

Abstract
The present study examined the interaction between early life stress and 5-HTT genotypes in predicting two risk factors for suicidal behavior – the brooding subtype of rumination and impulsivity, in the form of negative urgency – over time. Furthermore, we examined early life stress, brooding, and impulsivity as predictors of suicidal ideation over time. Participants with and without a history of early life stress were genotyped for the 5-HTTLPR polymorphism and completed assessments assessing brooding and negative urgency at baseline and 6-month follow up. Early life emotional abuse was associated with negative urgency at follow-up. We found an indirect effect of early life emotional abuse on negative urgency through brooding among individuals with 5-HTT low expressing genotypes but not among individuals with 5-HTT high expressing genotypes. Further, a logistic regression analysis revealed that negative urgency was associated with higher odds (O.R. = 16.2) of reporting suicide ideation (versus no ideation) at follow-up. Our findings suggest that brooding and negative urgency may result from the interaction between early life emotional abuse and 5-HTT low expressing genotypes. Further research is necessary to understand how early life stress interacts with 5-HTT genotypes to confer risk for suicidal behavior through psychological mechanisms.
Keywords: Early life emotional abuse, Serotonin transporter, Rumination, Impulsivity, Suicidal ideation
1. Introduction
Early life stress puts people at risk for various forms of psychopathology, including but not limited to post-traumatic stress disorder, panic disorder, obsessive-compulsive disorder, depression, and suicidal behavior (Santa Mina and Gallop, 1998; Yen et al., 2009; Wang et al., 2012; Carr et al., 2013). The extent to which it does so depends on a variety of factors, ranging from altered neurobiological functioning to maladaptive cognitive and behavioral responses to stress (Yang and Clum, 2000; Johnson et al., 2002; Caspi et al., 2003). Vulnerability to the effects of stress on cognition, affect, and suicidal behavior is impacted by genetic factors – specifically serotonin transporter (5-HTT) genotypes (Courtet et al., 2001; Antypa and Van der Does, 2010; Karg et al., 2011). However, the cognitive mechanisms through which stress affects risk for suicidal behavior, along with the moderating role of serotonin transporter genotypes, remain to be elucidated. The present pilot study sought to examine the interaction between early life stress and 5-HTT genotypes in predicting two risk factors for suicidal behavior – the brooding subtype of rumination and impulsivity, in the form of negative urgency – over time. Furthermore, this study sought to examine early life stress, ruminative subtypes, and impulsivity as predictors of suicide ideation over time.
1.1. Brooding, Negative Urgency, and Risk for Suicidal Behavior
Brooding rumination – a form of perseverative thinking that involves a passive dwelling on the reasons for one’s negative mood – has been found to be associated with depressive symptoms and suicide ideation, both concurrently and over time (Treynor et al., 2003; Chan et al., 2009; Miranda et al., 2013a; Miranda et al., 2013b). Previous research also suggests an association between early life stress and brooding. One study found that brooding partially explained the relationship between early life emotional abuse and current depressive symptoms among college students (Raes and Hermans, 2008). Another study found that brooding was associated with childhood emotional, physical, and sexual abuse among low-income pregnant women (O’Mahen et al., 2015). Finally, a recent study found that brooding and depressive symptoms partially explained the concurrent relationship between posttraumatic stress symptoms and suicide ideation among college students (Polanco-Roman et al., 2016). Thus, brooding may be one cognitive mechanism through which early life stress confers risk for suicidal thoughts and behavior.
Recent research also suggests a relationship between brooding and impulsivity, in the form of negative urgency (Valderrama et al., 2016). Negative urgency is a dimension of trait impulsivity characterized by the tendency to act rashly in response to negative emotions (Whiteside and Lynam, 2001; Cyders et al., 2007). Heterogeneity in measurement of impulsivity has led researchers to focus on identifying precise components of impulsivity that differentially predict psychopathology, with a recent meta-analysis suggesting negative urgency as the dimension of impulsivity with the strongest association to various types of psychopathology, including suicidal behavior (Berg et al., 2015). It is associated with endorsement of painful and provocative experiences among college students and clinical outpatients (Bender et al., 2011), with non-suicidal self-injury among adolescents and young adults (Glenn and Klonsky, 2010; Lockwood et al., 2017), and with suicide ideation in cross-sectional research with clinical and non-clinical samples (Johnson et al., 2017; Klonsky and May, 2010). However, no previous research of which we are aware has examined the association between negative urgency and early life stress in the context of risk for suicide ideation and/or behavior. We have previously suggested that brooding may increase risk for suicidal behavior by depleting the cognitive resources necessary to keep from responding impulsively to negative emotions (Valderrama et al., 2016). One possible mechanism through which early life stress may increase risk for suicidal behavior is through increases in brooding, and subsequently, through negative urgency.
1.2. Serotonin Transporter Genotype in Risk for Suicidal Behavior
Previous research has implicated abnormal serotonin functioning in mood disorders and suicidal behavior (Mann, 1999). Much of the research establishing an association between serotonin dysfunction, depression, and suicidal behavior has focused on the serotonin transporter (5-HTT) gene, whose promoter region is known as the 5-HTT gene-linked polymorphic region (5-HTTLPR). Genotypic variants of 5-HTTLPR include short and long forms (Helis et al., 1996). The short allele has been associated with abnormal and decreased serotonergic functioning, including lower serotonin reuptake (Karg et al., 2011).
A previous study found that the short allele of the 5-HTTLPR was most common in a group of previous violent suicide attempters with a history of major depressive disorder compared to those without major depression, but there were no differences among non-suicide-attempters with and without a history of major depression (Courtet et al., 2001). The short allele may thus relate to other variables that confer risk for depressive symptoms and suicidal behavior. Indeed, childhood maltreatment between ages 3 and 11 has been found to be associated with increased rates of major depression at ages 18–26; and number of stressful life events between ages 21 and 26 has been found to be associated with increased rate of major depression and increased probability of suicide ideation or an attempt at age 26. These associations are stronger among individuals possessing 5-HTT low expressing genotypes (one or two copies of the short allele) than 5-HTT high expressing genotypes (two copies of the long allele) (Caspi et al., 2003). Thus, 5-HTT genotypes may interact with stressful life events to moderate the association between stress and depression and/or suicidal behavior. Indeed, a meta-analysis of 54 studies reported the short allele to be significantly associated with increased stress sensitivity and subsequent greater likelihood of developing depression (Karg et al., 2011). Moreover, the short allele has been associated with other risk factors for suicidal behavior, including biased information processing, neuroticism, and HPA axis hyperactivity (Gonda et al., 2009; Pergamin-Hight et al., 2012; Miller et al., 2013).
