

Abstract
Study of the probiotic potential of microorganisms isolated from fermented foods has been increasing, especially studies related to lactobacilli. In intestinal models, lactobacilli have demonstrated beneficial properties, such as anti-inflammatory activity and increased antibody production, but the molecular mechanisms involving probiotic and antagonistic action as well as their effect on human vaginal cells have not yet been fully elucidated. The aim of this study was to evaluate the functional and antagonistic properties of three strains of lactobacilli isolated from cocoa fermentation (Lactobacillus fermentum 5.2, L. plantarum 6.2, and L. plantarum 7.1) against Gardnerella vaginalis. Our results show that the lactobacilli have potential use as probiotics, since they have high hydrophobicity and autoaggregation properties and effectively adhere to vaginal cells. Metabolites secreted into the culture medium and whole cells of the strains under study are capable of interfering with the growth of G. vaginalis to different degrees. The elucidation of the antagonistic mechanisms as well as their effect on human cells may be useful in the development of a product containing such microorganisms or products secreted by them.
1. Introduction
Probiotics are microorganisms capable of conferring health benefits to the host after correct administration. Lactic acid bacteria (LAB) are an integral part of the intestinal and genital microbiota of humans and other vertebrates [1].
Probiotic can act in different ways: (1) competitively occupying receptors on mucosal epithelial cells [2]; (2) inhibiting the adhesion of pathogens [3]; (3) producing antimicrobial substances such as bacteriocins, hydrogen peroxide, and organic acids [4, 5]; (4) inhibiting the synthesis of toxins or degrading cytotoxic compounds [6]; and (5) modulating the immune response [7, 8].
Most of the probiotics available on the market have human origin, due to the concept that (it was expected) their action would be accentuated in organisms of the same species from which the strain was primarily isolated. However, new studies have shown that microorganisms of extraintestinal origin, isolated from plants and fermented foods, exhibit promising effects in the treatment and prevention of numerous diseases [9–11].
Cocoa is the main agricultural product in southern Bahia, and Brazil is one of the world's largest producers, along with Ghana and Côte d'Ivoire in Africa [12]. The fermentation of cocoa beans is a process in which LAB plays an important role, because these microorganisms contribute in the formation of the sensory characteristics of the final product, chocolate [13, 14].
Interest in searching for new strains with probiotic potential has risen in the industrial and scientific sectors mainly due to the market demand for functional foods and therapeutics with lesser side effects and because of the numerous benefits attributed to these microorganisms [9, 15]. The role of LAB in cocoa fermentation has not been fully clarified, but the diversity of bacteria involved in this process makes this process/product a promising source for isolation of the prospecting strains for biotechnology applications [16].
Preliminary studies of our group showed that LAB isolated from this fermentative process were able to reduce intestinal inflammation induced in an experimental model of colitis in rats, decreasing the concentration of proinflammatory cytokines in serum, increasing level of IgA, and restoring tissue structure of the mucosa [10, 17].
Bacterial vaginosis is a clinical condition of disturbance of the native microbiota with decreased Lactobacillus counts and increased pathogenic microorganisms such as Gardnerella vaginalis [18]. Several LAB isolated from vaginal microenvironment exhibit inhibitory activity against vaginosis-associated pathogens, such as Candida albicans [19], Neisseria gonorrhoeae, G. vaginalis [20], and Enterobacteriaceae [4]. However, there are no investigations of the use of LAB isolated from cocoa fermentation in bacterial vaginosis models.
Thus, the aim of the present study was to evaluate in vitro functional and antagonistic probiotic features of three Lactobacillus strains isolated from the cocoa fermentation process against G. vaginalis.
2. Materials and Methods
2.1. Strains, Cell Lines, and Growth Conditions
Three strains of lactobacilli previously isolated and characterized by our research group [17] were used in this study: Lactobacillus fermentum 5.2, Lactobacillus plantarum 6.2, and Lactobacillus plantarum 7.1.
Lactobacillus strains were grown in de Man, Rogosa, and Sharpe (MRS) medium (HiMedia) for 18–24 h at 37°C under microaerophilic conditions. Gardnerella vaginalis ATCC 49154 was grown on 5% blood agar plates (HiMedia) or Brain and Heart Infusion (BHI) broth (Difco) for 18–24 h at 37°C in a 5% CO2 atmosphere.
HMVII, a vaginal epithelial cell line (BCRJ 0316), was grown in RPMI 1640 medium (HyClone) supplemented with 10% fetal bovine serum (FBS) (Gibco) and 1% antibiotic (penicillin and streptomycin) (Gibco) at 37°C in a 5% CO2 atmosphere.
2.2. Lactobacilli Supernatant Preparation
Lactobacilli were grown in MRS broth for 48 h at 37°C. After incubation, the supernatants were obtained by harvesting of cells by centrifugation for 15 min at 8,000 ×g. pH of supernatants was measured before lyophilization. Lyophilized supernatants were kept under refrigeration conditions until use. Before use they were reconstituted in sterile ultrapure water and filtered through 0.22 μm membranes.
2.3. Autoaggregation and Coaggregation Assays
Autoaggregation and coaggregation assays were adapted from Kos et al. [21]. For the autoaggregation assay, strains of lactobacilli were grown in MRS broth for 18 h. After centrifugation (8,000 ×g, 10 min), pellets of cells were resuspended, washed twice with 0.9% saline, and finally resuspended to 1 × 108 CFU mL−1 in the same solution. Then, suspensions were vortexed and incubated at 37°C for 5 h. Each hour, an aliquot (1 mL) from the top of the suspensions was carefully removed and its absorbance read at 600 nm in a spectrophotometer. Autoaggregation was calculated using the following formula: autoaggregation (%) = ((A0 − At)/A0) × 100, where A0 indicates the absorbance at time 0 h and At indicates the absorbance every hour, up to 5 h.
