

Abstract
The ribosome is a dynamic particle that undergoes many structural changes during translation. We show through chemical probing with dimethyl sulfate (DMS) that conformational changes occur at several nucleotides in the peptidyl transferase center upon alterations in pH, temperature, and monovalent ion concentration, consistent with observations made by Elson and coworkers over 30 years ago. Moreover, we have found that the pH-dependent DMS reactivity of A2451 in the center of the 23S rRNA peptidyl transferase region, ascribed to a perturbed pKa of this base, occurs only in inactive 50S and 70S ribosomes. The degree of DMS reactivity of this base in the inactive ribosomes depends on both the identity and amount of monovalent ion present. Furthermore, G2447, a residue proposed to be critical for the hypothesized pKa perturbation, is not essential for the conditional DMS reactivity at A2451. Given that the pH-dependent change in DMS reactivity at A2451 occurs only in inactive ribosomes, and that this DMS reactivity can increase with increasing salt (independently of pH), we conclude that this observation cannot be used as supporting evidence for a recently proposed model of acid/base catalyzed ribosomal transpeptidation.
The dynamics of ribosome translation is a challenging problem for structural biologists. Structural changes occur during assembly of the subunits, tRNA binding, and translocation. The recent wealth of information from cryoelectron microscopy and x-ray structures of different ribosomal complexes provides clear evidence for various conformational changes during the ribosomal cycle (1–5). Biochemical evidence for structural switches in rRNA comes from mutagenesis studies on 16S rRNA in the 912 region (6) and from probing 23S rRNA from pre- and posttranslocational complexes (7), where recurrent changes in rRNA structure have been detected during elongation. In addition, experiments from more than 30 years ago suggested reversible conformational changes in the Escherichia coli 50S subunit, where, upon removal of monovalent cations, these particles could no longer perform peptidyl transfer or bind the peptidyl transferase inhibitor chloramphenicol (8–10). This phenomenon was thought to represent a conformational change near the peptidyl transferase center (Fig. 1A), and chemical footprinting with the peptidyl transferase inhibitor chloramphenicol later revealed this region to include 23S rRNA residue A2451 (11). A2451 is a universally conserved nucleotide whose interaction with peptidyl transferase substrates has been implicated by several methods (12–14).
Figure 1.

The peptidyl transferase center. (A) Secondary structure of the peptidyl transferase center (PTC), with bases of interest in this study indicated. All numbering is for E. coli. (B) Schematic of the catalytic center taken from the H. marismortui 50S crystal structure, with the relative position of the tetrahedral intermediate analogue soaked into the crystal indicated (16).
The recent publication of the high-resolution crystal structure of the Haloarcula marismortui 50S ribosomal subunit has placed A2451 at the center of attention, as it was shown to be the closest nucleotide to a peptidyl transferase inhibitor (C-C-dA-phosphoramide-puromycin) designed to mimic the tetrahedral intermediate formed during transpeptidation (15–17). Based on this proximity, the authors proposed a mechanism for acid-base catalysis in which the N-3 position of A2451 sequentially accepts and then donates a proton in the formation and resolution of the intermediate (16). Alternatively, it was hypothesized that N-3 could participate in catalysis through oxyanion stabilization by hydrogen bonding to the tetrahedral intermediate's nonbridging oxygen. For either of these to happen, it was recognized that the pKa of the N-3 of A2451 would have to be significantly perturbed, from a pKa below 1 (18) to near physiological levels. This perturbation was proposed to stem from a charge relay network between A2451, residue G2447, and the backbone phosphate of A2450 (Fig. 1B). It was recognized by the authors that the identity of G2447 was critical for the charge relay system, and that a G-to-A substitution at this base would be incompatible with the contacts proposed to be essential in the network (16). Supporting evidence for this hypothesis was found by the interesting discovery that in E. coli the level of reactivity of the N-1 or N-3 position of A2451 to the modifying agent dimethyl sulfate (DMS) was pH dependent, suggesting a pKa of around 7.6 (19). Although this result was interpreted as convincing biochemical data that the proposed model was correct, an equally valid explanation was that a conformational change at A2451 alternatively exposed or shielded this base from chemical modification.
In our efforts to probe for structural changes at the peptidyl transferase region under different reaction conditions, we decided to test for a relationship between these conditions and the pH-dependent DMS reactivity of A2451. In particular, we tested for the influence of monovalent ions, given the proximity in the structure of A2451 to a coordinated monovalent K+ ion, as well as the previous proposal of a conformational change in the peptidyl transferase center upon active/inactive transition. In this study, we have found that pH-dependent reactivity of this base toward DMS occurs only in ribosomes that are in an inactive conformation. These ribosomes are unable to catalyze peptide bond formation, as assayed by the fragment reaction. Furthermore, we show that the conditional reactivity of A2451 is affected not only by pH and temperature but also by monovalent ion concentration and identity. We also observe additional pH-dependent changes in DMS reactivity in other nucleotides of both the 50S and 30S subunits of 70S ribosomes. These data indicate that upon ribosome activation a conformational change occurs at the peptidyl transferase region that protects A2451 from DMS modification. Finally, we show that mutation of residue G2447, proposed to be critical in the charge relay model for a perturbed pKa at A2451, has no effect on the latter base's pH-dependent DMS reactivity. We conclude that the conditional DMS reactivity of A2451 is due to a conformational change relating to the active/inactive transition rather than a perturbed pKa, and as such cannot be used as evidence to support the proposed model for ribosomal transpeptidation.
Materials and Methods
70S Preparation and Modification.
Reassociated E. coli 70S ribosomes from E. coli strain Can 20 were prepared as described (7). E. coli ribosomes (70 S, 7 pmol) were added to 30 μl buffer A [50 mM Hepes⋅KOH with pH 6.5, 7.5, or 8.5/100 mM KCl/10 mM Mg(OAc)2] at 0°C. When indicated in figures the ribosomes were incubated for 15 min at 37°C before DMS modification. RNA was usually methylated by the addition of 1.2 μl DMS diluted by 1:8 in ethanol and incubated at 0°C for 1 h or for 10 min at 37°C. The reaction was stopped with 15 μl stop solution (1 M Tris⋅Cl, pH 7.5/1 M β-mercapthoethanol/0.12 M EDTA), and ribosomes were precipitated with 500 μl ethanol. rRNA was purified and used as a template for primer extension analysis as described (7). Gels were scanned and analyzed with a Molecular Dynamics PhosphorImager.
50S Preparation and Modification.
E. coli 50S subunits from strains XL-1, MC250 pLK35 (wt), or MC250 pLK2447A were prepared as described (20) with the following modifications. After fractionation 50S subunits were dialyzed into 10 mM K-Hepes (pH 7.5), 10 mM MgCl2, and 5 mM β-mercapthoethanol, then pelleted and resuspended in the same buffer. The K+ ion concentration in this buffer, coming from the KOH used to equilibrate the Hepes, was between 2 and 3 mM. DMS modification was performed as described (21) in the buffers containing 50 mM K-Pipes (pH 6.5), 50 mM K-Hepes (pH 7.5), or 50 mM K-bicine (pH 8.5), with 15 mM MgCl2 and the relevant concentration of KCl, NH4Cl, or NaCl as described in the Results. Gels were scanned and analyzed with a Fuji BAS-2500 PhosphorImager.
Peptidyl Transferase Assay.
Peptidyl transferase assays were performed with the fragment reaction as described (22), with the modification that ribosomes were preincubated as described in the Results.
Results
Loss of pH-Dependent DMS Reactivity at A2451 upon Ribosome Activation.
Published data (19) showed that DMS modification of A2451 in 23S rRNA of 50S subunits was much greater at pH 8.5 than at pH 6.5, which was attributed to a perturbed pKa of this nucleotide. We decided to investigate this pH-dependent reactivity at A2451 in a variety of ribosomal complexes under varying ionic and pH conditions. Initially we probed reassociated, empty 70S ribosomes. Only DMS incubation for 1 h at 0°C (the published conditions) consistently gave the expected pH-dependent shift at this nucleotide (Fig. 2A). In addition to A2451, we also found A2453 to exhibit a similar pH-dependent shift in DMS reactivity in 70S ribosomes.
Figure 2.

pH-dependent changes in DMS reactivity at A2451 in 70S ribosomes under different conditions. Primer extension analysis of 23S rRNAs isolated from unmodified (DMS −) and DMS-modified (DMS +) ribosomes. A, C, and U are dideoxy sequencing lanes. (A) Influence of DMS incubation conditions: 70S ribosomes were modified with DMS either for 10 min at 37°C or for 1 h at 0°C. The asterisk denotes A2469, the modification of which is strongly enhanced at higher temperatures. (B) Influence of the activation of 70S ribosomes by preincubation for 15 min at 37°C (lanes 1–4) or 15 min at 0°C (lanes 5–8), before DMS modification (1 h at 0°C).
We were puzzled by the fact that DMS modification for 10 min at 37°C resulted in equally low modification of A2451 at pH 6.5 and 8.5, even though other nucleotides such as A2469 were hypermodified (Fig. 2A, asterisks in lanes 2 and 3). Therefore, 70S particles were preincubated for 15 min at 37°C and then DMS modified at 0°C for 1 h. The results in Fig. 2B show that the preincubation step abolished the enhanced DMS modification at higher pH. As ribosomal particles are known to need a temperature activation step in translation buffer, these results suggested an influence of the temperature on the pH-dependent DMS modification. Subsequently, ribosomal complexes constructed with tRNA and mRNA, which were incubated at elevated temperature for complex formation, also showed no pH-dependent DMS modification at A2451 (data not shown). As these results were consistent with the active/inactive state model described 30 years earlier by Elson and coworkers (10), in which monovalent ions play a crucial role in this transition, we decided to investigate the role of monovalent ions on the reactivity of A2451 to DMS more thoroughly.
Monovalent Ions and Heat Incubation Affect A2451 DMS Reactivity.
Both 50S and 70S ribosomes are inactivated by low levels of monovalent cations during their preparation (10) and can be reactivated by incubation in 150 mM KCl or 100 mM NH4Cl, but not 100 mM NaCl, for 10 min at 37°C. The reactivated ribosomes remain active when placed on ice. With this approach we inactivated 50S subunits by dialysis at low concentration of monovalent cation (≈2–3 mM K+) and then performed chemical modification in the presence of different monovalent ions at varying concentrations, with or without preincubation at 37°C. Inactivated subunits incubated and modified on ice exhibited pH-dependent DMS reactivity at A2451 in modification buffers containing either 150 mM KCl or 100 mM NH4Cl (Fig. 3A). However, incubation of these subunits at 37°C for 10 min (conditions that reactivate ribosomes; ref. 10) before modification at 0°C resulted in the loss of DMS reactivity of A2451 at all pHs, consistent with what we had observed in 70S ribosomes.
Figure 3.

Influence of changing monovalent ion concentration and temperature on the 50S subunit's pH-dependent DMS reactivity at A2451 and ability to perform peptidyl transfer. Unm, unmodified ribosomes. (A) Heat activation of 50S subunits in K+ (lanes 6–8) or NH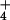 (lanes 12–14) results in loss of pH-dependent DMS reactivity at A2451. (B) Heat incubation in the absence of monovalent cation (lanes 6–8) or in the presence of Na+ (lanes 12–14) does not result in loss of pH-dependent DMS reactivity at A2451. (C) Increasing concentrations of K+ (lanes 3–5) or NH
(lanes 12–14) results in loss of pH-dependent DMS reactivity at A2451. (B) Heat incubation in the absence of monovalent cation (lanes 6–8) or in the presence of Na+ (lanes 12–14) does not result in loss of pH-dependent DMS reactivity at A2451. (C) Increasing concentrations of K+ (lanes 3–5) or NH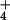 (lanes 6–8) at constant pH (6.5) at 0°C results in increasing DMS reactivity of A2451. (D) 50S subunits shown to exhibit pH-dependent DMS reactivity at A2451 cannot perform peptidyl transferase as assayed by the fragment reaction. 50S preincubation conditions are as shown.
(lanes 6–8) at constant pH (6.5) at 0°C results in increasing DMS reactivity of A2451. (D) 50S subunits shown to exhibit pH-dependent DMS reactivity at A2451 cannot perform peptidyl transferase as assayed by the fragment reaction. 50S preincubation conditions are as shown.
To test this correlation with the active/inactive transition further, we modified the 50S subunits on ice at different pHs either in the absence of monovalent cation or in the presence of 100 mM NaCl, with or without a 10-min preincubation at 37°C (conditions that do not reactivate ribosomes; Fig. 3B). 50S subunits incubated and modified on ice in the absence of monovalent cation displayed the pH-dependent reactivity to DMS at A2451. However, this effect was retained when the ribosomes were preincubated at 37°C, in contrast to the situation where modification was performed in NH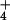 or K+ ions. Similarly, 50S subunits displayed pH-dependent DMS reactivity at A2451 when NaCl (100 mM) was the monovalent cation in the modification buffer, and this effect was retained in ribosomes preincubated at 37°C, although the level of reactivity was slightly diminished. We conclude that the inactivation of ribosomes by removal of monovalent cations yields particles with pH-dependent reactivity at A2451, and reactivation with the appropriate cation coincides with a conformational change that renders this base unreactive to chemical modification at any pH.
or K+ ions. Similarly, 50S subunits displayed pH-dependent DMS reactivity at A2451 when NaCl (100 mM) was the monovalent cation in the modification buffer, and this effect was retained in ribosomes preincubated at 37°C, although the level of reactivity was slightly diminished. We conclude that the inactivation of ribosomes by removal of monovalent cations yields particles with pH-dependent reactivity at A2451, and reactivation with the appropriate cation coincides with a conformational change that renders this base unreactive to chemical modification at any pH.
K+ or NH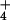 Concentration Affects DMS Reactivity of A2451 Independently of pH.
Concentration Affects DMS Reactivity of A2451 Independently of pH.
DMS modification of A2451 in inactive ribosomes at a given pH was greater in NH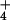 than in K+ (Fig. 3A). We asked whether DMS reactivity in inactive 50S subunits might be affected by the concentration (as well as the identity) of monovalent ion present during modification, even in the absence of a pH shift. As shown in Fig. 3C, increasing concentrations of K+ or NH
than in K+ (Fig. 3A). We asked whether DMS reactivity in inactive 50S subunits might be affected by the concentration (as well as the identity) of monovalent ion present during modification, even in the absence of a pH shift. As shown in Fig. 3C, increasing concentrations of K+ or NH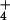 ion resulted in increasing DMS reactivity at A2451 when all modifications were performed at pH 6.5 and at 0°C. As the degree of DMS modification at A2451 in inactive ribosomes is influenced by the identity and concentration of monovalent ions independently of pH, this conditional reactivity is inconsistent with a perturbation of the native pKa and must result from structural change.
ion resulted in increasing DMS reactivity at A2451 when all modifications were performed at pH 6.5 and at 0°C. As the degree of DMS modification at A2451 in inactive ribosomes is influenced by the identity and concentration of monovalent ions independently of pH, this conditional reactivity is inconsistent with a perturbation of the native pKa and must result from structural change.
50S Subunits Displaying pH-Dependent DMS Reactivity at A2451 Are Inactive in Peptidyl Transferase.
To rigorously confirm the correlation between the Zamir/Elson transition and the conditional reactivity of A2451 to DMS, 50S subunits preincubated under the various conditions described above for the chemical modification were assayed for peptidyl transferase activity with the fragment reaction (23). Indeed, we found that only subunits preincubated in either K+ or NH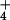 ions at 37°C were capable of performing ribosomal transpeptidation, as was originally described over 30 years ago (Fig. 3D). This finding confirmed that only inactive subunits, incapable of performing peptidyl transfer, display the pH-dependent change in DMS reactivity at residue A2451.
ions at 37°C were capable of performing ribosomal transpeptidation, as was originally described over 30 years ago (Fig. 3D). This finding confirmed that only inactive subunits, incapable of performing peptidyl transfer, display the pH-dependent change in DMS reactivity at residue A2451.
Effect of G2447A Mutation on pH-Dependent DMS Reactivity of A2451.
In the proposed model of ribosomal transpeptidation (16), the displacement of charge necessary for the perturbed pKa of A2451 is provided through a relay system passing through G2447 to the phosphate backbone of residue A2450 (Fig. 1B). It has been shown recently that the mutation of G2447 to A is viable in a strain of E. coli in which these mutants represent the sole source of ribosomes (20). As expected, these mutant ribosomes were shown to be active in peptide bond formation both in vivo and in vitro. However, we were interested in determining the effect of this mutation on the pH-dependent DMS reactivity of A2451, as the critical identity of this residue in the proposed model suggests that the mutation would abolish this effect. We found that the G2447A mutation had no discernible effect on the pH-dependent DMS reactivity of A2451 in inactive 50S subunits (Fig. 4), further indicating that the conditional DMS reactivity of A2451 is not due to a perturbed pKa. As predicted, activation of mutant 50S subunits resulted in loss of DMS reactivity at A2451 (data not shown).
Figure 4.

Mutation of G2447 to A has no effect on pH-dependent DMS reactivity of A2451. Unm, unmodified ribosomes. Lanes 3–5: WT ribosomes. Lanes 6–8: G2447A ribosomes. Sequencing lanes are given for the G2447A mutant.
Additional Sites of pH-Dependent DMS Reactivity in the Ribosome.
We were interested in identifying nucleotides in addition to A2451 that showed a pH-dependent change in DMS reactivity. Thus we screened the complete 16S and 23S rRNA of 70S ribosomes (except the very 3′ ends) for modification. As discussed above, only A2453 showed an enhancement of DMS reactivity at higher pH, similar to that observed with A2451, and this reactivity disappeared on subunit activation (Fig. 2). Interestingly, we found three adenosines (A908 in16S rRNA; A471 and A2572 in 23S rRNA) with a reverse pH dependence; these nucleotides were more highly modified by DMS at pH 6.5 than at pH 8.5. Among these, only A2572 showed a reduced reactivity at all pHs relative to that seen with inactive ribosomes (Fig. 5). As these data cannot be interpreted to result from a perturbed pKa of the adenosines, these changes in DMS reactivity must occur as a consequence of conformational changes at these residues upon changes in pH.
Figure 5.

Changes in DMS reactivity at additional nucleotides in the peptidyl transferase center on active/inactive transition. pH-dependent DMS reactivity of A2572 and U2584 changes on going from active (lanes 2–4) to inactive (lanes 6–8) ribosomes.
We also identified a uridine, U2584, the DMS modification of which at pH 8.5 depended on the ribosomes being in the active conformation (Fig. 5). This finding is of particular interest, as this nucleotide is also located in the peptidyl transferase region (Fig. 1A). We found several other uridines that were DMS reactive at pH 8.5 and not at pH 6.5, but the modification of these uridines was not affected by the active/inactive transition. These modifications were surprising, as DMS usually reacts with adenosine and cytosine at neutral pH, but not with uridine, due to a pKa of around 10 (N-3 of uridine; ref. 18). This observation indicates that these uridines probably have a perturbed pKa, and it is a shift from the keto to the enol tautomeric form that enables them to be modified at pH 8.5 but not at pH 6.5.
Taken together, the results from the differences in DMS modification of A2451, A2572, and U2584 show that defined structural changes occur at the peptidyl transferase center upon activation of the ribosome.
Discussion
We have shown that structural changes occur at specific nucleotides in the peptidyl transferase center upon transition between the inactive and active conformations. Of primary importance among these residues is A2451, the significance of which in peptide bond formation has been implicated by several biochemical and genetic experiments (12–14) and recently by the atomic resolution crystal structure of the H. marismortui 50S subunit (16). Our results show that in inactive ribosomes, the accessibility of A2451 to DMS modification is conditionally dependent on both pH and monovalent ion concentration. Upon activation, however, A2451 becomes unreactive to DMS under all conditions. We conclude from these observations that A2451 becomes shielded from chemical modification by a conformational change upon subunit activation, and that this base remains unreactive to DMS in ribosomes able to form peptide bonds.
The close proximity of A2451 to a transition state analogue in the crystal structure of the 50S subunit has led to the hypothesis that it participates in acid/base catalysis by virtue of a perturbed pKa at its N-3 position (16). Biochemical support for this hypothesis was provided by the interpretation that the pH-dependent DMS reactivity of A2451 resulted from a perturbed pKa (19). We have shown in this study, however, that the reported pH-dependent DMS modification at residue A2451 occurs only in inactive ribosomes and disappears upon activation. This observation accounts for the fact that changes in DMS reactivity are observed only at 0°C, as modification at 37°C in the appropriate ionic conditions results in ribosome activation. Inactivation can occur during the isolation of ribosomes in a low concentration of monovalent cations, and preparations may contain both active and inactive ribosomes. The active/inactive transition is reversible (10), and reactivation is accomplished by the incubation of ribosomes in a higher concentration of the appropriate monovalent cation (typically 100 mM NH4Cl) and a temperature shift, to 37°C in this work.
Support for a conformational change upon transition between active and inactive forms comes from the 50S subunit crystal structure. A2451 is in close proximity to a K+ ion that is coordinated to G2061 and G2447 (Fig. 1B). G2061 hydrogen bonds to the N-1 position of A2451, thereby protecting it from DMS modification. We propose that this K+ dissociates in the inactive conformation, making A2451 conditionally accessible to DMS. This dissociation might also account for the inability of inactive 50S subunits to bind chloramphenicol (9).
Several nucleotides in addition to A2451 demonstrated pH-dependent DMS modification. Interestingly, three adenosines (23S rRNA A471, A2572, and 16S rRNA A908) showed a pH-dependent DMS pattern that was the reverse of A2451: they were more strongly modified at pH 6.5 than at pH 8.5, indicating that a conformational change at lower pH exposes the N-1 position of these nucleotides. A907, the base adjacent to A908, has been shown to lose DMS reactivity upon heat activation of 30S (24), and both bases are adjacent to a conformational switch thought to occur during decoding (6). The pH-dependent DMS reactivity of A2572 is retained but is less intense in activated ribosomes. In the crystal structure, A2572 loops out in a helix connecting the A-loop with the peptidyl transferase center, resulting in a sharp bend at this nucleotide. This base had been shown to be cleaved by hydroxyl radicals generated by Fe(II)EDTA tethered to the 5′ end of A site tRNA and is hypermodified by DMS in erythromycin-resistant mutants of ribosomal protein L4 that harbor a deficiency in peptidyl transferase activity (25–27). Flexibility in this region was suggested by the observation that binding of a tRNA analogue to the A-loop of crystallized 50S subunits results in conformational changes in the peptidyl transferase region (16). Further corroboration of a flexible loop comes from our observation that activation of the ribosome changes the pH-dependent DMS modification pattern of A2572. However, at this time it is not possible to correlate pH-dependent structural changes with the observed pH dependency of ribosomal peptide bond formation.
Another interesting observation concerns nucleotide U2584, which is modified only upon activation of the ribosome. This residue is adjacent to U2585, which has been implicated in peptidyl transferase function (13, 14). Furthermore, it shifts in the 50S crystal structure on binding of the tetrahedral intermediate analogue to contact the terminal adenosine designed to mimic A76 of A-site tRNA. This conformational change at U2584, together with those observed at A2451 and A2572, provides strong evidence for a conformational rearrangement in the peptidyl transferase region upon activation of the ribosome.
The proposed model for acid/base catalyzed ribosomal peptidyl transfer has been the source of some debate among ribosomologists.§ Although the data presented here do not exclude the possibility of an acid/base catalytic mechanism, they remove the only direct biochemical support for a perturbed pKa at the proposed catalytic nucleotide. Taking into account the recent observations that the identity of A2451 or G2447 is not critical for ribosomal transpeptidation both in vitro and in vivo (20, 28), and the fact that mutagenesis of G2447 to A has no effect on the pH-dependent DMS reactivity of A2451, we believe that other models for peptide bond formation should also be entertained.
Our results show that differences in pH, temperature, and both the concentration and identity of monovalent cation can induce subtle changes in nucleotide conformation in the active site of the ribosome. These observations underline the difficulty of correlating structural data with biochemical data from flexible and dynamic RNA structures. The question remains whether the conformational changes described here have physiological relevance to the mechanism of peptide bond formation. Evidence that such could be the case comes from the recent 5.5-Å crystal structure of the Thermus thermophilus 70S ribosome complexed with mRNA and tRNA, in which several residues in the peptidyl transferase center, including A2451 and U2585, are in a conformation different from that described for the Haloarcula 50S subunit (5, 16). Complexed 70S crystal structures at higher resolution, together with more extensive biochemical characterization of the peptidyl transferase reaction, may yield definitive insights into this question.
Acknowledgments
We thank R. Green for her assistance in making tRNA fragments for the peptidyl transferase assay and N. Polacek and K. Nierhaus for discussions and ribosomes (Department of Molecular Biology and Genetics, The Johns Hopkins Univ. School of Medicine). We acknowledge S. Gregory, M. O'Connor, J. Thompson, S. Pond-Tor, and G. Q. Pennabble for helpful discussions and are indebted to K. Katze for her infallible instincts and suggestions. This work was supported by Grant P-13651-GEN from the Österreichischer Fonds zur Förderung der wissenschaftlichen Forschung (to A.B.) and U.S. National Institutes of Health Grant GM19756 (to A.E.D.).
Abbreviation
- DMS
dimethyl sulfate
Footnotes
Barta, A., Dorner, S., Polacek, N., Berg, J. M., Lorsch, J. R., Nissen, P., Hansen, J., Muth, G. W. & Ban, N. (2000) Science 291, 203a (abstr.).
References
- 1.Frank J, Agrawal R K. Nature (London) 2000;406:318–322. doi: 10.1038/35018597. [DOI] [PubMed] [Google Scholar]
- 2.Gabashvili I S, Agrawal R K, Grassucci R, Squires C L, Dahlberg A E, Frank J. EMBO J. 1999;18:6501–6507. doi: 10.1093/emboj/18.22.6501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ogle J M, Brodersen D E, Clemons W M, Jr, Tarry M J, Carter A P, Ramakrishnan V. Science. 2001;292:897–902. doi: 10.1126/science.1060612. [DOI] [PubMed] [Google Scholar]
- 4.Stark H, Rodnina M V, Wieden H-J, van Heel M, Wintermeyer W. Cell. 2000;100:301–309. doi: 10.1016/s0092-8674(00)80666-2. [DOI] [PubMed] [Google Scholar]
- 5.Yusopov M M, Yusopova G Z, Baucom A, Lieberman K, Earnest T N, Cate J H D, Noller H F. Science. 2001;292:883–896. doi: 10.1126/science.1060089. [DOI] [PubMed] [Google Scholar]
- 6.Lodmell J S, Dahlberg A E. Science. 1997;277:1262–1267. doi: 10.1126/science.277.5330.1262. [DOI] [PubMed] [Google Scholar]
- 7.Polacek N, Patzke S, Nierhaus K H, Barta A. Mol Cell. 2000;6:159–171. [PubMed] [Google Scholar]
- 8.Miskin R, Zamir A, Elson D. Biochem Biophys Res Commun. 1968;33:551–557. doi: 10.1016/0006-291x(68)90330-6. [DOI] [PubMed] [Google Scholar]
- 9.Vogel Z, Vogel T, Zamir A, Elson D. J Mol Biol. 1971;60:339–346. doi: 10.1016/0022-2836(71)90298-1. [DOI] [PubMed] [Google Scholar]
- 10.Zamir A, Miskin R, Vogel Z, Elson D. Methods Enzymol. 1974;30:406–426. doi: 10.1016/0076-6879(74)30042-0. [DOI] [PubMed] [Google Scholar]
- 11.Moazed D, Noller H F. Biochimie. 1987;69:879–884. doi: 10.1016/0300-9084(87)90215-x. [DOI] [PubMed] [Google Scholar]
- 12.Bocchetta M, Xiong L, Mankin A S. Proc Natl Acad Sci USA. 1998;95:3525–3530. doi: 10.1073/pnas.95.7.3525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Moazed D, Noller H F. Cell. 1989;57:585–597. doi: 10.1016/0092-8674(89)90128-1. [DOI] [PubMed] [Google Scholar]
- 14.Steiner G, Kuechler E, Barta A. EMBO J. 1988;7:3949–3955. doi: 10.1002/j.1460-2075.1988.tb03281.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ban N, Nissen P, Hansen J, Moore P B, Steitz T A. Science. 2000;289:905–919. doi: 10.1126/science.289.5481.905. [DOI] [PubMed] [Google Scholar]
- 16.Nissen P, Hansen J, Ban N, Moore P B, Steitz T A. Science. 2000;289:920–930. doi: 10.1126/science.289.5481.920. [DOI] [PubMed] [Google Scholar]
- 17.Welch M, Chastang J, Yarus M. Biochemistry. 1995;34:385–390. doi: 10.1021/bi00002a001. [DOI] [PubMed] [Google Scholar]
- 18.Saenger W. In: Principles of Nucleic Acid Structure. Charles R, Cantor, editors. New York: Springer; 1984. [Google Scholar]
- 19.Muth G W, Ortoleva-Donnely L, Strobel S. Science. 2000;289:947–950. doi: 10.1126/science.289.5481.947. [DOI] [PubMed] [Google Scholar]
- 20.Thompson J, Kim D F, O'Connor M, Lieberman K R, Bayfield M, Gregory S, Green R, Noller H F, Dahlberg A E. Proc Natl Acad Sci USA. 2001;98:9002–9007. doi: 10.1073/pnas.151257098. . (First Published July 24, 2001; 10.1073/pnas.151257098) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Van-Ryk D I, Dahlberg A E. Nucleic Acids Res. 1995;23:3563–3570. doi: 10.1093/nar/23.17.3563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Moazed D, Noller H F. Proc Natl Acad Sci USA. 1991;88:3725–3728. doi: 10.1073/pnas.88.9.3725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Monro R E, Cerna J, Marcker K A. Proc Natl Acad Sci USA. 1968;61:1042–1049. doi: 10.1073/pnas.61.3.1042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Moazed D, Van Stolk B J, Douthwaite S, Noller H F. J Mol Biol. 1986;191:483–493. doi: 10.1016/0022-2836(86)90143-9. [DOI] [PubMed] [Google Scholar]
- 25.Gregory S T, Dahlberg A E. J Mol Biol. 1999;289:827–834. doi: 10.1006/jmbi.1999.2839. [DOI] [PubMed] [Google Scholar]
- 26.Joseph S, Noller H F. EMBO J. 1996;15:910–916. [PMC free article] [PubMed] [Google Scholar]
- 27.Wittmann H G, Stoffler G, Apirion D, Rosen L, Tanaka K, Tamaki M, Takata R, Dekio S, Otaka E. Mol Gen Genet. 1973;127:175–189. doi: 10.1007/BF00333665. [DOI] [PubMed] [Google Scholar]
- 28.Polacek N, Gaynor M, Yassin A, Mankin A S. Nature (London) 2001;411:498–501. doi: 10.1038/35078113. [DOI] [PubMed] [Google Scholar]