

Abstract
Improved understanding of expression of immune checkpoint receptors (ICR) on tumor-infiltrating lymphocytes (TIL) may facilitate more effective immunotherapy in head and neck cancer (HNC) patients. A higher frequency of PD-1+ TIL has been reported in human papillomavirus (HPV)+ HNC patients, despite the role of PD-1 in T cell exhaustion. This discordance led us to hypothesize that the extent of PD-1 expression more accurately defines T cell function and prognostic impact, since PD-1high T cells may be more exhausted than PD-1low T cells and may influence clinical outcome and response to anti-PD-1 immunotherapy. In this study, PD-1 expression was indeed upregulated on HNC patient TIL, and the frequency of these PD-1+ TIL was higher in HPV+ patients (p = 0.006), who nonetheless experienced significantly better clinical outcome. However, PD-1high CD8+ TIL were more frequent in HPV− patients and represented a more dysfunctional subset with compromised IFN-γ secretion. Moreover, HNC patients with higher frequencies of PD-1high CD8+ TIL showed significantly worse disease free survival (DFS) and higher hazard ratio for recurrence (p<0.001), while higher fractions of PD-1low T cells associated with HPV positivity and better outcome. In a murine HPV+ HNC model, anti-PD-1 mAb therapy differentially modulated PD-1high/low populations, and tumor rejection associated with loss of dysfunctional PD-1high CD8+ T cells and a significant increase in PD-1low TIL. Thus, the extent of PD-1 expression on CD8+ TIL provides a potential biomarker for anti-PD-1 based immunotherapy.
Keywords: Immune checkpoint receptors, PD-1, CD8 T cells, Immunotherapy
INTRODUCTION
Immune checkpoint receptor (ICR) blockade has become a major focus of investigation in the field of cancer immunotherapy. Importantly, ICR blockade has shown beneficial results in the clinic for certain patient populations, however biomarkers of response have not been clearly identified. Programmed death-1 (PD-1) (1) and cytotoxic T-lymphocyte antigen-4 (CTLA-4) are two of the main clinical targets (2). While these have been increasingly investigated, less is known about their expression in tumor infiltrating lymphocytes (TIL) during anti-PD-1 therapeutic interventions. Given the promising data that have been reported in several malignancies (3–5), we investigated ICR levels on specific TIL subsets in patients with head and neck squamous cell carcinoma (HNC).
An increasingly important prognostic marker for HNC patients is HPV status, since the percentage of HPV+ oropharyngeal squamous cell carcinoma (OPSCC) in North America has increased from 30% in the 1980s to 80% at present (6). Although it is known that HPV+ HNC has a better prognosis than HPV− HNC, and better response to anti-PD-1 based immunotherapy (7), the mechanism(s) underlying these clinical differences remain elusive, as do differences in immune escape strategies (8). Because of these prognostic differences between HPV+ and HPV− in HNC patients, the distinct immunologic features of the two groups should be compared in order to advance knowledge regarding tumor immune evasion. Therefore, we investigated differences in ICR expression at the mRNA and protein level and characterized immunologic properties of tumor associated T lymphocytes, including CD8+ T effector cells, with respect to HPV status.
Promising results using PD-1 or PD-L1 blocking monoclonal antibodies (mAb) have emerged for advanced recurrent/metastatic HNC (7,9), but still only a minority of patients (15–20%) respond, despite elevated expression of PD-L1 in >50% of HNC patients (8). On the one hand, PD-1 ligation has been demonstrated to have a negative impact on T cells and the blockade of this ligation results in improvement of their function (10). On the other hand, PD-1 positivity has been shown to represent antigen experienced, TA-specific T cells (11) and has been correlated with better clinical outcome (12). Additionally, other checkpoint receptors such as T cell immunoglobulin-3 (Tim-3) (13,14), Lymphocyte activation gene-3 (LAG-3) and B and T lymphocyte attenuator (BTLA) are under investigation. Tim-3 has been identified as a specific marker of fully differentiated IFN-γ producing CD4+ and CD8+ T cells (15). Its expression is regulated by the transcription factor T-bet (16) and negatively regulates Th1 and CD8+ cytotoxic T cell responses (17). LAG-3 is upregulated on activated CD4+ and CD8+ T cells as well as in a subset of activated natural killer (NK) cells (18). Beside its capacity to bind to major histocompatibility complexes (MHC) class II and its role in T helper cell and regulatory T cell (Treg) signaling, direct inhibitory effects of LAG-3 on CD8+ T effector cells have been demonstrated (19). BTLA negatively regulates T cell activation by inhibiting T cell proliferation and cytokine production. In contrast to other checkpoint receptors, BTLA is expressed on naïve T cells and only transiently upregulated upon TCR engagement (17). Thus, BTLA is downregulated on highly activated T cells (20).
Little is known about the role of checkpoint receptors in TIL from HPV+ vs. HPV− HNC. Interestingly a prior report suggested that the presence of PD-1 T cells in HPV+ patients was associated with a beneficial effect on survival (12). Given that HPV+ patients demonstrate a much better outcome in the clinic (12), we hypothesized that intratumoral ICR expression and function might provide immunologic insight into this difference in prognosis. Thus, as opposed to solely analyzing the frequency of PD-1+ vs. PD-1− TIL, we investigated whether the intensity of PD-1 expression on TIL influences clinical outcome, providing a potential prognostic biomarker and for monitoring response to anti-PD-1 based immunotherapy.
Materials and Methods
TCGA data retrieval and analysis
RNAseq data from queried genes were downloaded from the UCSC cancer genomics browser (https://genome-cancer.ucsc.edu). The HNC gene expression profile from 500 HNC specimens was measured experimentally (21). The RSEM units to quantitate RNAseq expression data were described previously (22). Correlations from TCGA data were calculated using Pearson r test and linear regression curve fits were graphed using GraphPad PRISM software v6.
Patients and specimens
Peripheral venous blood samples and tumors were obtained from HNC patients seen in the Department of Otorhinolaryngology at the University of Pittsburgh Medical Center. No active patient exclusion to clinical stage or HPV status was performed other than restriction to all consented, surgically treated patients with primary tumors and curative treatment intention. Specimen acquisition was continuously performed by the same physician during 2011–2013, the patients’ HPV-status was defined from surgical specimen by immunohistochemical (IHC) determination of the p16 status (positive p16 expression was defined as strong and diffuse nuclear and cytoplasmic staining in at least 70% of the tumor cells). All subjects signed written informed consent approved by the Institutional Review Board of the University of Pittsburgh, patient studies were conducted in accordance with the Declaration of Helsinki.
Collection of PBMC and TIL
Blood samples and tumor specimen were collected at the day of and prior to surgery and therefore, at the beginning of the treatment. Blood samples from cancer patients (30–40 ml) were drawn into heparinized tubes and centrifuged on Ficoll–Hypaque gradients (GE Healthcare Bioscience, Piscataway, NJ, USA). Peripheral blood mononuclear cells (PBMC) were recovered, washed in RPMI-1640 (Invitrogen, Carlsbad, CA, USA) and immediately used for experiments or stored at −80 °C until further analysis. For TIL isolation, freshly isolated tumors from HNC patients were minced into small pieces in RPMI-1640 (Invitrogen, Carlsbad, CA, USA), which then were transferred to a cell strainer (70 mm Nylon) and mechanically separated using a syringe plunge. The cells passing through the cell strainer were collected and subjected to Ficoll–Hypaque gradient centrifugation. After centrifugation, mononuclear cells were recovered and stored at −80 °C until flow cytometry analysis.
Quantitative polymerase chain reaction (qPCR) of ICR
qPCR was performed to investigate tumor specimen from 20 HPV+ and 20 HPV− HNC patients. Acquisition of patient material was separately performed during 2007–2011, following the consistent standards as described in patients and samples. RNA was extracted using the RNeasy Kit (Qiagen, Valencia, CA, USA) following the manufacturer’s protocol. RNA concentration was determined by spectrophotometer measurement (NanoVue Plus, GE Healthcare, Piscataway, New Jersey, USA). Random hexamers and MultiScribe reverse transcriptase (Applied Biosystems, Foster City, CA) were used for first strand cDNA synthesis according to manufacturer’s instructions. The samples were added to a 20-μL reaction using 2X Taqman master mix (Applied Biosystems) and analyzed on a StepOne™ Real-Time PCR System (Applied Biosystems) using the following PCR probes: Hs00169472_m1 for PD-1, Hs00158563_m1 for LAG-3, Hs00262170_m1 for TIM-3, Hs03044418_m1 for CTLA-4, Hs99999905_m1for GAPDH as a reference gene (all Life Technologies, Grand Island, NY, USA).
