

Abstract
This study provides a brain-based account of how object concepts at an intermediate (basic) level of specificity are represented, offering an enriched view of what it means for a concept to be a basic-level concept, a research topic pioneered by Rosch and others (Rosch et al., 1976). Applying machine learning techniques to fMRI data, it was possible to determine the semantic content encoded in the neural representations of object concepts at basic and subordinate levels of abstraction. The representation of basic-level concepts (e.g. bird) was spatially broad, encompassing sensorimotor brain areas that encode concrete object properties, and also language and heteromodal integrative areas that encode abstract semantic content. The representation of subordinate-level concepts (robin) was less widely distributed, concentrated in perceptual areas that underlie concrete content. Furthermore, basic-level concepts were representative of their subordinates in that they were neurally similar to their typical but not atypical subordinates (bird was neurally similar to robin but not woodpecker). The findings provide a brain-based account of the advantages that basic-level concepts enjoy in everyday life over subordinate-level concepts: the basic level is a broad topographical representation that encompasses both concrete and abstract semantic content, reflecting the multifaceted yet intuitive meaning of basic-level concepts.
Keywords: basic level, level of abstraction, neural representation, object concept, fMRI, MVPA
Introduction
If shown a picture of a robin, most people would call it a bird, fewer people would call it a robin, and even fewer would call it an animal. This scenario is representative of the tendency to name objects at an intermediate (basic) level of specificity, even if an object can be named at a more specific (subordinate) or general (superordinate) level (Rosch et al., 1976; Tversky & Hemenway, 1984). Although people can flexibly categorize the same object at different levels of abstraction, the basic level occupies a privileged position in the knowledge hierarchy in semantic cognition. Basic-level terms are generated faster than terms at other levels, and are used earlier by children (Jolicoeur et al., 1984; Murphy & Brownell, 1985; Anglin, 1977). Furthermore, basic-level terms are the nouns most frequently used in text (Wisniewski & Murphy, 1989). Although there remains some controversy as to the definition of a basic-level concept, the basic level of abstraction seems fundamental in cognition and deserves the label of basic. This finding has led to several attempts to explain the basic-level advantage, summarized below, as well as to this study’s attempt to characterize basic-level concepts in neural terms.
One prominent account attributes the basic-level advantage to the number of properties a concept has at a given level of abstraction (informativeness) and their overlap with the properties of other concepts at the same level (distinctiveness). Concepts at the basic level are both informative and distinctive (Rosch et al., 1976; Murphy & Brownell, 1985). For example, the basic-level concept bird is informative because it refers to a large number of properties that are shared by many bird species. Bird is also distinctive because birds differ substantially from other basic-level categories, including other animals such as fish. On the other hand, superordinate-level concepts are less informative than basic-level concepts, and subordinate-level concepts are less distinctive. Superordinate-level concepts are vague and uninformative because their member categories share few properties (e.g. animal could refer to birds or mammals, categories that have few similarities), but superordinate-level concepts are distinct from each other (animals and manmade objects differ substantially). Conversely, subordinate-level concepts are informative because they refer to well-defined objects, but they are largely indistinct from each other (e.g. robins and cardinals are much more similar than dissimilar).
Another possible account of the basic-level advantage concerns the types of semantic content that characterize concepts at each level of abstraction, as opposed to structural properties of the levels such as distinctiveness. Specifically, access to both concrete sensorimotor and abstract content in a concept may enable greater versatility in the use of a concept. Abstract knowledge, such as abstract properties that define a basic- or superordinate-level category (e.g. a tree is alive), can be useful for navigating semantic networks and realizing connections between distant concepts, which may aid in problem-solving or learning (Chi et al., 1981; Chi and Ohlsson, 2005). On the other hand, concrete knowledge about a concept, such as knowing how an object looks or feels, is helpful for communicating about the same concept by grounding the concept in people’s common sensorimotor experience (Goldstone et al., 2005). There is some evidence that basic-level concepts emphasize both concrete and abstract information, whereas the content of subordinate-level concepts tends to be concrete and superordinate-level concepts emphasize abstract content (Rosch et al., 1976; Tversky & Hemenway, 1984). The advantages of basic-level concepts might therefore derive from their balanced combination of multifaceted content.
The evidence that basic-level concepts emphasize both concrete and abstract semantic content is limited and draws from both behavioral and neuroimaging studies. Behavioral studies have shown that, when asked to list properties of object categories from different levels of abstraction, people tend to list nouns and adjectives for basic-level categories (e.g. has cloth, is round), nouns and even more adjectives for subordinate-level categories, and phrases that describe an object’s function or purpose for superordinate-level categories (Rosch et al., 1976; Tversky & Hemenway, 1984; Tversky, 1989). These studies show that the content of basic-level concepts lies between the mostly concrete content of subordinate-level concepts and the abstract properties of superordinate-level concepts.
Neuroimaging studies have found that naming pictures of objects at the superordinate versus the lower levels elicits greater activation in brain regions associated with abstract semantic content, while naming at the subordinate level preferentially activates sensorimotor areas (Kosslyn et al., 1995; Gauthier et al., 1997; Tanaka et al., 1999; Tyler et al., 2004; Rogers et al., 2006). For example, object naming at the superordinate level evoked greater activation in left inferior frontal gyrus, a region of the language system that has been shown to underlie the neural representation of abstract concepts (e.g. soul) more than concrete concepts (e.g. pliers) (Binder et al., 2005; Wang et al., 2010; Wang et al., 2013). On the other hand, object naming at the subordinate level elicited greater activation primarily in visual perceptual areas.
A concurrent emphasis on concrete and abstract content at the basic level suggests that a basic-level concept summarizes its subordinates rather than exhaustively specifying the properties of all its subordinates. According to theories that advocate this view, the abstract and general properties partially instantiate the subordinate-level concepts, for example knowing the common functions and general shape of cars partially instantiates many specific cars (Barsalou, 1992; Barsalou, 2003). The more concrete properties of a basic-level concept might be specific properties that are typical of its subordinates, for example thinking about car may evoke the property that it has four black tires (Murphy, 2004). By this account, basic-level concepts are partly concrete and partly abstract summaries that partially instantiate their typical subordinate-level concepts. Indeed, basic-level concepts might not retain their advantages over other concepts if they were not representative of their subordinates.
The current fMRI study aims to determine the types of semantic content that characterize object concepts at the basic and subordinate levels of abstraction, based on the brain areas differentially associated with the contemplation of concepts at the different levels. This aim corresponds to an approach that attempts to explain the advantages enjoyed by basic-level concepts in terms of the underlying semantic content.
In the experimental task used here, participants were asked to think about objects and their properties to evoke rich semantic content associated with the concepts. The concepts that were presented included basic-level concepts referring to living and manmade objects, and also typical and atypical subordinate-level concepts belonging to the basic-level categories (hereafter referred to as basic and subordinate concepts). The neural representations of basic and subordinate concepts were anatomically localized by determining where in the brain the two types of concept were neurally dissociable, using multivoxel pattern classification analysis (MVPA). MVPA can detect an activation pattern by virtue of assessing the conjoint activations of many voxels regardless of their spatial distribution in the brain (Mur et al., 2009). Univariate analysis that compares brain activation levels averaged across voxels was also used in conjunction with MVPA. These methods together provided a sensitive measure for uncovering the concepts’ neural representations, augmenting and updating previous research that compared univariate activation levels between different levels of abstraction using an object naming task (e.g. Kosslyn et al., 1995; Tyler et al., 2004; Rogers et al., 2006).
The neural representation of basic concepts was hypothesized to encompass a broad set of brain regions that encode concrete or abstract concept properties, namely sensorimotor cortex (concrete content) and language and heteromodal integration areas (abstract) (Hypothesis 1A). Complex, abstract properties are thought to result from an integration of more concrete properties that correspond to various sensorimotor modalities (Pexman et al., 2007; Bonner et al., 2013). An abstraction from sensorimotor modalities might also rely more on verbal processing, such as deriving meaning from word associates (Paivio, 1986).
