

Abstract
Osteoclasts are bone-resorbing cells that are derived from hematopoietic precursor cells and require macrophage-colony stimulating factor and receptor activator of nuclear factor-κB ligand (RANKL) for their survival, proliferation, differentiation, and activation. The binding of RANKL to its receptor RANK triggers osteoclast precursors to differentiate into osteoclasts. This process depends on RANKL-RANK signaling, which is temporally regulated by various adaptor proteins and kinases. Here we summarize the current understanding of the mechanisms that regulate RANK signaling during osteoclastogenesis. In the early stage, RANK signaling is mediated by recruiting adaptor molecules such as tumor necrosis factor receptor-associated factor 6 (TRAF6), which leads to the activation of mitogen-activated protein kinases (MAPKs), and the transcription factors nuclear factor-κB (NF-κB) and activator protein-1 (AP-1). Activated NF-κB induces the nuclear factor of activated T-cells cytoplasmic 1 (NFATc1), which is the key osteoclastogenesis regulator. In the intermediate stage of signaling, the co-stimulatory signal induces Ca2+ oscillation via activated phospholipase Cγ2 (PLCγ2) together with c-Fos/AP-1, wherein Ca2+ signaling facilitates the robust production of NFATc1. In the late stage of osteoclastogenesis, NFATc1 translocates into the nucleus where it induces numerous osteoclast-specific target genes that are responsible for cell fusion and function.
Keywords: nuclear factor-κB, nuclear factor of activated T-cells cytoplasmic 1, osteoclasts, receptor activator of nuclear factor-κB, tumor necrosis factor receptor-associated factors
INTRODUCTION
The mature skeleton is maintained by bone remodeling, which is a dynamic process in which the bone is first resorbed by osteoclasts, followed by bone formation in the lacunae by osteoblasts (Zaidi, 2007). Thus, the coupling of osteoclasts and osteoblasts is strictly regulated with regard to both space and time. Osteoclasts are bone-resorbing multinucleated cells that originate from hematopoietic precursors that are formed in the bone marrow (Asagiri and Takayanagi, 2007; Boyle et al., 2003; Karsenty and Wagner, 2002; Teitelbaum, 2000). Macrophage-colony stimulating factor (M-CSF) and the receptor activator of nuclear factor-κB ligand (RANKL; encoded by Tnfsf11), which are expressed by osteoblast-lineage cells (Fumoto et al., 2014) or osteocytes (Nakashima et al., 2011; Xiong et al., 2011), are required for the differentiation of osteoclast precursors into mature osteoclasts (Teitelbaum and Ross, 2003). The released RANKL binds to its receptor RANK (encoded by Tnfrsf11a), which is expressed via M-CSF stimulation on the surface of osteoclast precursor cells. Mice with genetic deletions of Tnfsf11 or Tnfrsf11a exhibit severe osteopetrosis accompanied by defective tooth eruption owing to the complete death of their osteoclasts (Dougall et al., 1999; Kong et al., 1999; Li et al., 2000). RANKL-RANK signaling is also negatively regulated by osteoprotegerin (encoded by Tnfrsf11b), which is a soluble decoy receptor for RANKL that obstructs RANKL binding to RANK (Yasuda et al., 1998).
RANK is a member of the tumor necrosis factor receptor (TNFR) superfamily that lacks intrinsic enzymatic activity that is required for activating downstream signaling molecules (Cheng et al., 2003). Thus, RANK transduces intracellular signals by recruiting adaptor molecules such as TNFR-associated factors (TRAFs), which then activate mitogen activated protein kinases (MAPKs) and nuclear factor-κB (NF-κB) (Darnay et al., 1998; Kobayashi et al., 2001; Wong et al., 1998). RANK signaling is controlled via the spatiotemporal regulation of downstream signaling, which results in the commitment of monocyte/macrophage precursor cells to the osteoclast lineage and the activation of mature osteoclasts.
During the initiation step of RANK signaling, RANKL binding to RANK leads to the recruitment of adaptor proteins such as TRAF6 to the trimerized cytoplasmic tail of RANK and the rapid activation of MAPKs, NF-κB, and activator protein-1 (AP-1) (Kobayashi et al., 2001; Teitelbaum and Ross, 2003). The subsequent signaling process is characterized by amplifying the nuclear factor of activated T-cells cytoplasmic 1 (NFATc1) via orchestrated signaling of activated AP-1 and co-stimulatory signal-mediated intracellular Ca2+ oscillation (Takayanagi et al., 2002). Finally, in the late stage of signaling, the osteoclastogenic genes transcribed by the activated NFATc1 regulate the multi-nucleation and bone resorption function of osteoclasts. Here we describe the current understanding of the molecular mechanisms involved in this process and the recent findings regarding osteoclastogenic regulation of RANK signaling.
