

Abstract
Galangin (3,5,7-trihydroxyflavone) is a polyphenolic compound abundant in honey and medicinal herbs, such as Alpinia officinarum. In this study, we investigated the anti-inflammatory effects of galangin under in vitro and in vivo neuroinflammatory conditions caused by polyinosinic-polycytidylic acid (poly(I:C)), a viral mimic dsRNA analog. Galangin suppressed the production of nitric oxide, reactive oxygen species, and pro-inflammatory cytokines in poly(I:C)-stimulated BV2 microglia. On the other hand, galangin enhanced anti-inflammatory interleukin (IL)-10 production. Galangin also suppressed the expression of pro-inflammatory markers in poly(I:C)-injected mouse brains. Further mechanistic studies showed that galangin inhibited poly(I:C)-induced nuclear factor (NF)-κB activity and phosphorylation of Akt without affecting MAP kinases. Interestingly, galangin increased the expression and transcriptional activity of peroxisome proliferator-activated receptor (PPAR)-γ, known to play an anti-inflammatory role. To investigate whether PPAR-γ is involved in the anti-inflammatory function of galangin, BV2 cells were pre-treated with PPAR-γ antagonist before treatment of galangin. We found that PPAR-γ antagonist significantly blocked galangin-mediated upregulation of IL-10 and attenuated the inhibition of tumor necrosis factor (TNF)-α and IL-6 in poly(I:C)-stimulated microglia. In conclusion, our data suggest that PI3K/Akt, NF-κB, and PPAR-γ play a pivotal role in mediating the anti-inflammatory effects of galangin in poly(I:C)-stimulated microglia.
Keywords: Galangin, Poly(I:C), Microglia, PI3K/Akt, NF-κB, PPAR-γ signaling
INTRODUCTION
Microglia are resident immune cells in the brain and play a role in neuronal survival during brain development via growth factors and phagocytosis of apoptotic cells and debris throughout the entire brain (Graeber and Streit, 2010; Ulland et al., 2015). By challenge with various immunogens (e.g., lipopolysaccharides) and pathogens (e.g., bacteria, viruses), resting microglia transform into an active form and secrete reactive oxygen species, prostaglandins, matrix metalloproteinases (MMPs), and pro-inflammatory cytokines (Hoogland et al., 2015). If the acute inflammation shifts to an anti-inflammatory/resolution phase, it leads to tissue repair and homeostasis. However, a persistent and unresolved inflammation leads to neuronal cell death and contributes to the generation of neurodegenerative disorders (Glass et al., 2010; Cherry et al., 2014).
Recent studies reported that systemic inflammation plays an important role in the occurrence and progression of neuroinflammatory disorders. Systemic challenges with infectious stimuli such as lipopolysaccharide (LPS), polyinosinic-polycytidylic acid (poly(I:C)), lipoteichoic acid, and live bacteria result in microglial activation and upregulate inflammatory responses in the brain (Cunningham et al., 2007; Qin et al., 2007; Hoogland et al., 2015). These neuroinflammatory conditions may eventually lead to neurodegenerative diseases. Thus, the development of compounds that can control systemic inflammation and/or microglial activation has been suggested as a potential therapeutic target for neurodegenerative diseases.
Galangin (3,5,7-trihydroxyflavone) is a polyphenolic compound enriched in honey and medicinal herbs, such as Alpinia officinarum (Jaganathan and Mandal, 2009; Honmore et al., 2016). Galangin is known to have anti-tumor, anti-oxidant, anti-inflammatory, and anti-microbial activities (Meyer et al., 1997; Cushnie et al., 2007; Ruweler et al., 2008; Chien et al., 2015; Honmore et al., 2016). The anti-tumor effects of galangin have been reported in human leukemia, hepatoma, and breast cancer cells (Heo et al., 2001; Jaganathan and Mandal, 2009; Chien et al., 2015). In addition, the anti-inflammatory effects of galangin have been reported in several animal models such as paw edema, asthma, and rheumatoid arthritis (Huh et al., 2013; Liu et al., 2015; Honmore et al., 2016). Galangin also showed neuroprotective effects in a model of acute ischemic stroke by restoring cortical blood flow and ameliorating apoptosis and mitochondrial dysfunction (Li et al., 2012). Moreover, galangin inhibited β-amyloid production and acetylcholinesterase activity, suggesting therapeutic potential for Alzheimer disease as well as cerebral ischemia (Guo et al., 2010; Zeng et al., 2015).