For the coaggregation assay, a Lactobacillus suspension was prepared similar to the autoaggregation assay. A suspension of G. vaginalis cells after growth in BHI was made and finally standardized to 1 × 108 CFU mL−1 in 0.9% saline. One ml of each Lactobacillus suspension was mixed with the same volume of G. vaginalis cell suspension and the mixture was vortexed for 10 sec and left for gravity sedimentation. Control tubes containing 2 mL of each bacterial cell suspension alone were made. Absorbance of the suspensions was read at 600 nm in a spectrophotometer after 5 h of incubation at 37°C. Coaggregation was calculated using the following formula: coaggregation (%) = [(Ax + Ay)/2 − A(x + y)]/[(Ax + Ay)/2], where x and y indicate the absorbance of strains in the control tubes and (x + y) indicates the absorbance of the mixtures.
2.4. Microbial Hydrophobicity Assay
To determine the degree of hydrophobicity, we used microbial adhesion to hydrocarbons (MATH), adapted from Rodríguez et al. [22], using xylene as solvent. Lactobacilli strains were grown in MRS broth for 18 h. After centrifugation (8,000 ×g, 10 min), pellets were recovered, washed twice with 0.9% saline, and adjusted to an optical density (OD 600) of 0.7. The solvent (xylene; 1 mL) was then added to each bacterial suspension and the mixtures were vortexed vigorously for 2 min and incubated for 2 h at 37°C. The lower aqueous phase was carefully removed and read at 600 nm in a spectrophotometer. Hydrophobicity was calculated using the following formula: hydrophobicity (%) = ((A0 − A2)/A0) × 100, where A0 indicates the absorbance at time 0 h and A2 indicates the absorbance after 2 h.
2.5. Lactobacillus Adhesion to HMVII Cells
For the adhesion test, we used a methodology adapted from Santos et al. [7]. Vaginal epithelial cells (HMVII) were used at a concentration of 1 × 106 cells mL−1. Lactobacilli were grown in MRS broth for 18 h. After centrifugation (8,000 ×g, 10 min), pellets were recovered, washed twice with 0.9% saline, and adjusted to 1 × 108 CFU mL−1 in RPMI supplemented with 10% FBS. Lactobacilli cell suspension was added to wells containing HMVII cells (multiplicity of infection, MOI = 100) and incubated at 37°C in a 5% CO2 atmosphere. Medium was added to the wells containing HMVII cells as a negative control. After 2 h of interaction, the cell monolayer was washed three times with 0.9% saline and treated with 0.25% trypsin-EDTA for 5 min. The determination of adhered lactobacilli was performed by serial dilution followed by plating on MRS agar. Plates were incubated for 48 h at 37°C and the colony forming units (CFU mL−1) were counted. The percentage of adhered lactobacilli was calculated by the following formula: adhesion (%) = (CFUend/CFUinitial) × 100.
In addition, scanning electron microscopy (SEM) was performed to visualize lactobacilli adhered to the vaginal cells after interaction. HMVII cells (1 × 106 cells mL−1) were grown on glass coverslips with each one of the three strains of Lactobacillus tested in this study (1 × 108 UFC mL−1) and incubated for 2 h at 37°C in a 5% CO2 atmosphere. HMVII cells alone were used as control. Coverslips were washed three times with 0.9% saline and fixed in 2.5% glutaraldehyde in 0.1 M sodium cacodylate buffer pH 7.2. Next, dehydration was performed in series of increasing acetone concentrations (50–100%, 10 min each). The samples were subjected to critical point drying and metallized with an approximately 20 nm thick gold layer to be observed in the scanning electron microscope Quanta 250 (FEI Company).
2.6. Antimicrobial Activity of Lactobacillus Culture Supernatants
First, an evaluation of the antimicrobial activity was made through the agar diffusion technique. The strain of G. vaginalis was previously cultured for 18–24 h at 37°C in a 5% CO2 atmosphere onto blood agar plates. Afterwards, bacteria were harvested from the agar, washed with 0.9% saline, centrifuged, resuspended in the same solution, and adjusted in a spectrophotometer at the concentration of 1 × 108 CFU mL−1. The inoculum was spread over the surface of Petri dishes containing BHI agar (Difco); then wells were perforated in the agar in which culture supernatants of the different lactobacilli were added. Plates were incubated for 24 h at 37°C in a 5% CO2 atmosphere. After incubation, the presence or absence of inhibition halos around the wells was observed.
Microdilution technique was performed to determine minimum inhibitory concentration (MIC) in 96-well microtiter plates according to the recommendations of the Clinical and Laboratory Standards Institute, CLSI [23]. Serial dilutions were done starting from 40 mg mL−1 of the culture supernatants of the lactobacilli in Mueller-Hinton broth (MH) containing 5 × 105 CFU mL−1 of G. vaginalis. The same procedure was done with the following controls: lyophilized culture medium without lactobacilli (MRS control); MH without inoculum (control of sterility of the medium); MH containing 5 × 105 CFU mL−1 of G. vaginalis (positive control); and MH containing 5 × 105 CFU mL−1 of G. vaginalis and 12.5 μg mL−1 of chloramphenicol (negative control). The microtiter plates were incubated for 24 h at 37°C in a 5% CO2 atmosphere with inhibition being observed by the absence of turbidity in the wells. To confirm whether the supernatants had a bactericidal or bacteriostatic effect, the contents of the wells were plated onto blood agar and then incubated at 37°C in a 5% CO2 atmosphere for 24 h [24].
2.7. Coculture Assay
To evaluate the influence of lactobacilli on the growth of G. vaginalis, we used the bacterial coculture technique described by Coudeyras et al. [25]. The assay was performed in BHI medium supplemented with 1% yeast extract, 0.1% maltose, 0.1% glucose, and 10% fetal bovine serum. An inoculum of 1 × 108 CFU mL−1 was made for each microorganism. The strain of G. vaginalis was cultivated alone (control) or with each strain of the three lines of Lactobacillus, in a ratio of 1 : 1 at 37°C in a 5% CO2 atmosphere for 24 h. Aliquots were removed after 4, 8, and 24 h, serially diluted, and plated onto blood agar plates to determine the microbial count of G. vaginalis after interaction. Plates were also incubated at 37°C for 24 h in a 5% CO2 atmosphere.