Flow cytometry
For cell surface staining, PBMCs and TIL were washed twice in staining buffer (2% w/v fetal bovine serum) and stained for cell surface markers. Cells were incubated with relevant antibodies for 30 min at 4°C in the dark, washed twice and re-suspended in staining buffer. Flow cytometry was performed using a CyAn flow cytometer (Dako, Ft. Collins, CO, USA) and Fortessa cytometry (Becton Dickinson, Franklin Lakes, NJ, USA) and data analyzed using FlowJo software (TreeStar, Inc., Ashland, OR, USA). The acquisition and analysis gates were restricted to the lymphocyte gate based on characteristic properties of the cells in the forward and side scatter.
Antibodies and reagents
The following anti-human monoclonal antibodies (mAb) were used for staining: PD-1-APC (eBioscience, San Diego, CA, USA), CD3-PerCP-Cy5.5, CTLA-4-PE, PD-1-PerCP Cy5.5 (all Biolegend, San Diego, CA, USA), CCR7-FITC (R&D Systems Inc, Minneapolis, MN, USA) CD3-Alexa Fluor 405, CD4-PE-Texas Red, CD-8-PE-TR, CD45RA-PE-TR (all Life Technologies, Grand Island, NY, USA), CD3-APC-Cy7, CD8-APC, CD8-PE-Cy7 (all BD Biosciences, San Jose, CA, USA), granzyme B FITC (clone GB11, Biolegend) including their respective isotypes, which served as negative controls for surface as well as intracellular staining. Antibodies were pre-titrated using activated as well as non-activated PBMC to determine optimal staining dilutions. Viability was assessed by Zombie aqua (Biolegend)
Identification of PD-1 subsets
Isolated T cells of healthy donors were stimulated for 5 days with CD3/CD28 beads. CD3+ CD8+ T cells were analyzed by flow cytometry for expression of PD-1 as demonstrated in supplemental figure S1. Accordingly, TIL were analyzed by flow cytometry for CD8+ PD-1+ cells in reference to isotype controls. Ranging from highest detected PD-1 expression to the lower limit of PD-1+, the gated PD-1+ cells were divided into cells with high PD-1 (PD-1high), intermediate (PD-1int) and low (PD-1low) expression levels.
Granzyme B expression
Isolated TIL (n=3 HNC patients) were stained for PD-1 expression of CD3+ CD8+ T cells and then compared for granzyme B positivity of PD-1negative, PD-1low, PD-1int and PD-1high subgroups.
Sorting of TIL subsets and IFN-γ ELISPOT
After TIL isolation from HNC tumors (n = 3), T cells were purified using EasySep™ Human T Cell Enrichment Kit (Stemcell technologies, Vancouver, Canada). After staining, CD8+ T cells were sorted using MoFlo Astrios (Beckman Coulter, Brea, CA, USA) in PD-1 negative, PD-1 low-intermediate and PD-1 high groups. Cells were rested overnight in human serum and compared for functional differences in an IFN-γ ELISPOT assay. Therefore, MultiScreen-IP filter plates (Millipore, Billerica, MA) were coated overnight with anti-human IFN-γ mAb 1-D1K (Mabtech, Mariemont, OH) (10 μg/mL in PBS) at 4°C. After washing with PBS, plates were blocked for 1 h at 37°C with 10% human serum in RPMI. Sorted cells were added to wells in duplicates (5×103) and stimulated with anti-CD3/CD28 beads (bead: cell = 5:1). After incubation for 18 hours at 37°C, plates were washed with PBS/0.5% Tween 20, and incubated with biotinylated anti-IFN-γ mAb (Mabtech) (2 μg/ml in PBS/0.5% BSA) for 2 hours at 37°C. Plates were washed again and incubated with Streptavidin-HRP (Mabtech, 1:500 in PBS/0.5% BSA) for 1 hour at 37°C. After washing, TMB substrate solution (Vector Laboratories, Inc. Burlingame, CA) was added. Color development was stopped by extensive washing in tap water after 5 minutes. Plates were dried and spots were counted using CTL ImmunoSpot® Analyzer (CTL, Shaker Heights, OH) and evaluated by CTL Professional Double Color Software.
Murine HNC model
All animal experiments were performed following institutional guidelines for animal experimentation. The murine in vivo HNC model was established as previously described by Hoover et al (23). Briefly, C57BL/6 mice were obtained from the Jackson Laboratory, Bar Harbor, ME (age 1–2 months). Immortalized, E6/E7 plus H-ras transduced mouse tonsil epithelial cells (MTEC) were obtained from Hoover (23). After subcutaneous injection of 1×106 cells into the neck of the mice, 33 mice with tumor growth were randomized and assigned into different treatment groups. The purpose was to compare the radiation therapy (RT) + anti-PD-1 mAb therapy group (n=17) to the RT + isotype control (clone MOPC-21, mouse IgG1k, BioXcell) group (n=16). Anti-PD-1 mAb (clone 4H2, IgG1 isotype) was obtained from Bristol-Myers Squibb, New York, NY (24). Anti-PD-1 therapy (3mg/kg body weight) was administered on days 1, 4, 11 and 15. Radiotherapy was administered to induce PD-L1 expression (25) in the tumor microenvironment in order to enlighten potential protective effects of anti-PD-1 mAb therapy. In addition, RT increases tumor death, phagocytosis and antigen presentation. Fractionated radiotherapy was administered for 5 consecutive days at 2Gy/d during week 1 (D4-D8) and week 2 (D11-D15). Treatment response was analyzed by measurement of the tumor volume at intervals between 3 and 18 days. Additionally, mice spleen were collected and processed as described for lymphocyte isolation. PD-1high and PD-1int/low fractions were compared from CD3+ CD8+ PD-1+ cells and analyzed according to different treatment groups. Different clones for treatment (clone 4H2) and staining (BV421 conjugated anti-PD-1 Ab, IgG2a isotype, clone 29F.1A12) were utilized.
Statistics
Differences in phenotypic counts by HPV status were calculated by Wilcoxon signed rank test for paired data or the Wilcoxon test for independent groups. PD-1+ TIL were divided by flow cytometry into subsets of the proportions of cells classified as PD-1high, PD-1int and PD-1low. We investigated the impact of the proportions of PD-1high and PD-1low in a cohort of 56 HNC patients having curative resection and followed up for disease recurrence. We used Cox proportional hazards models and checked models for linearity and for adequacy of the proportional hazards assumptions. As a heuristic device Kaplan – Meier plots of time to recurrence were prepared by dividing proportions of PD-1high and PD-1low cells into tertiles. For the murine experiment, tumor volumes of day 18 were compared. By modeling the fixed and random coefficients of a polynomial regression model, we compared the slopes between the two treatment groups. Data represent observed mean with bootstrap 95% CI.
Study approval
Approval for study of specimens from informed and consented HNC patients was obtained by the institutional review board of University of Pittsburgh (#99-069). Study approval for animal experiments was obtained by the University of Pittsburgh (IACUC# IS00005860).
RESULTS
Expression of PD-1 and CD8A, CD8B is significantly higher in HPV+ than in HPV− tumor specimens
To investigate the quantitative expression of ICR in HNC patients, a cohort of 20 HPV+ and 20 HPV− tumors was used for analysis by qRT-PCR. ICR expression was normalized to the average level of pharyngeal mucosa of healthy donors (n=4, figure 1A). Immune checkpoint receptors show highly variable expression for HPV+ and HPV− tumors in qPCR in relation to reference tissue. In order to confirm and further investigate these results in a larger cohort, ICR expression data from TCGA was retrieved and mRNA expression of the ICR shown in figures 1B–D analyzed. We found that expression of all ICRs and CD8A, CD8B and CD4 were higher in HPV+ tumors than in HPV− ones. However, more importantly was the finding that PD-1 was the only ICR that was significantly higher in HPV+ tumors along with CD8 T cells markers: CD8A and CD8B (figures 1E–F). In addition to comparing expression data from the two cohort of patients, we correlated the expression of PD-1, CTLA-4, TIM-3 and LAG-3 with that of CD8A, given that this marker of CD8 T cells was highly upregulated in HNC tumors, especially in HPV+ samples. We found that all ICRs queried highly correlated with CD8A expression regardless HPV status. However, HPV+ tumors showed a higher correlation coefficient than HPV− samples for all ICRs (figure 1G). Interestingly, PD-1 expression was highly correlated with that of CD8A in both HPV+ and HPV− tumors, with CTLA-4 being the other ICR have a similar correlation coefficient in HPV+ tumors (figures 1G–H). Since mRNA expression of ICRs was significantly higher in tumors vs. control mucosa, and since PD-1 was a highly expressed gene common to both HPV+ and HPV− tumors and highly correlated with CD8A expression, we decided to further investigate protein expression of these ICRs by flow cytometry in different subsets of T cells in PBL and TIL form HNC patients.
Fig. 1. Expression of PD-1 and CD8A, CD8B is significantly higher in HPV+ than in HPV− tumor specimens.