RANK SIGNALING AND TRAF ADAPTOR PROTEINS
RANKL binding to RANK induces the trimerization of RANK and the recruitment of an adaptor protein, TRAF6, via three TRAF6-binding sites in its C-terminal cytoplasmic tail (Wong et al., 1998; Ye et al., 2002), thereby initiating the downstream signaling cascade (Fig. 1). Other TRAF family members such as TRAF2, TRAF3, and TRAF5 can bind to RANK and activate the transcription factors required for osteoclast differentiation. However, studies on TRAF6-deficient mice revealed that TRAF6 is the major adaptor protein responsible for mediating signaling cascades that are activated by RANKL (Lomaga et al., 1999; Naito et al., 1999). Activated TRAF6 stimulates NF-κB activity by activating the IκB kinase (IKK) complex, either via atypical protein kinase C (aPKC) or TGFβ-activated kinase 1 (TAK1)-dependent phosphorylation, which is a process that requires IKKγ (also called NEMO) ubiquitination to achieve optimal activation (Duran et al., 2004; Ghosh and Karin, 2002; Mizukami et al., 2002). The scaffolding protein p62 binds to TRAF6 and interacts with aPKCs, resulting in the formation of a multimeric protein complex that regulates IKKβ (Duran et al., 2004). TRAF6 also forms complexes with TAK1 and the adaptor proteins TAK1-binding protein 1 (TAB1) and TAB2 (Mizukami et al., 2002). After its activation, TAK1 phosphorylates the IKK complex, thereby activating the NF-κB pathway (Ninomiya-Tsuji et al., 1999). NF-κB signaling is essential for osteoclastogenesis, and NF-κB p50/p52 double-knockout mice exhibit marked osteopetrosis because of the failure in osteoclast formation (Franzoso et al., 1997; Iotsova et al., 1997). AP-1 transcription factor comprising Fos (c-Fos, FosB, Fra-1, and Fra-2), Jun (c-Jun, JunB, and JunD), and ATF (ATFa, ATF2, ATF4, and B-ATF) family members is also activated via the induction of c-Fos by adaptor proteins (Grigoriadis et al., 1994; Wagner, 2002). c-Fos knockout mice and transgenic mice that over-express dominant negative c-Jun exhibit severe osteopetrosis (Ikeda et al., 2004; Wang et al., 1992). These results show that transcription factors such as NF-κB and AP-1 are essential downstream targets of the early RANKL signaling pathway. The recruitment of adaptor proteins also leads to the activation of MAPKs such as c-Jun N-terminal kinase (JNK), p38, and extracellular signal-regulated kinase, and Akt/PKB (David et al., 2002; He et al., 2011; Li et al., 2002; Wong et al., 1999). We recently proposed that the receptor for activated C kinase 1 (RACK1), a member of the Trp-Asp40 (WD40) repeat protein family, is necessary for the differentiation of osteoclasts (Lin et al., 2015). By acting as a scaffold protein to link the TRAF6-TAK1 complex with MKK6 but not with MKK3, RACK1 selectively facilitates the activation of p38 during the RANKL-initiated differentiation of osteoclast precursor cells into osteoclasts. In addition, considering the expression pattern of RACK1 in response to RANKL and its associations with the β1 and β2 integrin subunits and c-Src, RACK1 may participate in the functional stage where the resorption activity of osteoclasts is required. Other studies indicate that the activation of Akt, JNK, and NF-κB pathways requires the phosphorylation of growth factor receptor-bound protein 2 (Grb2)-associated binder-2 (Gab2) and its subsequent recruitment to RANK (Wada et al., 2005). Moreover, phospholipase Cγ2 (PLCγ2) forms a complex with Gab2, which is required for Gab2 phosphorylation, and modulates the recruitment of Gab2 to RANK (Mao et al., 2006). Lee et al. (2005) reported that the reactive oxygen species produced by the RANK-TRAF6-Rac1-NADPH oxidase 1-dependent pathway are also required for MAPK activation and osteoclastogenesis.
Fig. 1. Initiation of RANK signaling is mediated by TRAF6.
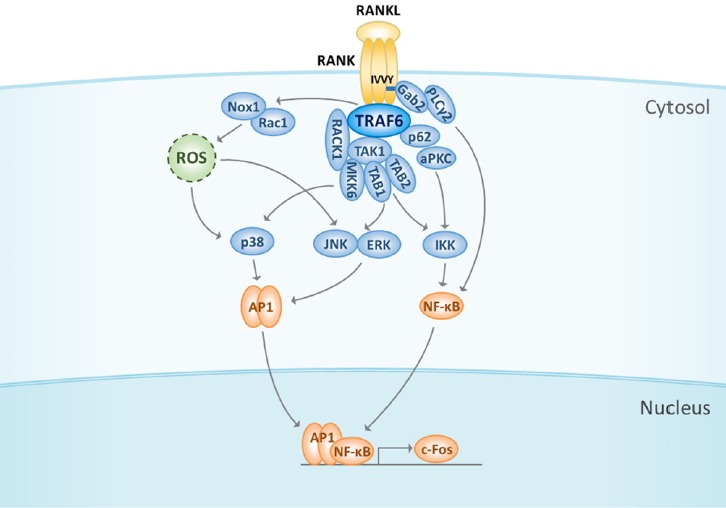
RANK stimulation by RANKL binding induces the recruitment and activation of a major adaptor protein, TRAF6. TRAF6 activates NF-κB either by interacting with p62 and aPKC or via TAK1 phosphorylation to regulate the IKK complex. Gab2 and PLCγ2 are other molecules that are required for NF-κB activation; they are recruited to RANK and activated. In addition, TRAF6 complexes with TAK1-TABs or TAK1-RACK1-MKK6 to facilitate the activation of MAPKs such as p38, JNK, and ERK. ROS produced by the RANK-TRAF6-Rac1-Nox1 cascades regulate MAPK activation. Activation of NF-κB and MAPKs leads to the induction of c-Fos at the initial stage of RANK signaling. RANK, receptor activator of nuclear factor-κB; RANKL, receptor activator of nuclear factor-κB ligand; TRAF6, TNF receptor-associated factors 6; NF-κB, nuclear factor-κB; aPCK, atypical protein kinase C; IKK, IκB kinase; TAK1, TGFβ-activated kinase 1; Gab2, growth factor receptor-bound protein 2 (Grb2)-associated binder-2; PLCγ2, phospholipase Cγ2; TAB, TAK1-binding protein; RACK1, receptor for activated C kinase 1; MAPKs, mitogen-activated protein kinases; JNK, c-Jun N-terminal kinase; ERK, extracellular signal-regulated kinase; ROS, reactive oxygen species; NOX1, NADPH oxidase 1; AP-1, activator protein-1; NFATc1, nuclear factor of activated T-cells cytoplasmic 1.
COSTIMULATORY SIGNALING
After the initial induction of NFATc1 by NF-κB, c-Fos/AP-1, and NFATc2 (Asagiri et al., 2005; Yamashita et al., 2007), RANK signaling functions together with the activation of immunoglobulin-like receptor/immunoreceptor tyrosine-based activation motif (ITAM) signaling (Fig. 2), which leading to the robust amplification of NFATc1 and its translocation to the nucleus where it binds with DNA in combination with other transcription factors to upregulate the transcription of NFATc1 target genes (Koga et al., 2004). NFATc1 may be a master transcription factor in osteoclastogenesis (Takayanagi et al., 2002), and numerous in vivo studies have demonstrated the essential role of NFATc1 in osteoclast formation. Osteoclast-specific conditional NFATc1-knockout mice exhibit osteopetrosis and inhibit osteoclastogenesis in vivo and in vitro (Aliprantis et al., 2008). Moreover, NFATc1 regulates not only osteoblasts and bone formation but also the activation and development of diverse cells in other biological systems such as the immune, cardiovascular, and skeletal muscular systems (Hogan et al., 2003; Koga et al., 2005).
Fig. 2. Cooperation of RANK signaling with costimulatory receptors.