On the other hand, compared to basic concepts, the neural representation of subordinate concepts was expected to reside to a greater extent in concrete sensorimotor areas (Hypothesis 1B). This representation was also expected to include anterior temporal cortex, a heteromodal area thought to bind together a concept’s properties to produce a unified concept (Patterson et al., 2007; Lambon Ralph, 2014). This area might be especially important for subordinate concepts due to their many specific properties that need to be bound to each other (Rogers et al., 2006). Hypotheses 1A–B are consistent with a body of research that has documented that concepts variously activate sensorimotor cortex, the language system, and heteromodal areas (Martin, 2007; Meteyard et al., 2010; Barsalou et al., 2008; Simmons et al., 2008).
A combination of concrete and abstract content at the basic level suggests that a basic concept is a representative summary of its subordinates that better describes its typical versus atypical subordinates. MVPA was used to test this corollary hypothesis that basic concepts are more neurally similar to their typical subordinates than to their atypical subordinates (e.g. the multivoxel activation pattern for bird was hypothesized to be more similar to that of robin than to woodpecker) (Hypothesis 2).
Materials and methods
Participants
Ten right-handed adults (seven males, three females; mean age of 25 years, ranging from 21 to 38) from Carnegie Mellon University and the Pittsburgh community participated and gave written informed consent approved by the Carnegie Mellon Institutional Review Board. Two additional participants’ data were discarded because of excessive head motion (greater than half the size of a voxel: 1.5mm total displacement in the x or y dimensions or 3mm in the z dimension). Two other participants’ data were discarded due to chance-level multivoxel pattern classification accuracy of the concepts (classification features were the 200 most “stable” voxels selected from anywhere in the brain excluding the occipital lobe; more detail concerning classification is provided below). This classification, which differed from the classification that tested the hypotheses, was used to check for systematicity in a participant’s activation patterns regardless of its correspondence to the hypotheses.
Experimental paradigm and task
The stimuli were 15 words which referred to a living or a manmade object at the basic or subordinate level, shown in Table I. There were five terms at the basic level and five typical and atypical subordinate terms. The terms at the basic level have been used in behavioral studies that examined the basic-level advantage (Rosch et al., 1976; Markman & Wisniewski, 1997). The typical and atypical subordinate terms were chosen using an independent group of seven participants who rated the typicality of various subordinates with respect to their basic categories, on a scale of 1–7. For example, out of four kinds of tree considered by the raters, oak received the highest mean typicality rating and palm the lowest. The typical (or atypical) subordinate terms selected as the stimuli were the most typical (or atypical) member of each of the five basic categories. The typicality ratings differed between the two sets of subordinate terms: the mean rating for typical subordinates was 6.34 (SD = 0.44 across terms), and for atypical subordinates it was 2.66 (SD = 0.39) (t(8) = 14.11, p < 0.001).
Table 1.
Basic and subordinate object concept stimuli.
| Basic concepts | Shoes | Car | Fish | Bird | Tree |
| Typical subordinate concepts | Sneakers | Taxi | Bass | Robin | Oak |
| Atypical subordinate concepts | Sandals | Limousine | Minnow | Woodpecker | Palm |
The stimulus words were presented six times, in six different random permutation orders. Each word was presented for 3s, followed by a 5s rest period, during which the participants were instructed to clear their minds and fixate on an “X” displayed in the center of the screen. A constant inter-trial interval was used rather than jittering, as only the mean activation across successive time points within a trial was submitted to the main analyses (described below); the precise temporal dynamics of the activation waveforms were not of interest in this study. Inter-trial intervals of 5s and similar duration have been found in previous studies of concept representation to provide sufficient time for the activation to return to baseline, as indicated by the high classification accuracies in these studies (e.g. a mean rank accuracy of 0.84, Chang et al., 2011). There were an additional five presentations of an “X” for 24s each, distributed evenly throughout the two scans, to provide a baseline measure for calculating percent signal change in the fMRI signal. The stimuli were presented using the CogLab presentation software.
When a word was presented, the participants’ task was to actively imagine and think about the properties of the object to which the word referred. The task is consistent with previous research that has treated a concept as a mental representation that picks out some of the properties of a real-world phenomenon (e.g. visual or functional properties of objects) (Cree & McRae, 2003; Wu & Barsalou, 2009). Several fMRI studies have used regression models to predict the activation patterns of object concepts, based on how different voxels are tuned to various properties of objects and on how important those properties are to defining a given object. For example, an object’s activation pattern can be predicted using its feature representation extracted from text corpora such as Wikipedia articles (Mitchell et al., 2008; Pereira et al., 2013). The participants in the current study generated properties for each object prior to the scanning session (for example, some properties generated for the basic concept tree were “is alive, is tall, has roots, has leaves where photosynthesis occurs”). Each participant was free to choose any properties for a given concept, and there was no attempt to impose consistency across participants in the choice of properties. The participants practiced the task prior to scanning to promote their contemplation of a consistent set of properties across the six presentations of a concept.
fMRI scanning parameters and data preprocessing
Functional blood oxygen level-dependent (BOLD) images were acquired on a 3T Siemens Verio Scanner and 32-channel phased-array head coil (Siemens Medical Solutions, Erlangen, Germany) at the Scientific Imaging and Brain Research Center of Carnegie Mellon University using a gradient echo EPI sequence with TR = 1000ms, TE = 25ms, and a 60° flip angle. Twenty 5mm-thick AC-PC-aligned slices were imaged with a gap of 1mm between slices, in an interleaved spatial order starting at the bottom. The acquisition matrix was 64 × 64 with 3.125 × 3.125 × 5mm in-plane resolution.
Data preprocessing was performed with the Statistical Parametric Mapping software (SPM8, Wellcome Department of Cognitive Neurology, London, UK). Images were corrected for slice acquisition timing, motion, and linear trend; temporally smoothed with a high-pass filter using a 190s cutoff; and normalized to the Montreal Neurological Institute (MNI) template without changing voxel size (3.125 × 3.125 × 6mm).
The percent signal change relative to the baseline condition was computed at each gray matter voxel for each stimulus presentation. The main input measure for the subsequent analyses consisted of the mean of the four brain images acquired within a 4s window, offset 5s from the stimulus onset (to account for the delay in hemodynamic response). The intensities of the voxels in this mean image for each stimulus presentation were then normalized (mean = 0, SD = 1).
Data analysis
Overview
A combination of MVPA and univariate analyses was used to test Hypothesis 1 regarding the topography of the neural representations of basic and subordinate concepts. The neural representations were anatomically localized by determining where in the brain the two types of concept were neurally dissociable. The identities of these brain regions indicated the types of semantic content encoded in the representations. MVPA was also used to test Hypothesis 2, namely that basic concepts are more neurally similar to their corresponding typical versus atypical subordinates.
Identifying an individual concept based on its multivoxel brain activation pattern
As a pre-condition for assessing the hypotheses, an initial classification analysis was performed to establish the systematicity of the multivoxel activation patterns underlying the concepts. A logistic regression classifier was implemented in MATLAB 7 (Mathworks, MA). Classification proceeded through three stages: algorithmic selection of a set of voxels (features) to be used for classification; training of a classifier on a subset of the data; and testing of the classifier on the remaining subset of the data. The training and testing used cross-validation procedures that iterated through all possible partitionings of the data into training and test sets, always keeping the training and test sets separate.
The voxels selected for use in the classification were the 200 most “stable” voxels drawn from any cortical region, excluding voxels in the occipital lobe that were correlated with the character length of the stimulus words. (Set sizes of 100–200 stable voxels resulted in similar outcomes.) The stability of a voxel was computed as the average pairwise correlation between its activation profiles (vector of its activation levels for the 15 stimulus words) across the repetitions in a training data subset (Just et al., 2010). For each partitioning into training and test data, the voxel selection criterion was applied to the training set and the classifier was trained to associate an activation pattern to each of the 15 words. Four (out of the six) repetitions of each word were used for training and the mean of the remaining two repetitions was used for testing, resulting in 15 total partitionings (folds) into training and test data. The activation values of the voxels were normalized (mean = 0, SD = 1) across all the words, separately for the training and test sets, to correct for possible drift in the signal across the six repetitions. Classification rank accuracy (referred to as accuracy) was the percentile rank of the correct word in the classifier’s ranked output (Mitchell et al., 2004).