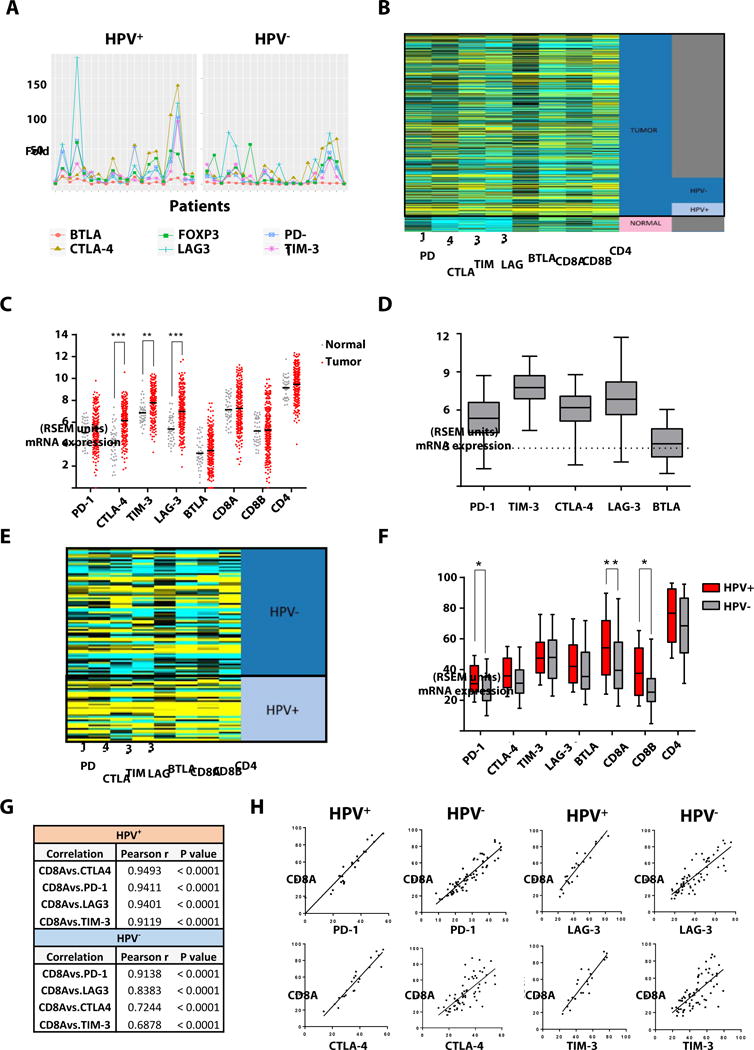
(A) QRT-PCR from clinical HNC tumor samples (n=40) was performed for several ICR: PD-1, CTLA-4, LAG-3, TIM-3, BTLA and the Treg transcription factor FOXP3. As a reference, the fold change of average expression of each ICR from healthy donor pharynx mucosa tissues (n=4) was utilized. Immune checkpoint receptors show highly variable expression for HPV+ and HPV− tumors in qPCR in relation to reference tissue. (B–D) Additionally, ICR mRNA expression data retrieved from TCGA was analyzed and compared to healthy donor mucosa. Expression of all ICRs and CD8A, CD8B and CD4 were higher in tumors as compared to normal TCGA reference mucosa. (E–F) The analysis of the TCGA database revealed that PD-1 was the only ICR that was significantly higher in HPV + tumors along with CD8 T cells markers: CD8A and CD8B. All ICRs highly correlated with CD8A expression regardless of HPV status. However, HPV+ tumors showed a higher correlation coefficient than HPV− ones for all ICRs (G). PD-1 expression highly correlated with that of CD8A in both HPV+ and HPV− tumors, CTLA-4 being the other ICR with a slightly higher correlation coefficient in HPV+ tumors (G, H).
PD-1 is predominantly expressed by CD8+ TEM and tumor-associated antigen-specific CD8+ T cells in the circulation of HPV+ and HPV− head and neck cancer patients
In order to consider inter-individual ICR expression of HPV+ and HPV− HNC patients’ cellular subsets, we analyzed CD8+ PBL for expression of PD-1 and CTLA-4, the best characterized ICR, in respect to traditional phenotypic markers (26). We investigated the proportions of native and memory T cells of HNC patients based on CCR7 (lymphoid tissue homing receptor) and CD45RA (a transmembrane tyrosine phosphatase) expression (figure 2A). As shown in figure 2B, the frequency of effector memory (TEM: CCR7−CD45RA−) was significantly higher than naive (CCR7+CD45RA+), central memory (TCM: CCR7+CD45RA−), and terminal effectors (TEMRA: CCR7−CD45RA+) in both HPV+ and HPV− patients (p < 0.001). The frequency of TEM was comparable between HPV+ and HPV− patients. Approximately 20% of TEM expressed PD-1, which was significantly higher than that expressed by naïve T cells in both HPV+ and HPV− patients (figure 2C, p<0.05 and p<0.01 respectively). In HPV− patients, PD-1+ cells in TCM were significantly higher than in naïve T cells (p<0.05). PD-1 single positive (PD-1+) T cells from both HPV+ and HPV− patients were significantly more frequent than double positive (PD-1+CTLA-4+) T cells or CTLA-4 single positive (CTLA-4+) T cells, indicating that PD-1 was more frequently expressed by T cells in peripheral blood, than CTLA-4 (p<0.001, figure 2D). However, in peripheral blood, the frequencies of PD-1+ T cells between HPV+ and HPV− patients were similar. Taken together, these results indicate that PD-1 rather than CTLA-4 is frequently expressed by CD8+ effector T cells in both HPV+ and HPV− patients. These results led us to investigate tumor infiltrating lymphocytes (TIL) by flow cytometry from these patient subgroups.
Fig. 2. PD-1 is predominantly expressed on antigen experienced peripheral TEM cells in both HPV+ and HPV− HNC patients.
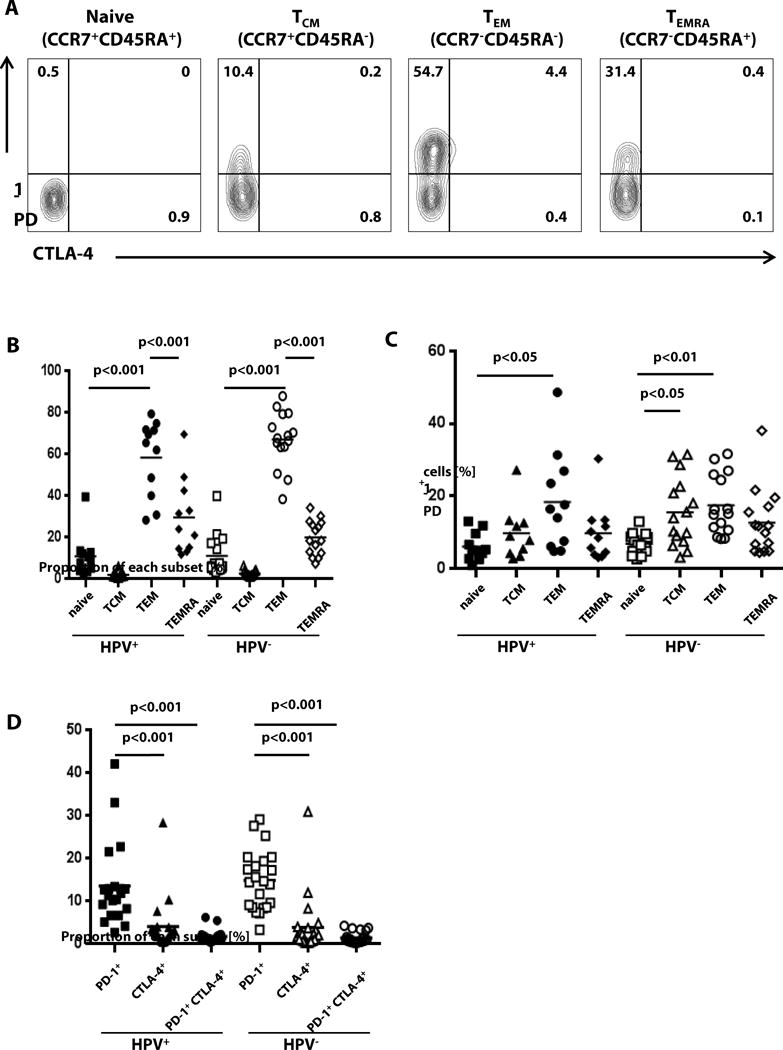
(A) PBL from HNC patients were analyzed for co-expression of PD-1 and CTLA-4 and phenotypic markers: effector memory (TEM: CCR7−CD45RA−) naive (CCR7+CD45RA+), central memory (TCM: CCR7+CD45RA−), and terminal effectors (TEMRA: CCR7−CD45RA+). (B) Comparison of phenotypic proportions between HPV+ and HPV− T cells (one-way ANOVA, Bonferroni’s mult. comparison test, HPV+ n = 11, HPV− n = 15). (C) Percentage of PD-1+ T cells were measured by flow cytometry depending on phenotype and HPV status (one-way ANOVA, Bonferroni’s multiple comparison test, HPV+ n = 11, HPV− n = 15). (D) Comparison of single positive (PD-1 or CTLA-4) and double positive T cells in PBL (one-way ANOVA, Bonferroni’s multiple comparison test, HPV+ n = 20, HPV− n = 22).