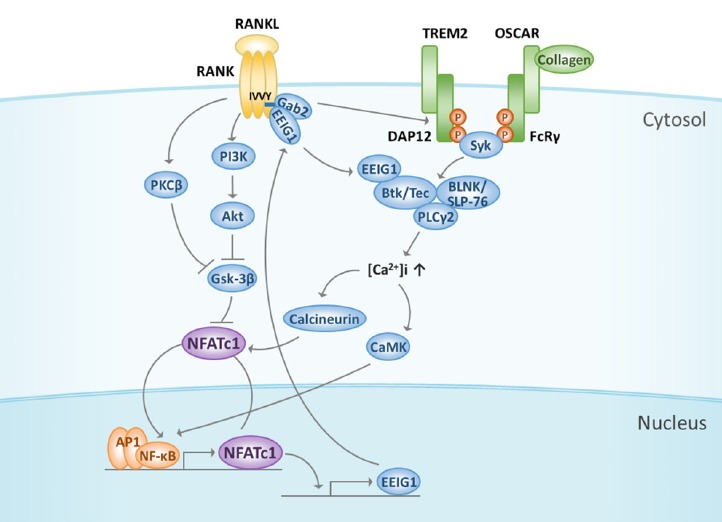
RANK signaling cooperates with immunoglobulin-like receptor/ITAM signals such as TREM-2/DAP12 and OSCAR/FcRγ, thereby leading to the amplification and translocation of NFATc1. When ITAM is tyrosine phosphorylated, Btk/Tec and BLNK/SLP-76 form a complex with PLCγ2 to activate PLCγ2 and Ca2+ signaling. EEIG1 induced by NFATc1 associates with Gab2 via the IVVY motif in RANK and then activates Btk/Tec followed by PLCγ2, suggesting that EEIG1 integrates RANK and ITAM signaling. Ca2+ oscillation induces calmodulin and calcineurin activation to regulate the nuclear translocation and amplification of NFATc1. The subcellular localization of NFATc1 is influenced by the phosphorylation of serine residues regulated by Gsk-3β, which is inhibited by either PI3K-Akt signaling or PKCβ. ITAM, immunoreceptor tyrosine-based activation motif; TREM-2, triggering receptor expressed in myeloid cells-2; DAP12, DNAX-activation protein 12; OSCAR, osteoclast-associated receptor; FcRγ, Fc receptor common γ subunit; Btk, Bruton’s tyrosine kinase; BLNK, B cell linker protein; SLP-76, Src homology 2 domain-containing leukocyte protein of 76 kD; EEIG1, early estrogen-induced gene 1; Gsk-3β, Glycogen synthase kinase-3β; PI3K, phosphoinositide 3-kinase; PKCβ, protein kinase Cβ.
In addition to RANK signaling, immunoglobulin-like receptors such as triggering receptor expressed in myeloid cells-2 (TREM-2) and osteoclast-associated receptor (OSCAR) transduce NFATc1 induction signals (Cella et al., 2003; Humphrey et al., 2006; Kim et al., 2002). These immune receptors have short cytoplasmic tails and are associated with adaptor proteins such as DNAX-activation protein 12 (DAP12) or the Fc receptor common γ subunit (FcRγ), which contains ITAM (Koga et al., 2004). When ITAM is tyrosine phosphorylated, possibly induced by RANKL stimulation, a complex is formed and contains the tyrosine kinases such as Bruton’s tyrosine kinase (Btk)/Tec, adaptor molecules B cell linker protein (BLNK), and Src homology 2 domain-containing leukocyte protein of 76 kDa (SLP-76), and this complex may integrate both RANK and ITAM downstream signaling, eventually leading to the activation of PLCγ2 by phosphorylation (Shinohara et al., 2008). Mice that are deficient in Btk and Tec develop an osteopetrotic phenotype because of severely impaired osteoclast differentiation, and the RANKL-induced tyrosine phosphorylation of PLCγ2 is highly suppressed (Shinohara et al., 2008). Thus, the RANKL-induced Ca2+ oscillation required for NFATc1 induction is abolished in cells that are defective in both Btk and Tec (Shinohara et al., 2008). This suggests that Btk and Tec are essential for osteoclastogenesis and that these kinases regulate NFATc1 activation via PLCγ2 and the Ca2+ signaling pathway. Furthermore, the formation of the complex comprising Btk/Tec and BNLK/SLP-76 cannot be detected in cells that are deficient in both DAP12 and FcRγ, suggesting that Tec kinases and BLNK both need to be activated by ITAM signaling to form this signaling complex, which may function as the molecular switch that links RANK and ITAM signaling (Shinohara et al., 2008). A recent study shows that early estrogen-induced gene 1 (EEIG1), which is induced by RANKL stimulation during osteoclast formation, physically interacts with RANK and associates with Gab2, PLCγ2, and Btk/Tec kinases after RANKL stimulation (Choi et al., 2013). EEIG1 knockdown leads to a marked reduction in RANK-induced osteoclast formation, which is caused by the impairment of RANKL-mediated PLCγ2 activation and NFATc1 induction. In response to RANKL stimulation, RANK forms a signaling complex that comprises EEIG1, Gab2, PLCγ2, and Btk/Tec, suggesting that EEIG1 is an adaptor protein and a linker protein that integrates RANK and ITAM signaling (Choi et al., 2013).
PLCγ2 also interacts with RANK via its highly conserved domain in the hyper-conserved region of RANK to form a complex with Gab2 and TRAF6, which results in its phosphorylation in a signal-dependent manner (Taguchi et al., 2009). Along with RANK signaling, ITAM signaling leads to the long-term induction of PLCγ2 activation. Activated PLCγ2 produces inositol-1,4,5-triphosphate (IP3) by digesting phosphatidylinositol-4,5-bisphosphate, and IP3 then binds to IP3 receptors on the surface of the endoplasmic reticulum (ER). Thus, the Ca2+ stored in ER is released into the cytoplasm, and Ca2+ oscillation is induced (Mao et al., 2006; Putney, 2005). The robust induction of NFATc1 is also regulated by its subcellular localization, which depends on the phosphorylation of its serine residues (Jang et al., 2011; Sheridan et al., 2002). In the absence of stimulation by RANKL, NFATc1 is heavily phosphorylated by active glycogen synthase kinase-3β (GSK-3β) and is localized in the cytoplasm. After RANK signaling induces GSK-3β phosphorylation at its inhibitory serine residue (Ser9 of GSK-3β), GSK-3β is inactivated. This enables NFATc1 dephosphorylation via calcineurin, followed by its translocation into the nucleus. In the nucleus, NFATc1 induces the expression of target genes. Transgenic mice, in which osteoclast specifically overexpress a constitutive active mutant of GSK-3β, exhibit osteopetrosis owing to marked reductions in the number of osteoclasts in vivo, as well as impaired osteoclast formation and NFATc1 induction in vitro (Jang et al., 2011). Furthermore, the inhibition of phosphoinositide 3-kinase reduces osteoclast formation and attenuates NFATc1 expression. Akt overexpression in osteoclast precursors strongly induces NFATc1 expression and its nuclear localization owing to the inactivation of GSK-3β via its increased phosphorylation (Moon et al., 2012). A recent study demonstrated that PKCβ controls the activity of NFATc1 by inactivating GSK-3β, thereby resulting in osteoclastogenesis (Shin et al., 2014). These results suggest that the GSK-3β/NFATc1 signaling cascade plays important roles in RANK signaling during osteoclast differentiation.
LATE STAGE OF SIGNALING IN OSTEOCLAST DIFFERENTIATION
In the late stage of RANK signaling, amplified NFATc1 activates and induces osteoclast-specific genes that encode proteins related to osteoclast differentiation, fusion, and function. Moreover, positive or negative regulators of NFATc1 are enhanced or suppressed during osteoclast differentiation (Fig. 3). The expression of negative regulators is downregulated during osteoclast differentiation in response to RANKL: the v-maf musculoaponeurotic fibrosarcoma oncogene family, protein B (MafB), interferon regulatory factor-8 (IRF-8), and B cell lymphoma 6 (Bcl6), which repress NFATc1 expression during osteoclast differentiation (Kim et al., 2007; Miyauchi et al., 2010; Zhao et al., 2009). All three of these negative factors are downregulated by the transcription repressor B-lymphocyte-induced maturation protein 1 (Blimp1) (Nishikawa et al., 2010). Blimp1 is induced by RANKL stimulation during osteoclast differentiation, and mice that lack Blimp1 in their osteoclasts exhibit osteopetrotic phenotypes owing to osteoclastogenesis aggravation (Miyauchi et al., 2010). Some evidence has recently suggested that sirtuin 6 (Sirt6), a nuclear histone deacetylase, functions as a co-repressor to directly inhibit anti-osteoclastogenic gene expression via Blimp1 (Park et al., 2016). Specific ablation of Sirt6 in hematopoietic cells increases the bone volume by decreasing the differentiation of osteoclasts. The expression of Sirt6 is upregulated by RANKL-induced NFATc1, and the induced Sirt6 positively regulates NFATc1 transcription by repressing MafB in combination with Blimp1 (Park et al., 2016). Therefore, NFATc1 maintains its own expression by inducing its target genes, Blimp1 and Sirt6, and they downregulate the expression of negative regulators.
Fig. 3. Late stage of RANK signaling.
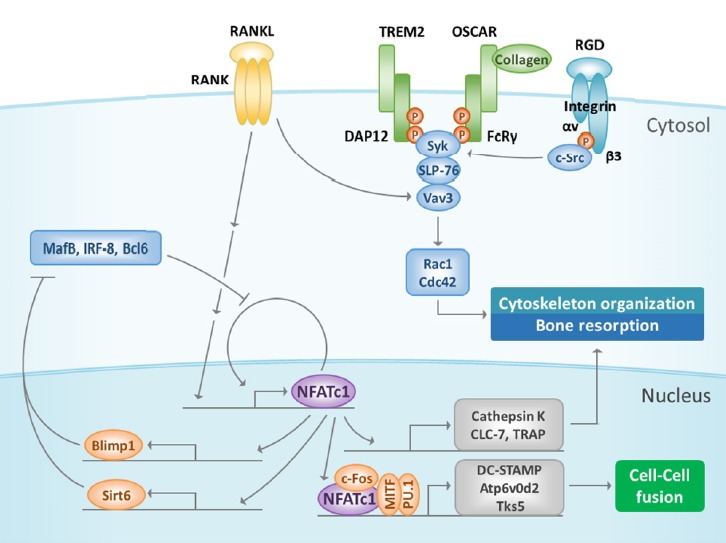
Amplified NFATc1 induces its target genes to regulate osteoclast differentiation, cell fusion, and function. NFATc1 represses negative regulators such as MafB, IRF-8, and Bcl6 during osteoclast differentiation by inducing Blimp1 and Sirt6, which act as transcription repressors. In addition, NFATc1 cooperates with other transcription factors such as MITF, c-Fos, and PU.1 to regulate osteoclast fusion by inducing DC-STAMP, Atp6v0d2, and Tks5. The αVβ3 integrin signal activates c-Src by binding to vitronectin. Next, c-Src phosphorylates Syk, which recruits DAP12 and SLP-76 to form a complex with Vav3. This complex activates Rac1 or Cdc42 and regulates the cytoskeleton organization that is important for regulating bone resorption. MafB, v-maf musculoaponeurotic fibrosarcoma oncogene family, protein B; IRF-8, interferon regulatory factor-8; Bcl6, B cell lymphoma 6; Blimp1, B-lymphocyte-induced maturation protein 1; Sirt6, sirtuin 6; MITF, microphthalmia-associated transcription factor; DC-STAMP, dendritic-cell-specific transmembrane protein; TRAP, tartrate-resistant acid phosphatase; Cdc42, cell division cycle 42.
Several proteins such as dendritic cell-specific transmembrane protein (DC-STAMP), vacuolar proton pump subunit Atp6v0d2, and c-Src substrate Tks5 are involved in osteoclast cell-cell fusion; their expression is regulated by NFATc1 together with PU.1, microphthalmia-associated transcription factor, and c-Fos (Kim et al., 2008; Oikawa et al., 2012; Yagi et al., 2007). DC-STAMP-deficient mice exhibit moderate osteopetrosis, which is distinguished by numerous tartrate-resistant acid phosphatase (TRAP)-positive mononuclear osteoclasts that can, although inefficiently, resorb bone (Yagi et al., 2007). Similarly, mice that lack Atp6v0d2 also exhibit mild osteopetrosis owing to the absence of multinuclear TRAP-positive osteoclasts; however, such mice mainly possess mononuclear TRAP-positive cells (Kim et al., 2008). αVβ3, the integrin vitronectin receptor, binds to vitronectin in the bone matrix, leading to bone resorption via the recruitment of c-Src tyrosine kinase (Chambers and Fuller, 2011; Zou et al., 2007). Next, c-Src phosphorylates Syk, which recruits DAP12 and SLP-76, which form an adaptor protein complex that activates the small Rho family GTPases, including Rac (Faccio et al., 2005). These interactions lead to the formation of the ruffled border membrane via the fusion of lysosomal secretory vesicles with the cytoplasmic membrane (Zou et al., 2007). To resorb bone, H+ ions are pumped out through the ruffled border and form HCl with Cl−, which demineralizes the bone matrix, and various hydrolases such as cathepsin K, CLC-7 chloride channel, and TRAP, which are NFATc1 target genes, are activated (Matsumoto et al., 2004; Takayanagi et al., 2002; Vaananen et al., 1990).
Kim et al. (2009) suggested that the cytoplasmic motif of RANK is specifically involved in osteoclast formation and function by regulating the actin cytoskeleton and survival of osteoclasts. They developed a cell-permeable RANK receptor inhibitor (RRI) to target the cytoplasmic motif of RANK. Treatment with the RRI peptide blocked RANKL-induced osteoclast formation and impaired the downstream RANK signaling via Vav3, Rac1, and cell division cycle 42, thereby disrupting the actin cytoskeleton in differentiated osteoclasts. Furthermore, RRI inhibited the bone-resorbing function of osteoclasts and enhanced osteoclast apoptosis. The RRI peptide did not abrogate any known protein kinase cascades mediated by TRAF6, and the induction of NFATc1 and osteoclast-specific genes were not changed by the peptide (Kim et al., 2009). These results suggest that the peptide targets a later stage of osteoclast formation after the NFATc1 action and the highly specific regulation of osteoclast formation, without affecting other signaling molecules that participate in various signaling pathways such as immune signaling.
CONCLUSION
The most important discovery in our understanding of osteoclasts is that RANKL is the key molecule that regulates the differentiation and function of osteoclasts. This observation allows the generation of virtually pure populations of osteoclasts, in vitro culture, thus facilitating the dissection of intracellular mechanisms and signaling networks that are involved in the cell’s differentiation and function. The signal is initiated by RANKL binding to RANK and is transduced via the recruitment of numerous adaptor proteins such as TRAF6 and kinases to the cytoplasmic tail of trimerized RANK, thereby forming a number of signaling complexes that facilitate the regulation of signal transduction. RANK signaling cooperates with the signal from the immune receptor/ITAM, which is another signaling pathway that is required for osteoclastogenesis. Together with ITAM-mediated Ca2+ signaling, RANK signaling finally induces the amplification of NFATc1 and the expression of osteoclastic genes. Many of the molecules that are involved in RANK signaling are also components of other signaling pathways such as the immune system; thus, the manner in which common molecules are regulated to ensure the specific signal transduction required for osteoclast formation remains unclear. Therefore, we suggest that various adaptor proteins, either novel for osteoclasts or already well-known in other systems, mediate the specificity of RANK signaling for osteoclast formation and function. Although targeting RANKL appears to be the most efficient and relevant approach for treating bone loss, other strategies might be required to minimize potential adverse effects on other organs such as the immune system. In addition, elucidating RANK signaling pathways that are specific to osteoclasts might identify potential new drug targets for preventing osteoporosis and other bone-related diseases.
ACKNOWLEDGMENTS
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea Government (MSIP) (No. 2016R1A2B3010699) and by the Soonchunhyang University Research Fund (to NKL).
REFERENCES
- Aliprantis A.O., Ueki Y., Sulyanto R., Park A., Sigrist K.S., Sharma S.M., Ostrowski M.C., Olsen B.R., Glimcher L.H. NFATc1 in mice represses osteoprotegerin during osteoclastogenesis and dissociates systemic osteopenia from inflammation in cherubism. J Clin Invest. 2008;118:3775–3789. doi: 10.1172/JCI35711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Asagiri M., Takayanagi H. The molecular understanding of osteoclast differentiation. Bone. 2007;40:251–264. doi: 10.1016/j.bone.2006.09.023. [DOI] [PubMed] [Google Scholar]
- Asagiri M., Sato K., Usami T., Ochi S., Nishina H., Yoshida H., Morita I., Wagner E.F., Mak T.W., Serfling E., et al. Autoamplification of NFATc1 expression determines its essential role in bone homeostasis. J Exp Med. 2005;202:1261–1269. doi: 10.1084/jem.20051150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boyle W.J., Simonet W.S., Lacey D.L. Osteoclast differentiation and activation. Nature. 2003;423:337–342. doi: 10.1038/nature01658. [DOI] [PubMed] [Google Scholar]
- Cella M., Buonsanti C., Strader C., Kondo T., Salmaggi A., Colonna M. Impaired differentiation of osteoclasts in TREM-2-deficient individuals. J Exp Med. 2003;198:645–651. doi: 10.1084/jem.20022220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chambers T.J., Fuller K. How are osteoclasts induced to resorb bone? Ann. N Y Acad Sci. 2011;1240:1–6. doi: 10.1111/j.1749-6632.2011.06249.x. [DOI] [PubMed] [Google Scholar]
- Cheng X., Kinosaki M., Murali R., Greene M.I. The TNF receptor superfamily: role in immune inflammation and bone formation. Immunol Res. 2003;27:287–294. doi: 10.1385/IR:27:2-3:287. [DOI] [PubMed] [Google Scholar]
- Choi H.K., Kang H.R., Jung E., Kim T.E., Lin J.J., Lee S.Y. Early estrogen-induced gene 1, a novel RANK signaling component, is essential for osteoclastogenesis. Cell Res. 2013;23:524–536. doi: 10.1038/cr.2013.33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darnay B.G., Haridas V., Ni J., Moore P.A., Aggarwal B.B. Characterization of the intracellular domain of receptor activator of NF-kappaB (RANK). Interaction with tumor necrosis factor receptor-associated factors and activation of NF-kappab and c-Jun N-terminal kinase. J Biol Chem. 1998;273:20551–20555. doi: 10.1074/jbc.273.32.20551. [DOI] [PubMed] [Google Scholar]
- David J.P., Sabapathy K., Hoffmann O., Idarraga M.H., Wagner E.F. JNK1 modulates osteoclastogenesis through both c-Jun phosphorylation-dependent and-independent mechanisms. J Cell Sci. 2002;115:4317–4325. doi: 10.1242/jcs.00082. [DOI] [PubMed] [Google Scholar]
- Dougall W.C., Glaccum M., Charrier K., Rohrbach K., Brasel K., De Smedt T., Daro E., Smith J., Tometsko M.E., Maliszewski C.R., et al. RANK is essential for osteoclast and lymph node development. Genes Dev. 1999;13:2412–2424. doi: 10.1101/gad.13.18.2412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duran A., Serrano M., Leitges M., Flores J.M., Picard S., Brown J.P., Moscat J., Diaz-Meco M.T. The atypical PKC-interacting protein p62 is an important mediator of RANK-activated osteoclastogenesis. Dev Cell. 2004;6:303–309. doi: 10.1016/s1534-5807(03)00403-9. [DOI] [PubMed] [Google Scholar]
- Faccio R., Teitelbaum S.L., Fujikawa K., Chappel J., Zallone A., Tybulewicz V.L., Ross F.P., Swat W. Vav3 regulates osteoclast function and bone mass. Nat Med. 2005;11:284–290. doi: 10.1038/nm1194. [DOI] [PubMed] [Google Scholar]
- Franzoso G., Carlson L., Xing L., Poljak L., Shores E.W., Brown K.D., Leonardi A., Tran T., Boyce B.F., Siebenlist U. Requirement for NF-kappaB in osteoclast and B-cell development. Genes Dev. 1997;11:3482–3496. doi: 10.1101/gad.11.24.3482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fumoto T., Takeshita S., Ito M., Ikeda K. Physiological functions of osteoblast lineage and T cell-derived RANKL in bone homeostasis. J Bone Miner Res. 2014;29:830–842. doi: 10.1002/jbmr.2096. [DOI] [PubMed] [Google Scholar]
- Ghosh S., Karin M. Missing pieces in the NF-kappaB puzzle. Cell. 2002;109(Suppl):S81–96. doi: 10.1016/s0092-8674(02)00703-1. [DOI] [PubMed] [Google Scholar]
- Grigoriadis A.E., Wang Z.Q., Cecchini M.G., Hofstetter W., Felix R., Fleisch H.A., Wagner E.F. c-Fos: a key regulator of osteoclast-macrophage lineage determination and bone remodeling. Science. 1994;266:443–448. doi: 10.1126/science.7939685. [DOI] [PubMed] [Google Scholar]
- He Y., Staser K., Rhodes S.D., Liu Y., Wu X., Park S.J., Yuan J., Yang X., Li X., Jiang L., et al. Erk1 positively regulates osteoclast differentiation and bone resorptive activity. PLoS One. 2011;6:e24780. doi: 10.1371/journal.pone.0024780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hogan P.G., Chen L., Nardone J., Rao A. Transcriptional regulation by calcium, calcineurin, and NFAT. Genes Dev. 2003;17:2205–2232. doi: 10.1101/gad.1102703. [DOI] [PubMed] [Google Scholar]
- Humphrey M.B., Daws M.R., Spusta S.C., Niemi E.C., Torchia J.A., Lanier L.L., Seaman W.E., Nakamura M.C. TREM2, a DAP12-associated receptor, regulates osteoclast differentiation and function. J Bone Miner Res. 2006;21:237–245. doi: 10.1359/JBMR.051016. [DOI] [PubMed] [Google Scholar]
- Ikeda F., Nishimura R., Matsubara T., Tanaka S., Inoue J., Reddy S.V., Hata K., Yamashita K., Hiraga T., Watanabe T., et al. Critical roles of c-Jun signaling in regulation of NFAT family and RANKL-regulated osteoclast differentiation. J Clin Invest. 2004;114:475–484. doi: 10.1172/JCI19657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iotsova V., Caamano J., Loy J., Yang Y., Lewin A., Bravo R. Osteopetrosis in mice lacking NF-kappaB1 and NF-kappaB2. Nat Med. 1997;3:1285–1289. doi: 10.1038/nm1197-1285. [DOI] [PubMed] [Google Scholar]
- Jang H.D., Shin J.H., Park D.R., Hong J.H., Yoon K., Ko R., Ko C.Y., Kim H.S., Jeong D., Kim N., et al. Inactivation of glycogen synthase kinase-3beta is required for osteoclast differentiation. J Biol Chem. 2011;286:39043–39050. doi: 10.1074/jbc.M111.256768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karsenty G., Wagner E.F. Reaching a genetic and molecular understanding of skeletal development. Dev Cell. 2002;2:389–406. doi: 10.1016/s1534-5807(02)00157-0. [DOI] [PubMed] [Google Scholar]
- Kim N., Takami M., Rho J., Josien R., Choi Y. A novel member of the leukocyte receptor complex regulates osteoclast differentiation. J Exp Med. 2002;195:201–209. doi: 10.1084/jem.20011681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim K., Kim J.H., Lee J., Jin H.M., Kook H., Kim K.K., Lee S.Y., Kim N. MafB negatively regulates RANKL-mediated osteoclast differentiation. Blood. 2007;109:3253–3259. doi: 10.1182/blood-2006-09-048249. [DOI] [PubMed] [Google Scholar]
- Kim K., Lee S.H., Ha Kim J., Choi Y., Kim N. NFATc1 induces osteoclast fusion via up-regulation of Atp6v0d2 and the dendritic cell-specific transmembrane protein (DC-STAMP) Mol Endocrinol. 2008;22:176–185. doi: 10.1210/me.2007-0237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim H., Choi H.K., Shin J.H., Kim K.H., Huh J.Y., Lee S.A., Ko C.Y., Kim H.S., Shin H.I., Lee H.J., et al. Selective inhibition of RANK blocks osteoclast maturation and function and prevents bone loss in mice. J Clin Invest. 2009;119:813–825. doi: 10.1172/JCI36809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kobayashi N., Kadono Y., Naito A., Matsumoto K., Yamamoto T., Tanaka S., Inoue J. Segregation of TRAF6-mediated signaling pathways clarifies its role in osteoclastogenesis. EMBO J. 2001;20:1271–1280. doi: 10.1093/emboj/20.6.1271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koga T., Inui M., Inoue K., Kim S., Suematsu A., Kobayashi E., Iwata T., Ohnishi H., Matozaki T., Kodama T., et al. Costimulatory signals mediated by the ITAM motif cooperate with RANKL for bone homeostasis. Nature. 2004;428:758–763. doi: 10.1038/nature02444. [DOI] [PubMed] [Google Scholar]
- Koga T., Matsui Y., Asagiri M., Kodama T., de Crombrugghe B., Nakashima K., Takayanagi H. NFAT and Osterix cooperatively regulate bone formation. Nat Med. 2005;11:880–885. doi: 10.1038/nm1270. [DOI] [PubMed] [Google Scholar]
- Kong Y.Y., Feige U., Sarosi I., Bolon B., Tafuri A., Morony S., Capparelli C., Li J., Elliott R., McCabe S., et al. Activated T cells regulate bone loss and joint destruction in adjuvant arthritis through osteoprotegerin ligand. Nature. 1999;402:304–309. doi: 10.1038/46303. [DOI] [PubMed] [Google Scholar]
- Lee N.K., Choi Y.G., Baik J.Y., Han S.Y., Jeong D.W., Bae Y.S., Kim N., Lee S.Y. A crucial role for reactive oxygen species in RANKL-induced osteoclast differentiation. Blood. 2005;106:852–859. doi: 10.1182/blood-2004-09-3662. [DOI] [PubMed] [Google Scholar]
- Li J., Sarosi I., Yan X.Q., Morony S., Capparelli C., Tan H.L., McCabe S., Elliott R., Scully S., Van G., et al. RANK is the intrinsic hematopoietic cell surface receptor that controls osteoclastogenesis and regulation of bone mass and calcium metabolism. Proc Natl Acad Sci USA. 2000;97:1566–1571. doi: 10.1073/pnas.97.4.1566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li X., Udagawa N., Itoh K., Suda K., Murase Y., Nishihara T., Suda T., Takahashi N. p38 MAPK-mediated signals are required for inducing osteoclast differentiation but not for osteoclast function. Endocrinology. 2002;143:3105–3113. doi: 10.1210/endo.143.8.8954. [DOI] [PubMed] [Google Scholar]
- Lin J., Lee D., Choi Y., Lee S.Y. The scaffold protein RACK1 mediates the RANKL-dependent activation of p38 MAPK in osteoclast precursors. Sci Signal. 2015;8:ra54. doi: 10.1126/scisignal.2005867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lomaga M.A., Yeh W.C., Sarosi I., Duncan G.S., Furlonger C., Ho A., Morony S., Capparelli C., Van G., Kaufman S., et al. TRAF6 deficiency results in osteopetrosis and defective interleukin-1, CD40, and LPS signaling. Genes Dev. 1999;13:1015–1024. doi: 10.1101/gad.13.8.1015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mao D., Epple H., Uthgenannt B., Novack D.V., Faccio R. PLCgamma2 regulates osteoclastogenesis via its interaction with ITAM proteins and GAB2. J Clin Invest. 2006;116:2869–2879. doi: 10.1172/JCI28775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsumoto M., Kogawa M., Wada S., Takayanagi H., Tsujimoto M., Katayama S., Hisatake K., Nogi Y. Essential role of p38 mitogen-activated protein kinase in cathepsin K gene expression during osteoclastogenesis through association of NFATc1 and PU.1. J Biol Chem. 2004;279:45969–45979. doi: 10.1074/jbc.M408795200. [DOI] [PubMed] [Google Scholar]
- Miyauchi Y., Ninomiya K., Miyamoto H., Sakamoto A., Iwasaki R., Hoshi H., Miyamoto K., Hao W., Yoshida S., Morioka H., et al. The Blimp1-Bcl6 axis is critical to regulate osteoclast differentiation and bone homeostasis. J Exp Med. 2010;207:751–762. doi: 10.1084/jem.20091957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mizukami J., Takaesu G., Akatsuka H., Sakurai H., Ninomiya-Tsuji J., Matsumoto K., Sakurai N. Receptor activator of NF-kappaB ligand (RANKL) activates TAK1 mitogen-activated protein kinase kinase kinase through a signaling complex containing RANK, TAB2, and TRAF6. Mol Cell Biol. 2002;22:992–1000. doi: 10.1128/MCB.22.4.992-1000.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moon J.B., Kim J.H., Kim K., Youn B.U., Ko A., Lee S.Y., Kim N. Akt induces osteoclast differentiation through regulating the GSK3beta/NFATc1 signaling cascade. J Immunol. 2012;188:163–169. doi: 10.4049/jimmunol.1101254. [DOI] [PubMed] [Google Scholar]
- Naito A., Azuma S., Tanaka S., Miyazaki T., Takaki S., Takatsu K., Nakao K., Nakamura K., Katsuki M., Yamamoto T., et al. Severe osteopetrosis, defective interleukin-1 signalling and lymph node organogenesis in TRAF6-deficient mice. Genes Cells. 1999;4:353–362. doi: 10.1046/j.1365-2443.1999.00265.x. [DOI] [PubMed] [Google Scholar]
- Nakashima T., Hayashi M., Fukunaga T., Kurata K., Oh-Hora M., Feng J.Q., Bonewald L.F., Kodama T., Wutz A., Wagner E.F., et al. Evidence for osteocyte regulation of bone homeostasis through RANKL expression. Nat Med. 2011;17:1231–1234. doi: 10.1038/nm.2452. [DOI] [PubMed] [Google Scholar]
- Ninomiya-Tsuji J., Kishimoto K., Hiyama A., Inoue J., Cao Z., Matsumoto K. The kinase TAK1 can activate the NIK-I kappaB as well as the MAP kinase cascade in the IL-1 signalling pathway. Nature. 1999;398:252–256. doi: 10.1038/18465. [DOI] [PubMed] [Google Scholar]
- Nishikawa K., Nakashima T., Hayashi M., Fukunaga T., Kato S., Kodama T., Takahashi S., Calame K., Takayanagi H. Blimp1-mediated repression of negative regulators is required for osteoclast differentiation. Proc Natl Acad Sci USA. 2010;107:3117–3122. doi: 10.1073/pnas.0912779107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oikawa T., Oyama M., Kozuka-Hata H., Uehara S., Udagawa N., Saya H., Matsuo K. Tks5-dependent formation of circumferential podosomes/invadopodia mediates cell-cell fusion. J Cell Biol. 2012;197:553–568. doi: 10.1083/jcb.201111116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park S.J., Huh J.E., Shin J., Park D.R., Ko R., Jin G.R., Seo D.H., Kim H.S., Shin H.I., Oh G.T., et al. Sirt6 cooperates with Blimp1 to positively regulate osteoclast differentiation. Sci Rep. 2016;6:26186. doi: 10.1038/srep26186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Putney J.W., Jr Capacitative calcium entry: sensing the calcium stores. J Cell Biol. 2005;169:381–382. doi: 10.1083/jcb.200503161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sheridan C.M., Heist E.K., Beals C.R., Crabtree G.R., Gardner P. Protein kinase A negatively modulates the nuclear accumulation of NF-ATc1 by priming for subsequent phosphorylation by glycogen synthase kinase-3. J Biol Chem. 2002;277:48664–48676. doi: 10.1074/jbc.M207029200. [DOI] [PubMed] [Google Scholar]
- Shin J., Jang H., Lin J., Lee S.Y. PKCbeta positively regulates RANKL-induced osteoclastogenesis by inactivating GSK-3beta. Mol Cells. 2014;37:747–752. doi: 10.14348/molcells.2014.0220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shinohara M., Koga T., Okamoto K., Sakaguchi S., Arai K., Yasuda H., Takai T., Kodama T., Morio T., Geha R.S., et al. Tyrosine kinases Btk and Tec regulate osteoclast differentiation by linking RANK and ITAM signals. Cell. 2008;132:794–806. doi: 10.1016/j.cell.2007.12.037. [DOI] [PubMed] [Google Scholar]
- Taguchi Y., Gohda J., Koga T., Takayanagi H., Inoue J. A unique domain in RANK is required for Gab2 and PLCgamma2 binding to establish osteoclastogenic signals. Genes Cells. 2009;14:1331–1345. doi: 10.1111/j.1365-2443.2009.01351.x. [DOI] [PubMed] [Google Scholar]
- Takayanagi H., Kim S., Koga T., Nishina H., Isshiki M., Yoshida H., Saiura A., Isobe M., Yokochi T., Inoue J., et al. Induction and activation of the transcription factor NFATc1 (NFAT2) integrate RANKL signaling in terminal differentiation of osteoclasts. Dev Cell. 2002;3:889–901. doi: 10.1016/s1534-5807(02)00369-6. [DOI] [PubMed] [Google Scholar]
- Teitelbaum S.L. Bone resorption by osteoclasts. Science. 2000;289:1504–1508. doi: 10.1126/science.289.5484.1504. [DOI] [PubMed] [Google Scholar]
- Teitelbaum S.L., Ross F.P. Genetic regulation of osteoclast development and function. Nat Rev Genet. 2003;4:638–649. doi: 10.1038/nrg1122. [DOI] [PubMed] [Google Scholar]
- Vaananen H.K., Karhukorpi E.K., Sundquist K., Wallmark B., Roininen I., Hentunen T., Tuukkanen J., Lakkakorpi P. Evidence for the presence of a proton pump of the vacuolar H(+)-ATPase type in the ruffled borders of osteoclasts. J Cell Biol. 1990;111:1305–1311. doi: 10.1083/jcb.111.3.1305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wada T., Nakashima T., Oliveira-dos-Santos A.J., Gasser J., Hara H., Schett G., Penninger J.M. The molecular scaffold Gab2 is a crucial component of RANK signaling and osteoclastogenesis. Nat Med. 2005;11:394–399. doi: 10.1038/nm1203. [DOI] [PubMed] [Google Scholar]
- Wagner E.F. Functions of AP1 (Fos/Jun) in bone development. Ann Rheum Dis. 2002;61(Suppl 2):ii40–42. doi: 10.1136/ard.61.suppl_2.ii40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Z.Q., Ovitt C., Grigoriadis A.E., Mohle-Steinlein U., Ruther U., Wagner E.F. Bone and haematopoietic defects in mice lacking c-fos. Nature. 1992;360:741–745. doi: 10.1038/360741a0. [DOI] [PubMed] [Google Scholar]
- Wong B.R., Josien R., Lee S.Y., Vologodskaia M., Steinman R.M., Choi Y. The TRAF family of signal transducers mediates NF-kappaB activation by the TRANCE receptor. J Biol Chem. 1998;273:28355–28359. doi: 10.1074/jbc.273.43.28355. [DOI] [PubMed] [Google Scholar]
- Wong B.R., Besser D., Kim N., Arron J.R., Vologodskaia M., Hanafusa H., Choi Y. TRANCE, a TNF family member, activates Akt/PKB through a signaling complex involving TRAF6 and c-Src. Mol Cell. 1999;4:1041–1049. doi: 10.1016/s1097-2765(00)80232-4. [DOI] [PubMed] [Google Scholar]
- Xiong J., Onal M., Jilka R.L., Weinstein R.S., Manolagas S.C., O’Brien C.A. Matrix-embedded cells control osteoclast formation. Nat Med. 2011;17:1235–1241. doi: 10.1038/nm.2448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yagi M., Ninomiya K., Fujita N., Suzuki T., Iwasaki R., Morita K., Hosogane N., Matsuo K., Toyama Y., Suda T., et al. Induction of DC-STAMP by alternative activation and downstream signaling mechanisms. J Bone Miner Res. 2007;22:992–1001. doi: 10.1359/jbmr.070401. [DOI] [PubMed] [Google Scholar]
- Yamashita T., Yao Z., Li F., Zhang Q., Badell I.R., Schwarz E.M., Takeshita S., Wagner E.F., Noda M., Matsuo K., et al. NF-kappaB p50 and p52 regulate receptor activator of NF-kappaB ligand (RANKL) and tumor necrosis factor-induced osteoclast precursor differentiation by activating c-Fos and NFATc1. J Biol Chem. 2007;282:18245–18253. doi: 10.1074/jbc.M610701200. [DOI] [PubMed] [Google Scholar]
- Yasuda H., Shima N., Nakagawa N., Yamaguchi K., Kinosaki M., Mochizuki S., Tomoyasu A., Yano K., Goto M., Murakami A., et al. Osteoclast differentiation factor is a ligand for osteoprotegerin/osteoclastogenesis-inhibitory factor and is identical to TRANCE/RANKL. Proc Natl Acad Sci USA. 1998;95:3597–3602. doi: 10.1073/pnas.95.7.3597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ye H., Arron J.R., Lamothe B., Cirilli M., Kobayashi T., Shevde N.K., Segal D., Dzivenu O.K., Vologodskaia M., Yim M., et al. Distinct molecular mechanism for initiating TRAF6 signalling. Nature. 2002;418:443–447. doi: 10.1038/nature00888. [DOI] [PubMed] [Google Scholar]
- Zaidi M. Skeletal remodeling in health and disease. Nat Med. 2007;13:791–801. doi: 10.1038/nm1593. [DOI] [PubMed] [Google Scholar]
- Zhao B., Takami M., Yamada A., Wang X., Koga T., Hu X., Tamura T., Ozato K., Choi Y., Ivashkiv L.B., et al. Interferon regulatory factor-8 regulates bone metabolism by suppressing osteoclastogenesis. Nat Med. 2009;15:1066–1071. doi: 10.1038/nm.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zou W., Kitaura H., Reeve J., Long F., Tybulewicz V.L., Shattil S.J., Ginsberg M.H., Ross F.P., Teitelbaum S.L. Syk, c-Src, the alphavbeta3 integrin, and ITAM immunoreceptors, in concert, regulate osteoclastic bone resorption. J Cell Biol. 2007;176:877–888. doi: 10.1083/jcb.200611083. [DOI] [PMC free article] [PubMed] [Google Scholar]