Uncovering the neural representations of basic and subordinate concepts
A two-way classification between basic and subordinate concepts revealed voxels whose brain activation levels were greater for either basic or subordinate concepts. A logistic regression classifier was used with the same cross-validation procedures described above. For each cross-validation fold, the trained classifier was applied to each concept in the test set to classify it as either basic or subordinate. Because basic concepts were hypothesized to be more neurally similar to their typical versus atypical subordinates, two separate classifications were performed: basic versus atypical subordinate concepts, and basic versus typical subordinate concepts.
The features of each classifier were the 800 voxels with the highest (absolute value) logistic regression weights from an independent identical classification performed on the classifier’s training data per cross-validation fold. The voxels were selected from any cortical region, excluding voxels whose activation correlated with word length. The accuracy in classifying each concept as either basic or subordinate reached its maximum at large set sizes of voxels, such as 800 voxels. This large set size of voxels was used to explore which brain areas exhibited activation differences between basic and subordinate concepts.
The neural representation of basic concepts was identified as the voxels that were most informative for the classifier to identify basic concepts, namely those that had the highest positively-valued logistic regression weights. The representation of subordinate concepts was identified as the voxels most informative for identifying subordinate concepts, which had the lowest negatively-valued weights. (This approach has previously been used to identify populations of voxels whose activation is selective for different classes of stimuli, e.g. Shinkareva et al., 2008; Wang et al., 2013.) A posteriori, the voxels weighted toward basic concepts were revealed to have higher brain activation levels (percent signal change) for basic versus subordinate concepts, whereas the voxels weighted toward subordinate concepts had higher activation for subordinate concepts.
The voxels weighted toward basic concepts were predicted to be broadly distributed in the brain, located in brain areas that represent concrete or abstract semantic content. The voxels weighted toward subordinate concepts were predicted to be located predominantly in sensorimotor regions that encode concrete content.
Assessing the neural similarity between basic concepts and their subordinate concepts
Basic concepts were hypothesized to be more neurally similar to their corresponding typical versus atypical subordinate concepts (Hypothesis 2). To test this hypothesis, a classifier was trained on the basic concepts (e.g. bird) to identify the basic category membership of the typical and atypical subordinate concepts (e.g. whether robin and woodpecker are classified as a bird). The voxels used were the 200 voxels most stable over the basic concepts, disregarding occipital voxels whose activation correlated with word length. (The choice of 200 voxels was guided by previous studies in which 100–200 stable voxels were the smallest set sizes that could still attain among the highest accuracies in classification of individual concepts, e.g. Just et al., 2010; Mason & Just, 2015). Cross-classification was expected to result in higher accuracies between basic concepts and their typical versus atypical subordinate concepts.
To provide a further test of the hypothesis, additional classifiers were trained on the basic category membership of the subordinate concepts (separately for typical and atypical subordinates) to identify the individual basic concepts. The voxels used for these classifiers were stable for either typical or atypical subordinate concepts (see above for the definition of a “stable” voxel). Each cross-classification iterated over 15 cross-validation folds, where the training data corresponded to four (out of the six) repetitions of each word in the training set of concepts, and the test data corresponded to the mean of two repetitions of each word in the test set of concepts.
Results
Classification of individual concepts
As a pre-condition for the main findings that follow, the results showed that an individual concept was identifiable from its multivoxel brain activation pattern. The classification yielded a mean accuracy of 0.73 across the 10 participants and accuracies as high as 0.87 for two participants, as shown in Figure 1. (The p < 0.001 probability threshold for a rank accuracy being greater than chance level is 0.55, determined using random permutation testing of simulated data.) The classifier used 200 stable voxels which were distributed throughout the brain. These results established the systematicity of the activation patterns evoked by the concepts, enabling an analysis of how the basic and subordinate levels are differentially neurally represented.
Figure 1. An individual object concept could be identified based on its multivoxel brain activation pattern.

The classification accuracies averaged across the 15 concepts are shown for the 10 participants (ordered by accuracy) and the group mean. The dashed line indicates the p < 0.001 probability threshold for a rank accuracy being greater than chance level.
Neural representation of basic versus subordinate concepts
The neural representations of basic and subordinate concepts were anatomically localized by determining where in the brain the two types of concept were neurally dissociable, using MVPA. The neural representation of basic concepts was hypothesized to encompass a spatially broad set of brain regions that encodes concrete and abstract semantic content, that is, sensorimotor cortex (concrete) and language and heteromodal areas (abstract) (Hypothesis 1A). The representation of subordinate concepts was expected to be located primarily in sensorimotor areas (Hypothesis 1B). Because basic concepts were hypothesized to be more neurally similar to their typical versus atypical subordinates (Hypothesis 2), basic concepts were compared to atypical and typical subordinate concepts separately in two different analyses.
Basic concepts compared to atypical subordinate concepts
A binary logistic regression classifier distinguished between basic and atypical subordinate concepts based on their neural signatures with a mean accuracy of 0.70 across participants (p < 0.001) (as shown in Figure 2A). The features of the classifier were the 800 voxels with the highest (absolute value) logistic regression weights from an independent identical classification performed on the classifier’s training data per cross-validation fold. The voxels with the highest logistic regression weights were the most informative for the classifier to distinguish between basic and atypical subordinate concepts.
Figure 2. The neural representations of basic and subordinate object concepts were anatomically localized by determining where in the brain the two types of concept were neurally dissociable.
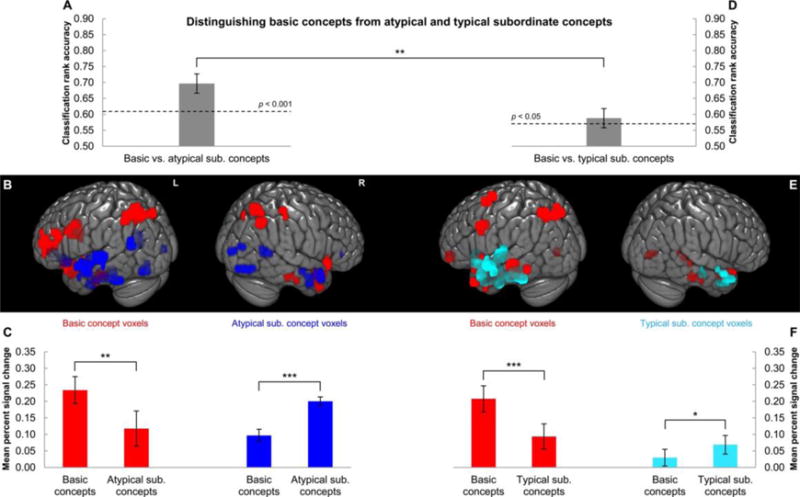
A, D: Mean accuracy (across participants) of distinguishing between (A) basic and atypical subordinate concepts and (D) basic and typical subordinate concepts. The dashed lines indicate the p < 0.05 and p < 0.001 probability thresholds for a rank accuracy being greater than chance level. B, E: The voxels that were most informative for the logistic regression classifiers to distinguish between (B) basic and atypical subordinate concepts and (E) basic and typical subordinate concepts (basic: red; atypical subordinate: blue; typical subordinate: cyan). These voxels had the highest or lowest 2.5% of the weights in the classification (basic: highest positively-valued weights; atypical/typical subordinate: lowest negatively-valued weights). The voxels were clustered at an extent threshold of 10 voxels. Rendering was performed on an MNI template brain using the 3D medical imaging software MRIcroGL (Rorden & Brett, 2000). C, F: The mean activation levels (percent signal change) across the clusters of voxels, elicited by (C) basic and atypical subordinate concepts and (F) basic and typical subordinate concepts. Note that the activation differences observed in these voxels are not independent from the finding that these voxels were weighted toward basic or atypical/typical subordinate concepts in the logistic regression classifications. All error bars are SEM across participants.
sub.: subordinate; L: left; R: right; MNI: Montreal Neurological Institute; *p < 0.05, **p < 0.01, ***p < 0.001 (paired-sample t-test)
The neural representation of basic concepts was identified as the voxels that were most informative for the classifier to identify basic concepts, and these voxels had positively-valued logistic regression weights. (This approach has previously been used to reveal populations of voxels whose activation is selective for different classes of stimuli, e.g. Shinkareva et al., 2008; Wang et al., 2013.) The voxels with the highest 2.5% of these weights were visualized across cross-validation folds and participants, generating a frequency map of the voxels with the highest weights for basic concepts. (Retaining different numbers of voxels in the frequency map showed a similar spatial distribution, as shown in Figure 3.) To reveal the voxels that were consistent across participants, the frequency map was thresholded by a simulated null-hypothesis distribution at p < 0.05 (FWE-corrected) after convolving each voxel with a 6mm Gaussian kernel (Kober et al., 2008).