Significantly higher frequencies of PD-1+ CD8+ TIL in HPV+ HNC patients’ tumors
To evaluate the relevance of ICR in effector cells infiltrating HPV+ and HPV− HNC, we compared CTLA-4 and PD-1 expression on CD8+ TIL isolated from both HPV+ and HPV− patient tumors. As shown in figure 3A, the frequencies of CTLA-4+ and PD-1+ cells were notably higher in CD8+ TIL for HPV+ and HPV− patients compared to those on CD8+ PBL. Most notably, the frequency of PD-1+ cells of CD8+ TIL was significantly higher in HPV+ patients than that in HPV− patients (p<0.006, figure 3B). PD-1 positivity was defined by isotype controls through flow cytometry as shown in figure 4A.
Fig. 3. HPV+ HNSCC patient TIL demonstrate higher overall expression of PD-1.
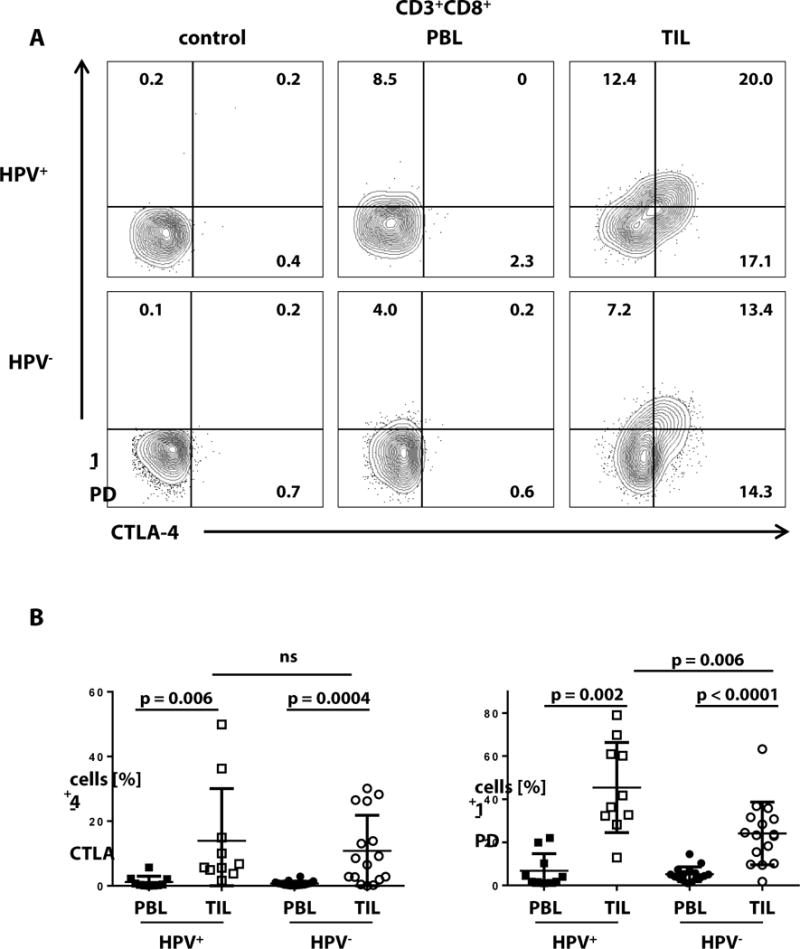
The frequency of CTLA-4+and PD-1+ cells was compared by flow cytometry in CD8+ TIL and PBL from HPV+ and HPV− patients. (A) Representative gating and stratification and sorting of TIL by extent of PD-1 level of expression. (B) Comparison of CTLA-4+ and PD-1+ in PBMC and TIL of HPV+ and HPV− (matched-pair Wilcoxon test for comparison of TIL vs. PBL, Mann-Whitney test for comparison HPV+ vs. HPV− TIL, HPV+ n = 10, HPV− n = 16).
Fig. 4. HPV− tumors contain significantly more PD-1high CD8+ TIL that manifest an exhaustion phenotype and produce little IFN-γ.
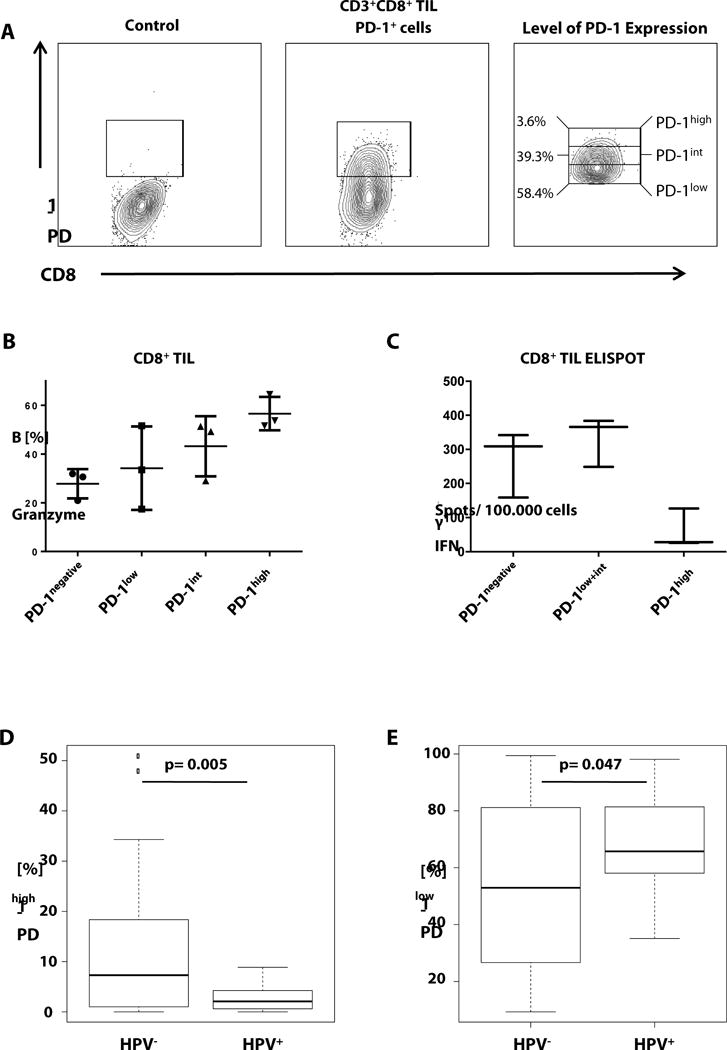
(A) The range from highest detected PD-1 to the negative control (PD-1negative) was divided into the three subgroups with low (PD-1low), intermediate (PD-1int) and high (PD-1high) PD-1 expressing cells. (B) Flow cytometric staining for CD3+, CD8+, and granzyme B on representative TIL (n=3, two HPV+ and one HPV−), gated on PD-1negative, PD-1low, PD-1int and PD-1high (n = 3, one-way ANOVA, p = 0.03). (C) ELISPOT analysis for IFN-γ secretion per 100,000 cells of CD8+ TIL (n=3, two HPV+ and one HPV−, distinct tumor samples from B), sorted into PD-1negative, PD-1low+int and PD-1high expressing subsets (n = 3, one-way ANOVA p = 0.01). (D, E) Box and Whiskers plots for PD-1high (D) and PD-1low (E) fractions of HPV+ and HPV− patients (Mann-Whitney comparison, HPV+ n = 20, HPV− n = 36).
