

Abstract
Rationale
3,4-methylenedioxymethamphetamine (MDMA) persistently improves symptoms of post-traumatic stress disorder (PTSD) when combined with psychotherapy. Studies in rodents suggest that these effects can be attributed to enhancement of fear memory extinction. Therefore, MDMA may improve the effects of exposure-based therapy for PTSD; particularly in treatment-resistant patients. However, given MDMA's broad pharmacological profile, further investigation is warranted before moving to a complex clinical population.
Objectives
We aimed to inform clinical research by providing a translational model of MDMA's effect, and elucidating monoaminergic mechanisms through which MDMA enhances fear extinction.
Methods
We explored the importance of monoamine transporters targeted by MDMA to fear memory extinction, as measured by reductions in conditioned freezing and fear-potentiated startle (FPS) in mice. Mice were treated with selective inhibitors of individual monoamine transporters prior to combined MDMA treatment and fear extinction training.
Results
MDMA enhanced the lasting extinction of FPS. Acute and chronic treatment with a 5-HT transporter (5-HTT) inhibitor blocked MDMA's effect on fear memory extinction. Acute inhibition of dopamine (DA) and norepinephrine (NE) transporters had no effect. 5-HT release alone did not enhance extinction. Blockade of MDMA's effect by 5-HTT inhibition also downregulated 5-HT2A-mediated behavior, and 5-HT2A antagonism disrupted MDMA's effect on extinction.
Conclusions
We validate enhancement of fear memory extinction by MDMA in a translational behavioral model, and reveal the importance of 5-HTT and 5-HT2A receptors to this effect. These observations support future clinical research of MDMA as an adjunct to exposure therapy, and provide important pharmacological considerations for clinical use in a population frequently treated with 5-HTT inhibitors.
Keywords: Fear extinction, serotonin, monoamines, MDMA, fear-potentiated startle
Introduction
For several decades 3,4-methylenedioxymethamphetmine (MDMA) has garnered interest as an adjunct to psychotherapy due to its profound acute mood-elevating and prosocial effects (Greer and Tolbert 1986). Recently, just two sessions of MDMA-assisted psychotherapy have been found to promote long-term reductions in symptoms of post-traumatic stress disorder (PTSD) (Mithoefer et al. 2011). Although the pharmacological and psychological mechanisms of this effect are not well understood, preclinical research suggests that MDMA enhances the extinction of fear memory (Young et al. 2015). Further mechanistic studies of this phenomenon are required to anticipate important considerations for the use of MDMA in a clinical setting.
Many PTSD patients continue to experience intensely fearful responses to traumatic memories long after the fear response serves a protective function, which may be due to impairments in the extinction of fear memory (Rothbaum and Davis 2003). The extinction of fear memory is a learning process in which the fear response to an aversive memory diminishes as it repeatedly fails to predict an expected threat. Fear extinction processes can be engaged by exposure-based therapies for PTSD through the repeated re-exposure to trauma-related stimuli in the absence of harmful consequences (Rothbaum and Schwartz 2002). Pharmacological adjuncts that enhance extinction learning have been hypothesized to improve outcomes in exposure-based therapy (de Kleine et al. 2013). In fear-conditioned mice, administering MDMA systemically or directly to the amygdala 30 min prior to fear extinction training results in lasting improvements in fear memory extinction (Young et al. 2015). This effect depends on signaling through brain derived neurotrophic factor (BDNF) initiated when MDMA and extinction training are combined. However, translating the extinction-enhancing properties of MDMA in rodents to exposure-based therapies in human populations requires a significantly greater understanding of the mechanisms by which MDMA enhances the extinction of fear memory.
In the brain, MDMA primarily targets monoaminergic transporters and receptors (Battaglia et al. 1988), several of which have been demonstrated to play a role in the extinction of fear memory. Like other phenethylamine derivatives, MDMA potently releases dopamine (DA), norepinephrine (NE), and serotonin (5-HT) through their respective transporters (Green et al. 2003). Treatments that increase 5-HT or NE neurotransmission – selective serotonin reuptake inhibitors (SSRIs), exogenous NE, and direct serotonergic/adrenergic agonists – all have been observed to enhance fear memory extinction in certain cases (Burghardt and Bauer 2013; Mueller and Cahill 2010; Zhang et al. 2013). MDMA's effects on transporter-mediated release can be obstructed by pre-treatment with selective reuptake inhibitors (Fitzgerald and Reid 1990; Hekmatpanah and Peroutka 1990; Hysek et al. 2012). Specifically, SSRIs and norepinephrine-reuptake inhibitors (NRIs) also inhibit some of MDMA's behavioral/subjective effects (Hysek et al. 2011; Liechti and Vollenweider 2000b), demonstrating that MDMA's unique behavioral profile depends on access to transporters of 5-HT and NE.
The primary purpose of the current study was to identify monoaminergic mechanisms through which MDMA enhances the extinction of fear memory. A second aim of this study was to validate the effect of MDMA on fear memory extinction using a measure of fear behavior more translatable to humans. Previous studies with MDMA have used conditioned freezing, which is a stereotypic fear behavior in rodents. Conversely, fear-potentiated startle (FPS) is a highly conserved fear behavior across species, wherein the presence of a fear-eliciting stimulus increases the startle reflex to a burst of noise (Davis 2006). FPS has been repeatedly used as an index of conditioned fear expression and extinction across animal and human investigations (Briscione et al. 2014; Norrholm et al. 2006). Though their expression relies on different neurological systems, extinction of both conditioned freezing and FPS depend on the amygdala, where MDMA has increased BDNF signaling important for learning (Myers and Davis 2007; Young et al. 2015). While similar patterns of freezing and FPS expression are observed when simultaneously measuring them in response to a conditioned aversive stimulus, FPS appears to extinguish more slowly than conditioned freezing (Daldrup et al. 2015).
Using auditory Pavlovian fear conditioning to instate a fear memory, we demonstrate a leftward shift in the dose-response relationship between MDMA and the extinction of FPS in comparison to observations in previous studies of the extinction of conditioned freezing. Using selective pharmacological inhibitors of DA, NE or 5-HT transporters, we reveal a crucial role for the 5-HT transporter (5-HTT) in MDMA's enhancing effect on fear memory extinction. This effect was also blocked by pharmacological treatments that inhibit 5-HT2A receptor (5-HT2AR). The behavioral and pharmacological data here provide further evidence of MDMA's enhancing effect on fear memory extinction and also underscore the importance of 5-HT neurotransmission in this effect.
Methods and Materials
Animals
Male C57BL/6 mice were from the Jackson Laboratory and bred at the Yerkes National Primate Research Center at Emory University. Mice were group-housed in ventilated cages and maintained on ad libitum food and water. Lights in the vivarium turned on at 7:00 A.M. and turned off at 9:00 P.M. All experiments were performed on male mice between postnatal day 49 and 112. Studies were performed during the lights-on phase, with experiments taking place between 9:00 A.M. and 5:00 P.M, as mice trained and tested during the lights-on phase, mice show significantly slower extinction (Chaudhury and Colwell 2002). Studies were in accordance with National Institutes of Health guidelines, and all procedures were approved by the Institutional Animal Care and Use Committee at Emory University.
Drugs
S,R(±)-MDMA (MDMA), M100907 (M100), RTI-336 were supplied by the National Institute on Drug Abuse (Research Technology Branch, Research Triangle Park, NC, USA). Citalopram was obtained from Eli Lilly and Co. (Indianapolis, IN, USA). Reboxetine was obtained from Tocris Bioscience (Avonmouth, Bristol, UK). (-)-1-(2,5-Dimethoxy-4-iodophenyl)-2-aminopropane hydrochloride (DOI) was obtained from Sigma-Aldrich (St. Louis, MO, USA). MDMA, citalopram, reboxetine and DOI were dissolved in 0.9% saline immediately before experimentation. RTI-336 was initially dissolved in 100% ethanol and diluted to less than a 2% concentration with saline. M100 was dissolved in saline and 0.1N HCl. Drug vehicle served as control for each respective experiment with a given drug. All solutions were injected intraperitoneally (i.p.) at a volume of 10 μL/g body weight. MDMA was administered 30 min prior to the behavioral procedure based on a previous study, and transporter inhibitors were administered 30 min before MDMA because all three drugs reach peak brain concentrations within 30 min of administration and remain elevated for at least 90 min (Kimmel et al. 2008; Kreilgaard et al. 2008; Strolin Benedetti et al. 1995).
Conditioning and Extinction of Freezing Behavior
The procedure for acquisition and sub-optimal extinction of conditioned freezing are described in detail previously (Young et al. 2015). Briefly, mice were exposed to cued fear conditioning on Day 1, fear extinction training on Day 3, and extinction testing on Day 4. Cued fear conditioning consisted of a 4 pairings of a CS-tone (75-80dB, 6.0 kHz, 30 s) and a US-footshock (0.6 mA, 1 s). Extinction training was carried out 48 h after fear conditioning (‘Day 3’) in a different context from conditioning, where they were exposed to 16 CS tones separated by 45 s each. Saline or MDMA was administered 30 min prior to extinction training, and mice were individually-housed during the period between drug administration and the behavioral procedure. Freezing was scored by video recording and FreezeView software (Coulbourn Instruments, San Diego, CA, USA). For experiments exploring the effects of chronically-administered citalopram, mice were treated daily with 10 mg/kg (i.p.) of citalopram for 22 days between fear conditioning and extinction training. Mice were habituated to handling and i.p. injection for two days prior to experimentation.
Fear-Potentiated Startle (FPS)
Studies exploring FPS were carried out using a startle response system from San Diego Instruments (San Diego, CA, USA). Two separate FPS experiments were performed, and the protocols were based on previous studies of FPS in mice (Smith et al. 2011). The first experiment tested the effects of a range of MDMA doses on the within- and between-session extinction of FPS. On Days 1–3 (‘Acclimation’) of the first experiment, animals were placed in the startle chamber, given a 5-min period during which no auditory stimuli were presented, followed by a semi-random presentation of fifty 20-ms white noise startle stimuli (ten each of 70, 80, 85, 90, and 100 dB) with a 30-s inter-trial interval (ITI). On Day 4 (‘PreTest’), study mice were given a 5-min acclimation period followed by presentation of 10 startle stimuli (85 dB; 1 min ITI), and then 20 ‘CS+startle’ trials consisting of a 30-s, 12-kHz, 70-dB tone. This CS tone used in the PreTest is identical to the one used as the CS on the following fear conditioning day. On Day 5, animals were fear conditioned to ten CS-US pairings (0.25 s, 0.4-mA) in the same fear conditioning chambers used in previously described experiments. On Day 6 (‘Memory Test’), FPS was assessed by presenting 4 interspersed startle stimulus alone and CS+startle stimuli. FPS was averaged for all mice during this trial, and treatment groups were counterbalanced for equal FPS during the Memory Test. On Day 7, the effects of MDMA on the within-session extinction of FPS was tested by administering vehicle or MDMA 20 minutes prior to placing mice in the startle chamber. Mice were individually-housed in familiar small covered translucent plastic buckets filled with bedding after drug administration and before the behavioral procedure. Sub-threshold extinction of FPS (Smith et al, 2011) consisted of the same protocol used during the PreTest on Day 4. Between-session extinction learning (or extinction retention) was tested on Day 8 using the same protocol.
The second FPS experiment tested the effects of chronic citalopram treatment on the effects of MDMA. This approach was identical in most ways to the previous experiment, except that fear conditioning occurred on Day 1, followed by 22 days of treatment. Memory testing, within-session extinction training, and between-extinction (retention) testing occurred on Days 25, 26, and 27, respectively. Startle chambers were cleaned with distilled water in between individual animal test sessions and then cleaned again with Quattricide at the end of each daily session. Separate chambers used for fear conditioning were cleaned with 70% EtOH. Startle was assessed in 100-ms blocks during the presentation of the startle stimulus. During acclimation periods, startle was assessed every 30 s. FPS was calculated by the following formula using average startle measurements across a session:
Statistical Analysis
Behavioral data were analyzed with SPSS 22.0 (IBM; Armonk, NY) and Prism 5.0 (GraphPad) using a two-tailed t-test, one- or two-way ANOVA, or a repeated-measures ANOVA with α = 0.05. Post hoc ANOVA comparisons were made using Dunnett's test for dose-response data and Bonferroni's test for all other data. Data in figures are presented as mean ± SE. The data variance was similar between the groups compared in each statistical analysis using Bartlett's test for equal variances.
Results
Pre-treatment with inhibitor of 5-HTT, but not with other transporter inhibitors, blocks MDMA's enhancing effect on the extinction of conditioned freezing
Administration of a range of doses of citalopram (SSRI), reboxetine (NRI), or RTI-336 (DAT inhibitor) 60 min prior to fear extinction training revealed that only 20 mg/kg of citalopram reduced conditioned freezing during extinction training (Figure 1a-1c) [citalopram: F(3,28)=11.22, p<0.001; n=8/group]. However, no effects of citalopram, reboxetine or RTI-336 treatment were observed the following day when conditioned freezing was tested (Figure 1a-1c) [citalopram: F(3,28)=0.18, p=0.098; n=8/group]. 10 mg/kg of each drug was used in all subsequent experiments because it was the highest dose administered that had no significant effect on conditioned freezing or fear extinction when administered alone 60 min prior to extinction training.
Fig 1. Effect of transporter inhibitors on fear extinction and MDMA-induced enhancement of fear extinction.
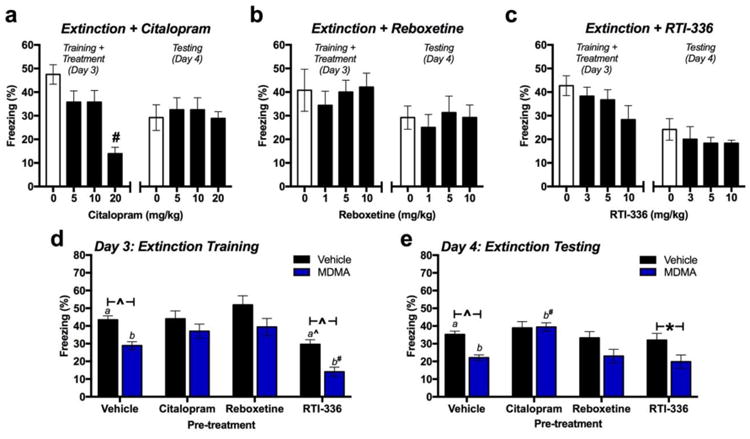
(a-c; left) Conditioned freezing during extinction training 60 min after receiving a dose of citalopram, reboxetine or RTI-336. (a-c-right) Conditioned freezing the following day (n=8/group). (d) Total average conditioned freezing during extinction training in vehicle- or MDMA-treated mice that received vehicle, citalopram, reboxetine, or RTI-336 60 min prior to extinction training. (n=15 in vehicle pre-treatment groups; n=9 in drug pre-treatment groups) (d) Total average conditioned freezing the day after treatment. *p<0.05; ^p<0.01, #p<0.001.
When all pre-treatment groups were analyzed together [nvehicle=15; ntreatment=9], a main effect of MDMA treatment was observed [F(1,76)=25.19, p<0.0001]. Individual two-way ANOVAs were performed for each transporter inhibitor-treated group in comparison to the group that received drug vehicle prior to MDMA. Only RTI-336 treatment had a main effect on conditioned freezing [F(1,44)=32.64, p<0.001], but it did not prevent MDMA from reducing conditioned freezing (Figure 1d) [RTI-336 × MDMA: F(1,44)=0.039, p=0.843]. Although both citalopram and reboxetine treatment prevented the MDMA-induced reductions in conditioned freezing observed in the vehicle-treated group, neither citalopram [F(1,44)=1.47, p = 0.232] nor reboxetine [F(1,44)=0.087, p = 0.769] treatment interacted with MDMA's effect (Figure 1d).
The following day, MDMA maintained a significant main effect on conditioned freezing [F(1,76)=17.61, p<0.001]. Post-hoc comparisons within each pre-treatment group that MDMA-induced reductions in freezing observed in animals treated with vehicle prior to MDMA [p=0.001] were prevented when mice were pre-treated with either citalopram [p>0.99] or reboxetine [p=0.08], but not with RTI-336 [p=0.03]. However, only mice treated with citalopram prior to MDMA exhibited significantly greater conditioned freezing that mice that received vehicle prior to MDMA [p<0.001].
MDMA's effects on fear extinction are blocked by chronic daily injections of citalopram
Daily citalopram administration for 22 days significantly altered the effect of MDMA on fear extinction. Extinction training was carried out two days after the final chronic treatment to avoid acute effects of treatment (Figure 2a). During extinction training, mice treated with MDMA exhibited significantly less freezing during the first two sets of 4 CS's if they had not been chronically treated with citalopram. Chronic treatment with citalopram abolished this effect across trials [one-way ANOVA for CS1: F(3,36)=31.46, p<0.001; one-way ANOVA for CS2: F(3,36)=5.59, p=0.003; n=10/group] (Figure 2b). Chronic citalopram treatment also abolished the effect of MDMA on average conditioned freezing for the whole extinction training session [chronic treatment × pre-treatment: F(1,36)=4.51, p=0.041; n=10/group] (Figure 2c). Similar effects were observed the following day during extinction testing, where MDMA-treated mice that received chronic vehicle exhibited significantly less conditioned freezing during the first 4 CS's than groups treated chronically with citalopram or without MDMA [one-way ANOVA for CS1: F(3,36)=3.95; p=0.016; n=10/group] (Figure 2b). On average, across the whole of extinction testing, chronic treatment with citalopram blocked MDMA-induced reductions in conditioned freezing (Figure 2d) [chronic treatment × pre-treatment: F(1,36)=7.74, p=0.009; n=10/group]. Here, chronic treatment with citalopram alone did not affect conditioned freezing [F(1,36)=3.49, p=0.07; n=10/group].
Fig 2. Chronically-administered citalopram inhibits MDMA's effect on fear extinction 48 h later.
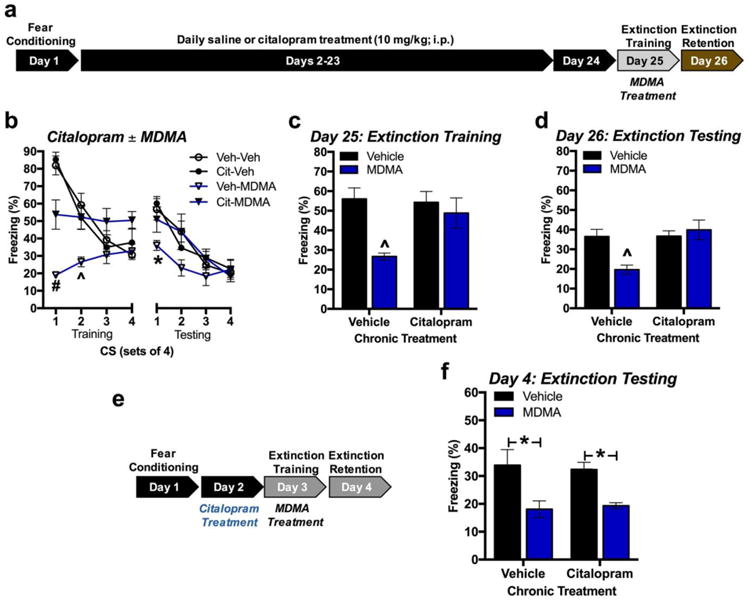
(a) Experimental model. (b) Conditioned freezing across groups of 4 CS re-exposures during extinction training (left) and testing (right) (n=10/group). (c) Average total conditioned freezing to CS during extinction training. Chronically administered citalopram blocked MDMA-induced reductions in conditioned freezing. (d) Average total conditioned freezing to CS during extinction testing. Chronically-administered citalopram significantly obstructed MDMA-induced reductions in conditioned freezing. (e) Experimental model. (f) Average total conditioned freezing during extinction testing. Acute citalopram treatment 24 h prior to extinction training did not alter MDMA‟s lasting effect on conditioned freezing during extinction testing (n=8/group). *p < 0.05; # p < 0.001
To rule out the possibility that residual citalopram remaining from the last day of chronic treatment was affecting fear extinction, a separate group of fear-conditioned mice was treated with vehicle or citalopram 24 h prior to extinction training and MDMA treatment (Figure 2e). Treatment with citalopram 24 h before extinction training did not affect the persistent reductions in conditioned freezing observed in MDMA-treated mice during extinction testing (Figure 2f) [F(1,28)=17.56, p=0.0003 for main effect of MDMA treatment; F(1,28)=0.001, p=0.970 for main effect of citalopram treatment ; F(1,28)=0.16, p=0.696 for citalopram × MDMA treatments].
Transporter-mediated 5-HT release is insufficient to enhance extinction: a role for the 5-HT2A receptor
Administration of a range of doses of fenfluramine – a compound that selectively releases 5-HT through the 5-HTT – 30 min prior to extinction training dose-dependently increased conditioned freezing during extinction training [F(3,28)=3.46, p=0.029; n=8/group]. These effects were not maintained during extinction training the following day when mice were re-exposed to 14 CS tones, as animals previously treated with vehicle exhibited similar levels of conditioned freezing in comparison to all fenfluramine treatment groups (Figure 3a) [F(3,28)=0.48, p = 0.696; n=8/group].
Fig 3. Effect of fenfluramine and the 5-HT2A receptor on fear memory extinction.
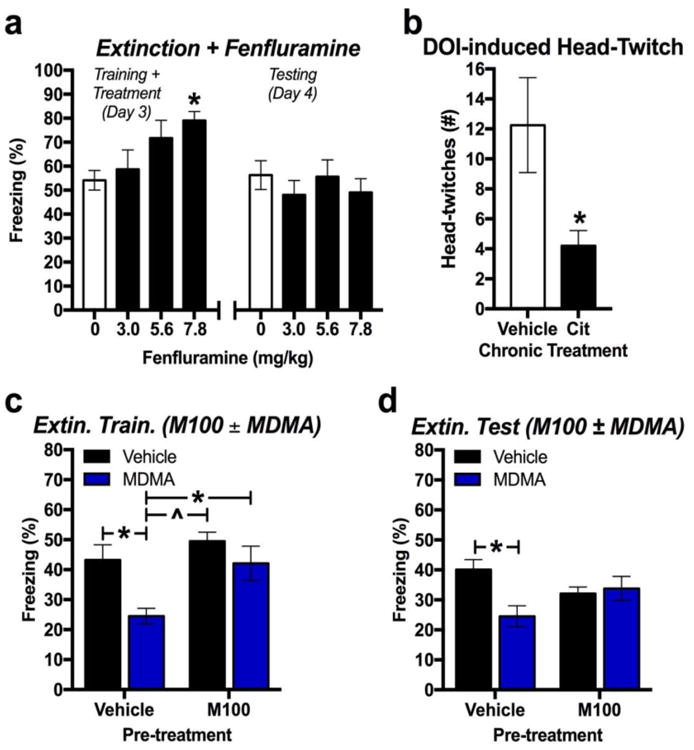
(a) Total conditioned freezing to CS re-exposure during extinction training (left) and testing (right) when fenfluramine was administered 30 min prior to extinction training (n=8/group). (b) Total number of head-twitches induced by the 5-HT2AR agonist DOI (1 mg/kg) 48 h after 22 days of daily vehicle or citalopram administration (n=5/group). (c) Total conditioned freezing to CS re-exposure during extinction training when the 5-HT2AR antagonist M100 was administered 30 min prior to MDMA treatment (n=8/group). (d) Total conditioned freezing to CS re-exposure during extinction testing the day after extinction training (n=8/group). *p < 0.05; ^p < 0.01; #p < 0.001
Next, we explored the effect of chronic citalopram treatment on a behavior dependent on the 5-HT2A receptor (5-HT2AR). Mice were treated with the 5-HT2AR agonist DOI (1 mg/kg, i.p.) 48 h after the last dose of chronic daily treatment with either vehicle or citalopram. Mice treated chronically with citalopram exhibited significantly less DOI-induced head-twitch behavior across a 10 min observation period (Figure 3b) [t(8)=2.41, p=0.042; n=5/group]. To explore the role of the 5-HT2AR in MDMA's effect on extinction, the selective 5-HT2AR antagonist M100 was administered 30 min prior to MDMA treatment. M100 (0.6 mg/kg) obstructed MDMA's effect during extinction training (Figure 3c) [F(1,28)=7.69, p=0.01 for main effect of M100; F(1,28)=9.17, p=0.005 for main effect of MDMA; F(1,28)=1.72, p=0.200 for interaction effect; n=8/group) and also significantly attenuated MDMA's effect on conditioned freezing 24 h later (Figure 3d) [F(1,28)=0.038, p=0.846 for main effect of M100; F(1,28)=4.39, p=0.044 for main effect of MDMA; F(1,28)=6.81, p=0.014 for M100 × MDMA treatments; p=8/group].
Chronically-administered citalopram blocks MDMA's enhancement of the extinction of fear-potentiated startle (FPS)
All three doses of MDMA administered prior to extinction training FPS during the training session. One-way ANOVA analysis of the average FPS exhibited during each group of 4 CS presentations during extinction training revealed significant effects of MDMA treatment across the extinction training session (Figure 4b) [CS1: F(3,60)=13.95, p<0.001; CS2: F(3,60)=8.00, p<0.001; CS5: F(3,60)=6.05, p=0.001 for main effects of treatment; n=16/group]. Total average FPS during extinction training was also reduced by all doses of MDMA (Figure 4c) [F(3,60)=8.91, p<0.0001; n=16/group]. However, when retention of extinction was tested the following day, only animals treated with 7.8 mg/kg of MDMA exhibited significantly less FPS (Figure 4d) [F(3,60)=3.45, p=0.022; n=16/group]. Analysis of each individual group of 4 CS presentations during extinction testing revealed a significant effect of treatment only in the final four CS presentations (Figure 4b) [CS10: F(3,60)=0.0024, p=0.002; n=16/group].
Fig 4. MDMA's effects on extinction of fear-potentiated startle (FPS).
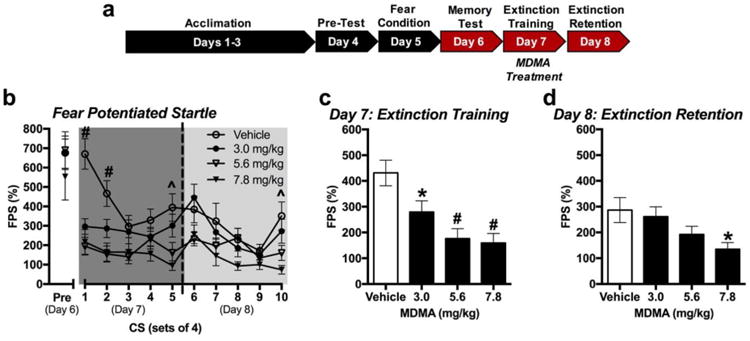
(a) Experimental model. MDMA was administered 20 min prior to extinction training. (b) Average FPS across sets of 4 CS re-exposures during memory testing (Pre), extinction training (Day 7), and extinction testing (Day 8). 7.8 mg/kg treatment resulted in significantly less FPS during CS trials 1,2, 5 and 10 compared to vehicle treatment (n=16/group). (c) Total FPS in response to CS re-exposure during extinction training when MDMA was administered 20 min prior to extinction training. All treatment groups had significantly less FPS than the control vehicle-treated group (n=16/group). (d) Total FPS in response to CS re-exposure during extinction testing. Only 7.8 mg/kg treatment resulted in significantly less FPS than the vehicle-treated group (n=16/group). *p < 0.05; ^p < 0.01; #p < 0.001.
Chronic daily injections of citalopram (10 mg/kg; i.p.) for 22 days did not alter FPS induced by 4 CS presentations during a brief memory test on Day 24 compared to vehicle (Figure 5b). The following day, MDMA treatment (7.8 mg/kg) prior to extinction training significantly reduced FPS regardless of whether mice had been chronically treated with either citalopram or vehicle (Figure 5c) [F(1,44=23.08, p<0.001 for main effect of acute treatment; F(1,44)=0.179, p=0.674 for main effect of chronic treatment; F(1,40)=0.403, p=0.529 for acute × chronic treatment, n=12/group]. When retention of fear extinction was tested in the mice the following day, reductions in FPS facilitated by MDMA were abolished in mice that had been chronically treated with citalopram (Figure 5d) [chronic treatment × acute treatment: F(1,44)=5.28, p=0.026; n=12/group].
Fig 5. Chronically-administered citalopram inhibits MDMA's effect on the extinction of FPS.
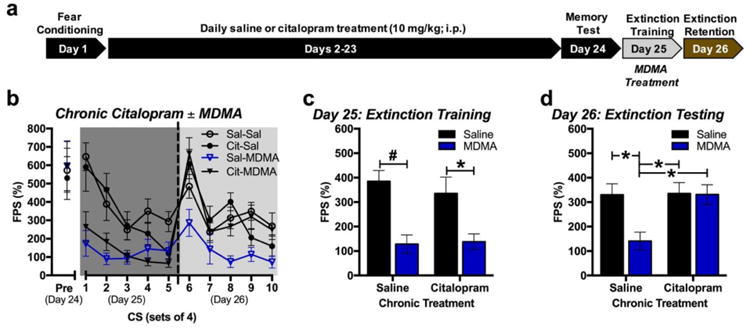
(a) Experimental model. (b) Average FPS across sets of 4 CS re-exposures during memory testing (Pre), extinction training (Day 25), and extinction testing (Day 26) when MDMA (7.8 mg/kg) was administered 20 min prior to extinction training (n=12/group). (c) Total FPS in response to CS re-exposure during extinction training when MDMA or vehicle was administered 20 min prior to extinction training. Both groups treated with MDMA exhibited significantly less FPS then vehicle-treated groups regardless of chronic treatment. (d) Total FPS in response to CS re-exposure during extinction testing. *p < 0.05; ^p < 0.01; #p < 0.001.
Discussion
The acute positive effects of MDMA on mood and affiliative behaviors have long driven interest in its usefulness as an adjunct to psychotherapy (Greer and Tolbert 1986). We have hypothesized that the lasting efficacy of MDMA as an adjunct to psychotherapy for PTSD may result in part from its effects on the extinction of powerful fear memories (Mithoefer et al. 2011; Mithoefer et al. 2013). Using both conditioned freezing and FPS as measures of fear memory extinction, we demonstrate that MDMA-induced enhancements of extinction can be impaired by pharmacologically inhibiting the 5-HTT. We propose that this effect is mediated by indirect inhibition of 5-HT2A receptors. The results of the present study support the hypothesis that MDMA enhances the extinction of fear memory, and also reveal an important role for serotonergic neurotransmission in this effect.
Our observation that administration of MDMA prior to extinction training prevented the expression of FPS during the extinction test is a critical indicator that MDMA improves extinction learning (Myers and Davis 2002), and supports previous observations where MDMA enhanced the extinction of conditioned freezing (Young et al. 2015). Though both freezing behaviors and the acoustic startle reflex rely on distinct neural pathways, the ability for fear memories to modulate both of these behaviors relies on the amygdala (Myers and Davis 2007). We have previously demonstrated that MDMA facilitates the extinction of conditioned freezing specifically in the amygdala, where it increases BDNF mRNA only after extinction training (Young et al. 2015). Interestingly, although expression of conditioned freezing and FPS follow similar trajectories of magnitude when measured simultaneously during fear memory retrieval, FPS appears to extinguish more slowly (Daldrup et al. 2015). We made similar observations in the present study, where extinction training to 16 CS in saline-treated mice resulted in greater relative reductions in conditioned freezing than in FPS. Therefore, it is important that we demonstrated an enhancing effect of 7.8 mg/kg MDMA on the lasting extinction of both conditioned freezing and FPS. Moreover, validation of MDMA's effect on the extinction of FPS is important because it is frequently used as an index of conditioned fear expression and extinction in human investigations, where MDMA is believed to have clinical utility (Briscione et al. 2014; Norrholm et al. 2006).
It is interesting to note that, although administering MDMA prior to extinction training significantly reduces both freezing and FPS during extinction training and testing sessions, these reductions in conditioned fear did not predict lasting improvements in extinction learning. Acute psychostimulant effects of MDMA unrelated to extinction learning are likely to mask conditioned freezing and FPS during extinction training via DA and/or NE. Amphetamine, which releases DA and NE in a similar manner as MDMA, also can also reduce conditioned fear during the extinction session without persistently improving the extinction of conditioned freezing or FPS (Borowski and Kokkinidis 1998; Carmack et al. 2010). 5-HT is unlikely to be involved in reductions of fear behavior during extinction training, as pre-extinction administration of fenfluramine dose-dependently increased conditioned freezing during extinction training. Although reduced fear expression during extinction training does not predict lasting improvements in extinction, those lasting improvements require that MDMA is on-board during the extinction session (Young et al. 2015). While it has been hypothesized that MDMA improves psychotherapy for PTSD through its acute positive effects on mood, these effects might be mitigated by MDMA's anxiogenic effects (Bershad et al. 2016; van Wel et al. 2012). The findings here and elsewhere suggest that subjective or behavioral responses to MDMA during extinction training may not predict lasting fear extinction outcomes.
Although systemically-administered MDMA potently increases extracellular levels of 5-HT in the brain, 5-HTT-mediated increases in 5-HT alone are unlikely to account for MDMA's effect on fear memory extinction (Gudelsky and Nash 1996; Murnane et al. 2010). While citalopram treatment itself selectively increases extracellular 5-HT in the brain (Balu et al. 2013), we and others have observed that neither acute nor chronic treatment with citalopram enhances fear memory extinction (Burghardt et al. 2013; Gunduz-Cinar et al. 2016). Even more strikingly, acute treatment with fenfluramine, which potently releases 5-HT through the 5-HTT (Crespi et al. 1997), did not enhance fear memory extinction on its own in our study. On the other hand, SSRIs that have significant binding affinity at other monoamine transporters can enhance fear extinction in rodents when chronically administered between fear conditioning and extinction training (Gunduz-Cinar et al. 2016; Karpova et al. 2011). While disrupting MDMA's access to 5-HTTs with the highly-selective SSRI citalopram (Sanchez and Hyttel 1999) actually blocked MDMA-induced extinction enhancement, the fact that drugs that specifically increase 5-HT do not improve extinction suggest the requirement of pharmacodynamic mechanisms beyond acute 5-HT release are required.
MDMA affects a variety of neuromodulatory systems and targets in the brain that have been previously associated with fear memory extinction (Green et al. 2003; Singewald et al. 2015). MDMA increases levels of extracellular NE in the prefrontal cortex (Lanteri et al. 2014), where inhibiting beta-adrenergic receptors in the infralimbic sub-region prior to extinction training impairs the retention of fear memory extinction the following day (Mueller et al. 2008). Conversely, extinction retention is enhanced by direct administration of MDMA into the infralimbic cortex (Young et al. 2015). Given that reboxetine, a selective inhibitor of NE transporters (NET), inhibits some physiological and behavioral effects of MDMA (Hysek et al. 2011), it follows that reboxetine pre-treatment might also inhibit MDMA's effect on extinction. However, reboxetine did not alter MDMA's effect on extinction in the current study, nor did it affect extinction learning on its own. Indeed, completely ablating NE signaling does not affect the extinction of cued fear memory (Murchison et al. 2004). MDMA's continued efficacy could be explained by direct binding of MDMA at beta-adrenergic receptors (Battaglia et al. 1988), blockade of which inhibits some of MDMA's cardiovascular effects (Hysek et al. 2010). Inhibition of beta-adrenergic receptors has been previously demonstrated to inhibit the effect of NE on prefrontal cortical neurons believed to be essential for extinction learning (Mueller et al. 2008).
Although MDMA also releases significant levels of DA, other transporter-mediated DA releasers such as amphetamine have not been demonstrated to promote extinction learning here or elsewhere (Borowski and Kokkinidis 1998; Carmack et al. 2010). While one study reported enhancement of contextual fear memory extinction by the DA transporter (DAT) inhibitor methylphenidate, this treatment also increases NE via NET inhibition (Abraham et al. 2012; Kuczenski and Segal 1997). RTI-336 was used block DAT in the current study because of its high selectivity for the DAT over 5-HTT and NET (Carroll et al. 2006). However, DAT only accounts for some of the DA released by MDMA. For example, activation of the 5-HT2A receptor by MDMA appears to contribute to increases in DA release in the striatum (Yamamoto et al. 1995). Given that we observed that 5-HT2A receptor inhibition blocked MDMA's effect on extinction, indirect DA release may in fact play an important role. One target of indirectly released DA that may be important to MDMA's effect on extinction is the D2 receptor. Although MDMA has comparatively very low affinity for the D2 receptor (Battaglia et al. 1988), the D2 receptor is required for some of MDMA's euphoric effects (Liechti and Vollenweider 2000a) and does contribute to normal fear memory extinction (Mueller et al. 2010).
Compensatory changes brought on by chronic SSRI treatment are likely to inhibit MDMA's effect on fear memory extinction because chronic citalopram treatment blocked MDMA-induced extinction enhancements even 48 h after the final treatment. This is expected to be a sufficient amount of time to clear systemic levels of citalopram, which has an estimated half-life of 90 min in mice (Fredricson Overo 1982). Moreover, the dose used in the present study is almost completely eliminated from serum within 6 h (Karlsson et al. 2013). One consistently-observed effect of chronic SSRI treatment is the downregulation of 5-HTT availability and function (Benmansour et al. 1999; Benmansour et al. 2002), which could diminish transporter-mediated release of 5-HT by MDMA. However, we also explored mechanisms besides 5-HT release, given that acutely increasing transporter-mediated 5-HT release with fenfluramine failed to enhance extinction. Pharmacological and genetic manipulations of the 5-HTT consistently alter 5-HT2A receptor function in the amygdala (Bocchio et al. 2015; Jennings et al. 2008). In addition to its effects on the 5-HTT, chronic SSRI treatment downregulates 5-HT2A receptor expression and/or function (Meyer et al. 2001; Sawyer et al. 2012). To determine the effect of our chronic citalopram treatment on 5-HT2A receptor function, we explored its effects on DOI-induced head-twitch behavior, which is mediated specifically through the 5-HT2A receptor (Dougherty and Aloyo 2011). Indeed, DOI-induced head-twitch behavior was significantly reduced 48 h after chronic citalopram treatment. Further supporting the requirement of 5-HT2ARs for MDMA's effect on extinction, blocking 5-HT2ARs with the highly selective antagonist M100 (Kehne et al. 1996) prevented MDMA's lasting enhancement of fear memory extinction. This is consistent with studies demonstrating that the 5HT2A receptor is required for MDMA's behavioral and physiological effects (Liechti et al. 2000; Murnane et al. 2012).
We did not observe lasting impairments in fear memory extinction in response to 5-HT2A receptor antagonist treatment alone, suggesting that 5-HT2A receptors may be sufficient, but not necessary for extinction. While others have observed impaired extinction acquisition in response to 5-HT2A receptor inhibition, they did not explore whether extinction was permanently impaired (Slipczuk et al. 2013; Zhang et al. 2013). Conversely, the extinction of fear memory is consistently enhanced by 5-HT2A receptor agonists (Catlow et al. 2013; Zhang et al. 2013), which also diminish the expression of FPS (Hughes et al. 2012). 5-HT axon terminals and 5-HT2A receptors are densely expressed in the amygdala (Asan et al. 2013; McDonald and Mascagni 2007), where they are particularly important for regulating fear behavior (Bocchio et al. 2015). Moreover, targeting MDMA to the amygdala facilitates fear memory extinction and increases brain-derived neurotrophic factor (Young et al. 2015), the latter of which can be stimulated specifically by 5-HT2A receptors. Besides 5-HT2A receptors, MDMA does bind with notably weaker affinity as a partial agonist at 5-HT1A and 5-HT2C receptor sites (Battaglia et al. 1988). Interestingly, 5-HT1A agonists have been observed to acutely reduce the expression of conditioned fear (de Paula and Leite-Panissi 2016; Risbrough et al. 2003), while antagonists block the psychomotor stimulant and pro-social effects of MDMA in mice (Hunt et al. 2011; Morley et al. 2005). Given the proposed role of therapeutic alliance in the effect of MDMA on PTSD psychotherapy, it is possible that 5-HT1A signaling is important in clinical context. In contrast, we have previously demonstrated here and elsewhere that fear expression during extinction training is irrelevant to lasting extinction improvements by MDMA (Young et al. 2015). Moreover, 5-HT1A antagonism did not inhibit MDMA's effect in the amygdala and has limited effects on the cognitive and subjective effects of MDMA in humans (Hasler et al. 2009).
We hypothesize that chronic SSRI treatment blocks MDMA's effect on fear memory extinction by downregulating 5-HTTs and 5-HT2A receptor signaling. However, it is unclear why acutely-administered citalopram also blocked MDMA's effect, when MDMA's acute 5-HT-releasing activity alone does not appear to enhance extinction. One potential explanation is rapid de-sensitization of the 5-HT2A receptor response to MDMA following citalopram treatment. Systemic injection of citalopram can increase extracellular 5-HT levels in the brain 3-5-fold within 20 min (Balu et al. 2013), and in vitro studies have demonstrated that increasing extracellular 5-HT diminishes the 5-HT2A receptor response to subsequently-administered 5-HT2A receptor agonists within minutes (Berg et al. 2001; Vouret-Craviari et al. 1995). Alternatively, acute and chronic citalopram may disrupt MDMA's effect through distinct non-serotonergic mechanisms, such as the downregulation of NMDA receptor function in the amygdala by chronically-administered citalopram, which would be expected to alter MDMA-induced activity important for extinction (Boyer et al. 1998). Another alternative hypothesis is that the effect of chronically-administered citalopram on MDMA's effect is due to withdrawal effects following cessation of chronic citalopram treatment. However, were this the case, we would expect chronic citalopram treatment alone to alter fear memory expression or extinction, which it did not. The interaction between chronic SSRI treatment and MDMA is of particular importance for translational purposes, given that SSRIs are a commonly-employed treatment for anxiety disorders characterized by impaired fear learning (Koen and Stein 2011).
Although several treatments have been observed to enhance the extinction of conditioned freezing and/or FPS, most of these compounds have not had similar efficacy in human clinical populations (Singewald et al. 2015). MDMA, however, has been observed to improve psychotherapeutic outcomes in PTSD patients (Mithoefer et al. 2011). Although these studies used unstructured psychotherapeutic approaches, and not exposure therapy, the observations in the present study support the hypothesis that reduction of PTSD symptoms by MDMA-assisted psychotherapy is facilitated by enhancing the extinction of the fear response to memories for trauma. Indeed, improvements in PTSD symptoms are achieved by exposure-based therapies that rely on principles of extinction learning (Rauch et al. 2015). However, a portion of patients remain unresponsive to that treatment. We speculate that treatment-resistant individuals may achieve improvements in PTSD symptoms with the combination of MDMA and exposure-based therapy, with the important caveat that concurrent SSRI treatment may obstruct efficacy.
Acknowledgments
With appreciation to the Abraham J. & Phyllis Katz Foundation for support of this research. We thank the animal care and veterinary staff at the Yerkes National Primate Research Center (YNPRC) for maintaining the health and well-being of our research subjects, whom we thank greatly for their contribution. M.B.Y. thanks Dr. Brian Dias for use of valuable behavioral equipment. M.B.Y. also thanks Daniel Curry and Karly Hampshire for assistance with behavioral experiments. The YNPRC is fully accredited by the American Association for Accreditation for Laboratory Animal Care. This research complied with all laws of the United States of America.
Funding and Disclosures: M.B.Y. became employed by Shire Pharmaceuticals (Lexington, MA, USA) after completion of the studies described herein. M.B.Y. was supported by a NIH/NIGMS IRACDA grant K21 GM000680 awarded to Emory University. L.L.H. was supported by NIH/NIDA K05 DA031246. B.O.R. has funding from Wounded Warrior Project, Department of Defense Clinical Trial Grant No.W81XWH-10-1-1045, “Enhancing Exposure Therapy for PTSD: Virtual Reality and Imaginal Exposure with a Cognitive Enhancer”, National Institute of Mental Health Grant No. 1R01MH094757-01, “Prospective Determination of Psychobiological Risk Factors for Posttraumatic Stress,” Brain and Behavior Research Foundation (NARSAD) Distinguished Investigator Grant, “Optimal Dose of early intervention to prevent PTSD”, and McCormick Foundation “Brave Heart: MLB's Welcome Back Veterans SouthEast Initiative.” Dr. Rothbaum receives royalties from Oxford University Press, Guilford, APPI, and Emory University and received one advisory board payment from Genentech. B.W.D. has received research support from Assurex, Axsome, Bristol-Myers Squibb, GlaxoSmithKline, Janssen, NIMH, Otsuka, Pfizer, and Takeda. He has served as a consultant to Pfizer and Medavante. S.D.N, L.M.K., T.J., S.A.R. and C.R. report no funding or conflicts of interest relevant to this manuscript.
References
- Abraham AD, Cunningham CL, Lattal KM. Methylphenidate enhances extinction of contextual fear. Learning & memory (Cold Spring Harbor, NY) 2012;19:67–72. doi: 10.1101/lm.024752.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Asan E, Steinke M, Lesch KP. Serotonergic innervation of the amygdala: targets, receptors, and implications for stress and anxiety. Histochemistry and cell biology. 2013;139:785–813. doi: 10.1007/s00418-013-1081-1. [DOI] [PubMed] [Google Scholar]
- Balu DT, Turner JR, Brookshire BR, Hill-Smith TE, Blendy JA, Lucki I. Brain monoamines and antidepressant-like responses in MRL/MpJ versus C57BL/6J mice. Neuropharmacology. 2013;67:503–10. doi: 10.1016/j.neuropharm.2012.11.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Battaglia G, Brooks BP, Kulsakdinun C, De Souza EB. Pharmacologic profile of MDMA (3,4-methylenedioxymethamphetamine) at various brain recognition sites. European journal of pharmacology. 1988;149:159–63. doi: 10.1016/0014-2999(88)90056-8. [DOI] [PubMed] [Google Scholar]
- Benmansour S, Cecchi M, Morilak DA, Gerhardt GA, Javors MA, Gould GG, Frazer A. Effects of chronic antidepressant treatments on serotonin transporter function, density, and mRNA level. J Neurosci. 1999;19:10494–501. doi: 10.1523/JNEUROSCI.19-23-10494.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benmansour S, Owens WA, Cecchi M, Morilak DA, Frazer A. Serotonin clearance in vivo is altered to a greater extent by antidepressant-induced downregulation of the serotonin transporter than by acute blockade of this transporter. J Neurosci. 2002;22:6766–72. doi: 10.1523/JNEUROSCI.22-15-06766.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berg KA, Stout BD, Maayani S, Clarke WP. Differences in rapid desensitization of 5-hydroxytryptamine2A and 5-hydroxytryptamine2C receptor-mediated phospholipase C activation. The Journal of pharmacology and experimental therapeutics. 2001;299:593–602. [PubMed] [Google Scholar]
- Bershad AK, Weafer JJ, Kirkpatrick MG, Wardle MC, Miller MA, de Wit H. Oxytocin receptor gene variation predicts subjective responses to MDMA. Social neuroscience. 2016;11:592–9. doi: 10.1080/17470919.2016.1143026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bocchio M, Fucsina G, Oikonomidis L, McHugh SB, Bannerman DM, Sharp T, Capogna M. Increased Serotonin Transporter Expression Reduces Fear and Recruitment of Parvalbumin Interneurons of the Amygdala. Neuropsychopharmacology. 2015;40:3015–26. doi: 10.1038/npp.2015.157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Borowski TB, Kokkinidis L. The effects of cocaine, amphetamine, and the dopamine D1 receptor agonist SKF 38393 on fear extinction as measured with potentiated startle: implications for psychomotor stimulant psychosis. Behavioral neuroscience. 1998;112:952–65. doi: 10.1037//0735-7044.112.4.952. [DOI] [PubMed] [Google Scholar]
- Boyer PA, Skolnick P, Fossom LH. Chronic administration of imipramine and citalopram alters the expression of NMDA receptor subunit mRNAs in mouse brain. A quantitative in situ hybridization study. Journal of molecular neuroscience: MN. 1998;10:219–33. doi: 10.1007/BF02761776. [DOI] [PubMed] [Google Scholar]
- Briscione MA, Jovanovic T, Norrholm SD. Conditioned fear associated phenotypes as robust, translational indices of trauma-, stressor-, and anxiety-related behaviors. Frontiers in psychiatry. 2014;5:88. doi: 10.3389/fpsyt.2014.00088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burghardt NS, Bauer EP. Acute and chronic effects of selective serotonin reuptake inhibitor treatment on fear conditioning: implications for underlying fear circuits. Neuroscience. 2013;247:253–72. doi: 10.1016/j.neuroscience.2013.05.050. [DOI] [PubMed] [Google Scholar]
- Burghardt NS, Sigurdsson T, Gorman JM, McEwen BS, LeDoux JE. Chronic antidepressant treatment impairs the acquisition of fear extinction. Biological psychiatry. 2013;73:1078–86. doi: 10.1016/j.biopsych.2012.10.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carmack SA, Wood SC, Anagnostaras SG. Amphetamine and extinction of cued fear. Neuroscience letters. 2010;468:18–22. doi: 10.1016/j.neulet.2009.10.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carroll FI, Howard JL, Howell LL, Fox BS, Kuhar MJ. Development of the dopamine transporter selective RTI-336 as a pharmacotherapy for cocaine abuse. The AAPS Journal. 2006;8:E196–203. doi: 10.1208/aapsj080124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Catlow BJ, Song S, Paredes DA, Kirstein CL, Sanchez-Ramos J. Effects of psilocybin on hippocampal neurogenesis and extinction of trace fear conditioning. Exp Brain Res. 2013;228:481–91. doi: 10.1007/s00221-013-3579-0. [DOI] [PubMed] [Google Scholar]
- Chaudhury D, Colwell CS. Circadian modulation of learning and memory in fear-conditioned mice. Behavioural brain research. 2002;133:95–108. doi: 10.1016/s0166-4328(01)00471-5. [DOI] [PubMed] [Google Scholar]
- Crespi D, Mennini T, Gobbi M. Carrier-dependent and Ca(2+)-dependent 5-HT and dopamine release induced by (+)-amphetamine, 3,4-methylendioxymethamphetamine, p-chloroamphetamine and (+)-fenfluramine. Br J Pharmacol. 1997;121:1735–43. doi: 10.1038/sj.bjp.0701325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daldrup T, Remmes J, Lesting J, Gaburro S, Fendt M, Meuth P, Kloke V, Pape HC, Seidenbecher T. Expression of freezing and fear-potentiated startle during sustained fear in mice. Genes, brain, and behavior. 2015;14:281–91. doi: 10.1111/gbb.12211. [DOI] [PubMed] [Google Scholar]
- Davis M. Neural systems involved in fear and anxiety measured with fear-potentiated startle. The American psychologist. 2006;61:741–56. doi: 10.1037/0003-066X.61.8.741. [DOI] [PubMed] [Google Scholar]
- de Kleine RA, Rothbaum BO, van Minnen A. Pharmacological enhancement of exposure-based treatment in PTSD: a qualitative review. European journal of psychotraumatology. 2013;4:21626. doi: 10.3402/ejpt.v4i0.21626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Paula BB, Leite-Panissi CR. Distinct effect of 5-HT1A and 5-HT2A receptors in the medial nucleus of the amygdala on tonic immobility behavior. Brain research. 2016;1643:152–8. doi: 10.1016/j.brainres.2016.04.073. [DOI] [PubMed] [Google Scholar]
- Dougherty JP, Aloyo VJ. Pharmacological and behavioral characterization of the 5-HT2A receptor in C57BL/6N mice. Psychopharmacology. 2011;215:581–93. doi: 10.1007/s00213-011-2207-6. [DOI] [PubMed] [Google Scholar]
- Fitzgerald JL, Reid JJ. Effects of methylenedioxymethamphetamine on the release of monoamines from rat brain slices. European journal of pharmacology. 1990;191:217–20. doi: 10.1016/0014-2999(90)94150-v. [DOI] [PubMed] [Google Scholar]
- Fredricson Overo K. Kinetics of citalopram in test animals; drug exposure in safety studies. Progress in neuro-psychopharmacology & biological psychiatry. 1982;6:297–309. doi: 10.1016/s0278-5846(82)80180-2. [DOI] [PubMed] [Google Scholar]
- Green AR, Mechan AO, Elliott JM, O'Shea E, Colado MI. The pharmacology and clinical pharmacology of 3,4-methylenedioxymethamphetamine (MDMA, “ecstasy”) Pharmacol Rev. 2003;55:463–508. doi: 10.1124/pr.55.3.3. [DOI] [PubMed] [Google Scholar]
- Greer G, Tolbert R. Subjective reports of the effects of MDMA in a clinical setting. J Psychoactive Drugs. 1986;18:319–27. doi: 10.1080/02791072.1986.10472364. [DOI] [PubMed] [Google Scholar]
- Gudelsky GA, Nash JF. Carrier-mediated release of serotonin by 3,4-methylenedioxymethamphetamine: implications for serotonin-dopamine interactions. Journal of neurochemistry. 1996;66:243–9. doi: 10.1046/j.1471-4159.1996.66010243.x. [DOI] [PubMed] [Google Scholar]
- Gunduz-Cinar O, Flynn S, Brockway E, Kaugars K, Baldi R, Ramikie TS, Cinar R, Kunos G, Patel S, Holmes A. Fluoxetine Facilitates Fear Extinction Through Amygdala Endocannabinoids. Neuropsychopharmacology. 2016;41:1598–609. doi: 10.1038/npp.2015.318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hasler F, Studerus E, Lindner K, Ludewig S, Vollenweider F. Investigation of serotonin-1A receptor function in the human psychopharmacology of MDMA. Journal of Psychopharmacology. 2009;23:923–935. doi: 10.1177/0269881108094650. [DOI] [PubMed] [Google Scholar]
- Hekmatpanah CR, Peroutka SJ. 5-hydroxytryptamine uptake blockers attenuate the 5-hydroxytryptamine-releasing effect of 3,4-methylenedioxymethamphetamine and related agents. European journal of pharmacology. 1990;177:95–8. doi: 10.1016/0014-2999(90)90555-k. [DOI] [PubMed] [Google Scholar]
- Hughes CR, Tran T, Keele NB. 5-HT2A Receptor Activation Normalizes Exaggerated Fear Behavior in p-Chlorophenylalanine (PCPA)-Treated Rats. Journal of Behavioral and Brain Science. 2012;2:454–462. doi: 10.4236/jbbs.2012.24053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hunt GE, McGregor IS, Cornish JL, Callaghan PD. MDMA-induced c-Fos expression in oxytocin-containing neurons is blocked by pretreatment with the 5-HT-1A receptor antagonist WAY 100635. Brain research bulletin. 2011;86:65–73. doi: 10.1016/j.brainresbull.2011.06.011. [DOI] [PubMed] [Google Scholar]
- Hysek CM, Simmler LD, Ineichen M, Grouzmann E, Hoener MC, Brenneisen R, Huwyler J, Liechti ME. The norepinephrine transporter inhibitor reboxetine reduces stimulant effects of MDMA (“ecstasy”) in humans. Clin Pharmacol Ther. 2011;90:246–55. doi: 10.1038/clpt.2011.78. [DOI] [PubMed] [Google Scholar]
- Hysek CM, Simmler LD, Nicola VG, Vischer N, Donzelli M, Krahenbuhl S, Grouzmann E, Huwyler J, Hoener MC, Liechti ME. Duloxetine inhibits effects of MDMA (“ecstasy”) in vitro and in humans in a randomized placebo-controlled laboratory study. PloS one. 2012;7:e36476. doi: 10.1371/journal.pone.0036476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hysek CM, Vollenweider FX, Liechti ME. Effects of a beta-blocker on the cardiovascular response to MDMA (Ecstasy) Emergency medicine journal: EMJ. 2010;27:586–9. doi: 10.1136/emj.2009.079905. [DOI] [PubMed] [Google Scholar]
- Jennings KA, Sheward WJ, Harmar AJ, Sharp T. Evidence that genetic variation in 5-HT transporter expression is linked to changes in 5-HT2A receptor function. Neuropharmacology. 2008;54:776–83. doi: 10.1016/j.neuropharm.2007.12.001. [DOI] [PubMed] [Google Scholar]
- Karlsson L, Carlsson B, Hiemke C, Ahlner J, Bengtsson F, Schmitt U, Kugelberg FC. Altered brain concentrations of citalopram and escitalopram in P-glycoprotein deficient mice after acute and chronic treatment. European neuropsychopharmacology: the journal of the European College of Neuropsychopharmacology. 2013;23:1636–44. doi: 10.1016/j.euroneuro.2013.01.003. [DOI] [PubMed] [Google Scholar]
- Karpova NN, Pickenhagen A, Lindholm J, Tiraboschi E, Kulesskaya N, Agustsdottir A, Antila H, Popova D, Akamine Y, Bahi A, Sullivan R, Hen R, Drew LJ, Castren E. Fear erasure in mice requires synergy between antidepressant drugs and extinction training. Science. 2011;334:1731–4. doi: 10.1126/science.1214592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kehne JH, Baron BM, Carr AA, Chaney SF, Elands J, Feldman DJ, Frank RA, van Giersbergen PL, McCloskey TC, Johnson MP, McCarty DR, Poirot M, Senyah Y, Siegel BW, Widmaier C. Preclinical characterization of the potential of the putative atypical antipsychotic MDL 100,907 as a potent 5-HT2A antagonist with a favorable CNS safety profile. The Journal of pharmacology and experimental therapeutics. 1996;277:968–81. [PubMed] [Google Scholar]
- Kimmel HL, Negus SS, Wilcox KM, Ewing SB, Stehouwer J, Goodman MM, Votaw JR, Mello NK, Carroll FI, Howell LL. Relationship between rate of drug uptake in brain and behavioral pharmacology of monoamine transporter inhibitors in rhesus monkeys. Pharmacology, biochemistry, and behavior. 2008;90:453–62. doi: 10.1016/j.pbb.2008.03.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koen N, Stein DJ. Pharmacotherapy of anxiety disorders: a critical review. Dialogues in clinical neuroscience. 2011;13:423–37. doi: 10.31887/DCNS.2011.13.4/nkoen. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kreilgaard M, Smith DG, Brennum LT, Sanchez C. Prediction of clinical response based on pharmacokinetic/pharmacodynamic models of 5-hydroxytryptamine reuptake inhibitors in mice. Br J Pharmacol. 2008;155:276–84. doi: 10.1038/bjp.2008.243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuczenski R, Segal DS. Effects of methylphenidate on extracellular dopamine, serotonin, and norepinephrine: comparison with amphetamine. Journal of neurochemistry. 1997;68:2032–7. doi: 10.1046/j.1471-4159.1997.68052032.x. [DOI] [PubMed] [Google Scholar]
- Lanteri C, Doucet EL, Hernandez Vallejo SJ, Godeheu G, Bobadilla AC, Salomon L, Lanfumey L, Tassin JP. Repeated exposure to MDMA triggers long-term plasticity of noradrenergic and serotonergic neurons. Molecular psychiatry. 2014;19:823–33. doi: 10.1038/mp.2013.97. [DOI] [PubMed] [Google Scholar]
- Liechti ME, Saur MR, Gamma A, Hell D, Vollenweider FX. Psychological and physiological effects of MDMA (“Ecstasy”) after pretreatment with the 5-HT(2) antagonist ketanserin in healthy humans. Neuropsychopharmacology. 2000;23:396–404. doi: 10.1016/S0893-133X(00)00126-3. [DOI] [PubMed] [Google Scholar]
- Liechti ME, Vollenweider FX. Acute psychological and physiological effects of MDMA (“Ecstasy”) after haloperidol pretreatment in healthy humans. European neuropsychopharmacology : the journal of the European College of Neuropsychopharmacology. 2000a;10:289–95. doi: 10.1016/s0924-977x(00)00086-9. [DOI] [PubMed] [Google Scholar]
- Liechti ME, Vollenweider FX. The serotonin uptake inhibitor citalopram reduces acute cardiovascular and vegetative effects of 3,4-methylenedioxymethamphetamine (‘Ecstasy’) in healthy volunteers. Journal of psychopharmacology (Oxford, England) 2000b;14:269–74. doi: 10.1177/026988110001400313. [DOI] [PubMed] [Google Scholar]
- McDonald AJ, Mascagni F. Neuronal localization of 5-HT type 2A receptor immunoreactivity in the rat basolateral amygdala. Neuroscience. 2007;146:306–20. doi: 10.1016/j.neuroscience.2007.01.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyer JH, Kapur S, Eisfeld B, Brown GM, Houle S, DaSilva J, Wilson AA, Rafi-Tari S, Mayberg HS, Kennedy SH. The effect of paroxetine on 5-HT(2A) receptors in depression: an [(18)F]setoperone PET imaging study. The American journal of psychiatry. 2001;158:78–85. doi: 10.1176/appi.ajp.158.1.78. [DOI] [PubMed] [Google Scholar]
- Mithoefer MC, Wagner MT, Mithoefer AT, Jerome L, Doblin R. The safety and efficacy of {+/-}3,4-methylenedioxymethamphetamine-assisted psychotherapy in subjects with chronic, treatment-resistant posttraumatic stress disorder: the first randomized controlled pilot study. Journal of psychopharmacology (Oxford, England) 2011;25:439–52. doi: 10.1177/0269881110378371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mithoefer MC, Wagner MT, Mithoefer AT, Jerome L, Martin SF, Yazar-Klosinski B, Michel Y, Brewerton TD, Doblin R. Durability of improvement in post-traumatic stress disorder symptoms and absence of harmful effects or drug dependency after 3,4-methylenedioxymethamphetamine-assisted psychotherapy: a prospective long-term follow-up study. Journal of psychopharmacology (Oxford, England) 2013;27:28–39. doi: 10.1177/0269881112456611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morley KC, Arnold JC, McGregor IS. Serotonin (1A) receptor involvement in acute 3,4-methylenedioxymethamphetamine (MDMA) facilitation of social interaction in the rat. Progress in neuro-psychopharmacology & biological psychiatry. 2005;29:648–57. doi: 10.1016/j.pnpbp.2005.04.009. [DOI] [PubMed] [Google Scholar]
- Mueller D, Bravo-Rivera C, Quirk GJ. Infralimbic D2 receptors are necessary for fear extinction and extinction-related tone responses. Biological psychiatry. 2010;68:1055–60. doi: 10.1016/j.biopsych.2010.08.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mueller D, Cahill SP. Noradrenergic modulation of extinction learning and exposure therapy. Behavioural brain research. 2010;208:1–11. doi: 10.1016/j.bbr.2009.11.025. [DOI] [PubMed] [Google Scholar]
- Mueller D, Porter JT, Quirk GJ. Noradrenergic signaling in infralimbic cortex increases cell excitability and strengthens memory for fear extinction. J Neurosci. 2008;28:369–75. doi: 10.1523/JNEUROSCI.3248-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murchison CF, Zhang XY, Zhang WP, Ouyang M, Lee A, Thomas SA. A distinct role for norepinephrine in memory retrieval. Cell. 2004;117:131–43. doi: 10.1016/s0092-8674(04)00259-4. [DOI] [PubMed] [Google Scholar]
- Murnane KS, Fantegrossi WE, Godfrey JR, Banks ML, Howell LL. Endocrine and neurochemical effects of 3,4-methylenedioxymethamphetamine and its stereoisomers in rhesus monkeys. The Journal of pharmacology and experimental therapeutics. 2010;334:642–50. doi: 10.1124/jpet.110.166595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murnane KS, Kimmel HL, Rice KC, Howell LL. The neuropharmacology of prolactin secretion elicited by 3,4-methylenedioxymethamphetamine (“ecstasy”): a concurrent microdialysis and plasma analysis study. Hormones and behavior. 2012;61:181–90. doi: 10.1016/j.yhbeh.2011.10.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Myers KM, Davis M. Behavioral and neural analysis of extinction. Neuron. 2002;36:567–84. doi: 10.1016/s0896-6273(02)01064-4. [DOI] [PubMed] [Google Scholar]
- Myers KM, Davis M. Mechanisms of fear extinction. Molecular psychiatry. 2007;12:120–50. doi: 10.1038/sj.mp.4001939. [DOI] [PubMed] [Google Scholar]
- Norrholm SD, Jovanovic T, Vervliet B, Myers KM, Davis M, Rothbaum BO, Duncan EJ. Conditioned fear extinction and reinstatement in a human fear-potentiated startle paradigm. Learning & memory (Cold Spring Harbor, NY) 2006;13:681–5. doi: 10.1101/lm.393906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rauch SA, King AP, Abelson J, Tuerk PW, Smith E, Rothbaum BO, Clifton E, Defever A, Liberzon I. Biological and symptom changes in posttraumatic stress disorder treatment: a randomized clinical trial. Depression and anxiety. 2015;32:204–12. doi: 10.1002/da.22331. [DOI] [PubMed] [Google Scholar]
- Risbrough VB, Brodkin JD, Geyer MA. GABA-A and 5-HT1A receptor agonists block expression of fear-potentiated startle in mice. Neuropsychopharmacology. 2003;28:654–63. doi: 10.1038/sj.npp.1300079. [DOI] [PubMed] [Google Scholar]
- Rothbaum BO, Davis M. Applying learning principles to the treatment of post-trauma reactions. Ann N Y Acad Sci. 2003;1008:112–21. doi: 10.1196/annals.1301.012. [DOI] [PubMed] [Google Scholar]
- Rothbaum BO, Schwartz AC. Exposure therapy for posttraumatic stress disorder. Am J Psychother. 2002;56:59–75. doi: 10.1176/appi.psychotherapy.2002.56.1.59. [DOI] [PubMed] [Google Scholar]
- Sanchez C, Hyttel J. Comparison of the effects of antidepressants and their metabolites on reuptake of biogenic amines and on receptor binding. Cellular and molecular neurobiology. 1999;19:467–89. doi: 10.1023/A:1006986824213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sawyer EK, Mun J, Nye JA, Kimmel HL, Voll RJ, Stehouwer JS, Rice KC, Goodman MM, Howell LL. Neurobiological changes mediating the effects of chronic fluoxetine on cocaine use. Neuropsychopharmacology. 2012;37:1816–24. doi: 10.1038/npp.2012.29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singewald N, Schmuckermair C, Whittle N, Holmes A, Ressler KJ. Pharmacology of cognitive enhancers for exposure-based therapy of fear, anxiety and trauma-related disorders. Pharmacology & therapeutics. 2015;149:150–90. doi: 10.1016/j.pharmthera.2014.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Slipczuk L, Tomaiuolo M, Garagoli F, Weisstaub N, Katche C, Bekinschtein P, Medina JH. Attenuating the persistence of fear memory storage using a single dose of antidepressant. Molecular psychiatry. 2013;18:7–8. doi: 10.1038/mp.2012.4. [DOI] [PubMed] [Google Scholar]
- Smith KS, Meloni EG, Myers KM, Van't Veer A, Carlezon WA, Jr, Rudolph U. Reduction of fear-potentiated startle by benzodiazepines in C57BL/6J mice. Psychopharmacology. 2011;213:697–706. doi: 10.1007/s00213-010-2026-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strolin Benedetti M, Frigerio E, Tocchetti P, Brianceschi G, Castelli MG, Pellizzoni C, Dostert P. Stereoselective and species-dependent kinetics of reboxetine in mouse and rat. Chirality. 1995;7:285–9. doi: 10.1002/chir.530070416. [DOI] [PubMed] [Google Scholar]
- van Wel JH, Kuypers KP, Theunissen EL, Bosker WM, Bakker K, Ramaekers JG. Effects of acute MDMA intoxication on mood and impulsivity: role of the 5-HT2 and 5-HT1 receptors. PloS one. 2012;7:e40187. doi: 10.1371/journal.pone.0040187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vouret-Craviari V, Auberger P, Pouyssegur J, Van Obberghen-Schilling E. Distinct mechanisms regulate 5-HT2 and thrombin receptor desensitization. The Journal of biological chemistry. 1995;270:4813–21. doi: 10.1074/jbc.270.9.4813. [DOI] [PubMed] [Google Scholar]
- Yamamoto BK, Nash JF, Gudelsky GA. Modulation of methylenedioxymethamphetamine-induced striatal dopamine release by the interaction between serotonin and gamma-aminobutyric acid in the substantia nigra. The Journal of pharmacology and experimental therapeutics. 1995;273:1063–70. [PubMed] [Google Scholar]
- Young MB, Andero R, Ressler KJ, Howell LL. 3,4-Methylenedioxymethamphetamine facilitates fear extinction learning. Translational psychiatry. 2015;5:e634. doi: 10.1038/tp.2015.138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang G, Asgeirsdottir HN, Cohen SJ, Munchow AH, Barrera MP, Stackman RW., Jr Stimulation of serotonin 2A receptors facilitates consolidation and extinction of fear memory in C57BL/6J mice. Neuropharmacology. 2013;64:403–13. doi: 10.1016/j.neuropharm.2012.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]