2.8. Lactobacilli Susceptibility to Antibiotics
Susceptibility of Lactobacillus strains to antimicrobials was determined by the modified agar diffusion method of CLSI [26]. Lactobacilli strains were grown in MRS broth for 18 h. After centrifugation (8,000 ×g, 10 min), pellets were recovered, washed twice with 0.9% saline, and adjusted to 0.5 on the McFarland scale. One hundred microliters of this suspension was spread onto MRS agar plates, followed by the arrangement of antibiotic disks. Plates were incubated at 37°C for 18–24 h and the diameters of the halos were measured and classified as sensitive (S), moderately sensitive (MS), and resistant (R), according to Charteris et al. [27]. The antimicrobials tested were amikacin (30 μg), amoxicillin (10 μg), ampicillin (10 μg), cefalotin (30 μg), ciprofloxacin (5 μg), clindamycin (2 μg), chloramphenicol (30 μg), erythromycin (10 μg), gentamicin (10 μg), nitrofurantoin (300 μg), norfloxacin (10 μg), penicillin G (10 μg), tetracycline (30 μg), and vancomycin (30 μg).
2.9. Statistical Analysis
All experiments were performed in triplicate. Quantitative data are presented by mean and standard deviations that were analyzed using GraphPad Prism 5.01. Statistical differences between mean values were determined using ANOVA and Tukey's posttest with p < 0.05.
3. Results and Discussion
3.1. Autoaggregation, Hydrophobicity, and Adhesion to Vaginal Epithelial Cells
All three strains of lactobacilli tested in this study showed percentage of autoaggregation around 30% after 5 h of incubation (Table 1). Autoaggregation is an important bacterial feature in several ecological niches, especially in human and animal mucosa, where probiotics display their activities. The ability to autoaggregate (form floccules) is a crucial factor for the maintenance of significant counts of the probiotic strain in the adverse conditions present in the oral cavity and the gastrointestinal and urogenital tracts [28]. Lactobacilli, in general, have an autoaggregation capacity ranging from low to moderate [29]. In the present study, lactobacilli showed moderate autoaggregation close to or above those found for lactobacilli isolated from other fermented foods, including cocoa. Two strains of L. plantarum isolated from cocoa fermentation showed autoaggregation values of 18.08 and 20.94% [30]. Similarly, seven L. fermentum strains isolated from fermented Chinese products presented autoaggregation ranging from 0.86 to 65.15% [31]
Table 1.
Characterization of surface properties, adhesion to vaginal cells, and antimicrobial properties of lactobacilli isolated from cocoa fermentation.
| Strain | Surface properties | Adhesion to HMVII cells (%) | Antimicrobial properties | ||
|---|---|---|---|---|---|
| Autoaggregation (%) | Hydrophobicity (%) | Coaggregation with G. vaginalis (%) | Acidification | ||
| L. fermentum 5.2 | 31.18 ± 4.39a | 53.96 ± 2.90a | 35.61 ± 2.98a | 43.15 ± 0.68a | 4.78 |
| L. plantarum 6.2 | 33.44 ± 1.53a | 55.52 ± 3.76a | 38.73 ± 2.87a | 44.61 ± 0.17a | 3.81 |
| L. plantarum 7.1 | 29.23 ± 1.14a | 71.20 ± 3.03b | 55.75 ± 3.72b | 44.15 ± 0.51a | 3.77 |
Presented values are means of triplicate determinations; ± indicates standard deviations from the mean. Mean values (±standard deviation) within the same column followed by different superscript letters differ significantly (p < 0.05).
Hydrophobicity, also known as microbial adhesion to hydrocarbons (MATH), together with autoaggregation, is considered an important bacterial surface feature and can be classified into 3 categories: low (MATH < 33%), medium (33% < 66%), or high (MATH > 66%) [29]. In this study, hydrophobicity was evaluated by the microbial adhesion to xylene (an apolar solvent) and, after 2 h of incubation, results obtained for the three strains were L. fermentum 5.2 and L. plantarum 6.2 showed moderate hydrophobicity (53.96% and 55.52%, resp.) while L. plantarum 7.1 was highly hydrophobic (71.20%). These values of hydrophobicity are much higher than those found for other lactobacilli isolated from cocoa fermentation. Ramos et al. [30], testing a strain of L. fermentum and three strains of L. plantarum, obtained hydrophobicity values ranging from zero to 1.4%. Santos and coauthors [16], analyzing hydrophobicity of 3 strains of L. fermentum and 6 strains of L. plantarum, obtained values that varied from 3.5 to 16.9%, with the highest value attributed to a strain of L. plantarum.
Regarding the adhesion of Lactobacillus to HMVII epithelial vaginal cells, the strains L. fermentum 5.2 and L. plantarum 6.2 showed similar or almost equal percentage (35.61% and 38.78%, resp.), whereas L. plantarum 7.1 was significantly more adhesive (55.75%). It was possible to confirm this result by scanning electron microscopy images. L. plantarum 7.1 presented more bacteria adhered to HMVII cells when compared to the other two strains (Figure 1). Several studies correlate the ability of a probiotic strain to bind to host mucosal cells with autoaggregation and hydrophobicity acting synergistically [4, 29, 31]. This fact corroborates our data, where L. plantarum 7.1 expressed higher adhesion because it had significantly higher hydrophobicity than the other strains tested (Table 1). Studies employing lactobacilli isolated from environmental or intestinal samples showed a low adhesion to epithelial cells, usually around 10% [32, 33], a value much lower than that found with strains isolated from cocoa fermentation.
Figure 1.

Scanning electron microscope images of vaginal epithelial cells treated for 2 h with lactobacilli isolated from cocoa fermentation. (a) Untreated HMVII cells (×2,500); (b) HMVII cells treated with L. fermentum 5.2 (×2,500); (c) HMVII cells treated with L. plantarum 6.2 (×2,500); (d) HMVII cells treated with L. plantarum 7.1 (×2,500; details in ×20,000).
Miljkovic et al. [3] have demonstrated that extraintestinal strains of L. paracasei subsp. paracasei express AggLb, an aggregation-promoting factor that contributes to the diverse functions and behavior of the carriers, including strong aggregation and hydrophobicity abilities and strong and specific interaction with collagen through changes to cell-surface properties. AggLb is also involved in protection of the host from pathogen infection by a mechanism of competitive exclusion.