Figure 3. The most informative voxels for classification between basic and atypical subordinate concepts were consistent across different feature selection thresholds.
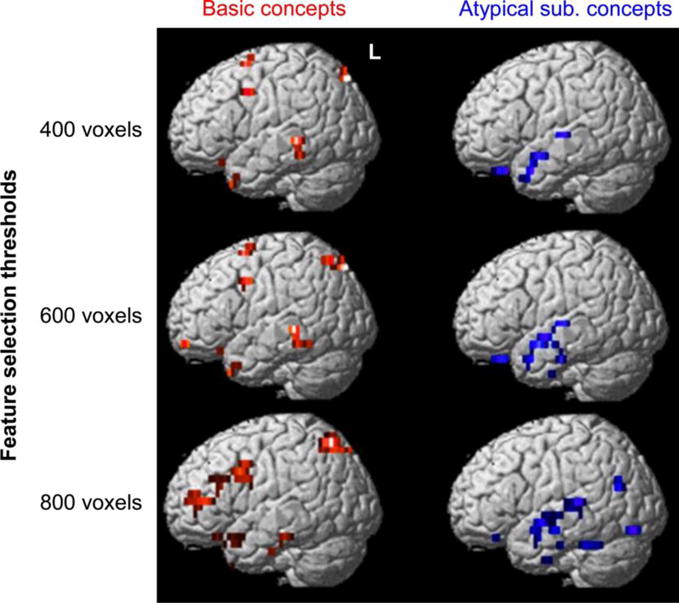
Logistic regression classifiers using different set sizes of voxels (400, 600, and 800) showed a similar distribution of the voxels most informative for decoding basic versus atypical subordinate concepts. The red color indicates the voxels weighted most toward basic concepts, with the highest 2.5% of the logistic regression weights. The blue color indicates the voxels weighted most toward atypical subordinate concepts, with the lowest 2.5% of the weights. The voxels are shown on the left hemisphere only, clustered at an extent threshold of 10 voxels. Rendering was performed on an MNI template brain using the Statistical Parametric Mapping software (SPM8, Wellcome Department of Cognitive Neurology, London, UK). Darker colors indicate greater anatomical depth.
sub.: subordinate; L: left; MNI: Montreal Neurological Institute
The voxels with the highest weights for basic concepts were distributed throughout frontal, parietal, and temporal lobes of the brain (red clusters in Figure 2B). There were clusters of voxels in language and heteromodal brain areas, including left inferior frontal gyrus (language), bilateral superior angular gyrus/intraparietal sulcus (heteromodal and visuospatial), and anterior temporal cortex (heteromodal). Additional clusters were located in sensorimotor areas, including bilateral anterior fusiform gyrus and right postcentral gyrus.
A posteriori, basic concepts were found to elicit greater activation levels than atypical subordinate concepts in the clusters of voxels with the highest weights for basic concepts. (Note that this finding of greater activation for basic concepts is not independent from the finding that these voxels were weighted toward basic concepts in the logistic regression classification.) In a cluster located in left inferior frontal gyrus, for example, basic concepts such as shoes and fish evoked greater activation (percent signal change averaged across voxels) than atypical subordinate concepts such as sandals and minnow, as shown in Figure 4 (t(9) = 6.36, p < 0.001). Higher activation elicited in this language region by basic concepts might indicate greater retrieval of verbal content that underlies abstract semantic content at the basic versus the subordinate level (Wang et al., 2010; Wang et al., 2013). The mean activation level across all the clusters weighted toward basic concepts was higher for basic versus atypical subordinate concepts (t(9) = 4.59, p < 0.01), as shown in Figure 2C.
Figure 4. Basic object concepts elicited greater activation (percent signal change) than atypical subordinate concepts in left inferior frontal gyrus.
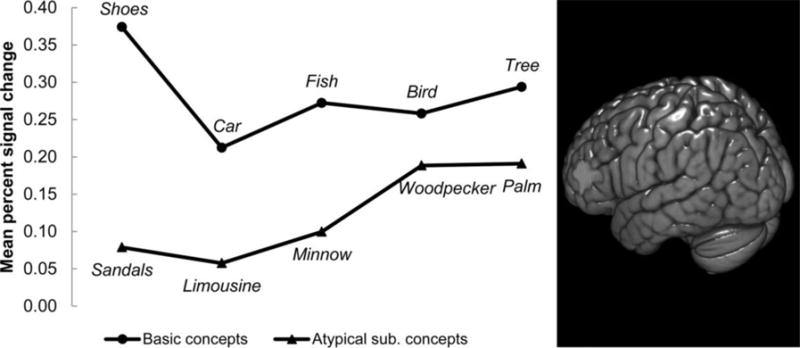
Right panel: The neural representation of basic concepts included a cluster of voxels in left inferior frontal gyrus, a language area. Left panel: Living and manmade object concepts at the basic level elicited greater activation levels in this cluster (averaged across voxels and participants) than their atypical subordinate counterparts. Rendering was performed on an MNI template brain using the 3D medical imaging software MRIcroGL (Rorden & Brett, 2000).
L: left; MNI: Montreal Neurological Institute
The spatial distribution of the representation of basic concepts was similar to the locations of voxels that exhibited greater activation for basic concepts as revealed by a group-level contrast using the general linear model (GLM). Figure 5 shows the clusters of voxels from the GLM contrast. Furthermore, the brain areas that showed higher activation for atypical subordinate concepts in the GLM contrast were similar to the voxels discovered by MVPA to underlie the representation of atypical subordinates.
Figure 5. Areas of heightened brain activation elicited by basic and atypical subordinate concepts.
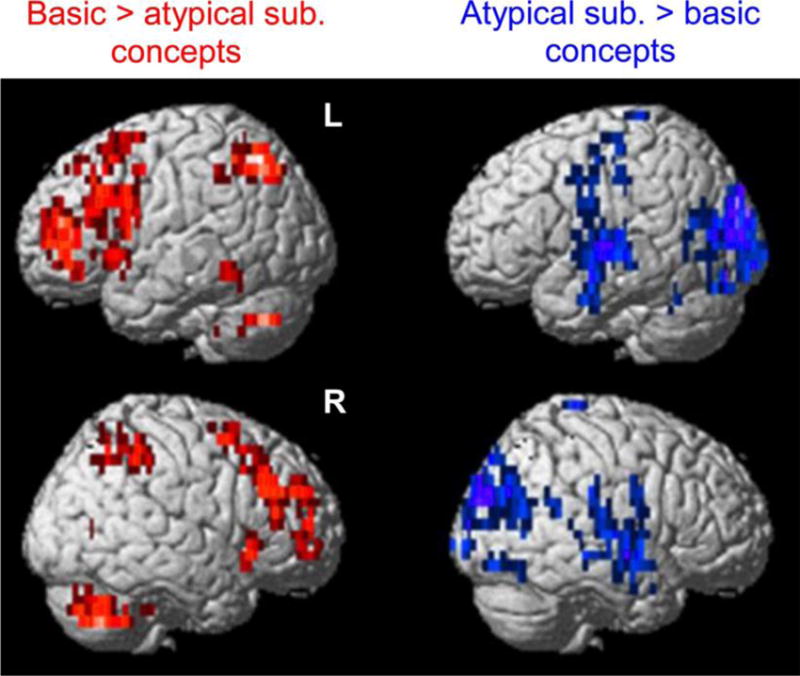
The red color indicates the voxels that exhibited greater activation for basic versus atypical subordinate concepts, as revealed by a whole-brain voxel-wise group-level GLM contrast (p < 0.05). The voxels were clustered at an extent threshold of 10 voxels. The blue color indicates the voxels that showed higher activation for atypical subordinate versus basic concepts. Rendering was performed on an MNI template brain using the Statistical Parametric Mapping software (SPM8, Wellcome Department of Cognitive Neurology, London, UK). Darker colors indicate greater anatomical depth.