Although the pharmacological activities of galangin have been reported in many papers, the anti-inflammatory effects of galangin in the brain have not been fully demonstrated. In this study, we examined the effects of galangin in poly(I:C)-induced neuroinflammatory conditions and analyzed its underlying molecular mechanisms. Our data showed that galangin exerts anti-inflammatory effects by modulating PI3K/Akt, NF-κB, and PPAR-γ signaling pathways.
MATERIALS AND METHODS
Reagent
Galangin (3,5,7-trihydroxyflavone), poly(I:C), and PPAR-γ antagonist (T0070907) were obtained from Sigma-Aldrich (St. Louis, MO, USA). All reagents used for cell culture were purchased from Welgene (Gyeongsan, Korea). Trizol reagent was obtained from Thermo Fisher Scientific (Waltham, MA, USA). Antibodies against phospho-/total forms of p38, JNK/SARK, ERK1/2, and Akt were supplied by Cell Signaling Technology (Beverley, CA, USA). The structure of galangin is shown in Fig. 1.
Fig. 1.
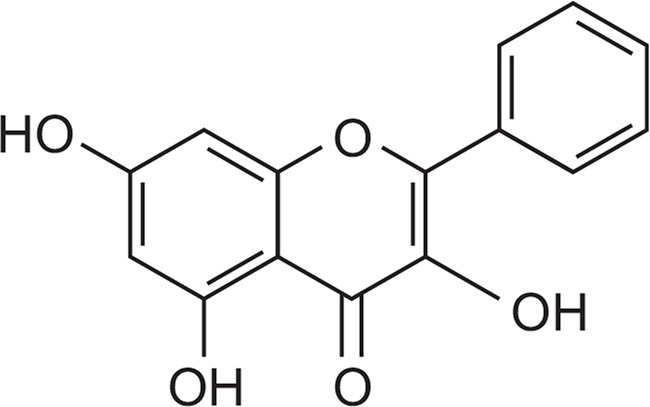
The chemical structure of galangin (3,5,7-trihydroxyflavone).
Microglial cell culture
The immortalized mouse BV2 microglial cell line (obtained from Dr. Hong JS, NIEHS, USA) was grown and maintained in Dulbecco’s modified Eagle’s medium supplemented with 10% heat-inactivated fetal bovine serum, streptomycin (10 μg/mL), and penicillin (10 U/mL) at 37°C under 5% CO2.
Measurement of nitric oxide, cytokine, and intracellular ROS levels
BV2 cells (1×105 cells per well in a 24-well plate) were pre-treated with galangin for 1 h and stimulated with poly(I:C) (10 μg/ml). After 16 h, the supernatants were collected and the accumulated nitrite was measured using the Griess reagent (Promega, Madison, WI, USA). The concentrations of TNF-α, IL-6, and IL-10 in the supernatants were measured by ELISA according to the procedure recommended by the supplier (BD Biosciences, San Jose, CA, USA). The intracellular accumulation of ROS was measured with 2′,7′-dichlorodihydrofluorescein diacetate (H2DCF-DA) (Sigma-Aldrich). In brief, microglia were stimulated with poly(I:C) for 16 h and stained with 20 mM H2DCF-DA in PBS for 1 h at 37°C. DCF fluorescence intensity was measured at 485-nm excitation and 535-nm emission, using a fluorescence plate reader (Molecular Devices, Sunnyvale, CA, USA).
RT-PCR
BV2 cells (7.5×105 cells per well in a 6-well plate) were treated with poly(I:C) in the presence or absence of galangin, and total RNA was extracted using TRI reagent (Thermo Fisher Scientific). Cortical brain tissue was homogenized using a homogenizer (Fisher Scientific, Pittsburgh, PA, USA), and total RNA was extracted using TRI reagent. For RT-PCR, total RNA (1 μg) was reverse-transcribed in a reaction mixture containing 0.1 μg of random primers, 3 mM MgCl2, 0.5 mM dNTP, 1× RT buffer, and 10 U reverse transcriptase (Promega). The synthesized cDNA was used as a template for the PCR reaction using Go Taq polymerase (Promega) and primers for the target gene. The primers used in the PCR reaction are shown in Table 1.
Table 1.
Primers used in RT-PCR reactions
| Gene | Forward Primer (5′→3′) | Reverse Primer (5′→3′) | Size |
|---|---|---|---|
| iNOS | GCTTGGGTCTTGTTCACTCC | GGCCTTGTGGTGAAGAGTGT | 385 bp |
| TNF-α | CCTATGTCTCAGCCTCTTCT | CCTGGTATGAGATAGCAAAT | 354 bp |
| IL-6 | CCACTTCACAAGTCGGAGGCTT | CCAGCTTATCTGTTAGGAGA | 214 bp |
| IL-1β | GGCAACTGTTCCTGAACTCAACTG | CCATTGAGGTGGAGAGCTTTCAGC | 447 bp |
| COX-2 | TTCAAAAGAAGTGCTGGAAAAGGT | GATCATCTCTACCTGAGTGTCTTT | 304 bp |
| IL-10 | GCCAGTACAGCCGGGAAGACAATA | GCCTTGTAGACACCTTGGTCTT | 185 bp |
| MMP8 | ATTCAGTCCCTCTATGGA | CTCCAGTATTTGTCCTCTAC | 245 bp |
| PPAR-γ | CCGAAGAACCATCCGATT | CGGGAAGGACTTTATGTA | 271 bp |
| GAPDH | TTCACCACCATGGAGAAGGC | GGCATGGACTGTGGTCATGA | 236 bp |
Mice
Adult male ICR mice (Mus musculus, 28–32 g, 7 weeks old) were purchased from Samtako Inc (Osan, Korea). All animal experiments were approved by the Institutional Animal Care and Research Ethics Committee at the School of Medicine, Ewha Womans University (IACUC #2014-0274), and were carried out in accordance with the guidelines of the National Institute of Heath’s Guide for the Care and Use of Laboratory Animals.
Poly(I:C)-induced inflammation and administration of galangin
Poly(I:C) (12 mg/kg) was administered intraperitoneally (i.p.) to induce neuroinflammation in male ICR mice, as previously described (Jeong et al., 2017). Galangin (50 mg/kg) dissolved in vehicle solution (normal saline containing 1% dimethylsulfoxide), was administered daily (i.p.) for 4 days before the poly(I:C) treatment. The brains were obtained 3 h after LPS administration and analyzed for the expression of inflammatory markers.
Electrophoretic mobility shift assay (EMSA)
BV2 cells were pretreated with galangin for 1 h and stimulated with poly(I:C) for 3 h. The nuclear extracts from the cells were prepared as previously described (Lee et al., 2015). Double-stranded DNA oligonucleotides containing consensus sequences of NF-κB were end-labeled using T4 polynucleotide kinase (New England Biolabs, Beverly, MA, USA) in the presence of [γ-32P] ATP. Nuclear proteins (5 μg) were incubated with a 32P-labeled probe on ice for 30 min, resolved on a 5% polyacrylamide gel, and visualized with autoradiography.
Transient transfection and luciferase assay
BV2 cells (2×105 cells per well on a 12-well plate) were transfected with 1 μg of plasmid DNA ([κB]3-luc or PPRE-luc) using the Metafectene transfection reagent (Biontex, Martinsried/Planegg, Germany). The effect of galangin on reporter gene activity was determined by pre-treating the cells with galangin prior to stimulation with poly(I:C) (10 μg/mL) for 6 h. After preparing the cell lysates, the luciferase assay was performed as previously described (Lee et al., 2015).
Western blot analysis
Whole cell protein lysates were prepared in a lysis buffer (10 mM Tris, pH 7.4, 30 mM NaCl, 1% Triton X-100, 0.1% SDS, 0.1% sodium deoxycholate, and 1 mM EDTA) containing a protease inhibitor cocktail. Protein samples were separated by SDS-PAGE, transferred to nitrocellulose membranes, and incubated with antibodies against the phospho- or the total form of MAP kinase (1:1000), Akt (1:1000) (Cell Signaling), and β-actin (1:1000) (Sigma). After thoroughly washing with TBST, HRP-conjugated secondary antibodies (1:2000 dilution in TBST; Bio-Rad, Hercules, CA, USA) were applied, and the blots were developed using an enhanced chemiluminescence detection kit (Thermo Fisher Scientific).
Statistical analysis
Data are expressed as mean ± standard error of the mean (SEM), and statistical analyses were performed using a one-way ANOVA followed by Newman-Keuls post-hoc tests or t-tests. The calculations were performed using the GraphPad Prism software (version 4.0; GraphPad Software, Inc., La Jolla, CA, USA). A p-value of less than 0.05 indicated statistical significance.
RESULTS
Galangin inhibits poly(I:C)-induced production of NO, pro-inflammatory cytokines, and ROS with enhancement of anti-inflammatory IL-10 in BV2 microglial cells
To examine the anti-inflammatory effects of galangin, BV2 cells were stimulated with poly(I:C) (10 μg/m) in the presence or absence of galangin. Poly(I:C) is a viral mimic dsRNA analog that activates the toll-like receptor 3 (TLR3) pathway (Cunningham et al., 2007). Galangin suppressed poly(I:C)-induced NO, TNF-α, and IL-6 production in a concentration-dependent manner (Fig. 2A). Moreover, galangin inhibited intracellular ROS production. On the other hand, galangin remarkably increased anti-inflammatory IL-10 production (Fig. 2A). Next, RT-PCR analysis was performed to determine the effect of galangin on mRNA expression of pro-/anti-inflammatory molecules in poly(I:C)-stimulated microglia. As shown in Fig. 2B, galangin significantly reduced the mRNA levels of iNOS, TNF-α, IL-6, IL-1β, and COX-2, induced by poly(I:C), while galangin increased IL-10 mRNA levels. The results suggest that galangin modulates the expression of iNOS, COX-2, and several pro-/anti-inflammatory cytokines at the transcriptional level.
Fig. 2.

Effects of galangin on the expression of pro-/anti-inflammatory molecules in poly(I:C)-stimulated BV2 microglia. (A) Cells were pre-treated with galangin (Gal) at the indicated concentrations (10, 30, 50 μM) for 1 h, and then subjected to treatment with poly(I:C) (10 μg/ml) for 16 h. The amounts of NO, TNF-α, IL-6, and IL-10 released into the medium were measured using Griess reagent or ELISA. Intracellular ROS levels were determined by the DCF-DA method. (B) Cells were pre-treated with galangin for 1 h, and then treated with poly(I:C) (10 μg/ml) for 6 h. An RT-PCR was performed to measure the mRNA expression of pro-/anti-inflammatory molecules. The quantification data are shown in the right panel. The data are expressed as the mean ± SEM of three independent experiments. *p<0.05, vs. control samples. #p<0.05, vs. poly(I:C)-treated samples.
Galangin suppresses the expression of pro-inflammatory markers in poly(I:C)-injected mouse brains
To confirm the effects of galangin under neuroinflammatory conditions in vivo, galangin was intraperitoneally injected into mice prior to the administration of poly(I:C). The expression levels of proinflammatory markers were determined in poly(I:C)-injected mouse brains 3 hours after injection. RT-PCR analysis showed that galangin inhibited the mRNA expression of iNOS, TNF-α, IL-6, and IL-1β in poly(I:C)-injected mouse brains (Fig. 3). Galangin also suppressed the expression of MMP-8, which plays a role as a pro-inflammatory mediator in activated microglia (Fig. 3) (Lee et al., 2015).
Fig. 3.

Effect of galangin on mRNA expression of inflammatory markers in poly(I:C)-injected mouse brains. (A, B) Effects of galangin (50 mg/kg) on mRNA levels of iNOS, pro-inflammatory cytokines (IL-6, IL-1β, TNF-α), and MMP-8 in the cortex of poly(I:C)-injected mice. Representative gels (A), and quantification data (B) are shown (n=5). *p<0.05, vs. control samples. #p<0.05, vs. poly(I:C)-treated samples.
Galangin inhibits poly(I:C)-induced NF-κB activity and phosphorylation of Akt without affecting MAPK signaling
To determine the molecular mechanism underlying the anti-inflammatory effects of galangin, we examined the effect of galangin on the TLR3 signaling pathway. After dsRNA binds to TLR3, it activates intracellular signaling molecules such as PI3K, MAPKs, and downstream transcription factors such as NF-κB, AP-1, and IRFs, which lead to the expression of various immune genes (Moynagh, 2005; Sen and Sarkar., 2005). Therefore, in the present study, we examined the effects of galangin on a key transcription factor NF-κB as well as signaling kinases such as PI3K/Akt and three types of MAP kinases. The EMSA data showed that galangin significantly inhibited poly(I:C)-induced NF-κB DNA binding activity (Fig. 4A). Moreover, galangin inhibited NF-κB reporter gene activity (Fig. 4B). We also found that galangin significantly inhibited poly(I:C)-induced phosphorylation of Akt, without affecting p38 MAPK, ERK, or JNK phosphorylation (Fig. 4C). The data suggest that PI3K/Akt and NF-κB signaling pathways may be involved in the anti-inflammatory mechanism of galangin.
Fig. 4.