HPV− tumors contain significantly more PD-1high CD8+ TIL, which are functionally impaired
Others have suggested that higher numbers of intratumoral PD-1+ T cells may correlate with better prognosis in HPV+ HNC (12), and the role of HPV in HNC patients’ response to anti-PD-1 therapy is controversial. Furthermore, Wherry has shown that the highest level of PD-1 expression reflects T cell exhaustion status (27). Thus, based on positive PD-1 expression, CD8+ TIL were divided into three subgroups, from highest PD-1 expressing cells (PD-1high), intermediate (PD-1int) to lowest PD-1 positive cells (PD-1low) (figure 4A). Definition of subgroups was confirmed with PD-1+ subpopulations of CD3/CD28 bead-stimulated healthy donor CD8+ T cells (supplementary figure 1). We previously showed that PD-1 expression level correlates with higher SHP-2 phosphatase, more potent dysfunction and impaired Th-1 signaling (28). Thus, we investigated the activation status and functional differences in subsets of PD-1 negative, PD-1low, PD-1int and PD-1high T cells, taken directly from freshly excised HNC tumors. As a marker of activation, granzyme B expression in each of the subsets was analyzed, indicating the highest activation for PD-1high subsets (figure 4B), enriched in HNC patient TIL (29). To analyze functional capabilities of these groups, sorted T cell populations of negative, moderate (PD-1low+PD-1int) or high (PD-1high) PD-1 expression were compared for their IFN-γ secretion capacity, after TCR stimulation using CD3/CD28 beads. ELISPOT analysis revealed significantly compromised IFN-γ secretion only in PD-1high CD8+ T cells, as compared to robust IFN-γ secretion by PD-1 negative and moderate PD-1(low/int) T cells (figure 4C). HPV+ and HPV− patients were compared based on the proportions of PD-1high and PD-1low TIL subsets. The fraction of PD-1high T-cells was greater in HPV− patients (7.3% vs. 2.1% of PD-1high cells, p = 0.0005 figure 4D). In contrast, the proportion of PD-1low expressing T cells was higher in HPV+ patients than in HPV− patients (65.7% vs. 53.0% p = 0.047, figure 4E).
PD-1high cells are associated with worse disease free survival
Based on the higher frequency of PD-1high T cells in HPV− patients, we investigated the impact of the extent of PD-1 expression by CD8+ T cells on time to disease recurrence. A cohort of HNC patients (n = 56, 36 HPV− and 20 HPV+) all of whom had curative therapy for locally advanced disease, were followed for disease recurrence. With a median follow up 19 months, 40 patients remained free of disease and 16 recurrences were observed, yielding a 2 year probability of recurrence-free survival of 70%, which is typical for our institution (30). A proportional hazards regression analysis revealed that the proportion of PD-1high CD8+ T cells potently increased the risk of disease recurrence in this cohort of HNC patients (hazard ratio = 2.15, 95% CI = 1.46 – 3.15, p < .0001). To validate this finding we examined the effect of the proportion of PD-1low cells upon recurrence and found the opposite impact. Indeed, greater fractions of PD-1low CD8+ T cells were protective of recurrence (hazard ratio = 0.19, 95% CI = .07 – .49, p = .0006). Figures 5A–D illustrate these findings showing Kaplan–Meier curves arbitrarily divided into tertiles of the proportions of PD-1high (figure 5A) and predicted log hazard ratio (figure 5B) and Kaplan–Meier curves arbitrarily divided into tertiles of the proportions of PD-1low (figure 5C) and predicted log hazard ratio (figure 5D).
Fig. 5. PD-1high TIL are associated with worse DFS whereas high levels of PD-1low TIL portend better clinical outcome.
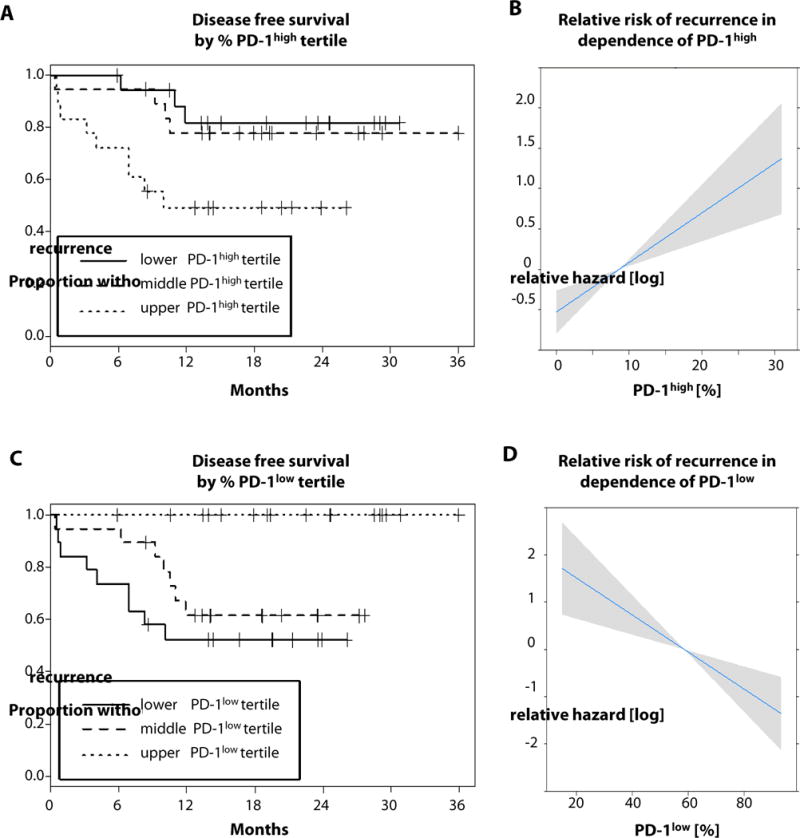
Kaplan-Meier curves portray DFS for HNC patients (HPV+ n=20, HPV− n=36) in relation to PD-1high vs. PD-1low fractions of CD8+ TIL. Tertiles for PD-1high fractions were distinguished and survival plotted as shown (A, ranges of lower tertile 0–1.75; middle tertile 1.75–8.7; upper tertile 8.7–51). Cox proportional hazards models were used to investigate the relative risk and models checked for linearity and adequacy of the proportional hazards assumptions. (B) Hazard ratio = 2.25 (95% CI = 1.46 – 3.15), p < .0001. Tertiles for PD-1low fractions were distinguished and survival plotted as shown (C, ranges of lower tertile 9.3–49.5; middle tertile 49.5–78.5; upper tertile 78.5–99.4). (D) Hazard ratio = 0.19 (95% CI = .07 – .49, p < .0006).
PD-1high CD8+ T cells are dramatically reduced in the setting of best treatment response to anti-PD-1 therapy
To investigate if PD-1 targeted Ab therapy influences PD-1 T cell fractions and how therapeutic modalities influence PD-1 expression levels, we used a murine HNC model investigating treatment response to anti-PD-1 mAb or its isotype in combination with radiation therapy (RT). Figure 6A demonstrates the development of tumor volume for the different therapeutic groups. As expected, better treatment response (i.e. lowest tumor volumes at day 30) was observed in the combination therapy with anti-PD-1 mAb. Moreover, anti-PD-1 mAb therapy plus RT significantly reduced the fraction of PD-1high CD8+ T cells and enhanced the frequency of PD-1low cells (figures 6B+C), in association with the best treatment response. This beneficial effect reversed the enrichment of PD-1high fractions that was observed in the RT + isotype control mAb group. This result demonstrates a valuable effect of anti-PD-1 mAb therapy, by dramatically reducing the poor prognostic subpopulation of PD-1high TIL cells, and permitting potent anti-tumor activity to develop via PD-1low/int, IFN-γ positive T cells.
Fig. 6. Dysfunctional PD-1high CD8+ T cells are dramatically reduced in the treatment response group to anti-PD-1 plus RT targeted therapy.
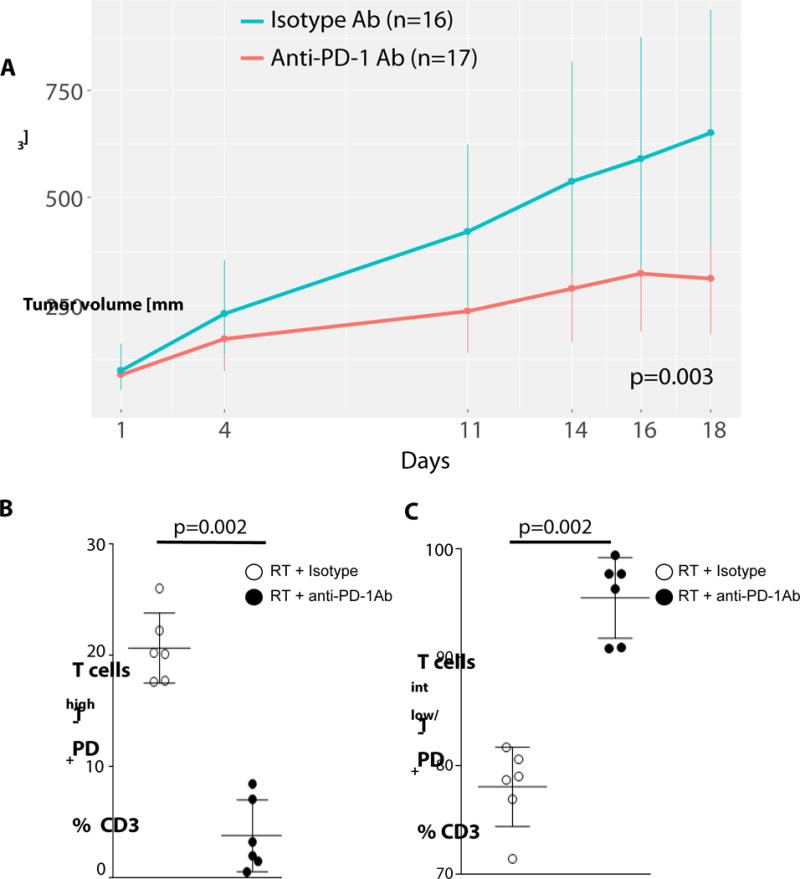
(A) 33 mice with positive tumor growth were randomized into two treatment groups (n= 16 for isotype + RT, and n=17 for anti-PD-1 Ab + RT). Treatment response was analyzed by measurement of the tumor volume on different days. A comparison of radiotherapy alone (isotype) and radiotherapy in combination with anti-PD-1 Ab treatment was performed. By modeling the fixed and random coefficients of a polynomial regression model, we conclude that the slopes between two treatment groups differ and that RT + anti-PD-1 Ab treated tumors grew more slowly than RT + isotype treated tumors. Data represent observed mean with bootstrap 95% CI (p = .0039). Anti-PD-1 Ab or isotype control (3mg/kg body weight) was administered at days 1, 4, 11, 15; radiotherapy was fractionated in 2 Gy ×10 days (on day 4,5,6,7,8,11,12,13,14,15). (B, C) PD-1 expression of isolated splenic CD3+ PD-1+ lymphocytes, PD-1high (B) and PD-1int/low fractions (C) of RT + isotype vs. RT + anti-PD-1 Ab were compared with non-parametric Mann-Whitney test, data are representative of +/− SEM.
DISCUSSION
In peripheral blood, we observed that PD-1 is mainly expressed on effector T cells rather than by naïve T cell populations. Despite an overall higher PD-1+ CD8+ T cell frequency in the TIL of HPV+ HNC patients, HPV− patients had larger fractions of PD-1high CD8+ T cells. We demonstrate that these PD-1high expressing CTL were highly activated (expressing the most granzyme B), but showed a severely dysfunctional phenotype, with compromised IFN-γ secretion capacity and negative prognostic impact in the clinic. In a murine PD-1 targeting immunotherapy model, we were able to observe that anti-PD-1 therapy overcomes the increase of PD-1high expressing cells that could be observed in mice treated with RT alone, which drives PD-L1 levels within 24 hrs. of treatment (25). Furthermore, in contrast to PD-1high CD8+ T cells, frequencies of PD-1low/int T cells were associated with a better DFS in our cohort of HNC patients. These beneficial, activated T cells were significantly increased upon anti-PD-1 mAb therapy in the murine HNC model, correlating with clinical efficacy.
Clinical trials targeting ICR have shown substantial results for multiple cancer subsets including melanoma, renal cell carcinoma, non-small cell lung cancer (31,32) and recently HNC (7). Nevertheless, the knowledge about ICR in HNC tumors and TIL is limited, but is of interest due to emerging immunotherapeutic targeting of these molecules in several cancer types (33). While cancer immunotherapy targeting ICR has emerged as a major therapeutic advance (34), biomarkers of response are inadequate and have so far focused on ligand expression (i.e. PD-L1) by tumor or immune cells. Here, we propose that response to anti-PD-1 mAb therapy may be influenced by levels of PD-1 on effector T cells in the circulation or the microenvironment. The effect we observe here may reflect salvage and protection of activated functional T cells during tumor response to anti-PD-1 therapy.
Upon ligation, PD-1 directly inhibits TCR-mediated effector functions and thereby, causes T cell dysfunction (35). Despite the immune-inhibitory pathways that are activated upon PD-1 engagement, reports of PD-1 positivity in TIL and its prognostic impact have been controversial to date. PD-1 expression reflects T cell activation but means only that a T cell is receptive to an immunoinhibitory signal, not that the T cell is necessarily exhausted and dysfunctional. Our results demonstrate that part of this controversy might be attributed to the limitations of grouping PD-1 positive cells despite heterogeneous functional capacity. Indeed, we show that PD-1high vs. PD-1low T cells possess distinct and important functional differences with significant prognostic importance.
In a recent meta-analysis of 29 studies investigating PD-1 expression and overall survival (OS) in patients with epithelial malignancies, PD-1 expression by TIL was associated with a shorter OS (36). In contrast, Badoual and colleagues reported PD-1 expressing tumor-infiltrating T cells as a favorable prognostic marker in HPV+ HNC (12). This conundrum intrigued us due to the potential for the level of PD-1 expression to more accurately mark different TIL subsets with unique behavior. Also distinct etiologies of HNC such as HPV+/− disease, with such dramatically different prognoses (37), provided an unparalleled window on this potential biomarker. HPV+ HNC represents a unique cancer subset (38), given its location in lymphoepithelial tissue with specific immunological properties (39), virally associated immune evasion properties (40) and yet better clinical prognosis than non-HPV associated HNC (37). Therefore, we investigated immunological properties like ICR expression of both patient subgroups in antigen specific cells from peripheral blood and tumor tissue. Our findings in CD8+ TIL confirm an overall increased number of PD-1+ T cells for HPV+ patients. Most importantly, when investigating the extent of PD-1 expression in PD-1+ cells, we observed significant differences between HPV+ and HPV− patients. HPV+ patients had a significant lower fraction of the PD-1high expressing cells in comparison to the HPV− patients. In contrast, the PD-1low fraction was significantly elevated in HPV+ patients in comparison to HPV− patients. In combination with the shown functional differences of PD-1low and PD-1high CD8+ TIL, these results might contribute to a better understanding of prognostic differences between HPV+ and HPV− patients, and additionally, account for correlations of PD-1+ cells and favorable prognostics. Therefore, our analysis of DFS focused on the impact of fractions of PD-1high and PD-1low CD8+ TIL.
Here, we show that the patients with higher PD-1high fractions had a significantly higher risk of recurrence (2.25 fold increased risk as this fraction of cells increased). On the contrary, high PD-1low fractions were associated with improved DFS. These data support an important role for T cell exhaustion in progression of disease, with the possibility that rescue of the PD-1low/int cells represents the mechanism of clinical benefit as well as a more accurate biomarker of clinical response to anti-PD-1/PD-L1 targeting immunotherapy. In the setting of chronic viral infection, Blackburn and colleagues were able to identify a subset of exhausted T cells, which can be rescued through anti-PD-1 targeting therapy (10). The data from our murine HPV+ model support these observations in the case of cancer associated T cell exhaustion as demonstrated in treatment groups with anti-PD-1 treatment, which showed an increase of PD-1low expressing T cell fractions. Moreover, radiotherapy treatment response was significantly increased through the addition of PD-1 targeting immunotherapy.
Our data show that the effector memory cell subset of HNC patients more highly express PD-1. As PD-1 has been shown to negatively regulate T cell phenotype, proliferation and instead induces apoptosis (13,14,28), the combination of our observations linking PD-1 and peripheral effector memory subsets in vivo supports the association of PD-1 with clonally expanded, tumor-reactive populations (11). In this situation ligation of PD-1 by its ligands is likely, and even more so for the PD-1high expressing populations. This may explain the vulnerability and functional insufficiency of antigen-specific T cell expansion in the tumor microenvironment. Indeed, Fourcade and colleagues report inverse correlations of expansion and PD-1 levels in a melanoma vaccine study (41). In our study, we observed a protective effect from these negative regulatory pathways in case of anti-PD-1 treatment, as demonstrated by the significant increase of the PD-1low/int fractions in the murine anti-PD1 therapy model. The antitumor activity of the PD-1low/int populations is also supported by the improved treatment response in this group. Additionally, analyzed CD8+ T cells were obtained from splenocytes which represent circulating T cells. Therefore, in our point of view, the increase in PD-1low fractions represents more likely a clonal expansion of PD-1+ anti-tumor effective T cell populations, perhaps detectable in the circulation of anti-PD-1 treated patients.
By separating different extent of PD-1 expression levels in CD8+ TIL, we observed that cells of high PD-1 expression levels possess a severely impaired IFN-γ secretion capacity, in contrast to PD-1low and PD-1int T cell fractions. This is supported by the fact that in freshly isolated TIL, the PD-1neg groups secrete lower IFN-γ after stimulation in the ELISPOT assays, with increasing differences of IFN between PD-1low/int and PD-1high after 18hr of stimulation during ELISPOT. These results are in congruence with reports of Flecken et al, who showed reduced IFN-γ secretion after expansion of tumor-associated antigen-specific CD8+ T-cells (42). This impairment did not resolve after Treg depletion and supports direct involvement of PD-1 ligation and thereby downregulation of functional cytokine production. A study of Zhang and colleagues identified alteration of IFN-γ levels depending on PD-1 polymorphisms, guiding to a direct involvement of PD-1 receptor pathways on IFN-γ production (43). Additionally, PD-1 blockade has been associated to an increased IFN-γ production and elevation of IFN-γ dependent associated chemokines (44). These relationships may contribute to difficulties in separating reasons for - and effects of - both PD-1 and PD-L1 expression. PD-1 positive cells have been associated with high TA specificity. The release of cytokines like IFN-y of these TA specific cells in the tumor microenvironment has been shown to induce PD-L1 with negative impact on function and cytokine production of PD-1+ cells. Therefore, a more sophisticated view of different PD-1 expression levels is needed.