sub.: subordinate; L: left; R: right; GLM: general linear model; MNI: Montreal Neurological Institute
The neural representation of atypical subordinate concepts was identified as the voxels most informative for the classifier to identify atypical subordinates, which had negatively-valued logistic regression weights. The voxels with the lowest 2.5% of these weights for atypical subordinates were less widely distributed in the brain than the voxels weighted toward basic concepts, and were concentrated in visual and auditory perceptual brain regions (blue clusters in Figure 2B). These areas were bilateral superior and inferior occipitotemporal cortex, bilateral anterior and posterior fusiform gyrus, and left superior temporal gyrus. There were also clusters of such voxels located bilaterally in anterior temporal cortex, a heteromodal area. The mean activation level across all the clusters was greater for atypical subordinate versus basic concepts (t(9) = 5.44, p < 0.001), as shown in Figure 2C.
Consistent with the hypothesis, the neural representation of basic concepts spanned frontal, parietal, and temporal lobes of the brain, encompassing sensorimotor brain areas that encode concrete content, and also language and heteromodal areas that encode abstract content. By contrast, the representation of atypical subordinate concepts was more focused in temporal and occipital perceptual regions that encode concrete content, although it also included a heteromodal element in anterior temporal cortex. These results were revealed using concepts referring to both living and manmade objects.
Basic concepts compared to typical subordinate concepts
A classifier distinguished between basic and typical subordinate concepts based on their neural signatures with a mean accuracy of 0.59 across participants (p < 0.05) (as shown in Figure 2D). This accuracy was lower than the accuracy of distinguishing between basic and atypical subordinate concepts (t(9) = 3.72, p < 0.01). This lower classification accuracy is consistent with the hypothesis that basic concepts are more neurally similar (less dissimilar) to their typical versus atypical subordinate concepts (Hypothesis 2). (Note that the difference between typical and atypical subordinate concepts in their neural separability from basic concepts is not attributable to a difference in the systematicity of their activation patterns. Classification of typical and atypical subordinate concepts yielded similar mean accuracies, which were 0.70 (SD = 0.15) and 0.66 (SD = 0.12), respectively; there was no statistically significant difference in their classification accuracies: t(9) = 1.02, p = 0.33.)
The anatomical distribution of the neural signatures of basic and typical subordinate concepts was similar to the distribution discovered in the classification between basic and atypical subordinate concepts. The voxels weighted toward basic concepts (i.e. voxels with the highest positively-valued weights in the classifier) were broadly distributed throughout the brain (red clusters in Figure 2E). Clusters of such voxels were located in language and heteromodal brain areas, including left inferior frontal gyrus and middle temporal gyrus (language), left superior angular gyrus/intraparietal sulcus (heteromodal and visuospatial), and bilateral anterior temporal cortex (heteromodal). There were also clusters in sensorimotor areas, including left premotor/supplementary motor area and bilateral anterior fusiform gyrus. The mean activation level across all the clusters was greater for basic versus typical subordinate concepts (t(9) = 6.02, p < 0.001) (Figure 2F).
The voxels weighted toward typical subordinate concepts (i.e. voxels with the lowest negatively-valued weights) were concentrated in visual and auditory perceptual areas, which included bilateral anterior fusiform gyrus and left superior temporal gyrus (cyan clusters in Figure 2E). Additional clusters were located bilaterally in anterior temporal cortex, a heteromodal area. The mean activation level across all the clusters was higher for typical subordinate versus basic concepts (t(9) = 2.31, p < 0.05) (Figure 2F).
In summary, the neural representation of basic concepts was topographically broader than the representation of typical subordinate concepts. The extra breadth included language and heteromodal brain areas that encode abstract semantic content. Like atypical subordinate concepts, typical subordinate concepts were neurally represented in a narrower set of brain areas that encode primarily concrete perceptual properties of the concepts. Despite this similarity in the spatial distribution of typical and atypical representations, typical subordinate concepts were more neurally similar to basic concepts than atypical subordinates were. This result indicates some commonality in the semantic content encoded in the representations of basic and typical subordinate concepts.
Neural similarity between basic and typical subordinate concepts
In addition to comparing the topography of the neural representations of basic and subordinate concepts, one can also consider pairwise comparisons of the concepts. Basic concepts were hypothesized to be more neurally similar to their corresponding typical versus atypical subordinate concepts (e.g. bird was hypothesized to be more neurally similar to robin than to woodpecker) (Hypothesis 2). The results of the classification analyses between basic and subordinate concepts reported above revealed that basic concepts were on average more neurally similar to typical than atypical subordinate concepts. Additional classification was performed to determine whether a given basic concept was more neurally similar to its typical versus atypical subordinate concept.
A classifier trained on the basic concepts (e.g. bird) could identify the basic category membership of the typical subordinate concepts (e.g. robin is a bird), yielding a mean classification accuracy of 0.60 across participants (p < 0.05) (as shown in Figure 6A). The classifier used the 200 most stable voxels in the brain, which were distributed throughout the brain areas discovered to underlie the representations of basic and subordinate concepts. By contrast, a classifier trained on the basic concepts could not identify the basic category membership of the atypical subordinate concepts (e.g. woodpecker could not be identified as a bird), yielding a mean accuracy of 0.52 which was below chance level. (The p < 0.05 probability threshold for a rank accuracy being greater than chance level is 0.57.) This accuracy was lower than the accuracy of the analogous cross-classification using typical subordinate concepts (t(9) = 2.40, p < 0.05). These results are consistent with the hypothesis that basic concepts are more neurally similar to their corresponding typical versus atypical subordinate concepts.
Figure 6. Basic object concepts were neurally similar to their corresponding typical, but not atypical, subordinate concepts.
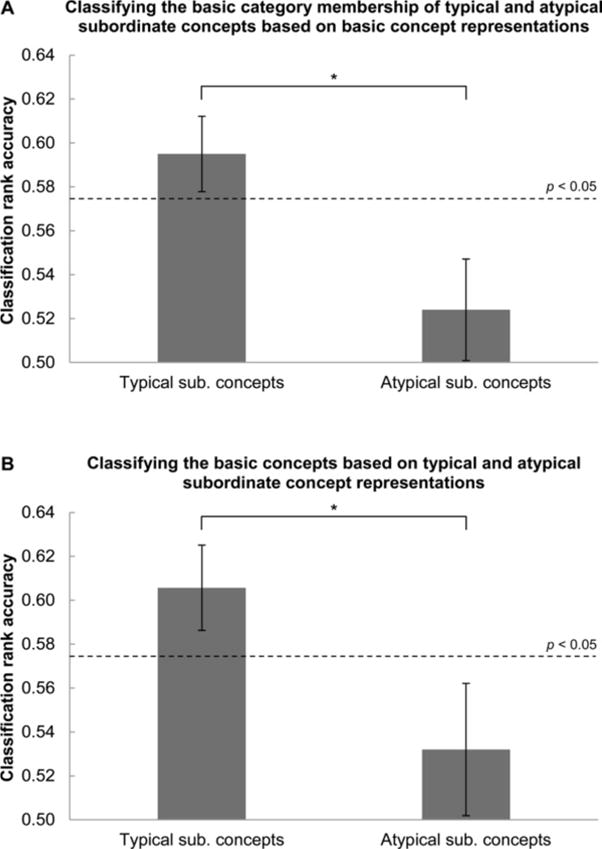
A: The basic category membership of the typical, but not atypical, subordinate concepts could be identified using a classifier trained on the neural representations of the basic concepts (e.g. robin but not woodpecker was identified as a bird). B: The individual basic concepts could be identified using a classifier trained on the neural representations of the typical, but not atypical, subordinate concepts. The classification accuracies shown above are averaged across participants, and the dashed lines indicate the p < 0.05 probability threshold for a rank accuracy being greater than chance level. Error bars are SEM across participants.
sub.: subordinate; *p < 0.05 (paired-sample t-test)
Additional cross-classification provided further support for the hypothesis. A classifier trained on the basic category membership of the typical subordinate concepts (e.g. robin is a bird) could identify the individual basic concepts with a mean classification accuracy of 0.61, using 200 voxels in the classification (p < 0.05) (as shown in Figure 6B). By contrast, a classifier trained on the basic category membership of the atypical subordinate concepts (e.g. woodpecker is a bird) could not identify the basic concepts, yielding a mean accuracy of 0.53, which was lower than the accuracy of the analogous cross-classification involving typical subordinate concepts (t(9) = 2.63, p < 0.05).