Some research has also indicated a relationship between low 5-HTT binding and impulsivity among previous suicide attempters (Courtet et al., 2004; Lindström et al., 2004), while other research has indicated no relationship (Baca Garcia et al., 2004). However, previous suicide attempters with 5-HTT low expressing genotypes have been found to be significantly more likely to reattempt suicide during a one-year follow up than individuals with 5-HTT high expressing genotypes (Courtet et al., 2004). Notably, these studies did not assess environmental influences, such as early life stress, nor examine specific dimensions of impulsivity. Finally, a study with college students found that there was a positive association between childhood adversity and impulsivity among individuals with 5-HTT low expressing genotypes but not among those with 5-HTT high expressing genotypes. This was only the case for forms of impulsivity involving the pervasive influence of feelings on one’s orientation towards the world and for impulsivity involving the tendency for emotions to influence actions, but not for impulsivity that was not related to emotion (Carver et el., 2011).
Taken together, evidence suggests that early life stress may interact with 5-HTT low expressing genotypes to confer risk for future suicidal behavior. However, whether brooding and negative urgency are mechanisms through which early life stress interacts with 5-HTT genotypes to predict suicidal behavior is unclear. The present study sought to examine whether early life stress would predict negative urgency through brooding, and whether this effect would be moderated by 5-HTT genotypes. Further, we examined whether self-reported early life stress, brooding, and negative urgency would predict suicide ideation over time.
2. Methods
2.1. Subjects
Fifty undergraduates (44 females), ages 18–27 (M = 19.1, SD = 3.28), with and without a history of early life stress, were selected from a larger sample of 452 undergraduates (Mage = 19.0, SD = 3.3) that completed an initial survey that screened for early life stress, suicide ideation, and lifetime suicide attempt history 6 months beforehand (as part of a research participation requirement in their introductory psychology course). The racial/ethnic composition of the sample was 46% Asian, 23% Latino/a, 16% White, 6% Black, 9% of other ethnicities and was representative of the larger sample from which it was recruited.
2.2. Procedures
Participants completed initial surveys assessing early life stress, impulsivity, rumination, suicide attempt history, and suicide ideation online. To compare individuals with and without a history of early life stress, an approximately equal number of individuals with high (n = 24) and low (n = 26) early life stress scores (see below) were invited to complete self-report measures assessing impulsivity, rumination, and suicide ideation in person six months later. Further, saliva samples were collected to indicate which 5-HTTLPR polymorphism individuals possessed. Participants received a $20 gift card for their participation. Written informed consent was obtained from all participants at the beginning of the study session. Note that any participant that endorsed recent/current ideation or a suicide attempt history received an e-mail from the PI (JV) that provided contact information for the college counseling center as well as the lab telephone number in case the participant chose to inquire about other resources outside of the counseling center. All procedures received full-board approval by an Institutional Review Board.
2.3. Measures
2.3.1. Early Life Stress
The Early Trauma Inventory – Self Report-Short Form Revised (ETISR-SF) (Bremner et al., 2000), administered at baseline, is a self-report questionnaire that assesses physical, emotional, sexual, and other trauma experienced before the age of 18. Sample items include: “Did you experience the divorce or separation of your parents?”, “Were you often put down or ridiculed?” The ETISR-SF has been shown to have both good internal consistency reliability and validity (Bremner et al., 2000). Cronbach’s alpha was 0.56, 0.73, 0.78, and 0.63 for general physical trauma, physical punishment, emotional abuse, and sexual abuse, respectively.
Participants were divided into those with high versus low ETISR-SF scores, based on an analysis of the 452 participants who completed the initial survey and from which the current sample was recruited. High ETISR-SF scores were those at or higher than 12, or 1 SD above the mean (7.45), and low/average scores were those below 12.
2.3.2. Suicide Attempt History and Recent Suicide Ideation
The Suicidal Behavior Screening (SBS), a self-report tool used to screen for history of suicide attempts and recent suicide ideation, was administered at baseline and follow up. Suicide attempt history was determined by the question, “Have you ever, in your whole life, tried to kill yourself or made a suicide attempt?” Suicide ideation was determined by the question, “In the past 6 months, have you thought about killing yourself?” Seventeen out of 50 participants reported suicide ideation at baseline, and 13 participants reported suicide ideation at follow up (of the 13 reporting ideation at follow up, 8 had reported ideation at baseline). Twelve participants reported a lifetime suicide attempt history at baseline, and 13 participants (26%) reported a lifetime suicide attempt history at follow up (including the 12 individuals that had previously reported a suicide attempt at baseline).
2.3.3. Rumination
The brooding and reflection subscales of the Ruminative Responses Scale (RRS) (Nolen-Hoeksema et al., 1999) were used to assess rumination at baseline and follow up. The brooding subscale, consisting of 5 items, assesses the tendency to passively dwell on the reasons for a person’s negative mood. The reflection subscale, consisting of 5 items, assesses the tendency to actively engage in cognitive problem solving in an effort to improve one’s mood. Cronbach’s alphas at baseline were .82 and .76 for brooding and reflection, respectively, and at follow up were 0.80 and 0.78 for brooding and reflection, respectively.
2.3.4. Negative Urgency
Negative urgency was assessed at baseline and follow up via a shortened version of the UPPS Impulsive Behavior Scale (UPPS) (Whiteside and Lynam, 2001). The UPPS is a 45-item scale that assesses impulsivity across four different dimensions: lack of premeditation (i.e., not thinking through the consequences of one’s actions), negative urgency (i.e., the tendency to act rashly under conditions of negative affect), sensation seeking (i.e., the tendency to seek excitement), and lack of perseverance (i.e., not following through with a task). We used a shortened version of the form, developed by Glenn and Klonsky (2010), consisting of 16 items – four items from each impulsivity dimension that had the highest item-total correlations in the initial study (Whiteside and Lynam, 2001). Cronbach’s alphas at baseline were 0.75, 0.75, 0.78, and 0.73 for lack of premeditation, negative urgency, sensation seeking, and lack of perseverance, respectively, and at follow up were 0.86, 0.79, 0.86, and 0.82 for lack of premeditation, negative urgency, sensation seeking, and lack of perseverance, respectively.