Effect of galangin on NF-κB activity and the phosphorylation of Akt and MAP kinases in poly(I:C)-stimulated BV2 cells. (A) Nuclear extracts were prepared from BV2 cells after treatment with poly(I:C) (10 μg/ml) for 3 h, in the absence or presence of galangin, and EMSA was performed to determine the DNA binding activity of NF-κB. The arrow indicates the DNA-protein complex of NF-κB. (B) BV2 cells transfected with the reporter plasmid ([κB]3-luc) were pretreated with galangin for 1 h prior to poly(I:C) treatment. After 6 h, cells were harvested and a luciferase assay was performed. (C) Western blot analysis for MAPKs and PI3K/Akt activities. Cell extracts were prepared from BV2 cells treated with poly(I:C) for 30 min, in the absence or presence of galangin, and a western blot analysis was performed using antibodies against phospho- or total forms of three types of MAPKs or Akt. The blots are representative of three independent experiments. Quantification of data from western blot analysis is shown in the bottom panel. The levels of the phosphorylated forms of MAPKs or Akt were normalized to the levels of each total form and expressed as relative fold changes versus the untreated control group. The results were obtained from three independent experiments and expressed as the mean ± SEM. *p<0.05, vs. control samples. #p<0.05, vs. poly(I:C)-treated samples.
PPAR-γ signaling appears to mediate anti-inflammatory activity of galangin in poly(I:C)-stimulated microglia
PPAR-γ is a nuclear receptor that acts as an anti-inflammatory regulator in macrophage and microglial activation (Bouhlel et al., 2007; Saijo et al., 2013). In addition, many papers reported that activation of PPAR-γ can suppress the inflammatory response through inhibition of NF-κB (Saijo et al., 2013). In the present study, we found that galangin increased PPAR-γ-mediated transcriptional activity in poly(I:C)-stimulated BV2 cells (Fig. 5A), and pre-treatment with PPAR-γ antagonist (T0070907) abolished the increased PPRE-luc activity (Fig. 5A). These data indicate that the upregulation of PPRE-luc activity by galangin was mediated by PPAR-γ. Furthermore, we found that the treatment of BV2 cells with poly(I:C) significantly suppressed PPAR-γ expression, and treatment with galangin restored PPAR-γ expression (Fig. 5B).
Fig. 5.

Galangin increases PPAR-γ transcriptional activity and expression, and PPAR-γ antagonist reverses the anti-inflammatory effect of galangin in poly(I:C)-stimulated BV2 cells. (A) BV2 cells were transfected with PPRE-luc and treated with poly(I:C) with or without galangin. After 6 h, the cells were harvested and the luciferase assay was performed. The PPAR-γ antagonist, T0070907 (5 μM), was added to BV2 cells 1 h before treatment with galangin. (B) BV2 cells were pretreated with galangin for 1 h prior to incubation with poly(I:C) for 6 h. The mRNA level of PPAR-γ was determined by RT-PCR. Quantification data are shown in the bottom panel. (C) The effect of T0070907 on the production of IL-10, IL-6, TNF-α, and NO in poly(I:C) +galangin-treated cells. BV2 cells were pre-treated with the indicated concentrations (1, 3, 5 μM) of T0070907 for 1 h, and then galangin (50 μM) for 1 h, followed by treatment with poly(I:C) (10 μg/ml) for 16 h. The amounts of IL-10, IL-6, TNF-α, and NO released into the medium were determined. Data are expressed as mean ± SEM from four independent experiments. *p<0.05, vs. control samples. #p<0.05, vs. poly(I:C)-treated samples. ##p<0.05, vs. poly(I:C) +galangin-treated samples.
To investigate whether PPAR-γ mediates the anti-inflammatory effects of galangin, BV2 cells were treated with T0070907 before the addition of poly(I:C) and/or galangin. As shown in Fig. 5C, treatment with T0070907 significantly blocked galangin-mediated upregulation of IL-10, and the inhibition of IL-6 and TNF-α in poly(I:C)-stimulated microglia. However, it did not affect NO production. These data collectively suggest that PPAR-γ is at least partly involved in anti-inflammatory mechanism of galangin in activated microglia.
DISCUSSION
The present study showed the anti-inflammatory effects of galangin in poly(I:C)-induced neuroinflammatory conditions in vitro and in vivo. Galangin suppressed the expression of pro-inflammatory molecules such as iNOS, COX-2, and proinflammatory cytokines, and enhanced anti-inflammatory IL-10 in poly(I:C)-stimulated microglia (Fig. 2). Moreover, galangin inhibited the expression of proinflammatory markers in poly(I:C)-injected mouse brains (Fig. 3). Detailed mechanistic studies showed that PI3K/Akt, NF-κB, and PPAR-γ are involved in mediating the anti-inflammatory effects of galangin in poly(I:C)-stimulated BV2 microglia (Fig. 4, 5). The data collectively suggest the therapeutic potential of galangin in neuroinflammatory disorders.
Previous studies demonstrated the role of PPAR-γ in neuroinflammation (Zolezzi et al., 2017). PPAR-γ is expressed in microglia and mediates anti-inflammatory activity by inducing microglial polarization into the M2-like phenotype (Saijo et al., 2013). Moreover, PPAR-γ is known to suppress the expression of pro-inflammatory cytokines, chemokines, and MMPs via inhibition of transcription factors, such as NF-κB, STAT, and AP-1 (Bernado and Minghetti, 2006). In particular, several mechanisms have been proposed underlying the NF-κB regulation by PPAR-γ. First, PPAR-γ competes with NF-κB for binding to coactivators, such as p300/CBP, and leads to inhibition of a NF-κB-mediated inflammatory response (Zhao et al., 2015). Second, PPAR-γ directly binds to NF-κB subunits p50/p65 and inhibits the DNA binding activity of NF-κB (Chung et al., 2008). Lastly, PPAR-γ inhibits NF-κB-mediated transcription via transrepression through preventing clearance of co-repressor complexes from NF-κB (Saijo et al., 2013). Therefore, at least one of these mechanisms might be related to PPAR-γ-mediated inhibition of NF-κB in galangin+poly(I:C)-treated microglial cells. Further studies are necessary to unravel the detailed mechanism.
In the present study, we found that galangin inhibited the phosphorylation of Akt in LPS-stimulated microglia (Fig. 4). Several papers have reported that PI3K/Akt signaling pathways play an important role in pro-inflammatory gene expression in activated microglia (Kim et al., 2004; Saponaro et al., 2012). Kim et al. (2004) reported that the PI3K inhibitor LY294002 suppressed the expression of iNOS, pro-inflammatory cytokines, and MMP-9 in LPS-stimulated BV2 cells. In addition, Saponaro et al. (2012) reported that the PI3K/Akt pathway plays a role in NF-κB-dependent inflammatory responses of activated microglia. Therefore, in addition to PPAR-γ, the PI3K/Akt signaling pathway appears to be involved in the inhibition of NF-κB and subsequent downregulation of proinflammatory gene expression by galangin in poly(I:C)-stimulated BV2 microglial cells.
Recent studies have reported on the therapeutic benefits of galangin in several neurological disorders. Galangin showed neuroprotective effects in a model of focal cerebral ischemia by improving regional cortical blood flow and attenuating mitochondrial dysfunction and apoptosis (Li et al., 2012). Moreover, galangin showed therapeutic potential for Alzheimer disease. It inhibited acetylcholine esterase activity (Guo et al., 2010) and decreased β-amyloid production by suppressing BACE activity, which may be related to PPAR- activation (Sastre et al., 2003; Descamps et al., 2013). Recently, galangin has been suggested as one of the candidate compounds for the treatment of Alzheimer disease (Ahmad et al., 2016). Therefore, the anti-inflammatory effects of galangin may provide therapeutic potential for various neurodegenerative diseases that are associated with neuroinflammation.
Acknowledgments
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF), funded by the Ministry of Science, ICT & Future Planning (Grant No. NRF-2010-0027945).
REFERENCES
- Ahmad SS, Akhtar S, Jamal QM, Rizvi SM, Kamal MA, Khan MK, Siddiqui MH. Multiple targets for the management of Alzheimer’s disease. CNS Neurol. Disord. Drug Targets. 2016;15:1279–1289. doi: 10.2174/1871527315666161003165855. [DOI] [PubMed] [Google Scholar]
- Bernado A, Minghetti L. PPAR-γ agonists as regulators of microglial activation and brain inflammation. Curr Pharm Des. 2006;12:93–109. doi: 10.2174/138161206780574579. [DOI] [PubMed] [Google Scholar]
- Bouhlel MA, Derudas B, Rigamonti E, Dièvart R, Brozek J, Haulon S, Zawadzki C, Jude B, Torpier G, Marx N, Staels B, Chinetti-Gbaguidi G. PPARγ activation primes human monocytes into alternative M2 macrophages with anti-inflammatory properties. Cell Metab. 2007;6:137–143. doi: 10.1016/j.cmet.2007.06.010. [DOI] [PubMed] [Google Scholar]
- Cherry JD, Olschowka JA, O’Banion MK. Neuroinflammation and M2 microglia: the good, the bad, and the inflamed. J. Neuroinflammation. 2014;11:98. doi: 10.1186/1742-2094-11-98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chien ST, Shi MD, Lee YC, Te CC, Shih YW. Galangin, a novel dietary flavonoid, attenuates metastatic feature via PKC/ERK signaling pathway in TPA-treated liver cancer HepG2 cells. Cancer Cell Int. 2015;15:15. doi: 10.1186/s12935-015-0168-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung JH, Seo AY, Chung SW, Kim MK, Leeuwenburgh C, Yu BP, Chung HY. Molecular mechanism of PPAR in the regulation of age-related inflammation. Ageing Res Rev. 2008;7:126–136. doi: 10.1016/j.arr.2008.01.001. [DOI] [PubMed] [Google Scholar]
- Cunningham C, Campion S, Teeling J, Felton L, Perry VH. The sickness behaviour and CNS inflammatory mediator profile induced by systemic challenge of mice with synthetic double-stranded RNA (poly I:C). Brain Behav Immun. 2007;21:490–502. doi: 10.1016/j.bbi.2006.12.007. [DOI] [PubMed] [Google Scholar]
- Cushnie TP, Hamilton VE, Chapman DG, Taylor PW, Lam AJ. Aggregation of Staphylococcus aureus following treatment with the antibacterial flavonol galangin. J Appl Microbiol. 2007;103:1562–1567. doi: 10.1111/j.1365-2672.2007.03393.x. [DOI] [PubMed] [Google Scholar]
- Descamps O, Spilman P, Zhang Q, Libeu CP, Poksay K, Gorostiza O, Campagna J, Jagodzinska B, Bredesen DE, John V. AβPP-selective BACE inhibitors (ASBI): novel class of therapeutic agents for alzheimer’s disease. J Alzheimers Dis. 2013;37:343–355. doi: 10.3233/JAD-130578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glass CK, Saijo K, Winner B, Marchetto MC, Gage FH. Mechanisms underlying inflammation in neurodegeneration. Cell. 2010;140:918–934. doi: 10.1016/j.cell.2010.02.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graeber MB, Streit WJ. Microglia: biology and pathology. Acta Neuropathol. 2010;119:89–105. doi: 10.1007/s00401-009-0622-0. [DOI] [PubMed] [Google Scholar]
- Guo AJ, Xie HQ, Choi RC, Zheng KY, Bi CW, Xu SL, Dong TT, Tsim KW. Galangin, a flavonol derived from Rhizoma Alpiniae Officinarum, inhibits acetylcholinesterase activity in vitro. Chem Biol Interact. 2010;187:246–248. doi: 10.1016/j.cbi.2010.05.002. [DOI] [PubMed] [Google Scholar]
- Heo MY, Sohn SJ, Au WW. Anti-genotoxicity of galangin as a cancer chemopreventive agent candidate. Mutat Res. 2001;488:135–150. doi: 10.1016/S1383-5742(01)00054-0. [DOI] [PubMed] [Google Scholar]
- Honmore VS, Kandhare AD, Kadam PP, Khedkar VM, Sarkar D, Bodhankar SL, Zanwar AA, Rojatkar SR, Natu AD. Isolates of Alpinia officinarum Hance as COX-2 inhibitors: Evidence from anti-inflammatory, antioxidant and molecular docking studies. Int Immunopharmacol. 2016;33:8–17. doi: 10.1016/j.intimp.2016.01.024. [DOI] [PubMed] [Google Scholar]
- Hoogland IC, Houbolt C, van Westerloo DJ, van Gool WA, van de Beek D. Systemic inflammation and microglial activation: systematic review of animal experiments. J. Neuroinflammation. 2015;12:114. doi: 10.1186/s12974-015-0332-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huh J, Jung I, Choi J, Baek Y, Lee J, Park D, Choi D. The natural flavonoid galangin inhibits osteoclastic bone destruction and osteoclastogenesis by suppressing NF-κB in collagen-induced arthritis and bone marrow-derived macrophages. Eur J Pharmacol. 2013;698:57–66. doi: 10.1016/j.ejphar.2012.08.013. [DOI] [PubMed] [Google Scholar]
- Jaganathan SK, Mandal M. Antiproliferative effects of honey and of its polyphenols: a review. J Biomed Biotechnol. 2009;2009:830616. doi: 10.1155/2009/830616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeong YH, Park JS, Kim DH, Kang JL, Kim HS. Anti-inflammatory mechanism of lonchocarpine in LPS- or poly(I:C)-induced neuroinflammation. Pharmacol Res. 2017;119:431–442. doi: 10.1016/j.phrs.2017.02.027. [DOI] [PubMed] [Google Scholar]
- Kim WK, Hwang SY, Oh ES, Pia HZ, Kim KW, Han IO. TGF-β1 represses activation and resultant death of microglia via inhibition of phosphatidylinositol 3-kinase activity. J Immunol. 2004;172:7015–7023. doi: 10.4049/jimmunol.172.11.7015. [DOI] [PubMed] [Google Scholar]
- Lee EJ, Ko HM, Jeong YH, Park EM, Kim HS. β-Lapachone suppresses neuroinflammation by modulating the expression of cytokines and matrix metalloproteinases in activated microglia. J. Neuroinflammation. 2015;12:133. doi: 10.1186/s12974-015-0355-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li S, Wu C, Zhu L, Gao J, Fang J, Li D, Fu M, Liang R, Wang L, Cheng M, Yang H. By improving regional cortical blood flow, attenuating mitochondrial dysfunction and sequential apoptosis galangin acts as a potential neuroprotective agent after acute ischemic stroke. Molecules. 2012;17:13403–13423. doi: 10.3390/molecules171113403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu YN, Zha WJ, Ma Y, Chen FF, Zhu W, Ge A, Zeng XN, Huang M. Galangin attenuates airway remodelling by inhibiting TGF-β1-mediated ROS generation and MAPK/Akt phosphorylation in asthma. Sci Rep. 2015;5:11758. doi: 10.1038/srep11758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyer JJ, Afolayan AJ, Taylor MB, Erasmus D. Antiviral activity of galangin isolated from the aerial parts of Helichrysum aureonitens. J Ethnopharmacol. 1997;56:165–169. doi: 10.1016/S0378-8741(97)01514-6. [DOI] [PubMed] [Google Scholar]
- Moynagh PN. TLR signalling and activation of IRFs: revisiting old friends from the NF-κB pathway. Trends Immunol. 2005;26:469–476. doi: 10.1016/j.it.2005.06.009. [DOI] [PubMed] [Google Scholar]
- Qin L, Wu X, Block ML, Liu Y, Breese GR, Hong JS, Knapp DJ, Crews FT. Systemic LPS causes chronic neuroinflammation and progressive neurodegeneration. Glia. 2007;55:453–462. doi: 10.1002/glia.20467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rüweler M, Anker A, Gülden M, Maser E, Seibert H. Inhibition of peroxide-induced radical generation by plant polyphenols in C6 astroglioma cells. Toxicol. In vitro. 2008;22:1377–1381. doi: 10.1016/j.tiv.2008.02.019. [DOI] [PubMed] [Google Scholar]
- Saijo K, Crotti A, Glass CK. Regulation of microglia activation and deactivation by nuclear receptors. Glia. 2013;61:104–111. doi: 10.1002/glia.22423. [DOI] [PubMed] [Google Scholar]
- Sastre M, Dewachter I, Landreth GE, Willson TM, Klockgether T, van Leuven F, Heneka MT. Nonsteroidal anti-inflammatory drugs and peroxisome proliferator-activated receptor-gamma agonists modulate immunostimulated processing of amyloid precursor protein through regulation of beta-secretase. J Neurosci. 2003;23:9796–9804. doi: 10.1523/JNEUROSCI.23-30-09796.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saponaro C, Cianciulli A, Calvello R, Dragone T, Iacobazzi F, Panaro MA. The PI3K/Akt pathway is required for LPS activation of microglial cells. Immunopharmacol Immunotoxicol. 2012;34:858–865. doi: 10.3109/08923973.2012.665461. [DOI] [PubMed] [Google Scholar]
- Sen GC, Sarkar SN. Transcriptional signaling by double-stranded RNA: role of TLR3. Cytokine Growth Factor Rev. 2005;16:1–14. doi: 10.1016/j.cytogfr.2005.01.006. [DOI] [PubMed] [Google Scholar]
- Ulland TK, Wang Y, Colonna M. Regulation of microglial survival and proliferation in health and diseases. Semin Immunol. 2015;27:410–415. doi: 10.1016/j.smim.2016.03.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeng H, Huang P, Wang X, Wu J, Wu M, Huang J. Galangin-induced down-regulation of BACE1 by epigenetic mechanisms in SH-SY5Y cells. Neuroscience. 2015;294:172–181. doi: 10.1016/j.neuroscience.2015.02.054. [DOI] [PubMed] [Google Scholar]
- Zhao XR, Gonzales N, Aronowski J. Pleiotropic role of PPARγ in intracerebral hemorrhage: an intricate system involving Nrf2, RXR, and NF-κB. CNS Neurosci Ther. 2015;21:357–366. doi: 10.1111/cns.12350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zolezzi JM, Santos MJ, Bastías-Candia S, Pinto C, Godoy JA, Inestrosa NC. PPARs in the central nervous system: roles in neurodegeneration and neuroinflammation. Biol Rev Camb Philos Soc. 2017;92:2046–2069. doi: 10.1111/brv.12320. [DOI] [PubMed] [Google Scholar]