Discussion
This study provides a brain-based account of how object concepts at different levels of abstraction are represented, offering an enriched view of what it means for a concept to be a basic-level concept, a research topic pioneered by Rosch and others (e.g. Rosch et al., 1976). Using sensitive machine learning techniques, it was possible to determine the semantic content encoded in the neural representations of living and manmade object concepts that are at different levels of abstraction. The findings provide a brain-based account of the advantages that basic concepts enjoy in everyday life over subordinate concepts: the basic level is a broad topographical representation that encompasses both concrete and abstract semantic content, reflecting the multifaceted yet intuitive meaning of basic concepts. Another novel result here is that basic concepts are more neurally similar to their typical than atypical subordinate concepts, which may further explain the greater utility of basic concepts in everyday life: basic concepts are representative of their subordinates, and thus are applicable in a wide variety of situations.
The semantic content encoded in the neural representation of basic concepts
The brain areas discovered to underlie the neural representation of basic concepts included sensorimotor cortex (concrete content), frontal and temporal language areas (abstract), and temporal and parietal heteromodal areas (abstract), consistent with Hypothesis 1A. Furthermore, as predicted basic concepts were representative of their subordinates in that they were neurally similar to their typical but not atypical subordinate concepts (Hypothesis 2).
The language areas involved in the representation of basic concepts included left inferior frontal gyrus and middle temporal gyrus, which have been widely implicated in language (Price, 2010). These areas have been shown to underlie the representation of abstract concepts (e.g. soul) more than concrete concepts (e.g. pliers) (Binder et al., 2005; Wang et al., 2010; Wang et al., 2013). Left inferior frontal gyrus has also been shown to activate more when naming objects at the superordinate versus the basic level, perhaps indicating greater retrieval of verbal content at successively higher levels of abstraction (Kosslyn et al., 1995; Tyler et al., 2004). Verbal content such as word associates might underlie concept properties that abstract away from sensorimotor modalities (Paivio, 1986). Abstract properties that define a basic or superordinate category may be difficult to instantiate in sensorimotor detail (e.g. a tree performs photosynthesis, and is alive) (Tversky, 1989). Left inferior frontal gyrus might be a retrieval site for verbal content, and might also provide executive control needed for the selection of concrete content that instantiates an abstract concept (Hoffman et al., 2010).
The representation of abstract content might also be subserved by anterior temporal cortex and inferior parietal cortex, which are thought to be “convergence zones” that bind and integrate the various properties of a concept corresponding to different sensorimotor modalities (Damasio, 1989; Binder & Desai, 2011; Pulvermüller, 2013). Both of these brain areas are situated neuroanatomically at a confluence of modal processing streams. Inferior parietal cortex, including angular gyrus, has been suggested to organize information from different modalities to construct higher-level concepts (Bonner et al., 2013). In particular, inferior parietal cortex might be especially important to the representation of events, due to its anatomical inclusion of brain networks that underlie action processing (Binder & Desai, 2011). The mental representation of an object’s function, an abstract property, may take the form of an event that draws on heteromodal content including action information about the object’s use.
Inferior parietal cortex, especially intraparietal sulcus, might also encode a coarse visuospatial description of a basic category’s subordinates, such as the objects’ shared shape and visually-guided use (Palmeri & Gauthier, 2004). These properties are likely to be represented at the basic level, as they have been shown in early landmark studies to be similar across subordinates belonging to the same basic category (e.g. Rosch et al., 1976).
The concrete sensorimotor areas identified in the representation of basic concepts included fusiform gyrus, premotor/supplementary motor area, and postcentral gyrus. Fusiform gyrus might represent specific visual properties of typical subordinates of the basic concepts. Furthermore, motor information (e.g. how to drive a car) might reside in the motor and somatosensory areas (Lewis, 2006).
The semantic content encoded in the neural representation of subordinate concepts
Compared to basic concepts, subordinate concepts were hypothesized to have a greater emphasis on concrete sensorimotor content (Hypothesis 1B). Consistent with the hypothesis, the neural representation of subordinate concepts (typical and atypical) was focused in perceptual areas, although it also contained a heteromodal element in anterior temporal cortex.
Anterior temporal cortex, and particularly anterior fusiform gyrus, is thought to play a critical role in binding together a concept’s various properties to produce a unified concept (Patterson et al., 2007; Lambon Ralph, 2014; Coutanche & Thompson-Schill, 2015). Although both subordinate and basic concepts have previously been found to activate these brain areas, subordinate concepts elicited greater activation than basic concepts (Rogers et al., 2006), which is consistent with the finding that subordinate concepts have a larger number of descriptive properties that need binding (Rosch et al., 1976; Tversky & Hemenway, 1984). The results reported here are consistent with this research, showing that anterior temporal cortex underlies the representation of both subordinate and basic concepts, with a somewhat greater focus on subordinate concepts.
The perceptual brain areas that were identified in the representation of subordinate concepts included occipitotemporal cortex, anterior and posterior fusiform gyrus, and superior temporal gyrus. Specific visual properties of the subordinate object concepts might be represented in occipitotemporal cortex. Multivoxel activation patterns in this area have been shown to contain information of specific objects that are subordinates of the same category (e.g. different chairs) (Eger et al., 2008). Visual properties might also be represented in fusiform gyrus, such as color information in posterior fusiform gyrus (Bramão et al., 2010). Previous research has found greater activity in these areas when naming depicted objects at the subordinate versus the basic level, possibly reflecting the need for greater processing of visual detail to provide specific object names (Gauthier et al., 1997; Gauthier et al., 2000). A “holistic” perceptual system implemented in fusiform gyrus could also be engaged when thinking about a complex interrelationship among the visual properties of a familiar subordinate concept (Bukach et al., 2006).
Acoustic information associated with the object concepts (such as sounds made by various birds or cars) could be represented in left superior temporal gyrus, an auditory perceptual area. Previous research has shown that reading words referring to objects with salient acoustic properties activates this region more than other words (Kiefer et al., 2008).
General discussion
The greater utility of basic concepts over other concepts in everyday life might be partly explained by the current brain-based findings. The lesser utility of subordinate concepts might be reflected in the comparatively small number of brain areas found here to constitute the focus of their neural representation, which had an unbalanced emphasis on perceptual content. These results suggest that the neural signature of concepts at the superordinate level should also be focused in a small number of brain areas, but these areas should emphasize abstract semantic content, given that superordinate categories tend to prompt abstract properties such as an object’s function in feature listing tasks (Rosch et al., 1976; Tversky & Hemenway, 1984). These areas ought to be language and heteromodal regions, as suggested by previous research on superordinate concepts (Kosslyn et al., 1995; Tyler et al., 2004). The advantage of basic concepts may therefore lie in their balanced activation of sensorimotor, verbal, and heteromodal integrative centers of the semantic system.