Bacterial surface properties (autoaggregation and hydrophobicity), as well as adhesion to host cells, are important criteria for the selection of probiotic bacteria strains [9, 21, 28]. Our findings show that the three tested strains have a good profile that could be used as vaginal probiotics.
3.2. Anti-Gardnerella Activity
Using the agar diffusion technique, we observed that only the supernatants of the L. plantarum strains used in this study (but not L. fermentum 5.2) showed antimicrobial activity against G. vaginalis, notable by the presence of inhibition halos around the wells. The supernatant halos of L. plantarum 6.2 and L. plantarum 7.1 were 12 and 11 mm, respectively (Figure 2(a)).
Figure 2.

Evaluation of antimicrobial activity of culture supernatants of lactobacilli isolated from cocoa fermentation against Gardnerella vaginalis. (a) Evaluation by agar diffusion. (b) Determination of minimum inhibitory concentrations. MRS: culture medium; Lf52: L. fermentum 5.2; Lp62: L. plantarum 6.2; and Lp71: L. plantarum 7.1.
Also in microdilution test where the minimum inhibitory concentration (MIC) was determined, inhibition of G. vaginalis was observed after exposure to the supernatants of both L. plantarum strains but not to L. fermentum. The minimum inhibitory concentration of both L. plantarum 6.2 and L. plantarum 7.1 supernatants was 10 mg mL−1. This effect was considered as bactericidal and confirmed by plating the contents of each well on the plate. After 24 h of incubation, there was no bacterial growth when G. vaginalis culture was exposed to 10 mg mL−1 or higher concentrations of culture supernatants (Figure 2(b)).
Antibacterial activity of L. plantarum supernatants alone may be related to their acidity, since the supernatants of L. plantarum 6.2 and L. plantarum 7.1 had pH of 3.81 and 3.77, respectively, while the pH of the supernatant of L. fermentum 5.2 was 4.78 (Table 1). The culture medium without any microbial growth had pH of 6.61. Some studies report that the difference in acid production is species-dependent in lactobacilli isolated from diverse sources. Supernatant of L. plantarum strain WSO, isolated from cucumber fermentation, had pH of 3.81 [34]. On the other hand, supernatant of a vaginal isolated L. fermentum with inhibitory potential against G. vaginalis had a pH of 4.16 [35]. Poppi et al. [36] showed that the pH of the supernatants of two L. plantarum strains (22c and 41b) isolated from poultry litter was 3.83 and 3.88, respectively.
Studies by other authors using the agar diffusion technique have shown that lactobacilli culture supernatants isolated from the vaginal microenvironment displayed inhibitory activity against Escherichia coli, Staphylococcus aureus, Proteus vulgaris, Klebsiella pneumoniae, and Gardnerella vaginalis [37–40]. After adjustment of pH to 6.5, the number of inhibitory strains was reduced to less than half of that observed when the supernatant was used without any treatment, indicating an important role of acids derived from the metabolism of lactobacilli in antibacterial activity [37]. The same effect was observed by Onwuakor et al. [38], where maize-isolated lactobacilli culture supernatants lost inhibitory activity against Salmonella typhimurium and Shigella dysenteriae when the pH was adjusted to values above 7.0. Antagonistic effects related to acid production (mainly lactic acid) have already been demonstrated for lactobacilli isolated from several sources. In these studies, exposure to high temperature or protease treatments did not significantly alter the antimicrobial activity of culture supernatants [39, 40].
In a study conducted by Melo et al. [24], the culture supernatant of an L. fermentum strain isolated from cocoa fermentation was able to inhibit the growth of S. aureus with an MIC of 20 mg mL−1. This effect, as found in our study for L. plantarum strains, was bactericidal and was confirmed by plating of treated culture. Similarly, a culture supernatant of a L. paracasei strain isolated from fermented milk was also shown to inhibit bacterial growth of pathogens, especially E. coli, with an MIC of 15.6 mg mL−1 [41]. However, to achieve the same effect on Serratia marcescens, values around 0.16 mg mL−1 of the culture supernatants from strains belonging to the species L. acidophilus and L. plantarum were required [5]. The activity of the culture supernatants against pathogens depends on several factors that include (1) the susceptibility of the target microorganism and (2) the composition of the lactobacilli supernatants, which differs in relation to the species, strain, and source of isolation, justifying the variation of MICs found in different studies [42].
The three strains of lactobacilli tested in our study showed high coaggregation values after incubation with G. vaginalis, greater than 40% (Table 1). Reduction of the adhesive activity of G. vaginalis bacteria by Lactobacillus strains is a well-known and desired effect of strains for potential vaginal probiotic application. In fact, other authors found that vaginal isolates of L. acidophilus, L. gasseri, and L. jensenii showed high coaggregation activity against C. albicans, E. coli, and G. vaginalis [4]. Mastromarino et al. [43] demonstrated high efficiency of coaggregation of L. salivarius and L. gasseri with G. vaginalis. In addition, strains of L. fermentum and L. plantarum isolated from cocoa fermentation efficiently coaggregated with E. coli, Shigella flexneri, Salmonella enterica, L. monocytogenes, and S. aureus [16, 30]. Coaggregation of probiotic microorganisms to pathogens generates a hostile environment for the pathogens implying the reduction of their growth, facilitation of the removal of the pathogen, and reestablishment of indigenous microbiota [44].
The coculture technique is able to assess the influence of one microorganism on the growth of another when both are incubated together. We observed that all Lactobacillus strains were able to reduce by one log unit the microbial counts of G. vaginalis after 24 hours of incubation when compared to G. vaginalis growing alone (Figure 3). Only L. fermentum 5.2 was able to maintain inhibitory activity against G. vaginalis during all the time period evaluated. It has been previously found that L. acidophilus, L. jensenii, L. gasseri, and L. crispatus isolated from the vaginal microbiota of healthy women showed inhibitory activity, demonstrated by the coculture technique, against G. vaginalis and Prevotella bivia with stable inhibition from the first hour [45]. These results were similar to those found by Coudeyras et al. [25] who, using a L. rhamnosus strain, demonstrated inhibition of G. vaginalis, P. bivia, and C. albicans after 8 hours, with significant inhibition of G. vaginalis after 24 hours. Other pathogens that are also capable of causing bacterial vaginosis, such as E. coli and S. aureus, also have their growth affected when cocultivated with strains of L. plantarum and L. fermentum: after 24 hours, a decrease of up to three logs was observed when compared to controls [46].
