transfer of hemodynamic loads (a) can be tuned via adaptations in material properties, such as the mechanical properties, geometry (b) and microstructure (d). Concurrently, cell behavior is defined by interdependent microstructural parameters (e.g., fiber diameter, alignment, pore size and topography; (d) and biochemical parameters (e.g., surface chemistry and bio-activation); (c). However, local loads and scaffold parameters change in time due to material degradation (e) and new tissue formation. Subfigures b and e are adapted from refs. 107 and 126, respectively
Biomechanical stimuli
The biomechanical environment plays an important role in cardiovascular development. Hemodynamic loads dictate the structural composition of valves and vessels, both in physiology and pathology.73, 74 For artificial grafts, the continuous exposure to cyclic pressures, shear stresses, and strains require an excellent fatigue behavior of a graft, in particular for heart valves. Moreover, the biomechanical environment dominantly influences the process of cell infiltration and tissue regeneration during in situ TE. To illustrate, implantation of a synthetic graft in the abdominal aorta of mice resulted in a well-organized ECM with native-like elastin content, while a similar graft resulted in only very sparse elastin formation after long-term implantation in the venous circulation.75, 76 Transfer of the biomechanical loads (e.g., shear stress, strain) on a cardiovascular scaffold to the infiltrating cells is dependent on the scaffold properties, such as geometry, microstructure, and mechanical properties. Moreover, the local loads change during scaffold degradation and tissue formation. By specifically tailoring these scaffold properties for the aspired clinical application, the hemodynamic load transmission to residing cells can be tuned in order to direct tissue regeneration towards homeostasis (Fig. 3).
Shear stress
Shear stresses play an important role in cardiovascular regeneration by affecting cell adhesion, activation and signaling. Moreover, fluid dynamics strongly influence developmental processes, such endothelial-to-mesenchymal transformation,77, 78 which are highly relevant for in situ TE strategies, as expertly reviewed elsewhere.79 At the cellular level, in vitro studies using 2D fluidic devices have demonstrated shear stress-dependent adhesion of various vascular cells (i.e., SMC, EC, and EPC) to various substrates in controlled shear flow.80–83 Shear stresses also have profound effects on the immune response. Increasing shear forces are known to influence platelet activation and adhesion,84 as well as the integrin-mediated adhesion and apoptosis of leukocytes.85, 86 Correspondingly, in vitro studies in our lab demonstrated that shear stress overruled the effect of biochemical stimulation on specific human monocyte recruitment into a 3D electrospun scaffold loaded with MCP-1.87
In vivo, the wall shear stress is notorious determinant of adverse remodeling. Aberrant wall shear stresses at the venous-arteriovenous synthetic graft anastomotic side are known to contribute to neointimal hyperplasia and early graft stenosis.88 Interestingly, differential healing of various grafts in dogs was suggested to be implant site-dependent due to variations in fluid dynamics.89 Taken together, these results highlight the importance of shear stress for in situ TE. For artificial heart valves, the shear stress profile depends on the valve’s geometrical design,90, 91 and as such represents an important design criterion.
Cyclic strain
Traditional in vitro TE studies have given us tremendous insights into the causal relationship between strains and ECM formation in 3D scaffolds. Strain is known to be a potent mediator of collagen production and turnover.92, 93 Gupta et al. proposed that GAGs and proteoglycan synthesis by valvular cells is dependent on cyclic strain.94 Recent studies highlight the importance of physiological strains on the synthesis and organization of the fibrous ECM, and specifically the maturation of elastic fibers.95, 96 Importantly, progenitor cells, such as EPCs and MSCs, have been shown to proliferate and differentiate into the cardiovascular cell lineage in response to strain, although the underlying processes of strain-driven differentiation are not well-understood.97, 98
For in situ TE, it is important to consider that strains also have an impact on the immunological environment. Straining of monocytes/macrophages was shown to contribute to monocyte-to-macrophage differentiation, a different inflammatory gene expression profile and the selective augmentation of related cytokines, matrix metalloproteinases, and scavenger receptors.99, 100 Interestingly, Matheson et al. demonstrated that macrophages display a differential response to uniaxial strain compared to biaxial strain.101 Moreover, they showed that cyclic biaxial strain may contribute to biomaterial degradation via augmentation of enzymatic activities (e.g., increased esterase production).102 In 3D culture, Ballotta et al. observed that moderate amounts of biaxial strain (7%) may contribute to a more anti-inflammatory macrophage phenotype (M2) when compared to high strains (12%).103 In a recent study, Battiston et al. demonstrated that 10% cyclic stretch on a co-culture of monocytes/macrophages and vascular SMC in a polyurethane scaffold contributed to synthesis of collagen type I and III and resulted in improved mechanical tissue properties (elastic modulus, tensile strength).48
The fundamental effects of strain on tissue regeneration directly translate to in vivo data of in situ TE vascular grafts. Numerous studies have stressed the importance of graft compliance for successful elastogenesis and tissue regeneration and reorganization.22, 104, 105 For electrospun meshes, the macromechanical properties can be predicted by multi-scale modeling, taking into account microstructural elements such as cross-link density, fiber alignment, and fiber diameter.106 The resulting strain distribution in the scaffold is determined by the microstructural organization in combination with the macroscopic geometry.107, 108 Multi-scale numerical models represent a valuable enabling tool to predict and tune local strains that will be experienced by recruited cells as a result of the hemodynamic loads.
Scaffold microstructural and material properties
Apart from determining the mechanical behavior, the physical properties of the scaffold have a profound influence on the behavior of colonizing cells. The scaffold microstructure is defined by multiple interdependent parameters, such as the fiber diameter and alignment, pore size, and surface topography and chemistry, and these parameters change during degradation of the scaffold. All these factors are known to have imposing effects on the cellular response to the scaffold, for example, in terms of infiltration, adhesion, and the immunogenic response.
Microstructure
The microstructural parameters of fibrous scaffolds provide essential physical cues, such as contact guidance, to infiltrating cells. Using an electrospun scaffold with aligned microfibers, De Jonge et al. demonstrated that newly formed collagen by human myofibroblasts was deposited in the direction of fiber alignment and that the contact-guiding cue overruled the effect of biomechanical stimuli until approximately 80% of the scaffold was resorbed.109 This suggests that the formation and organization of neotissue can be directed from the get-go by incorporating anisotropy in the scaffold, such as proposed by Sohier et al.110 Interestingly, Fioretta et al. observed a differential effect of electrospun microfibers with varying fiber diameter on cell alignment and ECM deposition between human EPC and mature EC, suggesting that the effect of contact guidance is cell type dependent.111
Apart from contact guidance, the microstructure has profound effects on cellular infiltration and the FBR. For electrospun scaffolds, which are most often used in cardiovascular TE, the fiber diameter is linearly correlated to the pore size. As such, fiber diameter and alignment directly influence the cell infiltration depth into the scaffold.112–114 Consequently, multi-layered scaffolds have been suggested not only to control the mechanical behavior,115 but also to control cell infiltration116, 117 (see Table 1). Additionally, it has been shown that electrospun nanofibers minimize blood activation and reduce macrophage activation when compared to microfibers.118, 119 Several studies revealed that the mechanism behind which porous materials have improved healing might involve a shift in the polarity of macrophages at the implant site.120, 121 Along this line, a positive correlation has been established between M2 polarization and pore size, with an increased expression of angiogenic factors (vascular endothelial growth factor (VEGF), bFGF, TGF-β) with increasing pore size.122 Madden et al. proposed a specific “sweet spot” in the scaffold’s microstructural pore size (20–40 μm) to optimally promote favorable M2 polarization.120 More specifically, it was demonstrated that this effect is associated with a change in macrophage shape. Upon identifying that M2 macrophages display an elongated, spindle-shaped morphology while M1 macrophages display a round pancake-like shape, McWhorter et al. controlled cell shape and polarization via engineered micro-patterned substrates. Remarkably, shape-induced and cytokine-induced M2 polarization occurred through distinct yet synergizing pathways, suggesting that biochemical cues compliment the effects of geometrical cues.123 Other important physical stimuli that may influence macrophage polarization include substrate stiffness, topology and surface chemistry, as recently reviewed in more detail by McWhorter et al.72
Material degradation
Material degradation will continuously alter the cellular microenvironment, as microstructure and mechanical loading will change in a time-dependent fashion. It is decisive for the success of the scaffold that functional tissue formation and degradation are well-balanced. Prolonged biomaterial presence will result in chronic inflammation and the formation of a tightly cross-linked fibrotic and calcific network that is unable to remodel. Too fast degradation, on the other hand, may result in the loss of structural integrity.
Degradable materials can be degraded within phagosomes after phagocytosis, or eroded via extracellular resorption, with or without the involvement of foreign body giant cells. After degradation, the monomeric components are removed by the natural (metabolic) pathways of the human body.124 The biodegradation mechanisms, i.e., (i) hydrolysis, (ii) oxidation, (iii) enzymatic degradation, and (iv) physical degradation are in part dependent on the chemical composition and the morphology of the biomaterial. Hong et al. described the tailored degradation rate of polyurethane scaffolds by partial substitution of polyester segments with polycarbonate segments in the polymer backbone.125 Brugmans et al. reported that supramolecular polycaprolactone (PCL)-based polymers are more susceptible to oxidative degradation in comparison to conventional PCL, which is more prone to hydrolytic and enzymatic degradation.126
Macrophages have been shown to play a pivotal role in material degradation, via the production of enzymes and reactive oxygen species that can accelerate degradation. Enzymes such as esterases and lipases endogenously produced by macrophages can accelerate the resorption process.126–128 Similarly, for the oxidative resorption pathway, macrophages with different polarization states can guide and accelerate the scaffold degradation via secretion of both reactive oxygen species and enzymes.126, 128 Understanding of the mechanisms of scaffold degradation is all but trivial, as it determines the extent to which cells (e.g., macrophages) will actively degrade scaffold material. Most importantly, scaffold degradation should be in balance with neotissue formation to maintain mechanical functionality at all times.
Taken together, there are multiple interdependent scaffold parameters, which affect the inflammatory and regenerative response. Table 3 provides an overview of suggested scaffold design principles to achieve an optimal pro-regenerative scaffold biomechanical and physical microenvironments.
Table 3.