The explanation of the basic-level advantage proposed here may account for changes in the basic level observed in experts, considered a major obstacle for strictly Roschean accounts (Medin et al., 2000). Experts in living categories, including biologists or traditional societies such as Mayan Amerindians, show a basic-level advantage at the specific level of abstraction (e.g. trout is a “basic” concept rather than fish) (Coley et al., 1997). These findings are inconsistent with the Roschean account that the basic level corresponds to the most abstract level at which category members share many properties, namely the intermediate level of abstraction (Rosch et al., 1976). The current study suggests instead that experts come to attach more conceptual knowledge to the detailed perceptual properties of specific-level concepts, thus lowering the basic level to the level of specific concepts. Indeed, experts in physics and computer science have been shown to group textbook exercises according to shared abstract principles (e.g. “conservation of energy”), while non-experts sort items based on surface features of the exercises (e.g. “presence of an incline”) (Chi et al., 1981; Weiser & Shertz, 1983). Top-down effects of expert knowledge have also been demonstrated in a simple visual categorization task (Harel et al., 2011). The brain-based account proposed here thus offers an enriched view of what it means for a concept to be a basic concept.
Atypical subordinate concepts were found here to differ neurally from basic concepts, but behavioral research has found atypical subordinates to exhibit some basic-level advantages, such as faster naming than typical subordinate and even basic concepts (Murphy & Brownell, 1985). Basic concepts are thought to have an advantage because they are both informative and distinctive, whereas subordinate concepts are informative but less distinct. Typical subordinates of the same basic category are largely indistinct from each other because they differ in only a small number of ways (e.g. robins and cardinals). Atypical subordinates, on the other hand, are the most distinctive members of a basic category (e.g. woodpeckers rather than robins), making them both informative and distinctive and thus making them more similar to basic concepts. However, the results reported here show that atypical subordinates differ from basic concepts in the same way that typical subordinates differ: the representation of subordinate concepts has an unbalanced focus on concrete semantic content. Future research is needed to determine whether different basic-level advantages have different underlying causes.
This first account of the advantages of basic object concepts in terms of their neural representation opens new perspectives. The representation of the basic level is a balanced activation of sensorimotor, verbal, and heteromodal integrative centers of the semantic system, perhaps revealing an optimal neural configuration for the processing or use of such concepts.
Acknowledgments
This research was supported by the National Institute of Mental Health grant MH029617. We thank Charles Kemp for many helpful suggestions regarding the data analysis and writing of the paper.
Contributor Information
Andrew James Bauer, Sidney Smith Hall, Dept. of Psychology, University of Toronto, 100 St. George Street, Toronto, ON M5S 3G3, Canada.
Marcel Adam Just, Center for Cognitive Brain Imaging, Baker Hall, Dept. of Psychology, Carnegie Mellon University, 5000 Forbes Avenue, Pittsburgh, PA 15213, USA, Phone: +1.412.268.2791.
References
- Anglin JM. Word, Object, and Conceptual Development. New York: W. W. Norton; 1977. [Google Scholar]
- Barsalou LW. Frames, concepts, and conceptual fields. In: Kittay E, Lehrer A, editors. Frames, fields, and contrasts: New essays in semantic and lexical organization. Hillsdale: Lawrence Erlbaum Associates; 1992. pp. 21–74. [Google Scholar]
- Barsalou LW. Abstraction in perceptual symbol systems. Philosophical Transactions of the Royal Society, Series B. 2003;358(1435):1177–1187. doi: 10.1098/rstb.2003.1319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barsalou LW, Santos A, Simmons WK, Wilson CD. Language and simulation in conceptual processing. In: De Vega M, Glenberg AM, AC Graesser A, editors. Symbols, embodiment, and meaning. Oxford, England: Oxford University Press; 2008. pp. 245–283. [Google Scholar]
- Binder JR, Westbury CF, McKiernan KA, Possing ET, Medler DA. Distinct brain systems for processing concrete and abstract concepts. Journal of Cognitive Neuroscience. 2005;17(6):905–917. doi: 10.1162/0898929054021102. [DOI] [PubMed] [Google Scholar]
- Binder JR, Desai RH. The neurobiology of semantic memory. Trends in Cognitive Sciences. 2011;15(11):527–536. doi: 10.1016/j.tics.2011.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonner MF, Peelle JE, Cook PA, Grossman M. Heteromodal conceptual processing in the angular gyrus. NeuroImage. 2013;71:175–186. doi: 10.1016/j.neuroimage.2013.01.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bramão I, Faísca L, Forkstam C, Reis A, Petersson KM. Cortical brain regions associated with color processing: an fMRI study. Open Neuroimaging Journal. 2010;4:164–173. doi: 10.2174/1874440001004010164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bukach CM, Gauthier I, Tarr MJ. Beyond faces and modularity: the power of an expertise framework. Trends in Cognitive Sciences. 2006;10(4):159–166. doi: 10.1016/j.tics.2006.02.004. [DOI] [PubMed] [Google Scholar]
- Chang KK, Mitchell T, Just MA. Quantitative modeling of the neural representation of objects: how semantic feature norms can account for fMRI activation. NeuroImage. 2011;56(2):716–727. doi: 10.1016/j.neuroimage.2010.04.271. [DOI] [PubMed] [Google Scholar]
- Chi MTH, Feltovich P, Glaser R. Categorization and representation of physics problems by experts and novices. Cognitive Science. 1981;5:121–152. [Google Scholar]
- Chi MTH, Ohlsson S. Complex declarative learning. In: Holyoak KJ, Morrison RG, editors. The Cambridge handbook of thinking and reasoning. New York: Cambridge University Press; 2005. pp. 371–399. [Google Scholar]
- Coley JD, Medin DL, Atran S. Does rank have its privilege? Inductive inferences within folkbiological taxonomies. Cognition. 1997;64:73–112. doi: 10.1016/s0010-0277(97)00017-6. http://doi.org/10.1016/S0010-0277(97)00017-6. [DOI] [PubMed] [Google Scholar]
- Coutanche MN, Thompson-Schill SL. Creating Concepts from Converging Features in Human Cortex. Cerebral Cortex. 2015;25(9):2584–2593. doi: 10.1093/cercor/bhu057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cree GS, McRae K. Analyzing the factors underlying the structure and computation of the meaning of chipmunk, cherry, chisel, cheese, and cello (and many other such concrete nouns) Journal of Experimental Psychology: General. 2003;132(2):163–201. doi: 10.1037/0096-3445.132.2.163. [DOI] [PubMed] [Google Scholar]
- Damasio AR. Time-locked multiregional retroactivation: a systems-level proposal for the neural substrates of recall and recognition. Cognition. 1989;33(1–2):25–62. doi: 10.1016/0010-0277(89)90005-x. [DOI] [PubMed] [Google Scholar]
- Eger E, Ashburner J, Haynes JD, Dolan RJ, Rees G. fMRI activity patterns in human LOC carry information about object exemplars within category. Journal of Cognitive Neuroscience. 2008;20(2):356–370. doi: 10.1162/jocn.2008.20019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gauthier I, Anderson AW, Tarr MJ, Skudlarski P, Gore JC. Levels of categorization in visual recognition studied using functional magnetic resonance imaging. Current Biology. 1997;7(9):645–651. doi: 10.1016/s0960-9822(06)00291-0. [DOI] [PubMed] [Google Scholar]
- Gauthier I, Tarr MJ, Moylan J, Anderson AW, Skudlarski P, Gore JC. Does visual subordinate categorisation engage the functionally defined fusiform face area? Cognitive Neuropsychology. 2000;17(1/2/3):143–163. doi: 10.1080/026432900380544. [DOI] [PubMed] [Google Scholar]