2.3.5 5-HTTLPR Genotyping
Saliva samples were collected and sent to DNA Genotek for DNA extraction and genotyping. DNA was extracted from 700μL of saliva for genotyping one Single Nucleotide Polymorphism (SNP); rs25531, and the long versus short 5-HTTLPR polymorphism, both found in the SLC6A4 gene. The SLC6A4 gene encodes the serotonin transporter (5-HTT) responsible for terminating the action of serotonin in the synaptic cleft and reuptaking serotonin to the presynaptic neuron. Quality checks, including PicoGreen analysis, Nandrop A260/280 absorbance readings, and agarose gel electrophoresis, were completed for each sample. One sample was not analyzed due to insufficient DNA yield. 5-HTTLPR genotypes using the rs25531 full genotype (triallelic) coding for lower (SA/LG, SA/SA, LG/LG) or higher (LA/LA, LA/LG, SA/LA) expressing 5-HTT genotypes was investigated (Hu et al., 2006; Zalsman et al., 2006).
2.4 Data Analysis
Relationships among study variables at times 1 and 2 were examined by computing bivariate correlations. Independent-samples t-tests were conducted to examine differences between the two early life stress groups at baseline (Low/Average versus High) on study measures and to examine differences between suicide ideators and non-ideators at 6-month follow up. Mediation and moderated mediation analyses were conducted via the PROCESS Procedure for SPSS (Hayes, 2013). Mediation may be tested when a predictor (early life stress) relates to a mediator (brooding) and the mediator (brooding) relates to an outcome variable (negative urgency). A relation between the predictor (early life stress) and outcome (negative urgency) may or may not be present (MacKinnon et al., 2002). Mediation was examined using model 4 of the PROCESS Procedure for SPSS. Moderated mediation conditional direct effects were estimated using model 8 of the PROCESS procedure (Hayes, 2012), and conditional indirect effects were tested using bias-corrected 95% confidence intervals, calculated using a bootstrapping procedure (with n = 1000 resamples). Conditional indirect effects were considered statistically significant when their confidence intervals did not include zero. Given that a higher proportion of individuals with a low expressing genotype were Asian (n = 16; 64%), versus of other racial/ethnic groups (n = 9; 36%), χ2(1) = 5.97, p < 0.05, analyses adjusted for race/ethnicity (Asian versus Other). Finally, a logistic regression analysis was performed to assess the effects of early life stress, brooding, negative urgency, and suicide ideation at baseline on the likelihood of ideation at follow up.
3. Results
3.1. Relationship between Early Life Stress, Rumination, and Impulsivity at Baseline
Correlations between early life stress and primary study measures at baseline and follow up are shown in Table 1. The early life stress subscale of emotional abuse was significantly and positively associated with brooding at baseline and with negative urgency at follow up. General trauma, physical punishment, and sexual abuse histories were not associated with brooding, but general trauma and overall early life stress were associated with reflection. Brooding (at baseline and follow up) was significantly and positively associated with negative urgency but not with lack of premeditation, lack of perseverance, or sensation seeking (at baseline and follow up). Reflection was not significantly associated with any of the impulsivity subscales.
Table 1.
Means, Standard Deviations, and Correlations among Self-Report Measures
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | M | SD | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. ETI Total1 | … | 10.92 | 4.73 | |||||||||||||||
| 2. General Trauma1 | 0.83** | … | 2.96 | 1.98 | ||||||||||||||
| 3. Physical Punishment1 | 0.65** | 0.3** | … | 2.98 | 1.63 | |||||||||||||
| 4. Emotional Abuse1 | 0.69** | 0.38** | 0.5** | … | 3.06 | 1.65 | ||||||||||||
| 5. Sexual Abuse1 | 0.38** | 0.31* | −0.14 | −0.10 | … | 0.94 | 1.39 | |||||||||||
| 6. Brooding1 | 0.08 | −0.21 | 0.23 | 0.35* | −0.19 | … | 13.36 | 3.79 | ||||||||||
| 7. Brooding2 | 0.06 | −0.15 | 0.21 | 0.27 | −0.19 | 0.6** | … | 12.88 | 3.99 | |||||||||
| 8. Reflection1 | 0.30* | 0.33** | 0.18 | 0.12 | 0.18 | 0.25 | 0.15 | … | 11.10 | 3.58 | ||||||||
| 9. Reflect ion2 | 0.07 | 0.04 | 0.00 | 0.12 | 0.09 | 0.3** | 0.40** | 0.6** | … | 12.24 | 3.62 | |||||||
| 10. Lack of premeditation1 | −0.06 | −0.18 | −0.15 | 0.11 | 0.03 | 0.12 | 0.23 | −0.40** | −0.14 | … | 1.89 | 0.57 | ||||||
| 11. Lack of Premeditation2 | 0.01 | 0.00 | −0.18 | −0.07 | 0.26 | 0.01 | 0.22 | −0.27 | −0.01 | 0.73** | … | 1.86 | 0.63 | |||||
| 12. Negative Urgency1 | 0.19 | −0.03 | 0.09 | 0.27 | 0.11 | 0.44* | 0.4** | −0.01 | 0.05 | 0.32* | 0.32* | … | 2.80 | 0.77 | ||||
| 13. Negative Urgency2 | 0.32* | 0.13 | 0.27 | 0.30* | 0.05 | 0.29* | 0.5** | −0.05 | −0.03 | 0.21 | 0.29* | 0.63** | … | 2.85 | 0.71 | |||
| 14. Sensation Seeking1 | 0.06 | 0.03 | −0.02 | −0.07 | 0.08 | −0.06 | −0.05 | −0.07 | −0.23 | 0.08 | 0.03 | 0.21 | −0.04 | … | 2.61 | 0.84 | ||
| 15. Sensation Seeking2 | 0.21 | 0.13 | 0.10 | −0.04 | 0.25 | −0.01 | −0.02 | −0.03 | −0.14 | 0.11 | 0.11 | 0.23 | 0.02 | 0.82** | … | 2.66 | 0.86 | |
| 16. Lack of Perseverance1 | −0.23 | −0.31* | −0.18 | 0.14 | −0.23 | 0.19 | 0.24 | 0.02 | 0.22 | 0.32* | 0.27 | 0.12 | −0.17 | 0.00 | −0.03 | … | 1.86 | 0.58 |
| 17. Lack of Perseverance2 | −0.14 | −0.12 | −0.16 | 0.11 | −0.18 | 0.02 | 0.17 | −0.03 | 0.13 | 0.23 | 0.37** | 0.10 | −0.01 | −0.14 | −0.17 | 0.7** | 1.98 | 0.63 |
Measured at baseline
Measured at follow up
p < .05
p < .01
There were no statistically significant group differences on the study variables (brooding, reflection, impulsivity subscales) at baseline or follow up.