In their analysis of tissue specific differences in PD-1 expression of T cells, Blackburn and colleagues describe that their ability to degranulate is dependent on the PD-1 expression levels, independent of anatomical location and thereby, PD-L1 expression (45). This is supported by our findings of higher granzyme B positivity for PD-1high expressing TIL. Therefore, the consideration of different PD-1 expression levels exemplifies the two disparate interpretations of PD-1 expression as a marker of activated, competent tumor reactive T cells on the one hand, and PD-1high expression as a marker of exhausted, dysfunctional cells with a negative influence in the tumor microenvironment on the other hand.
Our results are supported by the fact that anti-PD-1 therapy has been reported to be effective in HPV− than HPV+ patients (46,47). Thus, it is possible that higher frequencies of PD-1high T cells from HPV− patients may contribute to worse prognosis, as well as to a good response to anti-PD-1 immunotherapy. In consequence, we demonstrate that, PD-1 expression in CD8+ TIL of HNC represents (over-) activation and - at highest expression levels - exhaustion of effector CD8+ T cells that is associated with better clinical outcome. Thereby, we conclude two essential consequences for immunotherapy: first, low/intermediate but positive PD-1 expression levels are favorable in terms tumor specific functionality and prognosis and second, these data support that PD-1 blockade may allow the expansion of TA specific T cells, keeping their PD-1 expression at lower levels with enhanced anti-tumor response and reduced susceptibility to PD-L1 ligation, indicating a possible predictive role of the level of PD-1 expression on TIL. Whether this effect is due to active restoration of proliferation of PD-1high cells or enhanced proliferation and responses of PD-1low cells has to be investigated. Further studies must validate our finding using anti-PD-1 mAb treated cancer patients including HNC. Baseline PD-1high vs. PD-1low levels should be measured and correlated with responder status.
Supplementary Material
Acknowledgments
We would like to thank Bratislav Janjic and Michael Meyers from the Flow Cytometry Facility for excellent technical assistance.
Financial support: This work was supported by National Institute of Health grants R01 CA206517, DE019727, P50 CA097190, T32 CA060397 (R. L. Ferris), the University of Pittsburgh Cancer Institute award P30 CA047904 (R. L. Ferris), P50CA101942 (G. J. Freeman), DE024173 (Y. Lei) and the IFORES program of University of Duisburg-Essen (B. A. Kansy)
Robert L. Ferris: consulting or advisory role: AstraZeneca, Bristol-Myers Squibb, Merck, ONO Pharmaceutical and Celgene. Research funding: Amgen (Inst), Bristol-Myers Squibb (Inst.) AstraZeneca (Inst.) and VentiRx (Inst.). GF has patents/pending royalties on the PD-1 pathway from Bristol-Myers-Squibb, Roche, Merck, EMD-Serono, Boehringer-Ingelheim, AstraZeneca, and Novartis. GF has served on advisory boards for CoStim, Novartis, Roche, Eli Lilly, Bristol-Myers-Squibb, Seattle Genetics, Bethyl Laboratories, Xios, and Quiet.
Footnotes
Conflict of interest: The other authors declare no conflict of interest
References
- 1.Brahmer JR, Hammers H, Lipson EJ. Nivolumab: targeting PD-1 to bolster antitumor immunity. Future oncology (London, England) 2015:0. doi: 10.2217/fon.15.52. [DOI] [PubMed] [Google Scholar]
- 2.Peggs KS, Quezada SA, Chambers CA, Korman AJ, Allison JP. Blockade of CTLA-4 on both effector and regulatory T cell compartments contributes to the antitumor activity of anti-CTLA-4 antibodies. The Journal of experimental medicine. 2009;206:1717–25. doi: 10.1084/jem.20082492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Brahmer J, Reckamp KL, Baas P, Crino L, Eberhardt WE, Poddubskaya E, et al. Nivolumab versus Docetaxel in Advanced Squamous-Cell Non-Small-Cell Lung Cancer. The New England journal of medicine. 2015;373:123–35. doi: 10.1056/NEJMoa1504627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Larkin J, Chiarion-Sileni V, Gonzalez R, Grob JJ, Cowey CL, Lao CD, et al. Combined Nivolumab and Ipilimumab or Monotherapy in Untreated Melanoma. The New England journal of medicine. 2015;373:23–34. doi: 10.1056/NEJMoa1504030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.McDermott DF, Drake CG, Sznol M, Choueiri TK, Powderly JD, Smith DC, et al. Survival, Durable Response, and Long-Term Safety in Patients With Previously Treated Advanced Renal Cell Carcinoma Receiving Nivolumab. Journal of clinical oncology: official journal of the American Society of Clinical Oncology. 2015;33:2013–20. doi: 10.1200/JCO.2014.58.1041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Benson E, Li R, Eisele D, Fakhry C. The clinical impact of HPV tumor status upon head and neck squamous cell carcinomas. Oral oncology. 2014;50:565–74. doi: 10.1016/j.oraloncology.2013.09.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ferris RL, Blumenschein G, Jr, Fayette J, Guigay J, Colevas AD, Licitra L, et al. Nivolumab for Recurrent Squamous-Cell Carcinoma of the Head and Neck. N Engl J Med. 2016;375:1856–67. doi: 10.1056/NEJMoa1602252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Concha-Benavente F, Srivastava RM, Trivedi S, Lei Y, Chandran U, Seethala RR, et al. Identification of the cell-intrinsic and extrinsic pathways downstream of EGFR and IFNgamma that induce PD-L1 expression in head and neck cancer. Cancer research. 2015 doi: 10.1158/0008-5472.CAN-15-2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ferris RL. Immunology and Immunotherapy of Head and Neck Cancer. J Clin Oncol. 2015;33:3293–304. doi: 10.1200/JCO.2015.61.1509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Blackburn SD, Shin H, Freeman GJ, Wherry EJ. Selective expansion of a subset of exhausted CD8 T cells by alphaPD-L1 blockade. Proceedings of the National Academy of Sciences of the United States of America. 2008;105:15016–21. doi: 10.1073/pnas.0801497105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Gros A, Robbins PF, Yao X, Li YF, Turcotte S, Tran E, et al. PD-1 identifies the patient-specific CD8(+) tumor-reactive repertoire infiltrating human tumors. The Journal of clinical investigation. 2014;124:2246–59. doi: 10.1172/JCI73639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Badoual C, Hans S, Merillon N, Van Ryswick C, Ravel P, Benhamouda N, et al. PD-1-expressing tumor-infiltrating T cells are a favorable prognostic biomarker in HPV-associated head and neck cancer. Cancer research. 2013;73:128–38. doi: 10.1158/0008-5472.CAN-12-2606. [DOI] [PubMed] [Google Scholar]
- 13.Li J, Shayan G, Avery L, Jie H-B, Gildener-Leapman N, Schmitt N, Lu B, Kane LP, Ferris RL. Tumor-infiltrating Tim-3+ T cells proliferate avidly except when PD-1 is co-expressed: evidence for intracellular cross talk. Oncoimmunology. 2016 doi: 10.1080/2162402X.2016.1200778. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Shayan G, Srivastava R, Li J, Schmitt N, Kane LP, Ferris RL. Adaptive resistance to anti-PD1 therapy by Tim-3 upregulation is mediated by the PI3K-Akt pathway in head and neck cancer. Oncoimmunology. 2017;6:e1261779. doi: 10.1080/2162402X.2016.1261779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Monney L, Sabatos CA, Gaglia JL, Ryu A, Waldner H, Chernova T, et al. Th1-specific cell surface protein Tim-3 regulates macrophage activation and severity of an autoimmune disease. Nature. 2002;415:536–41. doi: 10.1038/415536a. [DOI] [PubMed] [Google Scholar]
- 16.Anderson AC, Lord GM, Dardalhon V, Lee DH, Sabatos-Peyton CA, Glimcher LH, et al. T-bet, a Th1 transcription factor regulates the expression of Tim-3. European journal of immunology. 2010;40:859–66. doi: 10.1002/eji.200939842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Le Mercier I, Lines JL, Noelle RJ. Beyond CTLA-4 and PD-1, the Generation Z of Negative Checkpoint Regulators. Frontiers in immunology. 2015;6:418. doi: 10.3389/fimmu.2015.00418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Anderson AC, Joller N, Kuchroo VK. Lag-3, Tim-3, and TIGIT: Co-inhibitory Receptors with Specialized Functions in Immune Regulation. Immunity. 2016;44:989–1004. doi: 10.1016/j.immuni.2016.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Grosso JF, Kelleher CC, Harris TJ, Maris CH, Hipkiss EL, De Marzo A, et al. LAG-3 regulates CD8+ T cell accumulation and effector function in murine self- and tumor-tolerance systems. The Journal of clinical investigation. 2007;117:3383–92. doi: 10.1172/JCI31184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Legat A, Speiser DE, Pircher H, Zehn D, Fuertes Marraco SA. Inhibitory Receptor Expression Depends More Dominantly on Differentiation and Activation than “Exhaustion” of Human CD8 T Cells. Frontiers in immunology. 2013;4:455. doi: 10.3389/fimmu.2013.00455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Comprehensive genomic characterization of head and neck squamous cell carcinomas. Nature. 2015;517:576–82. doi: 10.1038/nature14129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Li B, Dewey CN. RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC bioinformatics. 2011;12:323. doi: 10.1186/1471-2105-12-323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hoover AC, Spanos WC, Harris GF, Anderson ME, Klingelhutz AJ, Lee JH. The role of human papillomavirus 16 E6 in anchorage-independent and invasive growth of mouse tonsil epithelium. Archives of otolaryngology–head & neck surgery. 2007;133:495–502. doi: 10.1001/archotol.133.5.495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Li B, VanRoey M, Wang C, Chen TH, Korman A, Jooss K. Anti-programmed death-1 synergizes with granulocyte macrophage colony-stimulating factor–secreting tumor cell immunotherapy providing therapeutic benefit to mice with established tumors. Clinical cancer research: an official journal of the American Association for Cancer Research. 2009;15:1623–34. doi: 10.1158/1078-0432.CCR-08-1825. [DOI] [PubMed] [Google Scholar]
- 25.Deng L, Liang H, Burnette B, Beckett M, Darga T, Weichselbaum RR, et al. Irradiation and anti-PD-L1 treatment synergistically promote antitumor immunity in mice. The Journal of clinical investigation. 2014;124:687–95. doi: 10.1172/JCI67313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lanzavecchia A, Sallusto F. Understanding the generation and function of memory T cell subsets. Current opinion in immunology. 2005;17:326–32. doi: 10.1016/j.coi.2005.04.010. [DOI] [PubMed] [Google Scholar]
- 27.Wherry EJ, Ha SJ, Kaech SM, Haining WN, Sarkar S, Kalia V, et al. Molecular signature of CD8+ T cell exhaustion during chronic viral infection. Immunity. 2007;27:670–84. doi: 10.1016/j.immuni.2007.09.006. [DOI] [PubMed] [Google Scholar]
- 28.Li J, Jie HB, Lei Y, Gildener-Leapman N, Trivedi S, Green T, et al. PD-1/SHP-2 inhibits Tc1/Th1 phenotypic responses and the activation of T cells in the tumor microenvironment. Cancer research. 2015;75:508–18. doi: 10.1158/0008-5472.CAN-14-1215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Jie HB, Gildener-Leapman N, Li J, Srivastava RM, Gibson SP, Whiteside TL, et al. Intratumoral regulatory T cells upregulate immunosuppressive molecules in head and neck cancer patients. British journal of cancer. 2013;109:2629–35. doi: 10.1038/bjc.2013.645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Maxwell JH, Ferris RL, Gooding W, Cunningham D, Mehta V, Kim S, et al. Extracapsular spread in head and neck carcinoma: impact of site and human papillomavirus status. Cancer. 2013;119:3302–8. doi: 10.1002/cncr.28169. [DOI] [PubMed] [Google Scholar]
- 31.Topalian SL, Hodi FS, Brahmer JR, Gettinger SN, Smith DC, McDermott DF, et al. Safety, activity, and immune correlates of anti-PD-1 antibody in cancer. The New England journal of medicine. 2012;366:2443–54. doi: 10.1056/NEJMoa1200690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Das R, Verma R, Sznol M, Boddupalli CS, Gettinger SN, Kluger H, et al. Combination Therapy with Anti-CTLA-4 and Anti-PD-1 Leads to Distinct Immunologic Changes In Vivo. Journal of immunology (Baltimore, Md: 1950) 2015;194:950–9. doi: 10.4049/jimmunol.1401686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Ferris R. PD-1 targeting in cancer immunotherapy. Cancer. 2013;119:E1–3. doi: 10.1002/cncr.27832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Couzin-Frankel J. Breakthrough of the year 2013. Cancer immunotherapy. Science (New York, NY) 2013;342:1432–3. doi: 10.1126/science.342.6165.1432. [DOI] [PubMed] [Google Scholar]
- 35.Chikuma S, Terawaki S, Hayashi T, Nabeshima R, Yoshida T, Shibayama S, et al. PD-1-mediated suppression of IL-2 production induces CD8+ T cell anergy in vivo. Journal of immunology (Baltimore, Md: 1950) 2009;182:6682–9. doi: 10.4049/jimmunol.0900080. [DOI] [PubMed] [Google Scholar]
- 36.Zhang Y, Kang S, Shen J, He J, Jiang L, Wang W, et al. Prognostic significance of programmed cell death 1 (PD-1) or PD-1 ligand 1 (PD-L1) Expression in epithelial-originated cancer: a meta-analysis. Medicine. 2015;94:e515. doi: 10.1097/MD.0000000000000515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Ang KK, Harris J, Wheeler R, Weber R, Rosenthal DI, Nguyen-Tan PF, et al. Human papillomavirus and survival of patients with oropharyngeal cancer. The New England journal of medicine. 2010;363:24–35. doi: 10.1056/NEJMoa0912217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Rosales R, Rosales C. Immune therapy for human papillomaviruses-related cancers. World journal of clinical oncology. 2014;5:1002–19. doi: 10.5306/wjco.v5.i5.1002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Lyford-Pike S, Peng S, Young GD, Taube JM, Westra WH, Akpeng B, et al. Evidence for a role of the PD-1:PD-L1 pathway in immune resistance of HPV-associated head and neck squamous cell carcinoma. Cancer research. 2013;73:1733–41. doi: 10.1158/0008-5472.CAN-12-2384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Morales-Sanchez A, Fuentes-Panana EM. Human viruses and cancer. Viruses. 2014;6:4047–79. doi: 10.3390/v6104047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Fourcade J, Sun Z, Pagliano O, Chauvin JM, Sander C, Janjic B, et al. PD-1 and Tim-3 regulate the expansion of tumor antigen-specific CD8(+) T cells induced by melanoma vaccines. Cancer research. 2014;74:1045–55. doi: 10.1158/0008-5472.CAN-13-2908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Flecken T, Schmidt N, Hild S, Gostick E, Drognitz O, Zeiser R, et al. Immunodominance and functional alterations of tumor-associated antigen-specific CD8+ T-cell responses in hepatocellular carcinoma. Hepatology (Baltimore, Md) 2014;59:1415–26. doi: 10.1002/hep.26731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Zhang G, Li Z, Han Q, Li N, Zhu Q, Li F, et al. Altered TNF-alpha and IFN-gamma levels associated with PD1 but not TNFA polymorphisms in patients with chronic HBV infection. Infection, genetics and evolution: journal of molecular epidemiology and evolutionary genetics in infectious diseases. 2011;11:1624–30. doi: 10.1016/j.meegid.2011.06.004. [DOI] [PubMed] [Google Scholar]
- 44.Peng W, Liu C, Xu C, Lou Y, Chen J, Yang Y, et al. PD-1 blockade enhances T-cell migration to tumors by elevating IFN-gamma inducible chemokines. Cancer research. 2012;72:5209–18. doi: 10.1158/0008-5472.CAN-12-1187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Blackburn SD, Crawford A, Shin H, Polley A, Freeman GJ, Wherry EJ. Tissue-specific differences in PD-1 and PD-L1 expression during chronic viral infection: implications for CD8 T-cell exhaustion. Journal of virology. 2010;84:2078–89. doi: 10.1128/JVI.01579-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Tanguy Y, Seiwert BB, Weiss Jared, Gluck Iris, Paul Eder Joseph, Pai Sara I, Dolled-Filhart Marisa, Emancipator Kenneth, Pathiraja Kumudu, Gause Christine, Iannone Robert, Brown Holly, Houp Jennifer, Cheng Jonathan D, Chow Laura Quan Man. A phase Ib study of MK-3475 in patients with human papillomavirus (HPV)-associated and non-HPV–associated head and neck (H/N) cancer. Journal of Clinical Oncology, 2014 ASCO Annual Meeting Abstracts. 2014;32 [Google Scholar]
- 47.Fury SIO M, Balmanoukian AS, Hansen A, Massarelli E, Blake-Haskins A, Li X, Robbins PB, Vasselli J, Segal NH. 988PD - Clinical activity and safety of MEDI4736, an anti-PD-L1 antibody, in patients with head and neck cancer. Annals of Oncology. 2014;25(suppl_4):iv340–iv56. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.