- Goldstone RL, Feng Y, Rogosky B. Connecting concepts to the world and each other. In: Pecher D, Zwaan R, editors. Grounding cognition: The role of perception and action in memory, language, and thinking. Cambridge, England: Cambridge University Press; 2005. pp. 292–314. [Google Scholar]
- Harel A, Ullman S, Harari D, Bentin S. Basic-level categorization of intermediate complexity fragments reveals top-down effects of expertise in visual perception. Journal of Vision. 2011;11(8):1–13. doi: 10.1167/11.8.18. http://doi.org/10.1167/11.8.18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffman P, Jefferies E, Lambon Ralph MA. Ventrolateral prefrontal cortex plays an executive regulation role in comprehension of abstract words: convergent neuropsychological and repetitive TMS evidence. The Journal of Neuroscience. 2010;30(46):15450–15456. doi: 10.1523/JNEUROSCI.3783-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jolicoeur P, Gluck M, Kosslyn SM. Pictures and names: Making the connection. Cognitive Psychology. 1984;19:31–53. doi: 10.1016/0010-0285(84)90009-4. [DOI] [PubMed] [Google Scholar]
- Just MA, Cherkassky VL, Aryal S, Mitchell TM. A neurosemantic theory of concrete noun representation based on the underlying brain codes. PLoS ONE. 2010;5(1):e8622. doi: 10.1371/journal.pone.0008622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiefer M, Sim EJ, Herrnberger B, Grothe J, Hoenig K. The sound of concepts: four markers for a link between auditory and conceptual brain systems. The Journal of Neuroscience. 2008;28(47):12224–30. doi: 10.1523/JNEUROSCI.3579-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kober H, Barrett LF, Joseph J, Bliss-Moreau E, Lindquist K, Wager TD. Functional grouping and cortical-subcortical interactions in emotion: A meta-analysis of neuroimaging studies. NeuroImage. 2008;42(2):998–1031. doi: 10.1016/j.neuroimage.2008.03.059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kosslyn SM, Alpert NM, Thompson WL. Identifying objects at different levels of hierarchy: A positron emission tomography study. Human Brain Mapping. 1995;3(2):107–132. doi: 10.1002/hbm.460030207. [DOI] [Google Scholar]
- Lewis JW. Cortical networks related to human use of tools. The Neuroscientist. 2006;12(3):211–231. doi: 10.1177/1073858406288327. [DOI] [PubMed] [Google Scholar]
- Markman AB, Wisniewski EJ. Similar and different: The differentiation of basic-level categories. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1997;23(1):54–70. doi: 10.1037/0278-7393.23.1.54. [DOI] [Google Scholar]
- Martin A. The representation of object concepts in the brain. Annual Review of Psychology. 2007;58:25–45. doi: 10.1146/annurev.psych.57.102904.190143. [DOI] [PubMed] [Google Scholar]
- Mason RA, Just MA. Physics instruction induces changes in neural knowledge representation during successive stages of learning. NeuroImage. 2015;111:36–48. doi: 10.1016/j.neuroimage.2014.12.086. [DOI] [PubMed] [Google Scholar]
- Medin DL, Lynch EB, Solomon KO. Are There Kinds of Concepts? Annual Review of Psychology. 2000;51:121–147. doi: 10.1146/annurev.psych.51.1.121. [DOI] [PubMed] [Google Scholar]
- Meteyard L, Cuadrado SR, Bahrami B, Vigliocco G. Coming of age: A review of embodiment and the neuroscience of semantics. Cortex. 2010;48(7):788–804. doi: 10.1016/j.cortex.2010.11.002. [DOI] [PubMed] [Google Scholar]
- Mitchell T, Hutchinson R, Niculescu RS, Pereira F, Wang X, Just MA, Newman SD. Learning to decode cognitive states from brain images. Machine Learning. 2004;57:145–175. [Google Scholar]
- Mitchell TM, Shinkareva SV, Carlson A, Chang K-M, Malave VL, Mason RA, Just MA. Predicting human brain activity associated with the meanings of nouns. Science. 2008;320(5880):1191–1195. doi: 10.1126/science.1152876. [DOI] [PubMed] [Google Scholar]
- Mur M, Bandettini PA, Kriegeskorte N. Revealing representational content with pattern-information fMRI—an introductory guide. Social Cognitive and Affective Neuroscience. 2009;4(1):101–109. doi: 10.1093/scan/nsn044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murphy GL, Brownell HH. Category differentiation in object recognition: Typicality constraints on the basic category advantage. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1985;11(1):70–84. doi: 10.1037//0278-7393.11.1.70. [DOI] [PubMed] [Google Scholar]
- Murphy GL. The Big Book of Concept. Cambridge: MIT Press; 2004. [Google Scholar]
- Palmeri TJ, Gauthier I. Visual object understanding. Nature Reviews Neuroscience. 2004;5(4):291–303. doi: 10.1038/nrn1364. [DOI] [PubMed] [Google Scholar]
- Paivio A. Mental representations: A dual-coding approach. New York: Oxford University Press; 1986. [Google Scholar]
- Patterson K, Nestor PJ, Rogers TT. Where do you know what you know? The representation of semantic knowledge in the human brain. Nature Reviews Neuroscience. 2007;8(12):976–987. doi: 10.1038/nrn2277. [DOI] [PubMed] [Google Scholar]
- Pereira F, Botvinick M, Detre G. Using Wikipedia to learn semantic feature representations of concrete concepts in neuroimaging experiments. Artificial Intelligence. 2013;194:240–252. doi: 10.1016/j.artint.2012.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pexman PM, Hargreaves IS, Edwards JD, Henry LC, Goodyear BG. Neural correlates of concreteness in semantic categorization. Journal of Cognitive Neuroscience. 2007;19(8):1407–1419. doi: 10.1162/jocn.2007.19.8.1407. [DOI] [PubMed] [Google Scholar]
- Price CJ. The anatomy of language: a review of 100 fMRI studies published in 2009. Annals of the New York Academy of Sciences. 2010;1191:62–88. doi: 10.1111/j.1749-6632.2010.05444.x. [DOI] [PubMed] [Google Scholar]
- Pulvermüller F. How neurons make meaning: Brain mechanisms for embodied and abstract-symbolic semantics. Trends in Cognitive Sciences. 2013;17(9):458–470. doi: 10.1016/j.tics.2013.06.004. [DOI] [PubMed] [Google Scholar]
- Rogers TT, Hocking J, Noppeney U, Mechelli A, Gorno-Tempini ML, Patterson K, Price CJ. Anterior temporal cortex and semantic memory: reconciling findings from neuropsychology and functional imaging. Cognitive, Affective & Behavioral Neuroscience. 2006;6(3):201–213. doi: 10.3758/cabn.6.3.201. [DOI] [PubMed] [Google Scholar]
- Rorden C, Brett M. Stereotaxic display of brain lesions. Behavioural Neurology. 2000;12(4):191–200. doi: 10.1155/2000/421719. [DOI] [PubMed] [Google Scholar]
- Rosch E, Mervis CB, Gray W, Johnson D, Boyes-Braem P. Basic objects in natural categories. Cognitive Psychology. 1976;8:382–439. [Google Scholar]
- Shinkareva SV, Mason RA, Malave VL, Wang W, Mitchell TM, Just MA. Using fMRI brain activation to identify cognitive states associated with perception of tools and dwellings. PLoS ONE. 2008;3(1):e1394. doi: 10.1371/journal.pone.0001394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simmons WK, Hamann SB, Harenski CL, Hu XP, Barsalou LW. fMRI evidence for word association and situated simulation in conceptual processing. Journal of Physiology, Paris. 2008;102(1–3):106–119. doi: 10.1016/j.jphysparis.2008.03.014. [DOI] [PubMed] [Google Scholar]
- Tanaka J, Luu P, Weisbrod M, Kiefer M. Tracking the time course of object categorization using event-related potentials. NeuroReport. 1999;10(4):829–835. doi: 10.1097/00001756-199903170-00030. [DOI] [PubMed] [Google Scholar]
- Tversky B, Hemenway K. Objects, parts, and categories. Journal of Experimental Psychology: General. 1984;113:169–193. [PubMed] [Google Scholar]
- Tversky B. Parts, partonomies, and taxonomies. Developmental Psychology. 1989;25(6):983–995. doi: 10.1037/0012-1649.25.6.983. [DOI] [Google Scholar]
- Wang J, Conder JA, Blitzer DN, Shinkareva SV. Neural representation of abstract and concrete concepts: A meta-analysis of neuroimaging studies. