

Abstract
Objective:
Intrauterine adhesion (IUA) is a major health problem that causes infertility, menstrual irregularities, and recurrent pregnancy losses in women. Unfortunately, treatments for IUA are limited, and there are currently no effective strategies for preventing IUA recurrence. In this review, we introduced the role of Hippo signaling in the normal endometrium and IUA and described the mechanisms by which the Hippo pathway integrates with the Wnt and transforming growth factor-β (TGF-β) signaling pathways to form an intricate network governing the development of fibrosis.
Data Sources:
Original research articles in English that were published until July 2017 were collected from the PubMed database.
Study Selection:
Literature search was conducted using the search terms “endometrial fibrosis OR fibrosis AND or OR intrauterine adhesion OR Asherman syndrome OR IUA,” “Hippo AND or OR Hippo/TAZ,” “TGF-β,” and “Wnt.” Related original research articles were included in the comprehensive analysis.
Results:
Endometrial fibrosis is recognized as a key pathological event in the development of IUA, which is characterized by epithelial/fibroblast–myofibroblast transition. Myofibroblasts play crucial roles in the pathogenesis of fibrous scarring, and myofibroblast differentiation can be triggered by multiple signaling pathways. Hippo signaling is a critical regulator of the epithelial/fibroblast–myofibroblast transition and α-smooth muscle actin, which exhibits a specific spatiotemporal expression in the endometrium.
Conclusions:
Hippo signaling plays a critical role in fibrous diseases and participates in cross talks with Wnt and TGF-β signaling. Our findings not only contributed to knowledge on the pathogenesis of endometrial fibrosis, but can also serve as a useful resource for developing specific molecular inhibitors for IUA treatment and prevention.
Keywords: Endometrial Fibrosis, Hippo Signaling, Intrauterine Adhesion, Transforming Growth Factor-β; Wnt Signaling
INTRODUCTION
More than half a century has passed since Asherman[1] first described the frequency, etiology, and symptoms of intrauterine adhesion (IUA) in 1948. Since then, a numerous studies addressing the etiology and treatment of IUAs have been conducted. IUA refers to the destruction of the basal layer of the endometrium and is attributed to multiple causes.[2] These lead to adhesion between the uterine walls, which in turn results in clinical manifestations, such as amenorrhea, hypomenorrhea, pelvic pain, infertility, and habitual abortion.[3,4] IUAs generally occur after curettage and various forms of hysteroscopic surgery, including hysteroscopic resection of multiple fibroids and resection of the septa.[5] The primary causes of IUA include trauma to the basal layer of the endometrium and uterine infections.[6] Histologically, IUA is defined as the condition in which the endometrium becomes fibrosed, which leads to impaired endometrium function, uterine cavity deformation, and stenosis.[2] In IUA, the endometrial stroma is mainly replaced by large fibrous tissues. IUA is characterized by the destruction of the border between the basal and functional layers of the endometrium. In addition, the functional layer no longer responds to hormone stimulation, and fibrous scarring occurs across the uterine cavity. Vascularity might be abundant and may comprise thin-walled and dilated vessels; however, in most cases, the tissues become avascular.[7] Fibrosis may limit uterine myometrial activity and reduce the perfusion of estrogen and progesterone, eventually resulting in atrophy.[7]
The treatments for IUAs include adhesiolysis and preventing the recurrence of adhesion. The primary objective of treatment is to restore the normal shape and volume of the uterine cavity, and the secondary goals of intervention include the treatment of associated symptoms (i.e., infertility and amenorrhea) and prevention of recurrence of adhesion.[8] With the application of intrauterine devices and antibiotics, the probability of recurrence of uterine cavity adhesion after treatment is gradually reduced.[9] In the near future, improvements in infertility outcomes, including pregnancy rates, and live birth rates are expected to serve as the primary indicators for defining complete resolution of IUAs. However, for patients with moderate-to-severe IUAs, recurrence of adhesion after hysteroscopic adhesiolysis still has a high incidence and remains difficult to treat. Moreover, the effects of IUA treatment on reproductive outcomes remain to be established.
Intrauterine devices and high doses of estrogen therapy are often used to prevent the recurrence of uterine adhesions and to promote endometrial repair.[7,10] However, it is worth noting that estrogen therapy alone has poor efficacy and is usually accompanied by adverse effects, which significantly limit its widespread application. Endometrial fibrosis has been implicated in the development of IUA. Thus, the prevention of endometrial fibrosis is fundamental to IUA treatment. The Hippo signaling pathway has been previously demonstrated to participate in the fibrogenesis of various organs and cross talks with other signaling pathways, such as the transforming growth factor-β (TGF-β) and Wnt/β-catenin signaling pathways, through a variety of mechanisms.[11,12] In this review, we discuss the role of the Hippo pathway in endometrial fibrosis in IUAs and focus on the mechanisms by which the Hippo pathway integrates and interacts with other signaling pathways.
HIPPO SIGNALING PATHWAY
The Hippo pathway, which was first discovered in Drosophila, is a highly conserved signaling pathway that plays essential roles in growth control. The first core components of the Hippo pathway to be identified include the tumor suppressor genes such as Warts, Hippo, and Salvador, which belong to the hyperplastic group of Drosophila tumor suppressors.[13] Mutations in the Warts, Hippo, and Salvador genes lead to extensive tissue overgrowth.[13] Hippo interacts with Salvador to phosphorylate and activate the complex formed by Wts and Mats.[14] Yorkie is the transcriptional co-activator of the Hippo pathway. Subsequent studies have demonstrated that Yorkie can be directly phosphorylated and inhibited by Wts.[15,16]
The core components and downstream effectors of the Hippo pathway in Drosophila are highly conserved in mammals and include the following: Mst1/2 (Hippo homolog), Sav1 (Sav homolog), MOB1 (homolog of Mats), Lats1/2 (Wts homologs), TAZ (transcriptional co-activator with PDZ-binding motif), and YAP (Yorkie homolog).[13] Mst1/2 activates and phosphorylates Lats1/2 and MOB1, which in turn associate to form a complex that phosphorylates and inhibits the transcription of the co-factors TAZ and YAP.[17,18] As transcriptional co-activators, TAZ and YAP themselves cannot directly bind DNA but must form complexes with DNA-binding transcription factors, such as TEADs, Smads, and TBX5, to regulate the expression of downstream targets.[19,20] Activation of MST and LATS kinases is considered as an indicator of an activated Hippo pathway, which leads to the phosphorylation of TAZ and YAP. In turn, phosphorylated TAZ and YAP are exported to the nucleus and are retained in the cytoplasm, making them unable to bind TEAD factors and inhibiting the expression of target genes.[17,21] The active Hippo pathway inhibits the activities of TAZ and YAP and thus suppresses the expression of genes driven by TAZ/YAP. Conversely, when the Hippo pathway is repressed, TAZ and YAP accumulate in the nucleus and drive target gene expression by forming complexes with TEAD factors.[21,22] Thus, the Hippo pathway primarily acts by restraining the nuclear import of TAZ and YAP.
HIPPO PATHWAY AND NORMAL ENDOMETRIAL DEVELOPMENT
The human endometrium is a unique tissue that undergoes regular processes of substantial and periodic proliferation, differentiation, sloughing, and renewal during a woman's reproductive years.[23] Endometrial autocrine and paracrine factors change throughout the menstrual cycle and are thought to play vital roles in the regulation of endometrial physiology.[24] Previous studies have confirmed that members of the Hippo pathway are expressed in the endometrium and that their expression patterns change throughout the menstrual cycle, suggesting that the Hippo pathway is involved in regulating endometrial physiology.[25] Compared to the proliferative phase, Hippo and LATS kinases are upregulated during the secretory phase of the menstrual cycle.[25] Meanwhile, TAZ expression is downregulated during decidualization of endometrial stromal cells.[26,27]
Endometrial stromal cells undergo morphological and functional changes from the proliferative phase to the secretory phase. Stromal cells differentiate into decidual cells, and the endometrium undergoes decidualization during embryo implantation. Decidualization spreads from the implantation site to the whole uterine cavity, and the decidua serves as a contact inhibition barrier that prevents invasion by trophoblast cells for the whole duration of pregnancy.[27,28] Decidualization is vital for the establishment and maintenance of pregnancy. However, the molecular mechanisms that regulate the transformation of endometrial stromal cells during decidualization are still unknown. As a contact inhibitory signaling pathway, the Hippo pathway inhibits cell proliferation and promotes cell apoptosis when cells are completely surrounded by their neighbors.[25,29] Decidualization is the differentiation response of endometrial stromal cells, during which the Hippo pathway is activated and its downstream effector, TAZ, is downregulated along with its disappearance from the nucleus.[27] Moreover, another effector, YAP, was upregulated in human decidual cells and was found to potentially influence the decidualization of endometrial stromal cells.[28]
The Hippo pathway is required for the actual proliferation and differentiation of stem cells and is involved in processes, such as regulation of reproductive stem cell differentiation to improve ovarian aging[30] and epithelial stem cell differentiation to promote lung maturation.[31] Endometrial stem cells are responsible for regenerating the endometrium,[32] and this process is regulated by estrogen. However, estrogen does not directly act on endometrial mesenchymal stem cells, but influence stem cells via paracrine signaling to promote endometrium regrowth.[33] Moreover, TAZ can be activated by estrogen.[34] These observations suggested that estrogen regulates endometrial stem cell differentiation via the Hippo pathway.
HIPPO PATHWAY AND FIBROGENESIS
Recent studies have implicated the Hippo pathway and its transcriptional effectors, TAZ and YAP, as necessary for tissue fibrosis and myofibroblast activation.[12] The Hippo/TAZ signaling pathway has been proven to play a major role in regulating the epithelial–mesenchymal transition (EMT), to promote the expression of α-smooth muscle actin (α-SMA) and connective tissue growth factor (CTGF),[35,36] and to regulate the growth and differentiation of stem cells. As a contact inhibitory signaling pathway, the Hippo/TAZ pathway is related to cell differentiation and organ size control.[25] It is widely believed that the Hippo/TAZ pathway is activated when cells enter the differentiation phase or when an organ approaches its final size, which in turn inhibits cell proliferation and regulates the EMT. TAZ is an important transducer of the Hippo pathway that modulates the EMT and cell proliferation. Activated TAZ translocates from the cytoplasm into the nucleus and binds with various transcription factors, such as TEAD and Smads. TAZ has been suggested to regulate the expression of α-SMA and CTGF.[35,36,37] Activation of the Hippo pathway leads to TAZ phosphorylation and limits cytoplasmic retention,[37] which in turn inhibits TAZ-induced cell proliferation and the EMT process.
EMT has been well established as the key mechanism that mediates fibrosis during tissue repair. The most advanced form of EMT is the epithelial/fibroblast–myofibroblast transition, during which epithelial and stromal cells acquire a myofibroblast phenotype and is characterized by the expression of α-SMA and increased secretion of collagen I.[38,39] The mesenchymal–epithelial transition (MET), the reverse process of EMT, is known to be crucial in counteracting fibrotic processes.[40] The balance between EMT and MET critically determines the convalescence of organ fibrosis. A previous study reported that promotion of MET and reversal of EMT could relieve liver fibrosis.[40] EMT and MET were also demonstrated to be involved in the fibrosis of pelvic endometriosis,[41] conversion of endometriotic stromal cells into myofibroblasts, and the upregulation of α-SMA and collagen I.[39] Notably, the Hippo signaling pathway was also demonstrated to play a role in the development and progression of endometriosis, which modulates endometrial stromal cell activity through transcriptional regulation of CTGF.[42]
CROSS TALK BETWEEN HIPPO AND WNT SIGNALING
It is generally accepted that the Wnt gene family (especially Wnt4, Wnt5a, and Wnt7a) is highly involved in endometrial development.[43] The Wnt family of proteins comprises a large group of highly conserved and secreted glycoproteins that act via autocrine or paracrine mechanisms. Wnt proteins stimulate intracellular signaling pathways, regulate gene expression in the nucleus, and determine cell fate. Wnt4 and Wnt5a are involved in decidualization of endometrial stromal cells.[44] Wnt7a knockout mice were demonstrated to have thinner endometria and lack the endometrial glands.[44]
The involvement of Wnt in myofibroblast biology was first demonstrated one decade ago. Since then, numerous studies have suggested the critical role of the Wnt pathway in the fibrogenesis of organs, such as the heart and lungs.[45,46] The Wnt/β-catenin pathway is aberrantly activated in those fibrotic diseases. Aberrant activation of the Wnt/β-catenin pathway is implicated in endometriotic tissues and was demonstrated to mediate fibrogenesis in endometriosis.[47] Silence or inhibition of β-catenin attenuates TGF-β-induced fibrotic process[48] and decreases the expression of α-SMA, collagen I, and CTGF in endometriotic stromal cells.[39]
The first evidence of cross talk between Hippo and Wnt pathway was reported by Varelas et al.,[49] who demonstrated that TAZ inhibits DVL phosphorylation, which in turn inhibits the Wnt/β-catenin signaling pathway. Their findings also suggested that overexpression of LATS or MST leads to cytoplasmic localization of TAZ, which inhibits Wnt3a-mediated activity. Increased expression of nuclear β-catenin has been detected in the kidneys of TAZ knockout mice.[49,50] Moreover, increased heart size and higher levels of nuclear β-catenin were found in heart-specific Salvador knockout mice, which suggested the inhibitory role of Hippo pathway in the Wnt/β-catenin pathway.[51] However, Azzolin et al. demonstrated that β-catenin can regulate the expression of TAZ.[52,53] In the absence of Wnt, phosphorylated β-catenin forms a destruction complex with GSK3, Axin, and APC that is required for the association of TAZ with the β-TrCP/E3 ligase complex. In the presence of Wnt, β-catenin phosphorylation is blocked, which leads to dissociation of both TAZ and β-catenin from the destruction complex, which subsequently promotes the expression of the target genes.
CROSS TALK BETWEEN HIPPO AND TRANSFORMING GROWTH FACTOR-β SIGNALING
During the menstrual cycle, members of the TGF-β superfamily are dynamically expressed in the endometrium under hormonal regulation. The stroma and glands of endometrium secrete TGF-β into the uterine fluid.[54] In addition, the protein and mRNA expression of TGF-β is increased during menstruation, which suggests that TGF-β is involved in the initiation of menstruation.[54] Furthermore, TGF-β plays a specific role in the occurrence and aggravation of IUA. Aberrant upregulation of TGF-β is observed in the endometrium during IUA, and the observed increase in TGF-β was found to be positively correlated with the degree of IUA.[55]
TGF-β has been considered as a key mediator for fibrosis.[56] It plays a vital role in modulating wound healing and tissue repair. TGF-β activation is associated with excessive scaring and fibrosis, which are particularly pronounced at wound sites,[57] and blockade of TGF-β activity could reduce scaring.[58] TGF-β upregulates the expression of extracellular matrix (ECM) proteins and α-SMA via Smad2/3 and is implicated in the progression of multiple fibrotic diseases, including idiopathic pulmonary fibrosis and renal fibrosis.[59] It is well known that fibrosis is primarily mediated by the epithelial/fibroblast–myofibroblast transition.[60] Most ECM components, such as collagens, are produced by myofibroblasts. During fibrosis, myofibroblasts aberrantly assemble in the wound site, leading to the formation of fibrous scar tissue. In fibrous scar tissue, the highly crisscrossed structure of collagen makes the fibrous scar highly resistant to protease degradation, which results in irreversible fibrosis and damage of tissue architecture.[61] Although the biological functions of myofibroblast have been well recognized, to date, no strategies have been established to reverse fibrosis.
TGF-β signaling is involved in the differentiation of fibroblasts into myofibroblasts[62] and is a key regulator of myofibroblast biology.[61] Recent studies have also revealed cooperative interactions between YAP and TGF-β-regulated Smads that couple mechanotransduction in fibroblasts to TGF-β-induced profibrotic gene expression programs that are important in fibrosis.[63] Therefore, TGF-β signaling is likely to be regulated by Hippo signaling. When Hippo signaling is activated, cytoplasmic localization of TAZ prevents the interaction of TGF-β-regulated Smads with TAZ and restricts Smads’ nuclear accumulation and activity.[17] In turn, Smads blunts transcriptional responses to TGF-β, thereby inhibiting TGF-β-induced EMT. Hippo signaling is mediated by LATS kinases. Knockdown of LATS kinases was found to restore nuclear localization of TAZ and TGF-β/Smads signaling.[64] However, as a key aspect of this cross talk, TGF-β mediated TAZ upregulation via a Smad3-independent mechanism during experimental kidney fibrosis.[65] Moreover, TAZ-induced upregulation of collagen I could be potentiated by TGF-β.[12] Accumulating evidence has suggested that, during fibrosis, the TGF-β and Hippo signaling pathways work not as isolated entities but as integrated signaling networks that form the molecular framework.[61] TAZ knockdown or deficiency was found to attenuate myofibroblast response to TGF-β signaling, reduce matrix synthesis and contractile capability, and downregulate αSMA expression.[66]
CONCLUSIONS AND FUTURE PERSPECTIVES
Despite considerable advances in the etiology and treatment of IUAs, the cellular and molecular mechanisms underlying the development of IUAs remain unclear. Furthermore, no efficient therapy has been developed so far to prevent IUAs’ recurrence. Further research efforts must focus on elucidating the molecular mechanisms underlying endometrial fibrosis and strategies to reverse fibrogenesis. Recent advancements in the field of Hippo signaling have suggested its important role in fibrous diseases. Hippo signaling does not act alone but is involved in cross talk with Wnt and TGF-β signaling components [Figure 1]. The interaction among these pathways is the reason why inhibition of a single molecular target often does not achieve the desired therapeutic effect. Our findings not only contribute to the current knowledge on the pathogenesis of endometrial fibrosis, but also highlight the necessity to develop highly specific inhibitors that target the protein–protein interactions at the intersection of multiple signaling pathways.
Figure 1.
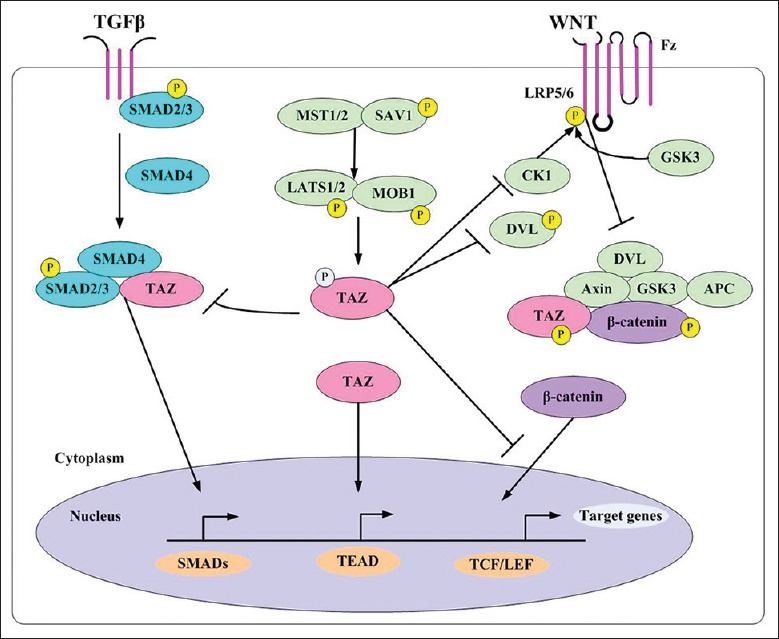
Cross talk among the Hippo, Wnt, and TGF-β signaling pathways. TGF-β: Transforming growth factor-β.
Financial support and sponsorship
This work was supported by grants from the National Natural Science Foundation of China (No. 81601236 and No. 81471505).
Conflicts of interest
There are no conflicts of interest.
Footnotes
Edited by: Qiang Shi
REFERENCES
- 1.Asherman JG. Amenorrhoea traumatica (atretica) J Obstet Gynaecol Br Emp. 1948;55:23–30. doi: 10.1111/j.1471-0528.1948.tb07045.x. doi: 10.1111/j.1471-0528.1948.tb07045.x. [DOI] [PubMed] [Google Scholar]
- 2.Deans R, Abbott J. Review of intrauterine adhesions. J Minim Invasive Gynecol. 2010;17:555–69. doi: 10.1016/j.jmig.2010.04.016. doi: 10.1016/j.jmig.2010.04.016. [DOI] [PubMed] [Google Scholar]
- 3.Myers EM, Hurst BS. Comprehensive management of severe Asherman syndrome and amenorrhea. Fertil Steril. 2012;97:160–4. doi: 10.1016/j.fertnstert.2011.10.036. doi: 10.1016/j.fertnstert.2011.10.036. [DOI] [PubMed] [Google Scholar]
- 4.Xiao S, Wan Y, Xue M, Zeng X, Xiao F, Xu D, et al. Etiology, treatment, and reproductive prognosis of women with moderate-to-severe intrauterine adhesions. Int J Gynaecol Obstet. 2014;125:121–4. doi: 10.1016/j.ijgo.2013.10.026. doi: 10.1016/j.ijgo.2013.10.026. [DOI] [PubMed] [Google Scholar]
- 5.Zhu Y, Hu J, Yu T, Ren Y, Hu L. High molecular weight hyaluronic acid inhibits fibrosis of endometrium. Med Sci Monit. 2016;22:3438–45. doi: 10.12659/MSM.896028. doi: 10.12659/MSM.896028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Healy MW, Schexnayder B, Connell MT, Terry N, DeCherney AH, Csokmay JM, et al. Intrauterine adhesion prevention after hysteroscopy: A systematic review and meta-analysis. Am J Obstet Gynecol. 2016;215:267–75.e7. doi: 10.1016/j.ajog.2016.05.001. doi: 10.1016/j.ajog.2016.05.001. [DOI] [PubMed] [Google Scholar]
- 7.Yu D, Wong YM, Cheong Y, Xia E, Li TC. Asherman syndrome – One century later. Fertil Steril. 2008;89:759–79. doi: 10.1016/j.fertnstert.2008.02.096. doi: 10.1016/j.fertnstert.2008.02.096. [DOI] [PubMed] [Google Scholar]
- 8.Zupi E, Centini G, Lazzeri L. Asherman syndrome: An unsolved clinical definition and management. Fertil Steril. 2015;104:1380–1. doi: 10.1016/j.fertnstert.2015.09.036. doi: 10.1016/j.fertnstert.2015.09.036. [DOI] [PubMed] [Google Scholar]
- 9.Pabuccu R, Onalan G, Kaya C, Selam B, Ceyhan T, Ornek T, et al. Efficiency and pregnancy outcome of serial intrauterine device-guided hysteroscopic adhesiolysis of intrauterine synechiae. Fertil Steril. 2008;90:1973–7. doi: 10.1016/j.fertnstert.2007.06.074. doi: 10.1016/j.fertnstert.2007.06.074. [DOI] [PubMed] [Google Scholar]
- 10.Römer T, Schmidt T, Foth D. Pre- and postoperative hormonal treatment in patients with hysteroscopic surgery. Contrib Gynecol Obstet. 2000;20:1–2. doi: 10.1159/000060283. [DOI] [PubMed] [Google Scholar]
- 11.Seo E, Kim WY, Hur J, Kim H, Nam SA, Choi A, et al. The Hippo-Salvador signaling pathway regulates renal tubulointerstitial fibrosis. Sci Rep. 2016;6:31931. doi: 10.1038/srep31931. doi: 10.1038/srep. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Jorgenson AJ, Choi KM, Sicard D, Smith KM, Hiemer SE, Varelas X, et al. TAZ activation drives fibroblast spheroid growth, expression of profibrotic paracrine signals, and context-dependent ECM gene expression. Am J Physiol Cell Physiol. 2017;312:C277–85. doi: 10.1152/ajpcell.00205.2016. doi: 10.1152/ajpcell.00205.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zhao B, Tumaneng K, Guan KL. The Hippo pathway in organ size control, tissue regeneration and stem cell self-renewal. Nat Cell Biol. 2011;13:877–83. doi: 10.1038/ncb2303. doi: 10.1038/ncb2303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wu S, Huang J, Dong J, Pan D. Hippo encodes a ste-20 family protein kinase that restricts cell proliferation and promotes apoptosis in conjunction with Salvador and warts. Cell. 2003;114:445–56. doi: 10.1016/s0092-8674(03)00549-x. doi: 10.1016/S0092-8674(03)00549-X. [DOI] [PubMed] [Google Scholar]
- 15.Dong J, Feldmann G, Huang J, Wu S, Zhang N, Comerford SA, et al. Elucidation of a universal size-control mechanism in Drosophila and mammals. Cell. 2007;130:1120–33. doi: 10.1016/j.cell.2007.07.019. doi: 10.1016/j.cell.2007.07.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Huang J, Wu S, Barrera J, Matthews K, Pan D. The Hippo signaling pathway coordinately regulates cell proliferation and apoptosis by inactivating Yorkie, the Drosophila Homolog of YAP. Cell. 2005;122:421–34. doi: 10.1016/j.cell.2005.06.007. doi: 10.1016/j.cell.2005.06.007. [DOI] [PubMed] [Google Scholar]
- 17.Narimatsu M, Samavarchi-Tehrani P, Varelas X, Wrana JL. Distinct polarity cues direct taz/Yap and TGFβ receptor localization to differentially control TGFβ-induced Smad signaling. Dev Cell. 2015;32:652–6. doi: 10.1016/j.devcel.2015.02.019. doi: 10.1016/j.devcel.2015.02.019. [DOI] [PubMed] [Google Scholar]
- 18.Praskova M, Xia F, Avruch J. MOBKL1A/MOBKL1B phosphorylation by MST1 and MST2 inhibits cell proliferation. Curr Biol. 2008;18:311–21. doi: 10.1016/j.cub.2008.02.006. doi: 10.1016/j.cub.2008.02.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Alarcón C, Zaromytidou AI, Xi Q, Gao S, Yu J, Fujisawa S, et al. Nuclear CDKs drive Smad transcriptional activation and turnover in BMP and TGF-beta pathways. Cell. 2009;139:757–69. doi: 10.1016/j.cell.2009.09.035. doi: 10.1016/j.cell.2009.09.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wu S, Liu Y, Zheng Y, Dong J, Pan D. The TEAD/TEF family protein scalloped mediates transcriptional output of the Hippo growth-regulatory pathway. Dev Cell. 2008;14:388–98. doi: 10.1016/j.devcel.2008.01.007. doi: 10.1016/j.devcel.2008.01.007. [DOI] [PubMed] [Google Scholar]
- 21.Johnson R, Halder G. The two faces of Hippo: Targeting the Hippo pathway for regenerative medicine and cancer treatment. Nat Rev Drug Discov. 2014;13:63–79. doi: 10.1038/nrd4161. doi: 10.1038/nrd4161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Yu FX, Zhao B, Guan KL. Hippo pathway in organ size control, tissue homeostasis, and cancer. Cell. 2015;163:811–28. doi: 10.1016/j.cell.2015.10.044. doi: 10.1016/j.cell.2015.10.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Shan X, Chan RW, Ng EH, Yeung WS. Spatial and temporal characterization of endometrial mesenchymal stem-like cells activity during the menstrual cycle. Exp Cell Res. 2017;350:184–9. doi: 10.1016/j.yexcr.2016.11.020. doi: 10.1016/j.yexcr.2016.11.020. [DOI] [PubMed] [Google Scholar]
- 24.Singh M, Chaudhry P, Asselin E. Bridging endometrial receptivity and implantation: Network of hormones, cytokines, and growth factors. J Endocrinol. 2011;210:5–14. doi: 10.1530/JOE-10-0461. doi: 10.1530/JOE-10-0461. [DOI] [PubMed] [Google Scholar]
- 25.Strakova Z, Kruss S, Morris K, Reed J. Members of the Hippo pathway are regulated in the uterus during the menstrual cycle. Biol Reprod. 2010;83:363. doi: 10.1093/biolreprod/83.s1.363. [Google Scholar]
- 26.Morris K, Reed J, Kruss S, Yanez D, Strakova Z. TAZ silencing affects diverse targets in uterine stromal cells. Biol Reprod. 2011;85:474. doi: 10.1093/biolreprod/85.s1. [Google Scholar]
- 27.Strakova Z, Reed J, Ihnatovych I. Human transcriptional coactivator with PDZ-binding motif (TAZ) is downregulated during decidualization. Biol Reprod. 2010;82:1112–8. doi: 10.1095/biolreprod.109.081844. doi: 10.1095/biolreprod.109.081844. PMID:20164440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Chen H, Song Y, Yang S, Fu J, Feng X, Huang W, et al. YAP mediates human decidualization of the uterine endometrial stromal cells. Placenta. 2017;53:30–5. doi: 10.1016/j.placenta.2017.03.013. doi: 10.1016/j.placenta.2017.03.013. [DOI] [PubMed] [Google Scholar]
- 29.Gumbiner BM, Kim NG. The Hippo-YAP signaling pathway and contact inhibition of growth. J Cell Sci. 2014;127:709–17. doi: 10.1242/jcs.140103. doi: 10.1242/jcs.140103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Li J, Zhou F, Zheng T, Pan Z, Liang X, Huang J, et al. Ovarian germline stem cells (OGSCs) and the Hippo signaling pathway association with physiological and pathological ovarian aging in mice. Cell Physiol Biochem. 2015;36:1712–24. doi: 10.1159/000430144. doi: 10.1159/000430144. [DOI] [PubMed] [Google Scholar]
- 31.Lange AW, Sridharan A, Xu Y, Stripp BR, Perl AK, Whitsett JA, et al. Hippo/Yap signaling controls epithelial progenitor cell proliferation and differentiation in the embryonic and adult lung. J Mol Cell Biol. 2015;7:35–47. doi: 10.1093/jmcb/mju046. doi: 10.1093/jmcb/mju046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Gargett CE, Ye L. Endometrial reconstruction from stem cells. Fertil Steril. 2012;98:11–20. doi: 10.1016/j.fertnstert.2012.05.004. doi: 10.1016/j.fertnstert.2012.05.004. [DOI] [PubMed] [Google Scholar]
- 33.Hyodo S, Matsubara K, Kameda K, Matsubara Y. Endometrial injury increases side population cells in the uterine endometrium: A decisive role of estrogen. Tohoku J Exp Med. 2011;224:47–55. doi: 10.1620/tjem.224.47. doi: 10.1620/tjem.224.47. [DOI] [PubMed] [Google Scholar]
- 34.Zhou X, Wang S, Wang Z, Feng X, Liu P, Lv XB, et al. Estrogen regulates Hippo signaling via GPER in breast cancer. J Clin Invest. 2015;125:2123–35. doi: 10.1172/JCI79573. doi: 10.1172/JCI79573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Speight P, Nakano H, Kelley TJ, Hinz B, Kapus A. Differential topical susceptibility to TGFβ in intact and injured regions of the epithelium: Key role in myofibroblast transition. Mol Biol Cell. 2013;24:3326–36. doi: 10.1091/mbc.E13-04-0220. doi: 10.1091/mbc.E13-04-0220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Wang Q, Xu Z, An Q, Jiang D, Wang L, Liang B, et al. TAZ promotes epithelial to mesenchymal transition via the upregulation of connective tissue growth factor expression in neuroblastoma cells. Mol Med Rep. 2015;11:982–8. doi: 10.3892/mmr.2014.2818. doi: 10.3892/mmr.2014.2818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lei QY, Zhang H, Zhao B, Zha ZY, Bai F, Pei XH, et al. TAZ promotes cell proliferation and epithelial-mesenchymal transition and is inhibited by the Hippo pathway. Mol Cell Biol. 2008;28:2426–36. doi: 10.1128/MCB.01874-07. doi: 10.1128/MCB.01874-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Guarino M, Tosoni A, Nebuloni M. Direct contribution of epithelium to organ fibrosis: Epithelial-mesenchymal transition. Hum Pathol. 2009;40:1365–76. doi: 10.1016/j.humpath.2009.02.020. doi: 10.1016/j.humpath.2009.02.020. [DOI] [PubMed] [Google Scholar]
- 39.Matsuzaki S, Darcha C. Involvement of the Wnt/β-catenin signaling pathway in the cellular and molecular mechanisms of fibrosis in endometriosis. PLoS One. 2013;8:e76808. doi: 10.1371/journal.pone.0076808. doi: 10.1371/journal.pone.0076808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Shen X, Cheng S, Peng Y, Song H, Li H. Attenuation of early liver fibrosis by herbal compound “Diwu Yanggan” through modulating the balance between epithelial-to-mesenchymal transition and mesenchymal-to-epithelial transition. BMC Complement Altern Med. 2014;14:418. doi: 10.1186/1472-6882-14-418. doi: 10.1186/1472-6882-14- [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Matsuzaki S, Darcha C. Epithelial to mesenchymal transition-like and mesenchymal to epithelial transition-like processes might be involved in the pathogenesis of pelvic endometriosis. Hum Reprod. 2012;27:712–21. doi: 10.1093/humrep/der442. doi: 10.1093/humrep/der442. [DOI] [PubMed] [Google Scholar]
- 42.Song Y, Fu J, Zhou M, Xiao L, Feng X, Chen H, et al. Activated Hippo/Yes-associated protein pathway promotes cell proliferation and anti-apoptosis in endometrial stromal cells of endometriosis. J Clin Endocrinol Metab. 2016;101:1552–61. doi: 10.1210/jc.2016-1120. doi: 10.1210/jc.2016-1120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Tulac S, Nayak NR, Kao LC, Van Waes M, Huang J, Lobo S, et al. Identification, characterization, and regulation of the canonical Wnt signaling pathway in human endometrium. J Clin Endocrinol Metab. 2003;88:3860–6. doi: 10.1210/jc.2003-030494. doi: 10.1210/jc.2003-030494. [DOI] [PubMed] [Google Scholar]
- 44.Matsuoka A, Kizuka F, Lee L, Tamura I, Taniguchi K, Asada H, et al. Progesterone increases manganese superoxide dismutase expression via a cAMP-dependent signaling mediated by noncanonical Wnt5a pathway in human endometrial stromal cells. J Clin Endocrinol Metab. 2010;95:E291–9. doi: 10.1210/jc.2010-0619. doi: 10.1210/jc.2010-0619. [DOI] [PubMed] [Google Scholar]
- 45.Henderson WR, Jr, Chi EY, Ye X, Nguyen C, Tien YT, Zhou B, et al. Inhibition of Wnt/beta-catenin/CREB binding protein (CBP) signaling reverses pulmonary fibrosis. Proc Natl Acad Sci U S A. 2010;107:14309–14. doi: 10.1073/pnas.1001520107. doi: 10.1073/pnas.1001520107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Surendran K, Schiavi S, Hruska KA. Wnt-dependent beta-catenin signaling is activated after unilateral ureteral obstruction, and recombinant secreted frizzled-related protein 4 alters the progression of renal fibrosis. J Am Soc Nephrol. 2005;16:2373–84. doi: 10.1681/ASN.2004110949. doi: 10.1681/ASN.2004110949. [DOI] [PubMed] [Google Scholar]
- 47.Matsuzaki S, Darcha C, Maleysson E, Canis M, Mage G. Impaired down-regulation of E-cadherin and beta-catenin protein expression in endometrial epithelial cells in the mid-secretory endometrium of infertile patients with endometriosis. J Clin Endocrinol Metab. 2010;95:3437–45. doi: 10.1210/jc.2009-2713. doi: 10.1210/jc.2009-2713. [DOI] [PubMed] [Google Scholar]
- 48.Akhmetshina A, Palumbo K, Dees C, Bergmann C, Venalis P, Zerr P, et al. Activation of canonical Wnt signalling is required for TGF-β-mediated fibrosis. Nat Commun. 2012;3:735. doi: 10.1038/ncomms1734. doi: 10.1038/ncomms1734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Varelas X, Miller BW, Sopko R, Song S, Gregorieff A, Fellouse FA, et al. The Hippo pathway regulates Wnt/beta-catenin signaling. Dev Cell. 2010;18:579–91. doi: 10.1016/j.devcel.2010.03.007. doi: 10.1016/j.devcel.2010.03.007. [DOI] [PubMed] [Google Scholar]
- 50.Imajo M, Miyatake K, Iimura A, Miyamoto A, Nishida E. A molecular mechanism that links Hippo signalling to the inhibition of Wnt/β-catenin signalling. EMBO J. 2012;31:1109–22. doi: 10.1038/emboj.2011.487. doi: 10.1038/emboj.2011.487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Heallen T, Zhang M, Wang J, Bonilla-Claudio M, Klysik E, Johnson RL, et al. Hippo pathway inhibits Wnt signaling to restrain cardiomyocyte proliferation and heart size. Science. 2011;332:458–61. doi: 10.1126/science.1199010. doi: 10.1126/science.1199010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Azzolin L, Panciera T, Soligo S, Enzo E, Bicciato S, Dupont S, et al. YAP/TAZ incorporation in the β-catenin destruction complex orchestrates the Wnt response. Cell. 2014;158:157–70. doi: 10.1016/j.cell.2014.06.013. doi: 10.1016/j.cell.2014.06.013. [DOI] [PubMed] [Google Scholar]
- 53.Azzolin L, Zanconato F, Bresolin S, Forcato M, Basso G, Bicciato S, et al. Role of TAZ as mediator of Wnt signaling. Cell. 2012;151:1443–56. doi: 10.1016/j.cell.2012.11.027. doi: 10.1016/j.cell.2012.11.027. [DOI] [PubMed] [Google Scholar]
- 54.Omwandho CO, Konrad L, Halis G, Oehmke F, Tinneberg HR. Role of TGF-betas in normal human endometrium and endometriosis. Hum Reprod. 2010;25:101–9. doi: 10.1093/humrep/dep382. doi: 10.1093/humrep/dep382. [DOI] [PubMed] [Google Scholar]
- 55.Li J, Du S, Sheng X, Liu J, Cen B, Huang F, et al. MicroRNA-29b inhibits endometrial fibrosis by regulating the sp1-TGF-β1/Smad-CTGF axis in a rat model. Reprod Sci. 2016;23:386–94. doi: 10.1177/1933719115602768. doi: 10.1177/1933719115602768. [DOI] [PubMed] [Google Scholar]
- 56.Leask A, Abraham DJ. TGF-beta signaling and the fibrotic response. FASEB J. 2004;18:816–27. doi: 10.1096/fj.03-1273rev. [DOI] [PubMed] [Google Scholar]
- 57.Branton MH, Kopp JB. TGF-beta and fibrosis. Microbes Infect. 1999;1:1349–65. doi: 10.1016/s1286-4579(99)00250-6. [DOI] [PubMed] [Google Scholar]
- 58.Mead AL, Wong TT, Cordeiro MF, Anderson IK, Khaw PT. Evaluation of anti-TGF-beta2 antibody as a new postoperative anti-scarring agent in glaucoma surgery. Invest Ophthalmol Vis Sci. 2003;44:3394–401. doi: 10.1167/iovs.02-0978. [DOI] [PubMed] [Google Scholar]
- 59.Khalil N, O’Connor R. Idiopathic pulmonary fibrosis: Current understanding of the pathogenesis and the status of treatment. CMAJ. 2004;171:153–60. doi: 10.1503/cmaj.1030055. doi: 10.1503/cmaj.1030055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Rybinski B, Franco-Barraza J, Cukierman E. The wound healing, chronic fibrosis, and cancer progression triad. Physiol Genomics. 2014;46:223–44. doi: 10.1152/physiolgenomics.00158.2013. doi: 10.1152/physiolgenomics.00158.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Piersma B, Bank RA, Boersema M. Signaling in fibrosis: TGF-β, WNT, and YAP/TAZ converge. Front Med (Lausanne) 2015;2:59. doi: 10.3389/fmed.2015.00059. doi: 10.3389/fmed.2015.000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Sun Y, Huang ZY, Wang ZH, Li CP, Meng XL, Zhang YJ, et al. TGF-β1 and TIMP-4 regulate atrial fibrosis in atrial fibrillation secondary to rheumatic heart disease. Mol Cell Biochem. 2015;406:131–8. doi: 10.1007/s11010-015-2431-1. doi: 10.1007/s11010-015-2431-1. [DOI] [PubMed] [Google Scholar]
- 63.Ayyaz A, Attisano L, Wrana JL. Recent advances in understanding contextual TGFβ signaling. F. 1000;es 2017;6:749. doi: 10.12688/f1000research.11295.1. doi: 10.12688/f1000research.11295.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Varelas X, Samavarchi-Tehrani P, Narimatsu M, Weiss A, Cockburn K, Larsen BG, et al. The crumbs complex couples cell density sensing to Hippo-dependent control of the TGF-β-SMAD pathway. Dev Cell. 2010;19:831–44. doi: 10.1016/j.devcel.2010.11.012. doi: 10.1016/j.devcel.2010.11.012. [DOI] [PubMed] [Google Scholar]
- 65.Miranda MZ, Bialik JF, Speight P, Dan Q, Yeung T, Szászi K, et al. TGF-β1 regulates the expression and transcriptional activity of TAZ protein via a smad3-independent, myocardin-related transcription factor-mediated mechanism. J Biol Chem. 2017;292:14902–20. doi: 10.1074/jbc.M117.780502. doi: 10.1074/jbc.M117.780502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Liu F, Lagares D, Choi KM, Stopfer L, Marinković A, Vrbanac V, et al. Mechanosignaling through YAP and TAZ drives fibroblast activation and fibrosis. Am J Physiol Lung Cell Mol Physiol. 2015;308:L344–57. doi: 10.1152/ajplung.00300.2014. doi: 10.1152/ajplung.00300.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]