Suggested scaffold design principles regarding physical and biomechanical microenvironment
| Parameter | May affect: | Design considerations: |
|---|---|---|
| Strain | ECM deposition | Stiffness tailored to: |
| Elastogenesis | Obtain appropriate mechanotransduction, which is essential for functional tissue regeneration | |
| GAGs production | Promote physiological, local and tissue-dependent strains for proliferation and differentiation of tissue producing cells and sustained synthesis and organization of ECM. | |
| Collagen maturation & organization | Avoid high cyclic strain levels (>8%) as this may induce a proteolytic milieu (pro-inflammatory macrophages polarization) | |
| Macrophage polarization | ||
| Shear stress | Platelet adhesion and activation | Stiffness and geometry mimicking the native counterpart to: |
| Leukocyte adhesion & activation | Promote low shear stresses and adequate cell infiltration | |
| Cell alignment | Avoid high laminar shear stresses or oscillatory shear stresses, responsible for aberrant cell proliferation, and hyperplasia | |
| Release kinetics of bioactivated grafts | Prioritization: In case of bioactivated grafts, high shear stresses might overrule the release kinetics of the bioactive compounds | |
| Fiber diameter | Cell infiltration | When compared to microfibers, nanofibers: |
| ECM deposition | Minimize blood activation | |
| Encapsulation | Reduce macrophage activation | |
| Mechanical properties | Reduce fibrous capsule thickness | |
| Better mimick the nanofibrous structure of ECM | ||
| Present an increased number of fibers per surface unit, and increased strength | ||
| Pore size | Encapsulation | Larger pore sizes (20–40 µm) are beneficial over small pore sizes (2–13 μm) for: |
| Macrophage polarization | Cell infiltration | |
| ECM deposition | Expression of angiogenic factors | |
| Polarization of macrophages into an anti-inflammatory phenotype | ||
| Prevention of calcification | ||
| Improved healing | ||
| Prioritization: | ||
| Pore size is a more critical regulator of macrophage polarization compared to the fiber diameter | ||
| Pore-size and cytokine-induced macrophage polarization occurs via distinct yet synergizing pathways. Biophysical cues presented by bio-materials may be used to compliment the effects of geometrical cues | ||
| Fiber alignment | Contact guidance to cells | Aligned fibers to be preferred over randomly aligned fibers because of the induced: |
| Contact guidance to ECM deposition | Collagen deposition via contact guidance | |
| Encapsulation | Stimulation of native-like SMC organization via contact guidance | |
| Mechanical properties | Formation of thinner fibrous capsule | |
| Layered constructs with different degree of fiber alignments as an effective tool to mimick the hierarchical organization and the variety of local strains exerted in complex native tissues | ||
| Topography | Cell adhesion | Rough surfaces to be preferred over smooth surfaces for the reduced adhesion of macrophages, FBGCs accumulation, and fibrotic capsule formation |
| FBGCs fomation | ||
| Surface chemistry | Monocyte adhesion | Hydrophilic and anionic surfaces can promote an anti-inflammatory response by: |
| Cytokine production | Down regulation of monocyte adhesion | |
| FBGCs formation | Selective production of anti-inflammatory cytokines | |
| Encapsulation | Inhibition of IL-4–mediated macrophage fusion into foreign body giant cells (FBGCs) | |
| Prioritization: In terms of fibrous capsule formation, geometric feature (fiber diameter and pore size) are a more critical regulator than topography and surface charge | ||
| Material degradation | Macrophage activation | Degradation of the scaffold tailored to: |
| Fibrosis | Obtain resorption rates compatible with ECM production | |
| Calcification | Avoid a too fast degradation, resulting in lack of adequate mechanical support and consequent tissue compaction and retraction avoid too slow degradation, which could promote prolonged macrophage activation, excessive fibrosis and calcification | |
| Implants degeneration |
ECM extracellular matrix, GAGs glycosaminoglycans, SMCs smooth-muscle cells, IL-4 interleukin-4, FBGCs foreign body giant cells
Biochemical stimuli
In several animal studies, implantation of vascular grafts in the form of cell-free, bare polymer led to remarkable formation of fully functional neovessel mimicking the native tissue,22, 129–131 posing unanswered questions on the necessity for bioactivation for tissue regeneration purposes. Nevertheless, bioactivated scaffolds which house chemotactic and/or trophic factors provide suitable biochemical and physico-chemical cues which, by mimicking the critical aspects of natural healing processes, might accelerate tissue regeneration (Table 4). Also, in patients with cardiovascular diseases, bioactivated scaffolds might augment the limited self-healing capacity by artificially accelerating the proliferation and differentiation of the recruited or implanted cells. On-the-fly preseeding of autologous bone marrow-derived cell fractions into biodegradable synthetic cardiovascular grafts stimulated in situ regeneration of autologous neovessels and valves.18, 132 Following studies revealed an interdependent effect on the secretion of trophic factors by preseeded MSC and the inflammatory environment created by circulating immune cells in vitro.62 To boost selected signaling molecules or tether endogenously released factors to promote a regenerative microenvironment, biodegradable scaffolds with controlled release of chemoattractants have been developed, which lead to enhanced infiltration of immune cells, de novo tissue formation and reduced fibrosis in several animal models.19, 36, 133, 134 Much research has been devoted to the modulation of the inflammatory response via macrophage polarization, with development of several bioactivated materials leading to successful nerve,135, 136 bone137, 138 and blood vessels regeneration,139 both in vitro and in vivo.
Table 4.
Selection of recent studies employing pre-seeded or biochemically functionalized grafts for regeneration in vitro and in vivo
| Goal | Application | In vivo model | Material | Bioactive | Approach | Main findings | Ref. |
|---|---|---|---|---|---|---|---|
| Crosstalk between preseeded cells and immune cells | Vascular regeneration | Human | PGA + P(CL/LA) | (Cell preseeding: BM-MNC) | Preseeding | No evidence of aneurysm formation, graft rupture, graft infection, or ectopic calcification. Graft stenosis as primary mode of graft failure | 18 |
| Heart valve regeneration | Non-human primates | PGA coated with P4HB | (Cell preseeding: BM-MNC) | Preseeding | Preserved valvular structures and adequate functionality up to 4 weeks post implantation. Substantial cellular remodeling and in-growth resulting in layered, endothelialized tissue | 32 | |
| Cardio-vascular regeneration | – | PCL-U4U | (Cell preseeding: BMSC, PBMCs) | Preseeding | PBMC’s secretion of a cocktail of immunomodulatory proteins; MSC’s secretion of trophic factors (MCP-1, VEGF and bFGF); synergistic upregulation of CXCL12 gene expression and bFGF protein production | 62 | |
| Vascular regeneration | Mouse | PGA + P(CL/LA) | (Cell preseeding: BMCs) | Preseeding | Early seeded hBMCs depletion; Significant secretion of MCP-1 by seeded cells; MCP-1 guided monocyte infiltration; scaffolds repopulation by SMCs and ECs | 19 | |
| Phased healing response via temporal release of exogenous growth factors | Vascular regeneration | Rat | PCL | MCP-1 | Fibrin coating | At 3 months medial layer with SMCs in an oriented collagen matrix, intimal layer with elastin fibers, and confluent endothelium | 36 |
| Angiogenesis | Mouse (subcutan-eous) | PLGA | SDF1-α | Physical adsorption/ osmotic pumps | SDF1-α guided MSC infiltration in vitro and in vivo; increased angiogenesis and reduced fibrosis in vivo | 134 | |
| Heart valve regeneration | Sheep | Bovine pericardium | SDF1-α, fibro-nectin | Coating | Improved homing of primitive cells; native-like matrix deposition; endothelial lining, and complete recellularisation after 5 months | 133 | |
| Heart valve regeneration | Dogs | Decellular-ized porcine valves | Fibro-nectin, HGF | Pre-Stirring, pressurization | Accelerated vascularization and endothelialization | 211 | |
| Vascular regeneration | Rabbit | Double layered PELCL | VEGF, PDGF | Coaxial electrospinning | Accelerated, sequential proliferation of VEC and VSMC in vitro. Enhanced revascularization and no thrombus formation in vivo | 212 | |
| Subcutaneous implantation | Rat | mPEGmA-Ac-TMPTA | Fibro-nectin-derived peptides | Covalent binding | Modulation of inflammatory response, adherent macrophage density, and FBGC formation | 213 | |
| Angiogenesis | Mouse | PLG microsphe-res | VEGF, PDGF | Encapsulation, gelification | Stable and sustained improvements in perfusion and angiogenesis | 214 | |
| Angiogenesis | Murine | Hydrogels | VEGF | Encapsulation, gelification | Stable and sustained improvements in perfusion and angiogenesis | 215 | |
| Angiogenesis | Mouse | GAG hydrogel | VEGF, Ang-1 | Non covalent mixing, crosslinking | Stimulation of localized microvessel growth at controlled rates | 216 | |
| Angiogenesis | Mouse | Hyaluronan hydrogels | VEGF, KGF | Crosslinking | Stimulation of microvessel growth; appropriate permeability and perfusion; no extravasation of red cells | 217 | |
| Angiogenesis | Rat | Dextran hydrogels | EGF, bFGF | Impregnation | Accelerated capillarization and wound healing | 218 | |
| Angiogenesis | Rats | Collagen-heparin scaffolds | FGF2, VEGF | Heparin binding | Increased production of collagen type IV and smooth muscle active; early mature vasculature formation | 146 | |
| Improved/selective cell adhesion | Subcutaneous implantantion | Mouse | PEG based hydrogels | Arg–Gly–Asp (RGD) peptide | Photopolymerization | Modulation of macrophage adhesion via selective engagement of integrins’ bindings | 219, 220 |
| Subcutaneous implantantion | Mouse | Polystyrene microparticles (in vitro); PET discs (in vivo) | Mac-1, Arg–Gly–Asp (RGD) peptide | Loading in EVA and incubation | Modulation of macrophage adhesion via selective engagement of integrins’ bindings; reduced fibrous capsule thickness | 221 | |
| Vascular regeneration | Rat | Supramol-ecular CE-UPy-PCL + UPy-PEG | - | UPy-UPy interactions (quadruple hydrogen bondings) | Reduced cell infiltration in the presence of only 10% of UPy-PEG for cell free vascular grafts. | 208 | |
| Modulation of inflamma-tory response via macrophage polarization | Vascular regeneration | Mouse | Alginate micro-particles | MCP-1, VEGF | Mixing with collagen/fibronectin gel | Promoted M2 polarization; Increased mature vessel formation via enhanced endothelial and smooth muscle cell recruitment | 141 |
| Nerve regeneration | Rat | Polysulfone/agarose hydrogel scaffolds | IL4 | Premixing | Promoted M2 polarization; accelerated downstream axon regeneration | 135 | |
| Bone regeneration | Mouse | Decellularized trabecular bone | IFN-γ, IL-4 | Physical adsorption (IFNγ), biotin-streptavidin binding (IL4) | Sequential pro-inflammatory M1—anti-inflammatory M2 polarization | 138 | |
| Bone regeneration | - | Silk films | IFN-γ, IL-4 | Covalent binding | Sequential pro-inflammatory M1 – anti-inflammatory M2 polarization | 137 | |
| Vascular regeneration | Rat | Multidomain peptide gels | MCP-1, IL-4 | Premixing | MCP-1 dependent cellular infiltration and IL-4 dictated M2 polarization | 139 | |
| Intraperitoneal implantation | Mouse | PLG microspheres | IL10 | Lentiviral vectors | Cytokine expression modulation of infiltrating leukocytes, decreased inflammation | 222 | |
| Nerve regeneration | Rat | Nanofibrous scaffolds | IL-10 | Covalent attachment | Promoted M2 polarization | 136 | |
| Vascular regeneration | Rat | Supramolecular UPy-PBU | SDF1-α-derived peptide (UPy-modified) | UPy-UPy interactions (quadruple hydrogen bondings) | Reduced expression of inflammatory markers in vitro, promoted macrophages infiltration and early in situ cellularization in vivo | 148 | |
| Subcutaneous implantation | Rat | PLCL | Heparin, SP | Covalent attachment | Recruitment of MSC and enhanced M2 polarization by heparin/SP functionalization | 209, 210 | |
| Tethering of endogenous-ly released factors | Angiogenesis | - | Heparin mimetic peptide nanofibers | VEGF, FGF-2, HGF | Heparin binding | Attachment of heparin-binding cytokines; improved cellular infiltration and angiogenesis | 144, 146, 147 |
PGA polyglycolic acid, PLGA poly(lactic-co-glycolic acid), PCL poly(ε-caprolactone), PCL-U4U polycaprolactone bisurea, PLA poly(L-lactic acid), PELCL poly(ethylene glycol)-b-poly(L-lactide-co-ε-caprolactone), P4HB poly-4-hydroxybutyrate, UPy ureido-pyrimidinone, CE-UPy-PCL chain-extended UPy-modified poly(ε-caprolactone), PEG poly(ethylene glycol), mPEGmA-Ac-TMPTA mono- methoxy polyethyleneglycol monoacrylate—mPEGmA—acrylic acid—Ac—trimethylolpropane triacrylate—TMPTA-, PLG poly(L-glutamate), PLCL poly (L-lactic co-ε-caprolactone), BMC bone marrow-derived cells, BM-MNC bone marrow-derived mononuclear cells, PBMC pheripheral blood mononuclear cells, EC endothelial cells, EPCs endothelial progenitor cells, SMC smooth muscle cells, MCP-1 monocyte chemoattractant protein-1, RGD arginine–glycine–aspartic acid, VEGF vascular endothelial growth factor, SDF-1α stromal cell-derived factor-1α, IL-4 interleukin-4, IL-10 interleukin-10, IFN-γ interferon- γ, HGF hepatocyte growth factor, PDGF platelet-derived growth factor, Ang-1 angiopoietin-1, KGF keratinocyte growth factor, EGF epidermal growth factor, bFGF basic fibroblast growth factor, FGF-2 fibroblast growth factor-2, Mac-1 macrophage-1 antigen, SP substance P.
Over the last decades various bioactivation methodologies have been developed, with direct adsorption of growth factors to scaffolds and hydrogels representing the simplest approach and the first to be extensively investigated (Table 4).135, 137, 139–141 However, when growth factors are administered in their native form, they are susceptible to biodegradation, inactivation due to a very short half-life, and insufficient delivery at the active site. In the search for methods overcoming these disadvantages, encapsulation of growth factors via non-covalent and covalent immobilization were investigated. These delivery systems hold a great deal of promise for localized administration, providing a solubilising and protective environment, minimizing the release to non-target sites, and serving also as an artificial ECM for cell penetration.130, 140, 142 Natural occurring GAGs have been identified to bind and modify inflammatory factors, with heparan sulfate being the most well- recognized natural binding site for several cytokines.35, 143, 144 In this respect, heparin-conjugated scaffolds145, 146 and heparin-mimetic peptide nanofibers144, 147 have been proven to indirectly boost cellular infiltration and angiogenesis. Ultimately, recent advances in supramolecular polymers allowed for the development of truly “smart”, cell-responsive scaffolds which can interact with their environment and mediate the host response to the biomaterial, reduce inflammation and promote early in situ re-cellularization.148
Current challenges
The primary goal for in situ cardiovascular TE is to recapitulate the complex structure and function of the native tissue, in a state of quiescent homeostasis (Fig. 4). Therein, the challenge is not to induce tissue formation, yet the regeneration of functional tissue. This can be achieved by presenting the optimal inflammatory, physical, and biomechanical microenvironments for the colonizing cells. These local microenvironments are constituted by the initial scaffold parameters in the hemodynamic environment, but will change over time due to scaffold degradation and the formation of new tissue. Temporal understanding and control of these processes is one of the main current challenges for in situ TE for cardiovascular applications. Consequently, the development of appropriate models is required in order to predict and tune the regenerative process, and to assess the robustness of the technique depending on patient demographics (Fig. 4).
Fig. 4.

a Schematic illustration displaying the three main interdependent challenges faced for successful, robust in-man application of in situ tissue engineered cardiovascular grafts. The development process is represented by a continuous feedback loop between the optimization of the graft design and the development of predictive models to understand and determine long-term in vivo performance, while taking into account graft recipient variability (e.g., age, gender, co-morbidities, and utility). b Optimization of graft design is visualized as a flowchart, in which interchangeable scaffold design parameters together with the hemodynamic loads and cells will determine if tissue homeostasis will be reached. Societal demands, including patient and physician wishes, should be taken into consideration during the (early) stages of graft development to determine the added value of these grafts for health care. HTA: Health Technology Assessment
Achieving tissue homeostasis
Tissue regeneration is considered the restoration of the original tissue function and structure. In contrast, post-natal wound healing is characterized by abundant deposition of collagen in the form of unorganized bundles within a non-functional matrix, which lacks elastic fibers and functionality: repair, rather than regeneration. The disproportionate accumulation of collagen in cardiovascular tissues, in the form of either a reactive or a reparative fibrosis, further increases stiffness. On the other hand, excessive collagen degradation can tether distortion in tissue architecture, excessive reduction in stiffness, and, ultimately, tissue rupture. For several years, research in the field has focused primarily on collagen deposition, and the possibility to develop cardiovascular grafts of suitable strength, with little regard for elasticity. However, the elastic network represents the hallmark factor distinguishing between the regeneration of functional tissue and fibrous scar tissue. Disturbances in elastin homeostasis have been pinpointed as the underlying causes of valvular grafts failure,149, 150 and aneurysma formation.151 Although elastin production during cardiovascular in situ TE has been reported, the homeostatic restoration of the native-like, organized elastic network during by adult endogenous cells is a major challenge. This is due to the proteolytic inflammatory milieu, which inhibits elastin expression,152 as well as the influence of hemodynamic loads on elastic network formation.95, 96, 153 While being essential for the development and remodeling of neotissue, the hemodynamic environment also provides a persistent cue for fibrosis.154 Many signaling proteins that are essential to tissue regeneration, such as MCP-1 and TGF-β, are also stimulatory factors for fibrosis. Similarly, on the cellular level, M2 macrophages and fibrocytes, for example, are correlated to both regeneration and fibrosis.155 This poses a paradoxal challenge, and the difference between physiological de novo tissue formation and excessive fibrosis is dependent on a rather delicate balance of factors. Developmental principles can provide important lessons for in situ TE. The growth factor profiles in embryonic and post-natal healing are very different,156, 157 with embryonic wounds displaying markedly reduced numbers of inflammatory cells compared to adult wounds, due to deficiencies in leukocyte infiltration. Martin et al. proposed that not the macrophage presence itself, but rather the macrophage polarization state influences the balance between scarring and healing, in accordance with previous literature.158
In vivo observations emphasize that timely resolution of inflammation is critical. In a recent study, Naito et al. characterized the time course of ECM development in an in situ TE venous graft in mice.159 They observed an initial surge in the production of fibrillary collagen, which was postulated to be part of an initial cellular FBR to isolate the polymer. This response was alleviated upon degradation of the scaffold, after which other ECM proteins, such as GAGs, elastin, and collagen type IV increased proportionally. As a result, the mechanical properties of the graft converged to those of the native vein.160 This was attributed to the transient changes in mechanotransduction of hemodynamic loads to the cells. In correspondence, delayed degradation and lack of mechanotransduction was pinpointed as the cause of insufficient tissue regeneration in arterial grafts in mice.161 Similarly, appropriate mechanotransduction and timely degradation of the scaffold were hypothesized to be the key factors underlying the successful in situ regeneration of a neoartery in rats.22 Accordingly, long-term follow-up of arterial PCL grafts in rats revealed extensive chondroid metaplasia, which was likely caused by the persistent presence of the PCL scaffold and poor mechanotransduction, up to 18-month follow-up.131 Sugiyura recently demonstrated that calcification of arterial grafts in mice could be avoided in fast-degrading grafts, in contrast to slow-degrading grafts.162 Correspondingly, Wu et al. advocated the importance of rapid resorption and appropriate mechanotransduction for the remodeling of fast degrading synthetic grafts toward physiological-like neoarteries.22
In summary, all these results corroborate that degradation is inextricably correlated to tissue formation and remodeling. The timely resolution of inflammation is a pivotal factor in the process of de novo tissue formation and the prevention of adverse remodeling. This resolution is governed by the biochemical and biomechanical microenvironment, which may be concerted by well-timed degradation of the synthetic scaffold (see also Table 3).
Development of predictive models
Perhaps one of the most underexposed challenges for in situ cardiovascular TE is the development of appropriate models to predict the long-term in vivo performance of grafts. When employed appropriately, animal models are a vital source of information to study and mimic the complex regenerative processes in vivo. Small animals (mice, rats) in particular are being used extensively to answer preliminary research questions in the developmental phase of new prostheses (e.g., regarding biocompatibility, material choice, and device design). However, although the bulk of animal studies for in situ vascular regeneration reports on promising patency and endothelialization rates, both the underlying mechanisms and the timescales are subject to strong interspecies variations, as well as implant-site variations,89, 163 which may mislead their conclusions in terms of clinical application. It is often overlooked that the predictive value of an animal model is only as useful as the context in which it is interpreted.164 For example, the mechanisms governing the cellularization of the graft with functional tissue cells (i.e., EC and SMC) differ between species. Endothelialization of vascular grafts in mice and rats is characterized by a rapid, progressive transanastomic overgrowth,14 which is typically not observed in humans, even after prolonged implantation periods. Therefore, refined animal models have recently been proposed by ourselves36 and others.165 Differences in critical hemodynamic parameters (e.g., shear stress) between different animal models and humans may further decrease the value and predictability of animal models.166
Another important consideration in the use of animal models is the immunological variance between species, especially considering that the process of in situ TE is highly dependent on the immune response to the implanted biomaterial. It has been demonstrated that the genomic response to acute inflammatory stimuli in mice poorly correlates to the human conditions, as well as between different mouse models.167 Genomic comparison of mice and human monocyte subpopulations has revealed that, although general expression patterns are conserved, significant and even opposing functional differences exist between species.168 Similarly, the characterization of the subsets based on marker expression is species-dependent. This calls for species-specific immunological marker panels to study the function of specific leukocyte populations in vivo.
Novel in vitro and in silico models that aim to predict the biomaterial-dependent host response are starting to gain attention as of recent.113, 169–171 Various recent studies defined the macrophage response and the resulting cytokine/chemokine profile (e.g., IL-6, TGF-β, TNF-α, MMP-9 secretion) as predictive parameters for the long-term host response on a biomaterial.170, 172, 173 In a recent study, Wolf et al. evaluated the static in vitro response of human peripheral blood mononuclear cells toward various synthetic and biological materials in vitro in terms of macrophage M1/M2 ratio and secretion of a small set of signaling proteins (MMP-2, MMP-9, IL-6, and interferon-γ-induced protein (IP)-10). Coupled with in silico principle component analysis, the in vitro data could be successfully correlated to the long-term tissue outcomes of the same materials implanted subcutaneously in a animal model.170 Enayati et al. highlighted the potential of a fibroblast-macrophage coculture model to increase the predictive value of in vitro models.174 Adding physiological complexity, bioengineered vascular tissues may serve as suitable in vitro models, as recently reviewed by Wolf et al.175
Taken together, advanced in vitro and in silico models mimicking the principal components of the host response to an implanted biomaterial or even “humanized” animal models in synergy with systematic in vitro studies, might represent an excellent and potentially superior alternative to animal models.
Stratification for clinical translation
Apart from aforementioned inter-species differences, patient-to-patient variability poses a challenge for stratification of in situ TE therapies. Even without considering any graft parameters, in situ TE is completely dependent on the natural regenerative potential of the graft recipient. The wound healing response, and thereby the intrinsic regenerative capacity are highly variable between patients, and even between healthy individuals. Mammalian wound healing is prone to genetic variability.176 Khosravi et al. who recently reported significant functional diversity in the long-term remodeling of identical arterial grafts, even in healthy laboratory animals.177 There is a strong natural variability in both the innate and adaptive immune response among humans, which, to a large extent, can be attributed to differences in age or gender.178–181 Consequently, young people and pre-menopausal women are known to have an increased risk of scarring than men and elderly, which is attributable to variations in the immune system.156 Common comorbidities of cardiovascular patients include for example diabetes and chronic kidney failure, further contributing to the variability of the regenerative capacity and thereby, the applicability of such techniques for specific patient cohorts. Krawiec et al. reported an increased risk of stenosis in vascular grafts engineered from human cells from diabetic patients, which was suggested to be due to a reduced remodeling capacity.182, 183 Wang et al. reported a significantly reduced regenerative capacity of diabetic rats.184 Together, these findings highlight the importance of risk stratification and the potential need for a personalized approach in translating these therapies to the clinic. Moreover, these are important considerations when selecting the appropriate patient cohort for first-in-man studies. Given the limitations of current alternative treatments, in situ TE would be most beneficial for children and young adults. Hence, pulmonary valved conduits for children with complex congenital heart disease would represent a most valuable first-in-man target, as is the case for the currently ongoing clinical trials by Xeltis BV.
Moreover, to translate toward in-man clinical application, it is important to remain focused on the clinical requirements, the wishes and needs of patients and physicians (cardiothoracic surgeons) as well as the societal demands. Given the societal demand for better, sustainable and more efficient health care it is important to search for strategies that have a higher return of investment and a reduced time to the market. Early Health Technology Assessment (HTA) is emerging as a research field focusing on the evaluation of medical, economic, social and ethical implications of a new medical device to determine the added value for health care.185, 186 As an example, conceptual models can be developed to determine the graft requirements to become cost-effective compared to golden standard treatment methods.187 Hence, these assessments can be used to continuously prioritize and guide design choices during development, to optimally introduce and use the grafts in the clinic. For example, patients with a limited natural regenerative capacity might benefit from the local incorporation of bioactive factors into an in situ TE scaffold (Table 4), and/or complementary systemic therapies, such as drugs to initiate stem cell mobilization.188
Conclusion
In situ TE of blood vessels and heart valves using resorbable synthetic grafts is rapidly progressing and first clinical trials are exemplary of its clinical potential. Here we described the underlying principles of biomaterial-driven regeneration, initiated by the host response to the material and governed by the interplay of immune cells (e.g., macrophages), stem/progenitor cells and tissue cells in the scaffold microenvironment. In enhancing our multi-disciplinary understanding of the fundamental processes underlying successful endogenous regeneration, it is important to consider the interdependent role of physical and biochemical cues in this process, hinging around the biomechanical cues exerted by the hemodynamic environment. The development of advanced predictive models will contribute to stratification of in situ cardiovascular TE as a robust clinical therapy.
Data availability
Data sharing not applicable to this article as no data sets were generated or analyzed during the current study.
Acknowledgements
This project has received funding from the European Union’s Seventh Framework Program for research, technological development and demonstration under grant agreement no 604514. We also acknowledge the support from the Netherlands Cardio Vascular Research Initiative: The Dutch Heart Foundation, Dutch Federation of University Medical Centres, the Netherlands Organization for Health Research and Development and the Royal Netherlands Academy of Sciences.
Author contributions
All authors were involved in the preparation, drafting, critical revision, and final approval of the manuscript.
Competing interests
Prof. Bouten is shareholder of Xeltis BV. Ms. Wissing, Ms. Bonito and Dr. Smits declare no potential competing financial interests.
Footnotes
Tamar B. Wissing and Valentina Bonito contributed equally to this work
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations
References
- 1.Mol A, Smits AIPM, Bouten CVC, Baaijens FPT. Tissue engineering of heart valves: advances and current challenges. Expert. Rev. Med. Devices. 2009;6:259–275. doi: 10.1586/erd.09.12. [DOI] [PubMed] [Google Scholar]
- 2.Bouten CVC, et al. Substrates for cardiovascular tissue engineering. Adv. Drug. Deliv. Rev. 2011;63:221–241. doi: 10.1016/j.addr.2011.01.007. [DOI] [PubMed] [Google Scholar]
- 3.van Loon, S. L. M., Smits, A. I. P. M., Driessen-Mol, A., Baaijens, F. P. T. & Bouten, C. V. C. The Immune Response in In Situ Tissue Engineering of Aortic Heart Valves. Calcific Aortic Valve Disease (ed. Aikawa, E.) 207–245 (2013).
- 4.MacGrogan, D. et al. How to make a heart valve: from embryonic development to bioengineering of living valve substitutes. Cold Spring Harb. Perspect. Med. 4, a013912 (2014). [DOI] [PMC free article] [PubMed]
- 5.Schoen FJ. Evolving concepts of cardiac valve dynamics: the continuum of development, functional structure, pathobiology, and tissue engineering. Circulation. 2008;118:1864–1880. doi: 10.1161/CIRCULATIONAHA.108.805911. [DOI] [PubMed] [Google Scholar]
- 6.Fioretta, E. S., Dijkman, P. E., Emmert, M. Y. & Hoerstrup, S. P. The future of heart valve replacement: recent developments and translational challenges for heart valve tissue engineering. J. Tissue Eng. Regen. Med. doi:10.1002/term.2326 (2016). [DOI] [PubMed]
- 7.Cheung, D. Y., Duan, B. & Butcher, J. T. Current progress in tissue engineering of heart valves: multiscale problems, multiscale solutions. Expert Opin. Biol. Ther. 15, 1155–1172. doi:10.1517/14712598.2015.1051527 (2015). [DOI] [PMC free article] [PubMed]
- 8.Pashneh-Tala, S., MacNeil, S. & Claeyssens, F. The tissue-engineered vascular graft—past, present, and future. Tissue. Eng. Part. B. Rev. 22, ten.teb.2015.0100 (2015). [DOI] [PMC free article] [PubMed]
- 9.Schoen FJ, Normann SJ, Brunswick RA, Diacoff GR. Can a small blood vessel prosthesis be derived from heterologous foreign body reactive tissue? J. Biomed. Mater. Res. 1979;13:149–154. doi: 10.1002/jbm.820130115. [DOI] [PubMed] [Google Scholar]
- 10.Sparks CH. Die-grown reinforced arterial grafts: observations on long-term animal grafts and clinical experience. Ann. Surg. 1970;172:787–794. doi: 10.1097/00000658-197011000-00003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.van der Lei B, Nieuwenhuis P, Molenaar I, Wildevuur CR. Long-term biologic fate of neoarteries regenerated in microporous, compliant, biodegradable, small-caliber vascular grafts in rats. Surgery. 1987;101:459–467. [PubMed] [Google Scholar]
- 12.Wildevuur CR, van der Lei B, Schakenraad JM. Basic aspects of the regeneration of small-calibre neoarteries in biodegradable vascular grafts in rats. Biomaterials. 1987;8:418–422. doi: 10.1016/0142-9612(87)90076-7. [DOI] [PubMed] [Google Scholar]
- 13.Harrington JK, et al. Determining the fate of seeded cells in venous tissue-engineered vascular grafts using serial MRI. FASEB. J. 2011;25:4150–4161. doi: 10.1096/fj.11-185140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hibino N, et al. Tissue-engineered vascular grafts form neovessels that arise from regeneration of the adjacent blood vessel. FASEB. J. 2011;25:2731–2739. doi: 10.1096/fj.11-182246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hibino N, et al. A critical role for macrophages in neovessel formation and the development of stenosis in tissue-engineered vascular grafts. FASEB. J. 2011;25:4253–4263. doi: 10.1096/fj.11-186585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Matsumura G, Hibino N, Ikada Y, Kurosawa H, Shin’oka T. Successful application of tissue engineered vascular autografts: clinical experience. Biomaterials. 2003;24:2303–2308. doi: 10.1016/S0142-9612(03)00043-7. [DOI] [PubMed] [Google Scholar]
- 17.Shinoka T, et al. Midterm clinical result of tissue-engineered vascular autografts seeded with autologous bone marrow cells. J. Thorac. Cardiovasc. Surg. 2005;129:1330–1338. doi: 10.1016/j.jtcvs.2004.12.047. [DOI] [PubMed] [Google Scholar]
- 18.Hibino N, et al. Late-term results of tissue-engineered vascular grafts in humans. J. Thorac. Cardiovasc. Surg. 2010;139:431–436. doi: 10.1016/j.jtcvs.2009.09.057. [DOI] [PubMed] [Google Scholar]
- 19.Roh JD, et al. Tissue-engineered vascular grafts transform into mature blood vessels via an inflammation-mediated process of vascular remodeling. Proc. Natl. Acad. Sci. USA. 2010;107:4669–4674. doi: 10.1073/pnas.0911465107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Syedain Z, et al. Tissue engineering of acellular vascular grafts capable of somatic growth in young lambs. Nat. Commun. 2016;7:12951. doi: 10.1038/ncomms12951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bockeria, L. A. et al. Total cavo-pulmonary connection with a new bio-absorbable vascular graft first clinical experience. J. Thorac. Cardiovasc. Surg. 153, 1542–1550 (2017). [DOI] [PubMed]
- 22.Wu W, Allen Ra, Wang Y. Fast-degrading elastomer enables rapid remodeling of a cell-free synthetic graft into a neoartery. Nat. Med. 2012;18:1148–1153. doi: 10.1038/nm.2821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zafar F, et al. Physiological growth, remodeling potential, and preserved function of a novel bioprosthetic tricuspid valve: Tubular bioprosthesis made of small intestinal submucosa-derived extracellular matrix. J. Am. Coll. Cardiol. 2015;66:877–888. doi: 10.1016/j.jacc.2015.06.1091. [DOI] [PubMed] [Google Scholar]
- 24.Ruiz CE, et al. Transcatheter placement of a low-profile biodegradable pulmonary valve made of small intestinal submucosa: A long-term study in a swine model. J. Thorac. Cardiovasc. Surg. 2005;130:477–484. doi: 10.1016/j.jtcvs.2005.04.008. [DOI] [PubMed] [Google Scholar]
- 25.Tudorache I, et al. Decellularized aortic allografts versus pulmonary autografts for aortic valve replacement in the growing sheep model: Haemodynamic and morphological results at 20 months after implantation. Eur. J. Cardio-thoracic Surg. 2016;49:1228–1238. doi: 10.1093/ejcts/ezv362. [DOI] [PubMed] [Google Scholar]
- 26.Neumann A, Cebotari S, Tudorache I, Haverich A, Sarikouch S. Heart valve engineering: Decellularized allograft matrices in clinical practice. Biomed. Tech. 2013;58:453–456. doi: 10.1515/bmt-2012-0115. [DOI] [PubMed] [Google Scholar]
- 27.Iop, L. et al. Decellularized allogeneic heart valves demonstrate self-regeneration potential after a long-term preclinical evaluation. PLoS. ONE. 9, e99593 (2014). [DOI] [PMC free article] [PubMed]
- 28.Syedain Z, et al. 6-Month aortic valve implantation of an off-the-shelf tissue-engineered valve in sheep. Biomaterials. 2015;73:175–184. doi: 10.1016/j.biomaterials.2015.09.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Driessen-Mol A, et al. Transcatheter implantation of homologous “off-the-shelf” tissue-engineered heart valves with self-repair capacity: Long-term functionality and rapid in vivo remodeling in sheep. J. Am. Coll. Cardiol. 2014;63:1320–1329. doi: 10.1016/j.jacc.2013.09.082. [DOI] [PubMed] [Google Scholar]
- 30.Dijkman PE, Driessen-Mol A, Frese L, Hoerstrup SP, Baaijens FPT. Decellularized homologous tissue-engineered heart valves as off-the-shelf alternatives to xeno- and homografts. Biomaterials. 2012;33:4545–4554. doi: 10.1016/j.biomaterials.2012.03.015. [DOI] [PubMed] [Google Scholar]
- 31.Reimer J, et al. Implantation of a tissue-engineered tubular heart valve in growing lambs. Ann. Biomed. Eng. 2016 doi: 10.1007/s10439-016-1605-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Weber B, et al. Injectable living marrow stromal cell-based autologous tissue engineered heart valves: first experiences with a one-step intervention in primates. Eur. Heart. J. 2011;32:2830–2840. doi: 10.1093/eurheartj/ehr059. [DOI] [PubMed] [Google Scholar]
- 33.Kluin J, et al. In situ heart valve tissue engineering using a bioresorbable elastomeric implant - From material design to 12 months follow-up in sheep. Biomaterials. 2017;125:101–117. doi: 10.1016/j.biomaterials.2017.02.007. [DOI] [PubMed] [Google Scholar]
- 34.Anderson JM, Rodriguez A, Chang DT. Foreign body reaction to biomaterials. Semin. Immunol. 2008;20:86–100. doi: 10.1016/j.smim.2007.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Franz S, Rammelt S, Scharnweber D, Simon JC. Immune responses to implants - a review of the implications for the design of immunomodulatory biomaterials. Biomaterials. 2011;32:6692–6709. doi: 10.1016/j.biomaterials.2011.05.078. [DOI] [PubMed] [Google Scholar]
- 36.Smits AIPM, et al. In situ tissue engineering of functional small-diameter blood vessels by host circulating cells only. Tissue. Eng. Part. A. 2015;21:2583–2594. doi: 10.1089/ten.tea.2015.0066. [DOI] [PubMed] [Google Scholar]
- 37.Hibino N, et al. The innate immune system contributes to tissue-engineered vascular graft performance. FASEB. J. 2015;29:2431–2438. doi: 10.1096/fj.14-268334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Mirza R, DiPietro La, Koh TJ. Selective and specific macrophage ablation is detrimental to wound healing in mice. Am. J. Pathol. 2009;175:2454–2462. doi: 10.2353/ajpath.2009.090248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Godwin JW, Pinto AR, Rosenthal NA. Macrophages are required for adult salamander limb regeneration. Proc. Natl. Acad. Sci. U. S. A. 2013;110:9415–9420. doi: 10.1073/pnas.1300290110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Danenberg HD. Macrophage depletion by clodronate-containing liposomes reduces neointimal formation after balloon injury in rats and rabbits. Circulation. 2002;106:599–605. doi: 10.1161/01.CIR.0000023532.98469.48. [DOI] [PubMed] [Google Scholar]
- 41.Mantovani, A., Biswas, S. K., Galdiero, M. R., Sica, A. & Locati, M. Macrophage plasticity and polarization in tissue repair and remodelling. J. Pathol. doi:10.1002/path.4133 (2012). [DOI] [PubMed]
- 42.Sica A, Mantovani A. Macrophage plasticity and polarization: in vivo veritas. J. Clin. Invest. 2012;122:787–795. doi: 10.1172/JCI59643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Mosser D, Edwards J. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008;8:958–969. doi: 10.1038/nri2448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Porcheray F, et al. Macrophage activation switching: an asset for the resolution of inflammation. Clin. Exp. Immunol. 2005;142:481–489. doi: 10.1111/j.1365-2249.2005.02934.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Spiller KL, et al. The role of macrophage phenotype in vascularization of tissue engineering scaffolds. Biomaterials. 2014;35:4477–4488. doi: 10.1016/j.biomaterials.2014.02.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Schnoor M, et al. Production of type VI collagen by human macrophages: a new dimension in macrophage functional heterogeneity. J. Immunol. 2008;180:5707–5719. doi: 10.4049/jimmunol.180.8.5707. [DOI] [PubMed] [Google Scholar]
- 47.Krettek A, Sukhova GK, Libby P. Elastogenesis in human arterial disease: A role for macrophages in disordered elastin synthesis. Arterioscler. Thromb. Vasc. Biol. 2003;23:582–587. doi: 10.1161/01.ATV.0000064372.78561.A5. [DOI] [PubMed] [Google Scholar]
- 48.Battiston, K. G., Labow, R. S., Simmons, C. a. & Santerre, J. P. Immunomodulatory polymeric scaffold enhances extracellular matrix production in cell co-cultures under dynamic mechanical stimulation. Acta Biomater. doi:10.1016/j.actbio.2015.05.038 (2015). [DOI] [PubMed]
- 49.Daniel J-M, Sedding DG. Circulating smooth muscle progenitor cells in arterial remodeling. J. Mol. Cell. Cardiol. 2011;50:273–279. doi: 10.1016/j.yjmcc.2010.10.030. [DOI] [PubMed] [Google Scholar]
- 50.Pilling D, Gomer RH. Differentiation of circulating monocytes into fibroblast-like cells. Methods. Mol. Biol. 2012;904:191–206. doi: 10.1007/978-1-61779-943-3_16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Swirski FK, Nahrendorf M. Do vascular smooth muscle cells differentiate to macrophages in atherosclerotic lesions? Circ. Res. 2015;115:605–606. doi: 10.1161/CIRCRESAHA.114.304925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Brown BN, et al. Macrophage phenotype as a predictor of constructive remodeling following the implantation of biologically derived surgical mesh materials. Acta Biomater. 2012;8:978–987. doi: 10.1016/j.actbio.2011.11.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Asahara T, et al. Isolation of putative progenitor endothelial cells for angiogenesis. Science. 1997;275:964–966. doi: 10.1126/science.275.5302.964. [DOI] [PubMed] [Google Scholar]
- 54.Simper D, Stalboerger PG, Panetta CJ, Wang S, Caplice NM. Smooth muscle progenitor cells in human blood. Circulation. 2002;106:1199–1204. doi: 10.1161/01.CIR.0000031525.61826.A8. [DOI] [PubMed] [Google Scholar]
- 55.Zilla P, Bezuidenhout D, Human P. Prosthetic vascular grafts: wrong models, wrong questions and no healing. Biomaterials. 2007;28:5009–5027. doi: 10.1016/j.biomaterials.2007.07.017. [DOI] [PubMed] [Google Scholar]
- 56.Deb A, et al. Bone marrow-derived myofibroblasts are present in adult human heart valves. J. Heart. Valve. Dis. 2005;14:674–678. [PubMed] [Google Scholar]
- 57.Hajdu Z, et al. Recruitment of bone marrow-derived valve interstitial cells is a normal homeostatic process. J. Mol. Cell. Cardiol. 2011;51:955–965. doi: 10.1016/j.yjmcc.2011.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Visconti RP, et al. An in vivo analysis of hematopoietic stem cell potential: hematopoietic origin of cardiac valve interstitial cells. Circ. Res. 2006;98:690–696. doi: 10.1161/01.RES.0000207384.81818.d4. [DOI] [PubMed] [Google Scholar]
- 59.Kouchi Y, et al. Apparent blood stream origin of endothelial and smooth muscle cells in the neointima of long, impervious carotid-femoral grafts in the dog. Ann. Vasc. Surg. 1998;12:46–54. doi: 10.1007/s100169900114. [DOI] [PubMed] [Google Scholar]
- 60.Shi Q, et al. Proof of fallout endothelialization of impervious Dacron grafts in the aorta and inferior vena cava of the dog. J. Vasc. Surg. 1994;20:546–556. doi: 10.1016/0741-5214(94)90279-8. [DOI] [PubMed] [Google Scholar]
- 61.Forbes SJ, Rosenthal N. Preparing the ground for tissue regeneration: from mechanism to therapy. Nat. Med. 2014;20:857–869. doi: 10.1038/nm.3653. [DOI] [PubMed] [Google Scholar]
- 62.Ballotta V, Smits AIPM, Driessen-Mol A, Bouten CVC, Baaijens FPT. Synergistic protein secretion by mesenchymal stromal cells seeded in 3D scaffolds and circulating leukocytes in physiological flow. Biomaterials. 2014;35:9100–9113. doi: 10.1016/j.biomaterials.2014.07.042. [DOI] [PubMed] [Google Scholar]
- 63.Aggarwal S, Pittenger MF. Human mesenchymal stem cells modulate allogeneic immune cell responses. Blood. 2005;105:1815–1822. doi: 10.1182/blood-2004-04-1559. [DOI] [PubMed] [Google Scholar]
- 64.Holt DJ, Chamberlain LM, Grainger DW. Cell–cell signaling in co-cultures of macrophages and fibroblasts. Biomaterials. 2010;31:9382–9394. doi: 10.1016/j.biomaterials.2010.07.101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Chen L, et al. Interaction of vascular smooth muscle cells and monocytes by soluble factors synergistically enhances IL-6 and MCP-1 production. Am. J. Physiol. Heart. Circ. Physiol. 2009;296:H987–H996. doi: 10.1152/ajpheart.01158.2008. [DOI] [PubMed] [Google Scholar]
- 66.Battiston KG, Ouyang B, Labow RS, Simmons C. a. & Santerre, J. P. Monocyte/macrophage cytokine activity regulates vascular smooth muscle cell function within a degradable polyurethane scaffold. Acta. Biomater. 2014;10:1146–1155. doi: 10.1016/j.actbio.2013.12.022. [DOI] [PubMed] [Google Scholar]
- 67.McBane JE, et al. The effect of degradable polymer surfaces on co-cultures of monocytes and smooth muscle cells. Biomaterials. 2011;32:3584–3595. doi: 10.1016/j.biomaterials.2011.01.069. [DOI] [PubMed] [Google Scholar]
- 68.Ploeger DT, et al. Cell plasticity in wound healing: paracrine factors of M1/ M2 polarized macrophages influence the phenotypical state of dermal fibroblasts. Cell. Commun. Signal. 2013;11:29. doi: 10.1186/1478-811X-11-29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Song E, et al. Influence of alternatively and classically activated macrophages on fibrogenic activities of human fibroblasts. Cell. Immunol. 2000;204:19–28. doi: 10.1006/cimm.2000.1687. [DOI] [PubMed] [Google Scholar]
- 70.McBane JE, Cai K, Labow RS, Santerre JP. Co-culturing monocytes with smooth muscle cells improves cell distribution within a degradable polyurethane scaffold and reduces inflammatory cytokines. Acta Biomater. 2012;8:488–501. doi: 10.1016/j.actbio.2011.09.018. [DOI] [PubMed] [Google Scholar]
- 71.Sridharan R, Cameron AR, Kelly DJ, Kearney CJ, O’Brien FJ. Biomaterial based modulation of macrophage polarization: a review and suggested design principles. Mater. Today. 2015;18:313–325. doi: 10.1016/j.mattod.2015.01.019. [DOI] [Google Scholar]
- 72.McWhorter FY, Davis CT, Liu WF. Physical and mechanical regulation of macrophage phenotype and function. Cell. Mol. Life. Sci. 2014;72:1303–1316. doi: 10.1007/s00018-014-1796-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Wagenseil JE, Mecham RP. Vascular extracellular matrix and arterial mechanics. Physiol. Rev. 2009;89:957–989. doi: 10.1152/physrev.00041.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Chester AH, et al. The living aortic valve: From molecules to function. Glob. Cardiol. Sci. Pract. 2014;2014:52–77. doi: 10.5339/gcsp.2014.52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Mirensky TL, et al. Tissue-engineered arterial grafts: long-term results after implantation in a small animal model. J. Pediatr. Surg. 2009;44:1127-32–3. doi: 10.1016/j.jpedsurg.2009.02.035. [DOI] [PubMed] [Google Scholar]
- 76.Mirensky TL, et al. Tissue-engineered vascular grafts: does cell seeding matter? J. Pediatr. Surg. 2010;45:1299–1305. doi: 10.1016/j.jpedsurg.2010.02.102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Mahler GJ, Frendl CM, Cao Q, Butcher JT. Effects of shear stress pattern and magnitude on mesenchymal transformation and invasion of aortic valve endothelial cells. Biotechnol. Bioeng. 2014;111:2326–2337. doi: 10.1002/bit.25291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Monaghan MG, et al. A spatiotemporal observation of EndMT and mesenchymal cell colonization at the onset of human cardiac valve development. Development. 2015;49:473–482. doi: 10.1242/dev.133843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Muylaert, D. E. P. et al. Environmental influences on endothelial to mesenchymal transition in developing implanted cardiovascular tissue-engineered grafts. Tissue. Eng. Part. B. Rev. 0, ten.teb.2015.0167 (2015). [DOI] [PubMed]
- 80.Angelos MG, et al. Dynamic adhesion of umbilical cord blood endothelial progenitor cells under laminar shear stress. Biophys. J. 2010;99:3545–3554. doi: 10.1016/j.bpj.2010.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Markway BD, et al. Capture of flowing endothelial cells using surface-immobilized anti-kinase insert domain receptor antibody. Tissue. Eng. Part. C. Methods. 2008;14:97–105. doi: 10.1089/ten.tec.2007.0300. [DOI] [PubMed] [Google Scholar]
- 82.Plouffe BD, Radisic M, Murthy SK. Microfluidic depletion of endothelial cells, smooth muscle cells, and fibroblasts from heterogeneous suspensions. Lab. Chip. 2008;8:462–472. doi: 10.1039/b715707j. [DOI] [PubMed] [Google Scholar]
- 83.Wang X, Cooper S. Adhesion of endothelial cells and endothelial progenitor cells on peptide-linked polymers in shear flow. Tissue. Eng. Part. A. 2013;19:1113–1121. doi: 10.1089/ten.tea.2011.0653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Ruggeri ZM. Platelet adhesion under flow. Microcirculation. 2009;16:58–83. doi: 10.1080/10739680802651477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Chen S, Springer TA. Selectin receptor-ligand bonds: Formation limited by shear rate and dissociation governed by the bell model. Proc. Natl. Acad. Sci. USA. 2001;98:950–955. doi: 10.1073/pnas.98.3.950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Shive MS, Brodbeck WG, Colton E, Anderson JM. Shear stress and material surface effects on adherent human monocyte apoptosis. J. Biomed. Mater. Res. 2002;60:148–158. doi: 10.1002/jbm.10035. [DOI] [PubMed] [Google Scholar]
- 87.Smits AIPM, Ballotta V, Driessen-Mol A, Bouten CVC, Baaijens FPT. Shear flow affects selective monocyte recruitment into MCP-1-loaded scaffolds. J. Cell. Mol. Med. 2014;18:2176–2188. doi: 10.1111/jcmm.12330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Fitts MK, Pike DB, Anderson K, Shiu Y-T. Hemodynamic shear stress and endothelial dysfunction in hemodialysis access. Int. Urol. Nephrol. 2014;7:33–44. doi: 10.2174/1874303X01407010033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Lyman DJ, Stewart SFC, Murray-Wijelath J, Wijelath E. Role of fluid dynamics on the healing of an in vivo tissue engineered vascular graft. J. Biomed. Mater. Res. B. Appl. Biomater. 2006;77:389–400. doi: 10.1002/jbm.b.30436. [DOI] [PubMed] [Google Scholar]
- 90.Claiborne TE, et al. In vitro evaluation of a novel hemodynamically optimized trileaflet polymeric prosthetic heart valve. J. Biomech. Eng. 2013;135:21021. doi: 10.1115/1.4023235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Gaudio CDel, Gasbarroni PL, Romano GP. Experimental investigations on the fluid-mechanics of an electrospun heart valve by means of particle image velocimetry. J. Mech. Behav. Biomed. Mater. 2016;64:229–239. doi: 10.1016/j.jmbbm.2016.07.030. [DOI] [PubMed] [Google Scholar]
- 92.Rubbens MP, et al. Intermittent straining accelerates the development of tissue properties in engineered heart valve tissue. Tissue. Eng. Part. A. 2009;15:999–1008. doi: 10.1089/ten.tea.2007.0396. [DOI] [PubMed] [Google Scholar]
- 93.van Geemen D, Driessen-Mol A, Baaijens FPT, Bouten CVC. Understanding strain-induced collagen matrix development in engineered cardiovascular tissues from gene expression profiles. Cell. Tissue. Res. 2013;352:727–737. doi: 10.1007/s00441-013-1573-2. [DOI] [PubMed] [Google Scholar]
- 94.Gupta V, Tseng H, Lawrence BD, Jane Grande-Allen K. Effect of cyclic mechanical strain on glycosaminoglycan and proteoglycan synthesis by heart valve cells. Acta Biomater. 2009;5:531–540. doi: 10.1016/j.actbio.2008.10.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Huang, A. H. et al. Biaxial stretch improves elastic fiber maturation, collagen arrangement and mechanical properties in engineered arteries. Tissue. Eng. Part. C. Methods. 1–51. doi:10.1089/ten.TEC.2015.0309 (2016). [DOI] [PMC free article] [PubMed]
- 96.Venkataraman L, Bashur CA, Ramamurthi A. Impact of cyclic stretch on induced elastogenesis within collagenous conduits. Tissue. Eng. Part. A. 2014;20:1403–1415. doi: 10.1089/ten.tea.2013.0294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Yamamoto K, Ando J. Differentiation of stem/progenitor cells into vascular cells in response to fluid mechanical forces. Biorheology. 2010;24:1–10. doi: 10.1007/s12573-010-0017-9. [DOI] [Google Scholar]
- 98.Kurpinski K, Chu J, Hashi C, Li S. Anisotropic mechanosensing by mesenchymal stem cells. Proc. Natl. Acad. Sci. U. S. A. 2006;103:16095–16100. doi: 10.1073/pnas.0604182103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Yang JH, Sakamoto H, Xu EC, Lee RT. Biomechanical regulation of human monocyte/macrophage molecular function. Am. J. Pathol. 2000;156:1797–1804. doi: 10.1016/S0002-9440(10)65051-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Ohki R, Yamamoto K, Mano H. Identification of mechanically induced genes in human monocytic cells by DNA microarrays. J. Hypertens. 2002;20:685–691. doi: 10.1097/00004872-200204000-00026. [DOI] [PubMed] [Google Scholar]
- 101.Matheson LA, Maksym GN, Santerre JP, Labow RS. Differential effects of uniaxial and biaxial strain on U937 macrophage-like cell morphology: Influence of extracellular matrix type proteins. J. Biomed. Mater. Res. Part A. 2007;81A:971–981. doi: 10.1002/jbm.a.31117. [DOI] [PubMed] [Google Scholar]
- 102.Matheson LA, Maksym GN, Santerre JP, Labow RS. Cyclic biaxial strain affects U937 macrophage-like morphology and enzymatic activities. J. Biomed. Mater. Res. Part A. 2006;76:52–62. doi: 10.1002/jbm.a.30448. [DOI] [PubMed] [Google Scholar]
- 103.Ballotta V, Driessen-Mol A, Bouten CVC, Baaijens FPT. Strain-dependent modulation of macrophage polarization within scaffolds. Biomaterials. 2014;35:4919–4928. doi: 10.1016/j.biomaterials.2014.03.002. [DOI] [PubMed] [Google Scholar]
- 104.Wise SG, et al. A multilayered synthetic human elastin/polycaprolactone hybrid vascular graft with tailored mechanical properties. Acta Biomater. 2011;7:295–303. doi: 10.1016/j.actbio.2010.07.022. [DOI] [PubMed] [Google Scholar]
- 105.Lee K-W, Stolz DB, Wang Y. Substantial expression of mature elastin in arterial constructs. Proc. Natl. Acad. Sci. U. S. A. 2011;108:2705–2710. doi: 10.1073/pnas.1017834108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Argento G, Simonet M, Oomens CWJ, Baaijens FPT. Multi-scale mechanical characterization of scaffolds for heart valve tissue engineering. J. Biomech. 2012;45:2893–2898. doi: 10.1016/j.jbiomech.2012.07.037. [DOI] [PubMed] [Google Scholar]
- 107.Loerakker S, Argento G, Oomens CWJ, Baaijens FPT. Effects of valve geometry and tissue anisotropy on the radial stretch and coaptation area of tissue-engineered heart valves. J. Biomech. 2013;46:1792–1800. doi: 10.1016/j.jbiomech.2013.05.015. [DOI] [PubMed] [Google Scholar]
- 108.Argento G, et al. Modeling the impact of scaffold architecture and mechanical loading on collagen turnover in engineered cardiovascular tissues. Biomech. Model. Mechanobiol. 2014 doi: 10.1007/s10237-014-0625-1. [DOI] [PubMed] [Google Scholar]
- 109.de Jonge N, et al. Degree of scaffold degradation influences collagen (re)orientation in engineered tissues. Tissue. Eng. Part. A. 2013;20:1747–1757. doi: 10.1089/ten.tea.2013.0517. [DOI] [PubMed] [Google Scholar]
- 110.Sohier J, et al. The potential of anisotropic matrices as substrate for heart valve engineering. Biomaterials. 2014;35:1833–1844. doi: 10.1016/j.biomaterials.2013.10.061. [DOI] [PubMed] [Google Scholar]
- 111.Fioretta ES, Simonet M, Smits AIPM, Baaijens FPTT, Bouten CVCC. Differential response of endothelial and endothelial colony forming cells on electrospun scaffolds with distinct microfiber diameters. Biomacromolecules. 2014;15:821–829. doi: 10.1021/bm4016418. [DOI] [PubMed] [Google Scholar]
- 112.Balguid A, et al. Tailoring fiber diameter in electrospun poly(epsilon-caprolactone) scaffolds for optimal cellular infiltration in cardiovascular tissue engineering. Tissue. Eng. Part. A. 2009;15:437–444. doi: 10.1089/ten.tea.2007.0294. [DOI] [PubMed] [Google Scholar]
- 113.Smits AIPM, Driessen-Mol A, Bouten CVC, Baaijens FPT. A mesofluidics-based test platform for systematic development of scaffolds for in situ cardiovascular tissue engineering. Tissue. Eng. Part. C. Methods. 2012;18:475–485. doi: 10.1089/ten.tec.2011.0458. [DOI] [PubMed] [Google Scholar]
- 114.Kurpinski KT, Stephenson JT, Janairo RRR, Lee H, Li S. The effect of fiber alignment and heparin coating on cell infiltration into nanofibrous PLLA scaffolds. Biomaterials. 2010;31:3536–3542. doi: 10.1016/j.biomaterials.2010.01.062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Vaz CM, van Tuijl S, Bouten CVC, Baaijens FPT. Design of scaffolds for blood vessel tissue engineering using a multi-layering electrospinning technique. Acta Biomater. 2005;1:575–582. doi: 10.1016/j.actbio.2005.06.006. [DOI] [PubMed] [Google Scholar]
- 116.de Valence S, et al. Advantages of bilayered vascular grafts for surgical applicability and tissue regeneration. Acta Biomater. 2012;8:3914–3920. doi: 10.1016/j.actbio.2012.06.035. [DOI] [PubMed] [Google Scholar]
- 117.Pham QP, Sharma U, Mikos AG. Electrospun poly(epsilon-caprolactone) microfiber and multilayer nanofiber/microfiber scaffolds: characterization of scaffolds and measurement of cellular infiltration. Biomacromolecules. 2006;7:2796–2805. doi: 10.1021/bm060680j. [DOI] [PubMed] [Google Scholar]
- 118.Milleret V, Hefti T, Hall H, Vogel V, Eberli D. Influence of the fiber diameter and surface roughness of electrospun vascular grafts on blood activation. Acta Biomater. 2012;8:4349–4356. doi: 10.1016/j.actbio.2012.07.032. [DOI] [PubMed] [Google Scholar]
- 119.Saino E, et al. Effect of electrospun fiber diameter and alignment on macrophage activation and secretion of proinflammatory cytokines and chemokines. Biomacromolecules. 2011;12:1900–1911. doi: 10.1021/bm200248h. [DOI] [PubMed] [Google Scholar]
- 120.Madden LR, et al. Proangiogenic scaffolds as functional templates for cardiac tissue engineering. Proc. Natl. Acad. Sci. U. S. A. 2010;107:15211–15216. doi: 10.1073/pnas.1006442107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Sussman EM, Halpin MC, Muster J, Moon RT, Ratner BD. Porous implants modulate healing and induce shifts in local macrophage polarization in the foreign body reaction. Ann. Biomed. Eng. 2014;42:1508–1516. doi: 10.1007/s10439-013-0933-0. [DOI] [PubMed] [Google Scholar]
- 122.Garg K, Pullen Na, Oskeritzian Ca, Ryan JJ, Bowlin GL. Macrophage functional polarization (M1/M2) in response to varying fiber and pore dimensions of electrospun scaffolds. Biomaterials. 2013;34:4439–4451. doi: 10.1016/j.biomaterials.2013.02.065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.McWhorter FY, Wang T, Nguyen P, Chung T, Liu WF. Modulation of macrophage phenotype by cell shape. Proc. Natl. Acad. Sci. U. S. A. 2013;110:17253–17258. doi: 10.1073/pnas.1308887110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Generali M, Dijkman PE, Hoerstrup SP. Bioresorbable Scaffolds for Cardiovascular Tissue Engineering. EMJ Interv. Cardiol. 2014;1:91–99. [Google Scholar]
- 125.Hong Y, et al. Tailoring the degradation kinetics of poly(ester carbonate urethane)urea thermoplastic elastomers for tissue engineering scaffolds. Biomaterials. 2010;31:4249–4258. doi: 10.1016/j.biomaterials.2010.02.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Brugmans MCP, et al. Hydrolytic and oxidative degradation of electrospun supramolecular biomaterials: In vitro degradation pathways. Acta Biomater. 2015;27:21–31. doi: 10.1016/j.actbio.2015.08.034. [DOI] [PubMed] [Google Scholar]
- 127.Zhang H, Zhou L, Zhang W. Control of scaffold degradation in tissue engineering: a review. Tissue. Eng. Part. B. Rev. 2014;20:492–502. doi: 10.1089/ten.teb.2013.0452. [DOI] [PubMed] [Google Scholar]
- 128.Peng H, et al. Controlled enzymatic degradation of poly(??-caprolactone)-based copolymers in the presence of porcine pancreatic lipase. Polym. Degrad. Stab. 2010;95:643–650. doi: 10.1016/j.polymdegradstab.2009.12.005. [DOI] [Google Scholar]
- 129.Matsumura G, et al. Long-term results of cell-free biodegradable scaffolds for in situ tissue engineering of pulmonary artery in a canine model. Biomaterials. 2013;34:6422–6428. doi: 10.1016/j.biomaterials.2013.05.037. [DOI] [PubMed] [Google Scholar]
- 130.Mrówczyński W, et al. Porcine carotid artery replacement with biodegradable electrospun poly-e-caprolactone vascular prosthesis. J. Vasc. Surg. 2014;59:210–219. doi: 10.1016/j.jvs.2013.03.004. [DOI] [PubMed] [Google Scholar]
- 131.de Valence S, et al. Long term performance of polycaprolactone vascular grafts in a rat abdominal aorta replacement model. Biomaterials. 2012;33:38–47. doi: 10.1016/j.biomaterials.2011.09.024. [DOI] [PubMed] [Google Scholar]
- 132.Weber B, Zeisberger SM, Hoerstrup SP. Prenatally harvested cells for cardiovascular tissue engineering: fabrication of autologous implants prior to birth. Placenta. 2011;32:S316–S319. doi: 10.1016/j.placenta.2011.04.001. [DOI] [PubMed] [Google Scholar]
- 133.De Visscher G, et al. The remodeling of cardiovascular bioprostheses under influence of stem cell homing signal pathways. Biomaterials. 2010;31:20–28. doi: 10.1016/j.biomaterials.2009.09.016. [DOI] [PubMed] [Google Scholar]
- 134.Thevenot PT, et al. The effect of incorporation of SDF-1alpha into PLGA scaffolds on stem cell recruitment and the inflammatory response. Biomaterials. 2010;31:3997–4008. doi: 10.1016/j.biomaterials.2010.01.144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Mokarram N, Merchant A, Mukhatyar V, Patel G, Bellamkonda RV. Effect of modulating macrophage phenotype on peripheral nerve repair. Biomaterials. 2012;33:8793–8801. doi: 10.1016/j.biomaterials.2012.08.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Potas JR, Haque F, Maclean FL, Nisbet DR. Interleukin-10 conjugated electrospun polycaprolactone (PCL) nanofibre scaffolds for promoting alternatively activated (M2) macrophages around the peripheral nerve in vivo. J. Immunol. Methods. 2015;420:38–49. doi: 10.1016/j.jim.2015.03.013. [DOI] [PubMed] [Google Scholar]
- 137.Reeves ARD, Spiller KL, Freytes DO, Vunjak-Novakovic G, Kaplan DL. Controlled release of cytokines using silk-biomaterials for macrophage polarization. Biomaterials. 2015;73:272–283. doi: 10.1016/j.biomaterials.2015.09.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Spiller KL, et al. Sequential delivery of immunomodulatory cytokines to facilitate the M1-to-M2 transition of macrophages and enhance vascularization of bone scaffolds. Biomaterials. 2015;37:194–207. doi: 10.1016/j.biomaterials.2014.10.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Kumar VA, et al. Self-assembling multidomain peptides tailor biological responses through biphasic release. Biomaterials. 2015;52:71–78. doi: 10.1016/j.biomaterials.2015.01.079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Chen F-M, Zhang M, Wu Z-F. Toward delivery of multiple growth factors in tissue engineering. Biomaterials. 2010;31:6279–6308. doi: 10.1016/j.biomaterials.2010.04.053. [DOI] [PubMed] [Google Scholar]
- 141.Jay SM, et al. Dual delivery of VEGF and MCP-1 to support endothelial cell transplantation for therapeutic vascularization. Biomaterials. 2010;31:3054–3062. doi: 10.1016/j.biomaterials.2010.01.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Szentivanyi A, Chakradeo T, Zernetsch H, Glasmacher B. Electrospun cellular microenvironments: Understanding controlled release and scaffold structure. Adv. Drug. Deliv. Rev. 2011;63:209–220. doi: 10.1016/j.addr.2010.12.002. [DOI] [PubMed] [Google Scholar]
- 143.Salek-Ardakani S, Arrand JR, Shaw D, Mackett M. Heparin and heparan sulfate bind interleukin-10 and modulate its activity. Blood. 2000;96:1879–1888. [PubMed] [Google Scholar]
- 144.Rajangam K, et al. Heparin binding nanostructures to promote growth of blood vessels. Nano. Lett. 2006;6:2086–2090. doi: 10.1021/nl0613555. [DOI] [PubMed] [Google Scholar]
- 145.Lee J, Yoo JJ, Atala A, Lee SJ. The effect of controlled release of PDGF-BB from heparin-conjugated electrospun PCL/gelatin scaffolds on cellular bioactivity and infiltration. Biomaterials. 2012;33:6709–6720. doi: 10.1016/j.biomaterials.2012.06.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Nillesen STM, et al. Increased angiogenesis and blood vessel maturation in acellular collagen-heparin scaffolds containing both FGF2 and VEGF. Biomaterials. 2007;28:1123–1131. doi: 10.1016/j.biomaterials.2006.10.029. [DOI] [PubMed] [Google Scholar]
- 147.Mammadov R, Mammadov B, Guler MO, Tekinay AB. Growth factor binding on heparin mimetic peptide nanofibers. Biomacromolecules. 2012;13:3311–3319. doi: 10.1021/bm3010897. [DOI] [PubMed] [Google Scholar]
- 148.Muylaert DEP, et al. Early in-situ cellularization of a supramolecular vascular graft is modified by synthetic stromal cell-derived factor-1α derived peptides. Biomaterials. 2015;76:187–195. doi: 10.1016/j.biomaterials.2015.10.052. [DOI] [PubMed] [Google Scholar]
- 149.Lee TC, Midura RJ, Hascall VC, Vesely I. The effect of elastin damage on the mechanics of the aortic valve. J. Biomech. 2001;34:203–210. doi: 10.1016/S0021-9290(00)00187-1. [DOI] [PubMed] [Google Scholar]
- 150.Schoof PH, et al. Degeneration of the pulmonary autograft: an explant study. J. Thorac. Cardiovasc. Surg. 2006;132:1426–1432. doi: 10.1016/j.jtcvs.2006.07.035. [DOI] [PubMed] [Google Scholar]
- 151.Anidjar S, et al. Elastase-induced experimental aneurysms in rats. Circulation. 1990;82:973–981. doi: 10.1161/01.CIR.82.3.973. [DOI] [PubMed] [Google Scholar]
- 152.Saitow CB, Wise SG, Weiss AS, Castellot JJ, Kaplan DL. Elastin biology and tissue engineering with adult cells. Biomol. Concepts. 2013;4:173–185. doi: 10.1515/bmc-2012-0040. [DOI] [PubMed] [Google Scholar]
- 153.Huang AH, et al. Design and use of a novel bioreactor for regeneration of biaxially stretched tissue-engineered vessels. Tissue. Eng. Part. C. Methods. 2015;21:841–851. doi: 10.1089/ten.tec.2014.0287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Nejad SP, Blaser MC, Paul Santerre J, Caldarone CA, Simmons CA. Biomechanical conditioning of tissue engineered heart valves: Too much of a good thing? Adv. Drug. Deliv. Rev. 2015 doi: 10.1016/j.addr.2015.11.003. [DOI] [PubMed] [Google Scholar]
- 155.Keeley EC, Mehrad B, Strieter RM. Fibrocytes: bringing new insights into mechanisms of inflammation and fibrosis. Int. J. Biochem. Cell. Biol. 2010;42:535–542. doi: 10.1016/j.biocel.2009.10.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Ferguson MWJ, O’Kane S. Scar-free healing: from embryonic mechanisms to adult therapeutic intervention. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2004;359:839–850. doi: 10.1098/rstb.2004.1475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Harty M, Neff AW, King MW, Mescher AL. Regeneration or scarring: an immunologic perspective. Dev. Dyn. 2003;226:268–279. doi: 10.1002/dvdy.10239. [DOI] [PubMed] [Google Scholar]
- 158.Martin P, et al. Wound Healing in the PU.1 Null Mouse—Tissue Repair Is Not Dependent on Inflammatory Cells. Curr. Biol. 2003;13:1122–1128. doi: 10.1016/S0960-9822(03)00396-8. [DOI] [PubMed] [Google Scholar]
- 159.Naito Y, et al. Characterization of the natural history of extracellular matrix production in tissue-engineered vascular grafts during neovessel formation. Cells. Tissues. Organs. 2012;195:60–72. doi: 10.1159/000331405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Naito Y, et al. Beyond burst pressure: initial evaluation of the natural history of the biaxial mechanical properties of tissue-engineered vascular grafts in the venous circulation using a murine model. Tissue. Eng. Part. A. 2014;20:346–355. doi: 10.1089/ten.tea.2012.0613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Udelsman BV, et al. Characterization of evolving biomechanical properties of tissue engineered vascular grafts in the arterial circulation. J. Biomech. 2014;47:2070–2079. doi: 10.1016/j.jbiomech.2014.03.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Sugiura, T. et al. Fast-degrading bioresorbable arterial vascular graft with high cellular infiltration inhibits calcification of the graft. J. Vasc. Surg. 1–8. doi:10.1016/j.jvs.2016.05.096 (2016). [DOI] [PMC free article] [PubMed]
- 163.Wu MH, et al. Implant site influence on arterial prosthesis healing: a comparative study with a triple implantation model in the same dog. J. Vasc. Surg. 1997;25:528–536. doi: 10.1016/S0741-5214(97)70264-7. [DOI] [PubMed] [Google Scholar]
- 164.McGonigle P, Ruggeri B. Animal models of human disease: challenges in enabling translation. Biochem. Pharmacol. 2014;87:162–171. doi: 10.1016/j.bcp.2013.08.006. [DOI] [PubMed] [Google Scholar]
- 165.Pennel T, Zilla P, Bezuidenhout D. Differentiating transmural from transanastomotic prosthetic graft endothelialization through an isolation loop-graft model. J. Vasc. Surg. 2013;58:1053–1061. doi: 10.1016/j.jvs.2012.11.093. [DOI] [PubMed] [Google Scholar]
- 166.Byrom MJ, Bannon PG, White GH, Ng MKC. Animal models for the assessment of novel vascular conduits. J. Vasc. Surg. 2010;52:176–195. doi: 10.1016/j.jvs.2009.10.080. [DOI] [PubMed] [Google Scholar]
- 167.Seok J, et al. Genomic responses in mouse models poorly mimic human inflammatory diseases. Proc. Natl. Acad. Sci. U. S. A. 2013;110:3507–3512. doi: 10.1073/pnas.1222878110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Ingersoll MA, et al. Comparison of gene expression profiles between human and mouse monocyte subsets. Blood. 2010;115:e10–e19. doi: 10.1182/blood-2009-07-235028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Miller KS, Khosravi R, Breuer CK, Humphrey JD. A hypothesis-driven parametric study of effects of polymeric scaffold properties on tissue engineered neovessel formation. Acta. Biomater. 2015;11:283–294. doi: 10.1016/j.actbio.2014.09.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Wolf MT, Vodovotz Y, Tottey S, Brown BN, Badylak SF. Predicting in vivo responses to biomaterials via combined in vitro and in silico analysis. Tissue. Eng. Part. C. Methods. 2015;21:148–159. doi: 10.1089/ten.tec.2014.0167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Jannasch, M. et al. In vitro chemotaxis and tissue remodeling assays quantitatively characterize foreign body reaction. ALTEX (2016). [DOI] [PubMed]
- 172.Damanik FFR, Rothuizen TC, van Blitterswijk C, Rotmans JI, Moroni L. Towards an in vitro model mimicking the foreign body response: tailoring the surface properties of biomaterials to modulate extracellular matrix. Sci. Rep. 2014;4:6325. doi: 10.1038/srep06325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Boersema GSA, Grotenhuis N, Bayon Y, Lange JF, Bastiaansen-Jenniskens YM. The effect of biomaterials used for tissue regeneration purposes on polarization of macrophages. Biores. Open Access. 2016;5:6–14. doi: 10.1089/biores.2015.0041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Enayati M, et al. Biocompatibility assessment of a new biodegradable vascular graft via in vitro co-culture approaches and in vivo model. Ann. Biomed. Eng. 2016 doi: 10.1007/s10439-016-1601-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Wolf, F., Vogt, F., Schmitz-Rode, T., Jockenhoevel, S. & Mela, P. Bioengineered vascular constructs as living models for in vitro cardiovascular research. Drug. Discov. Today. 0, (2016). [DOI] [PubMed]
- 176.McBrearty Ba, Clark LD, Zhang XM, Blankenhorn EP, Heber-Katz E. Genetic analysis of a mammalian wound-healing trait. Proc. Natl. Acad. Sci. U. S. A. 1998;95:11792–11797. doi: 10.1073/pnas.95.20.11792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Khosravi R, et al. Biomechanical diversity despite mechanobiological stability in tissue engineered vascular grafts two years post-implantation. Tissue. Eng. Part. A. 2015;21:1529–1538. doi: 10.1089/ten.tea.2014.0524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Bouman A, Schipper M, Heineman MJ, Faas MM. Gender difference in the non-specific and specific immune response in humans. Am. J. Reprod. Immunol. 2004;52:19–26. doi: 10.1111/j.1600-0897.2004.00177.x. [DOI] [PubMed] [Google Scholar]
- 179.Goetzl EJ, et al. Gender specificity of altered human immune cytokine profiles in aging. FASEB. J. 2010;24:3580–3589. doi: 10.1096/fj.10-160911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Hodkinson CF, et al. Whole blood analysis of phagocytosis, apoptosis, cytokine production, and leukocyte subsets in healthy older men and women: the ZENITH study. J. Gerontol. A. Biol. Sci. Med. Sci. 2006;61:907–917. doi: 10.1093/gerona/61.9.907. [DOI] [PubMed] [Google Scholar]
- 181.Imahara SD, Jelacic S, Junker CE, O’Keefe GE. The influence of gender on human innate immunity. Surgery. 2005;138:275–282. doi: 10.1016/j.surg.2005.03.020. [DOI] [PubMed] [Google Scholar]
- 182.Krawiec JT, et al. In vivo functional evaluation of tissue-engineered vascular grafts fabricated using human adipose-derived stem cells from high cardiovascular risk populations. Tissue. Eng. Part. A. 2016;22:765–775. doi: 10.1089/ten.tea.2015.0379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Krawiec JT, et al. A cautionary tale for autologous vascular tissue engineering: impact of human demographics on the ability of adipose-derived mesenchymal stem cells to recruit and differentiate into smooth muscle cells. Tissue. Eng. Part. A. 2015;21:426–437. doi: 10.1089/ten.tea.2014.0208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Wang Z, et al. Differences in the performance of PCL-based vascular grafts as abdominal aorta substitutes in healthy and diabetic rats. Biomater. Sci. 2016 doi: 10.1039/c6bm00178e. [DOI] [PubMed] [Google Scholar]
- 185.Markiewicz K, van Til JA, IJzerman MJ. Medical devices early assessment methods: Systematic literature review. Int. J. Technol. Assess. Health. Care. 2014;30:137–146. doi: 10.1017/S0266462314000026. [DOI] [PubMed] [Google Scholar]
- 186.IJzerman MJ, Steuten MG. Early assessment of medical technologies to inform product development and market access. A review of methods and applications. Appl. Health. Econ. Health. Policy. 2011;9:331–347. doi: 10.2165/11593380-000000000-00000. [DOI] [PubMed] [Google Scholar]
- 187.Huygens SA, et al. Conceptual model for early health technology assessment of current and novel heart valve interventions. Open Heart. 2016;3:e000500. doi: 10.1136/openhrt-2016-000500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Hopman RK, DiPersio JF. Advances in stem cell mobilization. Blood. Rev. 2014;28:31–40. doi: 10.1016/j.blre.2014.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Tara S, et al. Evaluation of remodeling process in small-diameter cell-free tissue-engineered arterial graft. J. Vasc. Surg. 2015;62:734–743. doi: 10.1016/j.jvs.2014.03.011. [DOI] [PubMed] [Google Scholar]
- 190.Fukunishi T, et al. Tissue-engineered small diameter arterial vascular grafts from cell-free nanofiber PCL/Chitosan scaffolds in a sheep model. PLoS. ONE. 2016;11:e0158555. doi: 10.1371/journal.pone.0158555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Tara S, et al. Well-organized neointima of large-pore poly(L-lactic acid) vascular graft coated with poly(L-lactic-co-ε-caprolactone) prevents calcific deposition compared to small-pore electrospun poly(L-lactic acid) graft in a mouse aortic implantation model. Atherosclerosis. 2014;237:684–691. doi: 10.1016/j.atherosclerosis.2014.09.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Sugiura, T. et al. Novel bioresorbable vascular graft with sponge-type scaffold as a small-diameter arterial graft. Ann. Thorac. Surg. 1–8. doi:10.1016/j.athoracsur.2016.01.110 (2016). [DOI] [PMC free article] [PubMed]
- 193.Wang, K. et al. Three-layered pcl grafts promoted vascular regeneration in a rabbit carotid artery model. Macromol. Biosci. 608–618. doi:10.1002/mabi.201500355 (2016). [DOI] [PubMed]
- 194.Wang Z, et al. The effect of thick fibers and large pores of electrospun poly(ε-caprolactone) vascular grafts on macrophage polarization and arterial regeneration. Biomaterials. 2014;35:5700–5710. doi: 10.1016/j.biomaterials.2014.03.078. [DOI] [PubMed] [Google Scholar]
- 195.Zhu M, et al. Circumferentially aligned fibers guided functional neoartery regeneration in vivo. Biomaterials. 2015;61:85–94. doi: 10.1016/j.biomaterials.2015.05.024. [DOI] [PubMed] [Google Scholar]
- 196.Bergmeister H, et al. Biodegradable, thermoplastic polyurethane grafts for small diameter vascular replacements. Acta Biomater. 2015;11:104–113. doi: 10.1016/j.actbio.2014.09.003. [DOI] [PubMed] [Google Scholar]
- 197.Yang X, Wei J, Lei D, Liu Y, Wu W. Appropriate density of PCL nano-fiber sheath promoted muscular remodeling of PGS/PCL grafts in arterial circulation. Biomaterials. 2016;88:34–47. doi: 10.1016/j.biomaterials.2016.02.026. [DOI] [PubMed] [Google Scholar]
- 198.Khosravi R, et al. Long-term functional efficacy of a novel electrospun poly(glycerol sebacate)-based arterial graft in mice. Ann. Biomed. Eng. 2016;44:1–15. doi: 10.1007/s10439-015-1545-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Pektok E, et al. Degradation and healing characteristics of small-diameter poly(epsilon-caprolactone) vascular grafts in the rat systemic arterial circulation. Circulation. 2008;118:2563–2570. doi: 10.1161/CIRCULATIONAHA.108.795732. [DOI] [PubMed] [Google Scholar]
- 200.Zavan B, et al. Neoarteries grown in vivo using a tissue-engineered hyaluronan-based scaffold. FASEB. J. 2008;22:2853–2861. doi: 10.1096/fj.08-107284. [DOI] [PubMed] [Google Scholar]
- 201.Pandis L, et al. Hyaluronan-based scaffold for in vivo regeneration of the rat vena cava: Preliminary results in an animal model. J. Biomed. Mater. Res. A. 2010;93:1289–1296. doi: 10.1002/jbm.a.32626. [DOI] [PubMed] [Google Scholar]
- 202.Lepidi S, et al. In vivo regeneration of small-diameter (2 mm) arteries using a polymer scaffold. FASEB. J. 2006;20:103–105. doi: 10.1096/fj.05-4802fje. [DOI] [PubMed] [Google Scholar]
- 203.Rothuizen TC, et al. Development and evaluation of in vivo tissue engineered blood vessels in a porcine model. Biomaterials. 2016;75:82–90. doi: 10.1016/j.biomaterials.2015.10.023. [DOI] [PubMed] [Google Scholar]
- 204.Rothuizen TC, et al. Tailoring the foreign body response for in situ vascular tissue engineering. Tissue. Eng. Part. C. Methods. 2015;21:436–446. doi: 10.1089/ten.tec.2014.0264. [DOI] [PubMed] [Google Scholar]
- 205.Koobatian MT, et al. Successful endothelialization and remodeling of a cell-free small-diameter arterial graft in a large animal model. Biomaterials. 2016;76:344–358. doi: 10.1016/j.biomaterials.2015.10.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Row S, et al. Arterial grafts exhibiting unprecedented cellular infiltration and remodeling in vivo: The role of cells in the vascular wall. Biomaterials. 2015;50:115–126. doi: 10.1016/j.biomaterials.2015.01.045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Dahl SLM, et al. Readily available tissue-engineered vascular grafts. Sci. Transl. Med. 2011;3:68ra9. doi: 10.1126/scitranslmed.3001426. [DOI] [PubMed] [Google Scholar]
- 208.Van Almen GC, et al. Development of non-cell adhesive vascular grafts using supramolecular building blocks. Macromol. Biosci. 2016;16:350–362. doi: 10.1002/mabi.201500278. [DOI] [PubMed] [Google Scholar]
- 209.Shafiq M, Jung Y, Kim SH. Covalent Immobilization of Stem Cell Inducing/Recruiting Factor and Heparin on Cell-Free Small-Diameter Vascular Graft for Accelerated In Situ Tissue Regeneration. J. Biomed. Mater. Res. A. 2016 doi: 10.1002/jbm.a.35666. [DOI] [PubMed] [Google Scholar]
- 210.Shafiq M, Jung Y, Kim SH. In situ vascular regeneration using substance P-immobilised poly(L-Lactide-co-??-caprolactone) scaffolds: Stem cell recruitment, angiogenesis, and tissue regeneration. Eur. Cells Mater. 2015;30:282–302. doi: 10.22203/eCM.v030a20. [DOI] [PubMed] [Google Scholar]
- 211.Ota T, et al. Fibronectin-hepatocyte growth factor enhances reendothelialization in tissue-engineered heart valve. Ann. Thorac. Surg. 2005;80:1794–1801. doi: 10.1016/j.athoracsur.2005.05.002. [DOI] [PubMed] [Google Scholar]
- 212.Zhang H, et al. Dual-delivery of VEGF and PDGF by double-layered electrospun membranes for blood vessel regeneration. Biomaterials. 2013;34:2202–2212. doi: 10.1016/j.biomaterials.2012.12.005. [DOI] [PubMed] [Google Scholar]
- 213.Kao WJ, Lee D. In vivo modulation of host response and macrophage behavior by polymer networks grafted with fibronectin-derived biomimetic oligopeptides: the role of RGD and PHSRN domains. Biomaterials. 2001;22:2901–2909. doi: 10.1016/S0142-9612(01)00037-0. [DOI] [PubMed] [Google Scholar]
- 214.Sun Q, et al. Sustained release of multiple growth factors from injectable polymeric system as a novel therapeutic approach towards angiogenesis. Pharm. Res. 2010;27:264–271. doi: 10.1007/s11095-009-0014-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Silva EA, Mooney DJ. Spatiotemporal control of vascular endothelial growth factor delivery from injectable hydrogels enhances angiogenesis. J. Thromb. Haemost. 2007;5:590–598. doi: 10.1111/j.1538-7836.2007.02386.x. [DOI] [PubMed] [Google Scholar]
- 216.Riley CM, et al. Stimulation of in vivo angiogenesis using dual growth factor-loaded crosslinked glycosaminoglycan hydrogels. Biomaterials. 2006;27:5935–5943. doi: 10.1016/j.biomaterials.2006.08.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Peattie RA, et al. Dual growth factor-induced angiogenesis in vivo using hyaluronan hydrogel implants. Biomaterials. 2006;27:1868–1875. doi: 10.1016/j.biomaterials.2005.09.035. [DOI] [PubMed] [Google Scholar]
- 218.Dogan AK, Gümüşderelioglu M, Aksöz E. Controlled release of EGF and bFGF from dextran hydrogels in vitro and in vivo. J. Biomed. Mater. Res. B. Appl. Biomater. 2005;74:504–510. doi: 10.1002/jbm.b.30231. [DOI] [PubMed] [Google Scholar]
- 219.Lynn AD, Kyriakides TR, Bryant SJ. Characterization of the in vitro macrophage response and in vivo host response to poly(ethylene glycol)-based hydrogels. J. Biomed. Mater. Res. A. 2010;93:941–953. doi: 10.1002/jbm.a.32595. [DOI] [PubMed] [Google Scholar]
- 220.Lynn AD, Blakney AK, Kyriakides TR, Bryant SJ. Temporal progression of the host response to implanted poly(ethylene glycol)-based hydrogels. J. Biomed. Mater. Res. A. 2011;96:621–631. doi: 10.1002/jbm.a.33015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221.Zaveri TD, Lewis JS, Dolgova NV, Clare-Salzler MJ, Keselowsky BG. Integrin-directed modulation of macrophage responses to biomaterials. Biomaterials. 2014;35:3504–3515. doi: 10.1016/j.biomaterials.2014.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222.Gower, R. M. et al. Modulation of leukocyte infiltration and phenotype in microporous tissue engineering scaffolds via vector induced IL-10 expression. Biomaterials35, 2024–2031 (2014). [DOI] [PMC free article] [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Data sharing not applicable to this article as no data sets were generated or analyzed during the current study.