3.2. Examining Brooding as a Mediator of the Relationship between Early Life Stress and Negative Urgency and the Moderating Role of 5-HTTLPR Genotypes
We examined whether brooding, in association with 5-HTT low expressing genotypes, would mediate the relationship between early life emotional abuse at baseline and negative urgency at follow up (Note that early life stress total scores did not predict brooding and were thus not used in these analyses). We hypothesized that the relationship between early life emotional abuse and negative urgency would be mediated by brooding among individuals with the 5-HTT low expressing genotypes (n = 25) and not among individuals with the 5-HTT high expressing genotypes (n = 24). There was an indirect effect of early life emotional abuse on negative urgency through brooding (indirect effect = 0.05; 95% CI = 0.01 – 0.13).
We then examined whether this indirect relationship was moderated by the triallelic 5-HTTLPR genotypes. We found an indirect effect of early life emotional abuse [indirect effect = 0.08; 95% CI = 0.02 – 0.18] on negative urgency through brooding among individuals with the 5-HTT low expressing genotypes (See Figure 1). However, we did not find an indirect effect of early life emotional abuse [indirect effect = −0.004; 95% CI = −0.09 – 0.06] on negative urgency through brooding among individuals with the 5-HTT high expressing genotypes. Thus, 5-HTT low expressing genotypes moderated the indirect relationship between early life emotional abuse and negative urgency though brooding (Index of moderated mediation = −0.09; 95% CI = −0.22 – −0.01).
Figure 1.
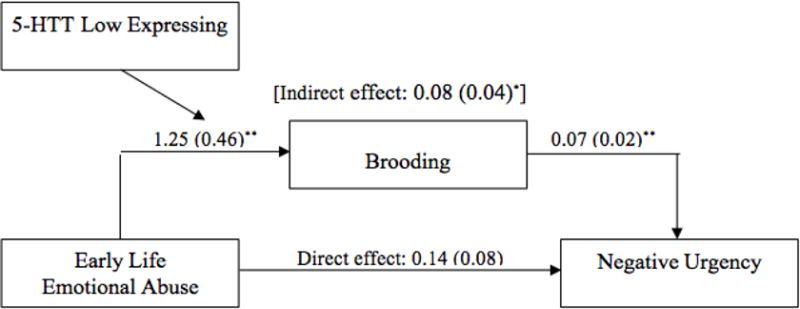
Early life emotional abuse does not predict negative urgency. Early life emotional abuse significantly predicts brooding. There was an indirect effect of early life emotional abuse on negative urgency through brooding for individuals possessing 5-HTT low expressing genotypes. There was no indirect effect of early life emotional abuse on negative urgency through brooding for individuals possessing 5-HTT high expressing genotypes (not shown; indirect effect: −0.004 (0.04)). Values shown are unstandardized coefficients, with standard errors in parentheses. *p < 0.05; ** p < 0.01.
3.3. Suicide Ideation at 6-month follow-up
Recent ideators (as assessed at 6-month follow up) had significantly higher lack of premeditation and negative urgency scores at follow up than did non-ideators (see Table 2). No other significant differences between ideators and non-ideators were found on measures of early life stress, rumination, and trait impulsivity. Furthermore, recent suicide ideators with a previous suicide attempt history (n = 4) had significantly higher early life stress scores (M = 15.50, SD = 3.11) than did recent ideators without a suicide attempt history (n = 9) (M = 9.44, SD = 3.81), t(11) = 2.77, p < 0.05. No other significant differences were found in measures of rumination and trait impulsivity among recent ideators with and without a suicide attempt history.
Table 2.
Differences between Suicide Ideators and Non-Ideators (assessed at 6-month follow up) on Study Measures
| Group | ||||
|---|---|---|---|---|
|
| ||||
| Current Suicide Ideators (n = 13) |
Non-Ideators (n = 37) |
|||
|
| ||||
| Measures | M (SD) | p | d | |
| ETI | ||||
| ETI Total1 | 11.31 (4.53) | 10.78 (4.85) | 0.74 | 0.11 |
| RRS | ||||
| Brooding2* | 14.92 (3.95) | 12.16 (3.80) | < 0.05 | 0.71 |
| Reflection2* | 14.08 (3.35) | 11.59 (3.53) | < 0.05 | 0.72 |
| UPPS-P SF Impulsivity Scale2 | ||||
| Lack of Premeditation** | 2.33 (0.52) | 1.69 (0.58) | <0.01 | 1.16 |
| Negative Urgency* | 3.21 (0.58) | 2.72 (0.72) | <0.05 | 0.75 |
| Sensation Seeking | 2.89 (0.88) | 2.57 (0.86) | 0.27 | 0.37 |
| Lack of Perseverance | 2.13 (0.64) | 1.93 (0.62) | 0.31 | 0.32 |
Measured at baseline
Measured at follow up
p < 0.05
p < 0.01
We also examined the effects of early life stress, brooding, negative urgency, and suicide ideation at baseline on suicide ideation at follow up. The logistic regression model explained 49.6% (Nagelkerke R2) of the variance in ideation and correctly classified 80% of cases. Both negative urgency (p < 0.01) and ideation at baseline (p < 0.01) significantly contributed to the model (see Table 3). Each unit increase in negative urgency was associated with 16.2 times higher odds of reporting suicide ideation at follow up, versus no ideation. Suicide ideation at baseline was associated with 13.8 times higher odds of reporting suicide ideation at follow up, versus no ideation. Brooding and early life stress were not significantly associated with odds of suicide ideation at followup, after adjusting for baseline suicide ideation.
Table 3.
Logistic Regression Predicting Suicide Ideation at Follow up from Ideation, Early Life Stress, Negative Urgency, and Brooding at Baseline
| Predictor | O.R. | 95 % CI | p |
|---|---|---|---|
| Baseline Ideation* | 13.81 | 1.87 – 101.60 | < 0.05 |
| Early Life Stress | 0.96 | 0.80 – 1.15 | 0.65 |
| Negative Urgency** | 16.20 | 2.34 – 112.19 | < 0.01 |
| Brooding | 0.77 | 0.57 – 1.04 | 0.09 |
p < 0.05;
p < 0.01
4. Discussion
The present study examined whether the relationship between early life stress and negative urgency – through the brooding subtype of rumination – would be moderated by 5-HTTLPR genotype variants. Early life emotional abuse was associated with brooding at baseline and with negative urgency at 6-month follow up, and brooding was associated with negative urgency (but not with lack of premeditation, lack of perseverance, or sensation seeking). Further, we found an indirect effect of early life emotional abuse on negative urgency through brooding for individuals possessing 5-HTT low expressing genotypes, but not for individuals possessing 5-HTT high expressing genotypes. Finally, we found that negative urgency predicted suicide ideation at 6-month follow up.
We found early life stress in the form of emotional abuse to be significantly and positively associated with brooding, but not with reflection. This is in contrast with a previous finding that early life emotional abuse was associated with both brooding and reflection among college students (Raes and Hermans, 2008). However, it is consistent with other previous research suggesting that while brooding, but not reflection, was associated with childhood emotional, physical, and sexual abuse, only childhood emotional abuse prospectively predicted brooding (O’Mahen, 2015). Early life emotional abuse proved to be unique among the different aspects of early life stress in the present study, with respect to its association with 5-HTT genotypes, brooding, and negative urgency.
These findings are also consistent with previous research linking low 5-HTT binding with impulsivity (Lindström et al., 2004) and with suicidal behavior (Courtet et al., 2004) but extends such research by examining the role of early life stress in this relationship and by examining a specific dimension of impulsivity in the form of negative urgency. To our knowledge, the present study is the first to reveal an association between early life stress and negative urgency through brooding that is moderated by 5-HTTLPR – specifically, an indirect relationship between early life emotional abuse and negative urgency via brooding among individuals possessing 5-HTT low expressing genotypes. We found no such relationships with the other forms of early life stress assessed in our study. Importantly, higher negative urgency, not brooding, was significantly related to increased risk of suicide ideation at follow-up, providing the first preliminary evidence of which we are aware that negative urgency predicts suicide ideation over time. Our findings suggest that rumination, in the form of brooding, and impulsivity, in the form of negative urgency, may result from the interaction between early life emotional abuse and 5-HTT low expressing genotypes. Moreover, 5-HTT high expressing genotypes may provide a buffer, such that early life emotional abuse may have less of an impact on later emotion regulation strategies. The interaction between 5-HTT low expressing genotypes and early life stress has previously been linked with depression, anxiety sensitivity, suicidal behavior, PTSD, smaller hippocampal volume, and deficits in cognitive and emotional processing (Gibb et al., 2006; Kaufman et al., 2006; Roy et al., 2007; Xie et al., 2009; Frodl et al., 2010; Owens et al., 2012; Sharpley et al., 2014), and 5-HTT may interact with certain types of stress in relation to outcomes such as history of depressive symptoms and suicide attempts (Gibb et al., 2006; Grabe et al., 2012). The present pilot study extends such research by suggesting psychological factors that may arise as a result of early life stress and that should be studied as potential mechanisms in the relationship between early life stress and risk for suicidal ideation using a larger sample.
Consistent with previous research, our study found no direct relationship between 5-HTTLPR allele groups and rumination (Beevers et al., 2009). However, a ruminative cognitive style has previously been found to be associated with early life emotional abuse and 5-HTT low expressing genotypes among young adults (Antypa and Van der Does, 2010). Interestingly, possessing a 5-HTT low expressing genotype is associated with dysregulated neural activity in emotion regulatory circuits (Murphy et al., 2013; Outhred et al., 2016). Such dysregulation has been associated with negatively biased information processing that is more likely to occur when an individual has experienced early life stress (Williams et al. 2009; Vrijsen et al., 2015). Attending toward negative information might contribute to maladaptive cognitive and behavioral patterns in the form of brooding and negative urgency. However, how exactly 5-HTT low expressing genotypes and early life stress confer risk for negative psychological outcomes such as brooding or negative urgency require further study, given numerous potential factors that may impact how people cope with stress. Previous research has established the important role of serotonin in modulating emotional behavior (Hariri and Holmes, 2006), but further research is necessary to understand the relationship between specific types of early life stress, 5-HTT genotypes, brooding, and negative urgency.
Some study limitations should be noted. First, the present sample was small and ethnically heterogeneous, and thus, our genetic results may not extrapolate to a larger sample of individuals. Indeed, 5-HTTLPR genotype and allele frequencies differ across ethnic populations (Kunugi et al., 1997; Serretti et al., 2007). Second, the present sample was non-clinical and primarily female, and thus, our results may not generalize to a clinical setting or to males. Third, our measure of early life stress did not examine the exact year(s) in which early life adversity occurred. Studies involving animal models of stress suggest that the impact of stress depends on timing of the stressor during development (Parker et al., 2004; Lyons et al., 2010). We could not assess how stress interacted with any potential critical time periods of central nervous system, neuroendocrine system, behavioral, or cognitive development. Relatedly, early life stress was self-reported in adulthood rather than examined in childhood or adolescence, and this measure may thus be subject to self-report bias and not an accurate indicator of abuse that occurred prior to adulthood. Additionally, given that brooding and negative urgency are traits, the present research, which involved only a 6-month follow-up period during adulthood, did not capture a time period in which one might expect changes in brooding and negative urgency to develop in response to stress experienced earlier in life. Future research should capture change over critical periods of development associated with earlier stressors. Finally, limited statistical power prevented us from having greater reliability in our logistic regression assessing suicide ideation at follow-up. Limited statistical power further prevented us from testing a fuller meditational model examining whether genotype would moderate an indirect relationship between early life stress and suicide ideation through both brooding and negative urgency. This question should be examined in future research with a larger sample and statistical models that adjust for additional factors, such as depression, that were not measured in the present study.
5. Concluding Comments
The present study suggests that early life emotional abuse contributes to negative urgency via its relationship to brooding and low-expressing 5-HTT genotypes. These findings have both clinical and research implications. Whether early life adversity – particularly emotional abuse, has an impact on factors that increase risk for suicide ideation and behavior depends on genotype, and clinical risk assessments should thus consider both. Further, cognitive therapies should focus on reducing negative urgency in an effort to prevent suicidal behavior in at-risk individuals. From a research perspective, future investigations should continue to examine the relationship between serotonergic circuitry in corticolimbic structures, emotion regulation, and epigenetic mechanisms.
Highlights.
We examine early life stress, brooding, negative urgency, and the 5-HTTLPR polymorphism
Early life emotional abuse associated with negative urgency
Indirect effect of early life emotional abuse on negative urgency through brooding among 5-HTT low expressing genotypes
Acknowledgments
This project was funded by RISE Grant GM060665 and a CUNY Graduate Center Doctoral Student Research Grant. Thanks to Erika Kincaid, Diana Ribeiro, Alliya Shafi, Audrey Sheppard, and Sylvia Tesseyman for their assistance with data collection.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Antypa N, Van der Does AJW. Serotonin transporter gene, childhood emotional abuse and cognitive vulnerability to depression. Genes Brain Behav. 2010;9:615–620. doi: 10.1111/j.1601-183X.2010.00593.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baca-Garcia E, Vaquero C, Diaz-Sastre C, García-Resa E, Saiz-Ruiz J, Fernandez-Piqueras J, de Leon J. Lack of association between the serotonin transporter promoter gene polymorphism and impulsivity or aggressive behavior among suicide attempters and healthy volunteers. Psychiatry Res. 2004;126:99–106. doi: 10.1016/j.psychres.2003.10.007. [DOI] [PubMed] [Google Scholar]
- Beevers CG, Wells TT, McGeary JE. The BDNF Val66Met polymorphism is associated with rumination in healthy adults. Emotion. 2009;9:579. doi: 10.1037/a0016189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berg JM, Latzman RD, Bliwise NG, Lilienfeld S. Parsing the heterogeneity of impulsivity: A meta-analytic review of the behavioral implications of the UPPS for psychopathology. Psychol Assess. 2015;27:1129–1146. doi: 10.1037/pas0000111. [DOI] [PubMed] [Google Scholar]
- Bender TW, Gordon KH, Bresin K, Joiner TE. Impulsivity and suicidality: the mediating role of painful and provocative experiences. J Affect Disord. 2011;129:301–307. doi: 10.1016/j.jad.2010.07.023. [DOI] [PubMed] [Google Scholar]
- Bremner JD, Vermetten E, Mazure CM. Development and preliminary psychometric properties of an instrument for the measurement of childhood trauma: the early trauma inventory. Depress Anxiety. 2000;12:1–12. doi: 10.1002/1520-6394(2000)12:1<1::AID-DA1>3.0.CO;2-W. [DOI] [PubMed] [Google Scholar]
- Carr CP, Martins CMS, Stingel AM, Lemgruber VB, Juruena MF. The role of early life stress in adult psychiatric disorders: A systematic review. J Nerv Ment Dis. 2013;201:1007–1020. doi: 10.1097/NMD.0000000000000049. [DOI] [PubMed] [Google Scholar]
- Carver CS, Johnson SL, Joormann J, Kim Y, Nam JY. Serotonin transporter polymorphism interacts with childhood adversity to predict aspects of impulsivity. Psychol Sci. 2011;22:589–595. doi: 10.1177/0956797611404085. [DOI] [PubMed] [Google Scholar]
- Caspi A, Sugden K, Moffitt TE, Taylor A, Craig IW, Harrington H, McClay J, Mill J, Martin J, Braithwaite A, Poulton R. Influence of life stress on depression: moderation by a polymorphism in the 5-HTT gene. Science. 2003;301:386–389. doi: 10.1126/science.1083968. [DOI] [PubMed] [Google Scholar]
- Chan S, Miranda R, Surrence K. Subtypes of rumination in the relationship between negative life events and suicidal ideation. Arch Suicide Res. 2009;13:123–135. doi: 10.1080/13811110902835015. [DOI] [PubMed] [Google Scholar]
- Courtet P, Baud P, Abbar M, Boulenger JP, Castelnau D, Mouthon D, Malafosse A, Buresi C. Association between violent suicidal behavior and the low activity allele of the serotonin transporter gene. Mol Psychiatry. 2001;6:338–341. doi: 10.1038/sj.mp.4000856. [DOI] [PubMed] [Google Scholar]
- Courtet P, Picot MC, Bellivier F, Torres S, Jollant F, Michelon C, Castelnau C, Astruc B, Buresi C, Malafosse A. Serotonin transporter gene may be involved in short-term risk of subsequent suicide attempts. Biol Psychiatry. 2004;55:46–51. doi: 10.1016/j.biopsych.2003.07.004. [DOI] [PubMed] [Google Scholar]
- Cyders MA, Smith GT, Spillane NS, Fischer S, Annus AM, Peterson C. Integration of impulsivity and positive mood to predict risky behavior: development and validation of a measure of positive urgency. Psychol Assess. 2007;19:107–118. doi: 10.1037/1040-3590.19.1.107. [DOI] [PubMed] [Google Scholar]
- Frodl T, Reinhold E, Koutsouleris N, Donohoe G, Bondy B, Reiser M, Möller HJ, Meisenzahl EM. Childhood stress, serotonin transporter gene and brain structures in major depression. Neuropsychopharmacology. 2010;35:1383–1390. doi: 10.1038/npp.2010.8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibb BE, McGeary JE, Beevers CG, Miller IW. Serotonin transporter (5-HTTLPR) genotype, childhood abuse, and suicide attempts in adult psychiatric inpatients. Suicide Life Threat Behav. 2006;36:687–693. doi: 10.1521/suli.2006.36.6.687. [DOI] [PubMed] [Google Scholar]
- Gonda X, Fountoulakis KN, Juhasz G, Rihmer Z, Lazary J, Laszik A, Akiskal HS, Bagdy G. Association of the s allele of the 5-HTTLPR with neuroticism-related traits and temperaments in a psychiatrically healthy population. Eur Arch Psychiaty Clin Neurosci. 2009;259:106–113. doi: 10.1007/s00406-008-0842-7. [DOI] [PubMed] [Google Scholar]
- Glenn CR, Klonsky ED. A multimethod analysis of impulsivity in non-suicidal self-injury. Pers Disord: Theory Res Treat. 2010;1:67–75. doi: 10.1037/a0017427. [DOI] [PubMed] [Google Scholar]
- Grabe HJ, Schwahn C, Mahler J, Appel K, Schulz A, Spitzer C, Fenske K, Barnow S, Freyberger HJ, Teumer A, Petersmann A. Genetic epistasis between the brain-derived neurotrophic factor Val66Met polymorphism and the 5-HTT promoter polymorphism moderates the susceptibility to depressive disorders after childhood abuse. Prog Neuropsychopharmacol Biol Psychiatry. 2012;36:264–270. doi: 10.1016/j.pnpbp.2011.09.010. [DOI] [PubMed] [Google Scholar]
- Hariri AR, Holmes A. Genetics of emotional regulation: the role of the serotonin transporter in neural function. Trends Cogn Sci. 2006;10:182–191. doi: 10.1016/j.tics.2006.02.011. [DOI] [PubMed] [Google Scholar]
- Hayes AF. Introduction to mediation, moderation, and conditional process analysis: A regression-based approach. New York: Guilford Press; 2013. [Google Scholar]
- Hu XZ, Lipsky RH, Zhu G, Akhtar LA, Taubman J, Greenberg BD, Xu K, Arnold PD, Richter MA, Kennedy JL, Murphy DL. Serotonin transporter promoter gain-of-function genotypes are linked to obsessive-compulsive disorder. Am J Hum Genet. 2006;78:815–826. doi: 10.1086/503850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson JG, Cohen P, Gould MS, Kasen S, Brown J, Brook JS. Childhood adversities, interpersonal difficulties, and risk for suicide attempts during late adolescence and early adulthood. Arch Gen Psychiatry. 2002;59:741–749. doi: 10.1001/archpsyc.59.8.741. [DOI] [PubMed] [Google Scholar]
- Johnson SL, Carver CS, Tharp JA. Suicidality in Bipolar Disorder: The role of emotion-triggered impulsivity. Suicide Life Threat Behav. 2017;47:177–192. doi: 10.1111/sltb.12274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karg K, Burmeister M, Shedden K, Sen S. The serotonin transporter promoter variant (5-HTTLPR), stress, and depression meta-analysis revisited: evidence of genetic moderation. Arch Gen Psychiatry. 2011;68:444–454. doi: 10.1001/archgenpsychiatry.2010.189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaufman J, Yang BZ, Douglas-Palumberi H, Grasso D, Lipschitz D, Houshyar S, Krystal JH, Gelernter J. Brain-derived neurotrophic factor–5-HTTLPR gene interactions and environmental modifiers of depression in children. Biol Psychiatry. 2006;59:673–680. doi: 10.1016/j.biopsych.2005.10.026. [DOI] [PubMed] [Google Scholar]
- Klonsky ED, May AM. Rethinking impulsivity in suicide. Suicide Life Threat Behav. 2010;40:612–619. doi: 10.1521/suli.2010.40.6.612. [DOI] [PubMed] [Google Scholar]
- Kunugi H, Hattori M, Kato T, Tatsumi M, Sakai T, Hirose T, Nanko S. Serotonin transporter gene polymorphisms: Ethnic difference and possible association with bipolar affective disorder. Mol Psychiatry. 1997;2:457–462. doi: 10.1038/sj.mp.4000334. [DOI] [PubMed] [Google Scholar]
- Lindström MB, Ryding E, Bosson P, Ahnlide JA, Rosén I, Träskman-Bendz L. Impulsivity related to brain serotonin transporter binding capacity in suicide attempters. Eur Neuropsychopharmacol. 2004;14:295–300. doi: 10.1016/j.euroneuro.2003.11.001. [DOI] [PubMed] [Google Scholar]
- Lockwood J, Daley D, Townsend E, Sayal K. Impulsivity and self-harm in adolescence: A systematic review. Eur Child Adolesc Psychiatry. 2017;26:387–402. doi: 10.1007/s00787-016-0915-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lyons DM, Parker KJ, Schatzberg AF. Animal models of early life stress: implications for understanding resilience. Dev Psychobiol. 2010;52:402–410. doi: 10.1002/dev.20429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacKinnon DP, Lockwood CM, Hoffman JM, West SG, Sheets V. A comparison of methods to test mediation and other intervening variable effects. Psychol Methods. 2002;7:83–104. doi: 10.1037/1082-989x.7.1.83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mann JJ. Role of the serotonergic system in the pathogenesis of major depression and suicidal behavior. Neuropsychopharmacology. 1999;21:99S–105S. doi: 10.1016/S0893-133X(99)00040-8. [DOI] [PubMed] [Google Scholar]
- Miller R, Wankerl M, Stalder T, Kirschbaum C, Alexander N. The serotonin transporter gene-linkedpolymorphic region (5-HTTLPR) and cortisol stress reactivity: A meta-analysis. Mol Psychiatry. 2013;18:1018–1024. doi: 10.1038/mp.2012.124. [DOI] [PubMed] [Google Scholar]
- Miranda R, Tsypes A, Gallagher M, Rajappa K. Rumination and hopelessness as mediators of the relation between perceived emotion dysregulation and suicidal ideation. Cognit Ther Res. 2013a;37:786–795. [Google Scholar]
- Miranda R, Valderrama J, Tsypes A, Gadol E, Gallagher M. Cognitive inflexibility and suicidal ideation: mediating role of brooding and hopelessness. Psychiatry Res. 2013b;210:174–181. doi: 10.1016/j.psychres.2013.02.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murphy SE, Norbury R, Godlewska BR, Cowen PJ, Mannie ZM, Harmer CJ, Munafo MR. The effect of the serotonin transporter polymorphism (5-HTTLPR) on amygdala function: a meta-analysis. Mol Psychiatry. 2013;18:512–520. doi: 10.1038/mp.2012.19. [DOI] [PubMed] [Google Scholar]
- Nolen-Hoeksema S, Larson J, Grayson C. Explaining the gender difference in depressive symptoms. J Pers Soc Psychol. 1999;77:1061–1072. doi: 10.1037//0022-3514.77.5.1061. [DOI] [PubMed] [Google Scholar]
- O’Mahen HA, Karl A, Moberly N, Fedock G. The association between childhood maltreatment and emotion regulation: Two different mechanisms contributing to depression? J Affect Disord. 2015;174:287–295. doi: 10.1016/j.jad.2014.11.028. [DOI] [PubMed] [Google Scholar]
- Outhred T, Das P, Dobson-Stone C, Felmingham KL, Bryant RA, Nathan PJ, Gin SM, Kemp AH. Impact of 5-HTTLPR on SSRI serotonin transporter blockade during emotion regulation: A preliminary fMRI study. J Affect Disord. 2016;196:11–19. doi: 10.1016/j.jad.2016.02.019. [DOI] [PubMed] [Google Scholar]
- Owens M, Goodyer IM, Wilkinson P, Bhardwaj A, Abbott R, Croudace T, Dunn V, Jones PB, Walsh ND, Ban M, Sahakian BJ. 5-HTTLPR and early childhood adversities moderate cognitive and emotional processing in adolescence. PLoS ONE. 2012;7 doi: 10.1371/journal.pone.0048482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parker KJ, Buckmaster CL, Schatzberg AF, Lyons DM. Prospective investigation of stress inoculation in young monkeys. Arch Gen Psychiatry. 2004;61:933–941. doi: 10.1001/archpsyc.61.9.933. [DOI] [PubMed] [Google Scholar]
- Pergamin-Hight L, Bakermans-Kranenburg MJ, Van Ijzendoorn MH, Bar-Haim Y. Variations in the promoter region of the serotonin transporter gene and biased attention for emotional information: a meta-analysis. Biol Psychiatry. 2012;71:373–379. doi: 10.1016/j.biopsych.2011.10.030. [DOI] [PubMed] [Google Scholar]
- Polanco-Roman L, Gomez J, Miranda R, Jeglic E. Stress-related symptoms and suicidal ideation: The roles of rumination and depressive symptoms vary by gender. Cognit Ther Res. 2016;40:606–616. doi: 10.1007/s10608-016-9782-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raes F, Hermans D. On the mediating role of subtypes of rumination in the relationship between childhood emotional abuse and depressed mood: brooding versus reflection. Depress Anxiety. 2008;25:1067–1070. doi: 10.1002/da.20447. [DOI] [PubMed] [Google Scholar]
- Roy A, Hu XZ, Janal MN, Goldman D. Interaction between childhood trauma and serotonin transporter gene variation in suicide. Neuropsychopharmacology. 2007;32:2046–2052. doi: 10.1038/sj.npp.1301331. [DOI] [PubMed] [Google Scholar]
- Santa Mina EE, Gallop RM. Childhood sexual and physical abuse and adult self-harm and suicidal behaviour: a literature review. Can J Psychiatry. 1998;43:793–800. doi: 10.1177/070674379804300803. [DOI] [PubMed] [Google Scholar]
- Serretti A, Kato M, De Ronchi D, Kinoshita T. Meta-analysis of serotonin transporter gene promoter polymorphism (5-HTTLPR) association with selective serotonin reuptake inhibitor efficacy in depressed patients. Mol Psychiatry. 2007;12:247–257. doi: 10.1038/sj.mp.4001926. [DOI] [PubMed] [Google Scholar]
- Sharpley CF, Palanisamy SK, Glyde NS, Dillingham PW, Agnew LL. An update on the interaction between the serotonin transporter promoter variant (5- HTTLPR), stress and depression, plus an exploration of non-confirming findings. Behav Brain Res. 2014;273:89–105. doi: 10.1016/j.bbr.2014.07.030. [DOI] [PubMed] [Google Scholar]
- Treynor W, Gonzalez R, Nolen-Hoeksema S. Rumination reconsidered: a psychometric analysis. Cognit Ther Res. 2003;27:247–259. [Google Scholar]
- Valderrama J, Miranda R, Jeglic E. Ruminative Subtypes and Impulsivity in Risk for Suicidal Behavior. Psychiatry Res. 2016;236:15–21. doi: 10.1016/j.psychres.2016.01.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vrijsen JN, Tendolkar I, Arias-Vásquez A, Franke B, Schene AH, Fernández G, van Oostrom I. Interaction of the 5-HTTLPR and childhood trauma influences memory bias in healthy individuals. J Affect Disord. 2015;186:83–89. doi: 10.1016/j.jad.2015.06.008. [DOI] [PubMed] [Google Scholar]
- Wang Y, Sareen J, Afifi TO, Bolton SL, Johnson EA, Bolton JM. Recent Stressful Life Events and Suicide Attempt. Psychiatr Ann. 2012;42:101–108. [Google Scholar]
- Williams LM, Gatt JM, Schofield PR, Olivieri G, Peduto A, Gordon E. ‘Negativity bias’ in risk for depression and anxiety: Brain–body fear circuitry correlates, 5-HTT-LPR and early life stress. Neuroimage. 2009;47:804–814. doi: 10.1016/j.neuroimage.2009.05.009. [DOI] [PubMed] [Google Scholar]
- Whiteside SP, Lynam DR. The five factor model and impulsivity: Using a structural model of personality to understand impulsivity. Pers Individ Dif. 2001;30:669–689. [Google Scholar]
- Xie P, Kranzler HR, Poling J, Stein MB, Anton RF, Brady K, Weiss RD, Farrer L, Gelernter J. Interactive effect of stressful life events and the serotonin transporter 5-HTTLPR genotype on posttraumatic stress disorder diagnosis in 2 independent populations. Arch Gen Psychiatry. 2009;66:1201–1209. doi: 10.1001/archgenpsychiatry.2009.153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang B, Clum GA. Childhood stress leads to later suicidality via its effect on cognitive functioning. Suicide Life Threat Behav. 2000;30:183–198. [PubMed] [Google Scholar]
- Yen S, Shea MT, Sanislow CA, Skodol AE, Grilo CM, Edelen MO, Stout RL, Morey LC, Zanarini MC, Markowitz JC, McGlashan TH, Daversa MT, Gunderson JG. Personality traits as prospective predictors of suicide attempts. Acta Psychiatr Scand. 2009;120:222–229. doi: 10.1111/j.1600-0447.2009.01366.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zalsman G, Huang YY, Oquendo MA, Burke AK, Hu XZ, Brent DA, Ellis SP, Goldman D, Mann JJ. Association of a triallelic serotonin transporter gene promoter region (5-HTTLPR) polymorphism with stressful life events and severity of depression. Am J Psychiatry. 2006;163:1588–1593. doi: 10.1176/ajp.2006.163.9.1588. [DOI] [PubMed] [Google Scholar]