Figure 3.
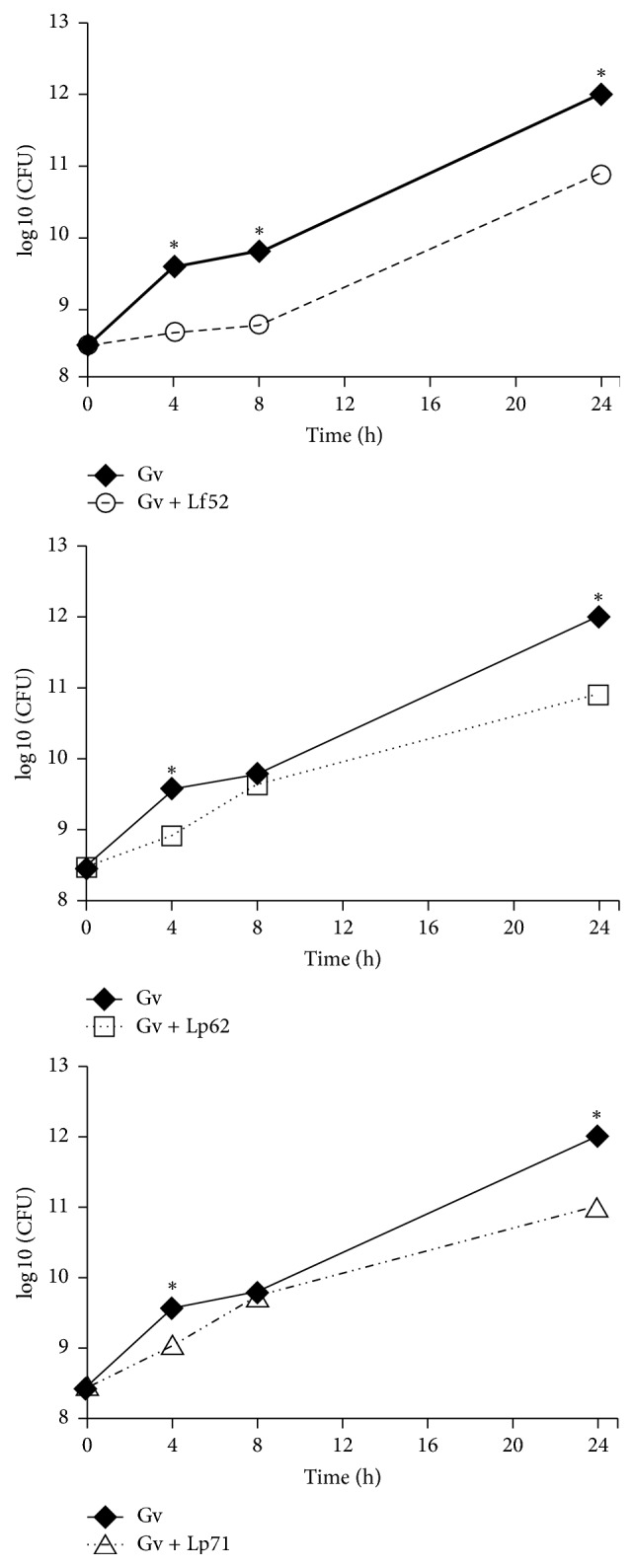
Effect of lactobacilli isolated from cocoa fermentation on the viability of Gardnerella vaginalis (Gv) as a function of the time of coculture. The pathogen was incubated without (filled shape) or with (empty shape) different lactobacilli (L. fermentum 5.2: Lf52; L. plantarum 6.2: Lp62; or L. plantarum 7.1: Lp71) for 24 hours and CFU mL−1 was determined after 4, 8, and 24 hours of incubation by plating onto appropriate media. Each value shown is the mean ± SD. ∗Statistically significant differences (p < 0.05).
In the present study, a concentration of 108 lactobacilli per mL was used in the coaggregation and coculture assays. Results found were satisfactory and promising, since such concentration was able to inhibit the growth of G. vaginalis after interaction. Commercial formulations and in vivo studies show that a concentration ranging from 108 to 109 CFU is required to achieve the same result [47–49].
3.3. Antimicrobial Susceptibility
Susceptibility of lactobacilli isolated from cocoa fermentation to different antimicrobials is shown in Table 2. Although lactobacilli have a long history of safe use, under certain host conditions they may cause rare bacteremia and endocarditis. Thus, some safety tests should be performed, such as antimicrobial susceptibility [4, 50]. The three strains of lactobacilli were sensitive to most antimicrobials tested and resistant to following antibiotics: vancomycin (a glycopeptide), aminoglycosides, and quinolones. Lactobacilli are generally resistant to antimicrobial inhibitors of nucleic acid synthesis, such as quinolones, whereas they are sensitive to cell wall inhibitors and protein synthesis inhibitors, except for vancomycin and aminoglycosides, respectively. It is important to emphasize that resistance to such antimicrobials is intrinsic to the genus Lactobacillus and does not present a risk of being transferred through horizontal genetic transfer to the bacteria of the native intestinal microbiota [9, 16, 51, 52].
Table 2.
Susceptibility profile of Lactobacillus strains isolated from cocoa fermentation.
| Antimicrobial | Susceptibilitya | ||||
|---|---|---|---|---|---|
| Group | Name | Disc conc. (µg) | L. fermentum 5.2 | L. plantarum 6.2 | L. plantarum 7.1 |
| Inhibitors of cell wall synthesis | |||||
| Penicillin | Amoxicillin | 10 | S | S | S |
| Ampicillin | 10 | S | S | S | |
| Penicillin G | 10 | S | MS | MS | |
| Cephalosporins | Cefalotin | 30 | S | S | S |
| Glycopeptides | Vancomycin | 30 | R | R | R |
| Inhibitors of protein synthesis | |||||
| Aminoglycosides | Amikacin | 30 | R | R | R |
| Gentamicin | 10 | R | R | R | |
| Streptomycin | 10 | R | R | R | |
| Tetracyclines | Tetracycline | 30 | S | S | MS |
| Single antibiotics | Chloramphenicol | 30 | S | S | S |
| Macrolides | Erythromycin | 15 | S | S | S |
| Lincosamides | Clindamycin | 2 | S | S | S |
| Inhibitors of nucleic acid synthesis | |||||
| Quinolones | Ciprofloxacin | 5 | R | R | R |
| Norfloxacin | 10 | R | R | R | |
| Other urinary tract antiseptics | |||||
| Single antibiotics | Nitrofurantoin | 300 | S | S | S |
aSusceptibility expressed as sensitive (S), moderately sensitive (MS), or resistant (R) [18].
4. Conclusion
Lactobacilli used in this study may protect the vaginal environment through multiple mechanisms, including adhesion to the epithelium, coaggregation with potential pathogens, and production of antagonistic molecules. They are promissory strains for the development of prophylactic agents. These results may serve as a basis for further studies aimed at investigating molecular mechanisms related to the inhibition of G. vaginalis by lactobacilli and their metabolites, as well as evaluating the immunomodulatory capacity of lactobacilli isolated from cocoa fermentation.
Conflicts of Interest
The authors declare that there are no conflicts of interest regarding the publication of this paper.
References
- 1.Ouwehand A. C., Salminen S., Isolauri E. Probiotics: an overview of beneficial effects. Antonie van Leeuwenhoek-Journal of Microbiology. 2002;82(1–4):279–289. doi: 10.1023/a:1020620607611. [DOI] [PubMed] [Google Scholar]
- 2.Cross M. L. Microbes versus microbes: Immune signals generated by probiotic lactobacilli and their role in protection against microbial pathogens. FEMS Immunology & Medical Microbiology. 2002;34(4):245–253. doi: 10.1016/S0928-8244(02)00377-2. [DOI] [PubMed] [Google Scholar]
- 3.Miljkovic M., Strahinic I., Tolinacki M., et al. AggLb is the largest cell-aggregation factor from Lactobacillus paracasei subsp. paracasei bgnj1-64, functions in collagen adhesion, and pathogen exclusion in vitro. PLoS ONE. 2015;10(5) doi: 10.1371/journal.pone.0126387.e0126387 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Hütt P., Lapp E., Štšepetova J., et al. Characterisation of probiotic properties in human vaginal lactobacilli strains. Microbial Ecology in Health & Disease. 2016;27:1–9. doi: 10.3402/mehd.v27.30484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Vahedi Shahandashti R., Kasra Kermanshahi R., Ghadam P. The inhibitory effect of bacteriocin produced by Lactobacillus acidophilus ATCC 4356 and Lactobacillus plantarum ATCC 8014 on planktonic cells and biofilms of Serratia marcescens. TURKISH JOURNAL OF MEDICAL SCIENCES. 2016;46(4):1188–1196. doi: 10.3906/sag-1505-51. [DOI] [PubMed] [Google Scholar]
- 6.Ahlberg S. H., Joutsjoki V., Korhonen H. J. Potential of lactic acid bacteria in aflatoxin risk mitigation. International Journal of Food Microbiology. 2015;207:87–102. doi: 10.1016/j.ijfoodmicro.2015.04.042. [DOI] [PubMed] [Google Scholar]
- 7.Santos T. F., Melo T. A., Almeida M. E., et al. Immunomodulatory effects of Lactobacillus plantarum Lp62 on intestinal epithelial and mononuclear cells. BioMed Research International. 2016;2016 doi: 10.1155/2016/8404156.8404156 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Jang S. E., Jeong J. J., Choi S. Y., et al. Lactobacillus rhamnosus HN001 and Lactobacillus acidophilus La-14 Attenuate Gardnerella vaginalis-Infected Bacterial Vaginosis in Mice. Nutrients. 2017;9:1–14. doi: 10.3390/nu9060531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Melo T. A., dos Santos T. F., Pereira L. R., et al. Functional profile evaluation of Lactobacillus fermentum TCUESC01: a new potential probiotic strain isolated during cocoa fermentation. BioMed Research International. 2017;2017:7. doi: 10.1155/2017/5165916.5165916 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Todorov S. D., von Mollendorff J. Evaluation of potential probiotic properties of Enterococcus mundtii, its survival in boza and in situ bacteriocin production. Food Technology and Biotechnology. 2005;47:178–191. [Google Scholar]
- 11.Todorov S. D., Wachsman M. B., Knoetze H., Meincken M., Dicks L. M. T. An antibacterial and antiviral peptide produced by Enterococcus mundtii ST4V isolated from soya beans. International Journal of Antimicrobial Agents. 2005;25(6):508–513. doi: 10.1016/j.ijantimicag.2005.02.005. [DOI] [PubMed] [Google Scholar]
- 12.Franzen M., Borgerhoff Mulder M. Ecological, economic and social perspectives on cocoa production worldwide. Biodiversity and Conservation. 2007;16(13):3835–3849. doi: 10.1007/s10531-007-9183-5. [DOI] [Google Scholar]
- 13.Schwan R. F. Cocoa fermentations conducted with a defined microbial cocktail inoculum. Applied and Environmental Microbiology. 1998;64(4):1477–1483. doi: 10.1128/aem.64.4.1477-1483.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Schwan R. F., Wheals A. E. The microbiology of cocoa fermentation and its role in chocolate quality. Critical Reviews in Food Science and Nutrition. 2004;44(4):205–221. doi: 10.1080/10408690490464104. [DOI] [PubMed] [Google Scholar]
- 15.Kumar H., Salminen S., Verhagen H., et al. Novel probiotics and prebiotics: Road to the market. Current Opinion in Biotechnology. 2015;32:99–103. doi: 10.1016/j.copbio.2014.11.021. [DOI] [PubMed] [Google Scholar]
- 16.Santos T. T., Ornellas R. M. S., Arcucio L. B., et al. Characterization of lactobacilli strains derived from cocoa fermentation in the south of Bahia for the development of probiotic cultures. LWT- Food Science and Technology. 2016;73:259–266. doi: 10.1016/j.lwt.2016.06.003. [DOI] [Google Scholar]
- 17.Dos Santos T. F., Melo T. A., Santos D. S., Rezende R. P., Dias J. C. T., Romano C. C. Efficacy of oral administration of lactic acid bacteria isolated from cocoa in a fermented milk preparation: Reduction of colitis in an experimental rat model. Genetics and Molecular Research. 2016;15(3) doi: 10.4238/gmr.15038097. [DOI] [PubMed] [Google Scholar]
- 18.Onderdonk A. B., Delaney M. L., Fichorova R. N. The human microbiome during bacterial vaginosis. Clinical Microbiology Reviews. 2016;29(2):223–238. doi: 10.1128/CMR.00075-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wang S., Wang Q., Yang E., Yan L., Li T., Zhuang H. Antimicrobial compounds produced by vaginal lactobacillus crispatus are able to strongly inhibit candida albicans growth, hyphal formation and regulate virulence-related gene expressions. Frontiers in Microbiology. 2017;8:1–11. doi: 10.3389/fmicb.2017.00564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Breshears L. M., Edwards V. L., Ravel J., Peterson M. L. Lactobacillus crispatus inhibits growth of Gardnerella vaginalis and Neisseria gonorrhoeae on a porcine vaginal mucosa model. BMC Microbiology. 2015;2015:1–12. doi: 10.1186/s12866-015-0608-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kos B., Šušković J., Vuković S., Sǐmpraga M., Frece J., Matošić S. Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilus M92. Journal of Applied Microbiology. 2003;94(6):981–987. doi: 10.1046/j.1365-2672.2003.01915.x. [DOI] [PubMed] [Google Scholar]
- 22.Rodríguez C., Cofré J. V., Sánchez M., Fernández P., Boggiano G., Castro E. Lactobacilli isolated from vaginal vault of dairy and meat cows during progesteronic stage of estrous cycle. Anaerobe. 2011;17(1):15–18. doi: 10.1016/j.anaerobe.2010.12.001. [DOI] [PubMed] [Google Scholar]
- 23.CLSI. Methods for diluition antimicrobial susceptibility tests for bacteria that grow aerobically; approved standard. 9th. Wayne, PA, USA: Clinical Laboratory Standards Institute; 2012. (CLSI document M07-A9). [Google Scholar]
- 24.Melo T. A., Dos Santos T. F., De Almeida M. E., et al. Inhibition of Staphylococcus aureus biofilm by Lactobacillus isolated from fine cocoa. BMC Microbiology. 2016;16(1):1–9. doi: 10.1186/s12866-016-0871-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Coudeyras S., Jugie G., Vermerie M., Forestier C. Adhesion of human probiotic Lactobacillus rhamnosus to cervical and vaginal cells and interaction with vaginosis-associated pathogens. Infectious Diseases in Obstetrics and Gynecology. 2008;2008:5. doi: 10.1155/2008/549640.549640 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.CLSI. Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Fifth Informational Supplement. Wayne, PA, USA: Clinical Laboratory Standards Institute; 2015. (CLSI document M100-S25). [Google Scholar]
- 27.Charteris W. P., Kelly P. M., Morelli L., Collins J. K. Antibiotic susceptibility of potentially probiotic Lactobacillus species. Journal of Food Protection. 1998;61(12):1636–1643. doi: 10.4315/0362-028X-61.12.1636. [DOI] [PubMed] [Google Scholar]
- 28.Nikolic M., Jovcic B., Kojic M., Topisirovic L. Surface properties of Lactobacillus and Leuconostoc isolates from homemade cheeses showing auto-aggregation ability. European Food Research and Technology. 2010;231(6):925–931. doi: 10.1007/s00217-010-1344-1. [DOI] [Google Scholar]
- 29.Bouchard D. S., Seridan B., Saraoui T., et al. Lactic acid bacteria isolated from bovine mammary microbiota: Potential allies against bovine mastitis. PLoS ONE. 2015;10(12) doi: 10.1371/journal.pone.0144831.e0144831 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ramos C. L., Thorsen L., Schwan R. F., Jespersen L. Strain-specific probiotics properties of Lactobacillus fermentum, Lactobacillus plantarum and Lactobacillus brevis isolates from Brazilian food products. Food Microbiology. 2013;36(1):22–29. doi: 10.1016/j.fm.2013.03.010. [DOI] [PubMed] [Google Scholar]
- 31.Li Q., Liu X., Dong M. Aggregation and adhesion abilities of lactic acid bacteria strains isolated from traditional fermented food. International Journal of Agricultural Policy and Research. 2015;3:84–92. [Google Scholar]
- 32.Duary R. K., Rajput Y. S., Batish V. K., Grover S. Assessing the adhesion of putative indigenous probiotic lactobacilli to human colonic epithelial cells. Indian Journal of Medical Research. 2011;137(11):664–671. doi: 10.4103/0971-5916.90992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.García-Cayuela T., Korany A. M., Bustos I., et al. Adhesion abilities of dairy Lactobacillus plantarum strains showing an aggregation phenotype. Food Research International. 2014;57:44–50. doi: 10.1016/j.foodres.2014.01.010. [DOI] [Google Scholar]
- 34.McDonald L. C., Fleming H. P., Hassan H. M. Acid tolerande of Leuconostoc mesenterioides and Lactobacillus plantarum. Applied and Environmental Microbiology. 1990;56:2120–2124. doi: 10.1128/aem.56.7.2120-2124.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Mclean N. W., Mcgroarty J. A. Growth inhibition of metronidazole-susceptible and metronidazole-resistant strains of Gardnerella vaginalis by lactobacilli in vitro. Applied and Environmental Microbiology. 1996;6:1089–1092. doi: 10.1128/aem.62.3.1089-1092.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Poppi L. B., Rivaldi J. D., Coutinho T. S., Astolfi-Ferreira C. S., Ferreira A. J. P., Mancilha I. M. Effect of Lactobacillus sp. isolates supernatant on Escherichia coli O157: H7 enhances the role of organic acids production as a factor for pathogen control. Pesquisa Veterinária Brasileira. 2015;35(4):353–359. doi: 10.1590/S0100-736X2015000400007. [DOI] [Google Scholar]
- 37.Andreeva P., Shterev A., Danova S. Antimicrobial activity of vaginal lactobacilli against Gardnerella vaginalis and pathogens. International Journal of Advanced Research in Biological Sciences. 2016;3:200–207. [Google Scholar]
- 38.Onwuakor C. E., Nwaugo V. O., Nnadi C. J., Emetole J. M. Effect of varied culture conditions on crude supernatant (bacteriocin) production from four Lactobacillus species isolated from locally fermented maize (ogi) American Journal of Microbiological Research. 2014;2(5):125–130. [Google Scholar]
- 39.Annuk H., Shchepetova J., Kullisaar T., Songisepp E., Zilmer M., Mikelsaar M. Characterization of intestinal lactobacilli as putative probiotic candidates. Journal of Applied Microbiology. 2003;94(3):403–412. doi: 10.1046/j.1365-2672.2003.01847.x. [DOI] [PubMed] [Google Scholar]
- 40.Hütt P., Shchepetova J., Lõivukene K., Kullisaar T., Mikelsaar M. Antagonistic activity of probiotic lactobacilli and bifidobacteria against entero- and uropathogens. Journal of Applied Microbiology. 2006;100(6):1324–1332. doi: 10.1111/j.1365-2672.2006.02857.x. [DOI] [PubMed] [Google Scholar]
- 41.Miao J., Guo H., Ou Y., et al. Purification and characterization of bacteriocin F1, a novel bacteriocin produced by Lactobacillus paracasei subsp. tolerans FX-6 from Tibetan kefir, a traditional fermented milk from Tibet, China. Food Control. 2014;42:48–53. doi: 10.1016/j.foodcont.2014.01.041. [DOI] [Google Scholar]
- 42.Servin A. L. Antagonistic activities of lactobacilli and bifidobacteria against microbial pathogens. FEMS Microbiology Reviews. 2004;28(4):405–440. doi: 10.1016/j.femsre.2004.01.003. [DOI] [PubMed] [Google Scholar]
- 43.Mastromarino P., Brigidi P., Macchia S., et al. Characterization and selection of vaginal Lactobacillus strains for the preparation of vaginal tablets. Journal of Applied Microbiology. 2002;93(5):884–893. doi: 10.1046/j.1365-2672.2002.01759.x. [DOI] [PubMed] [Google Scholar]
- 44.Younes J. A., van der Mei H. C., van den Heuvel E., Busscher H. J., Reid G. Adhesion forces and coaggregation between vaginal staphylococci and lactobacilli. PLoS ONE. 2012;7(5) doi: 10.1371/journal.pone.0036917.e36917 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Atassi F., Brassart D., Grob P., Graf F., Servin A. L. Lactobacillus strains isolated from the vaginal microbiota of healthy women inhibit Prevotella bivia and Gardnerella vaginalis in coculture and cell culture. FEMS Immunology & Medical Microbiology. 2006;48(3):424–432. doi: 10.1111/j.1574-695X.2006.00162.x. [DOI] [PubMed] [Google Scholar]
- 46.Shah N., Patel A., Ambalam P., Holst O., Ljungh A., Prajapati J. Determination of an antimicrobial activity of Weissella confusa, Lactobacillus fermentum, and Lactobacillus plantarum against clinical pathogenic strains of Escherichia coli and Staphylococcus aureus in co-culture. Annals of Microbiology. 2016;66(3):1137–1143. doi: 10.1007/s13213-016-1201-y. [DOI] [Google Scholar]
- 47.Pascual L., Ruiz F., Giordano W., Barberis I. L. Vaginal colonization and activity of the probiotic bacterium Lactobacillus fermentum L23 in a murine model of vaginal tract infection. Journal of Medical Microbiology. 2010;59(3):360–364. doi: 10.1099/jmm.0.012583-0. [DOI] [PubMed] [Google Scholar]
- 48.Daniele M., Pascual L., Barberis L. Curative effect of the probiotic strain lactobacillus fermentum L23 in a murine model of vaginal infection by gardnerella vaginalis. Letters in Applied Microbiology. 2014;59(1):93–98. doi: 10.1111/lam.12249. [DOI] [PubMed] [Google Scholar]
- 49.Vicariotto F., Mogna L., Del Piano M. Effectiveness of the two microorganisms Lactobacillus fermentum LF15 and Lactobacillus plantarum LP01, formulated in slow-release vaginal tablets, in women affected by bacterial vaginosis: A pilot study. Journal of Clinical Gastroenterology. 2014;48:S106–S112. doi: 10.1097/MCG.0000000000000226. [DOI] [PubMed] [Google Scholar]
- 50.Salminen M. K., Rautelin H., Tynkkynen S., et al. Lactobacillus bacteremia, species identification, and antimicrobial susceptibility of 85 blood isolates. Clinical infectious diseases : an official publication of the Infectious Diseases Society of America. 2006;42(5):e35–44. doi: 10.1086/500214. [DOI] [PubMed] [Google Scholar]
- 51.Shao Y., Zhang W., Guo H., Pan L., Zhang H., Sun T. Comparative studies on antibiotic resistance in Lactobacillus casei and Lactobacillus plantarum. Food Control. 2015;50:250–258. doi: 10.1016/j.foodcont.2014.09.003. [DOI] [Google Scholar]
- 52.Sharma P., Tomar S. K., Sangwan V., Goswami P., Singh R. Antibiotic Resistance of Lactobacillus sp. Isolated from Commercial Probiotic Preparations. Journal of Food Safety. 2016;36(1):38–51. doi: 10.1111/jfs.12211. [DOI] [Google Scholar]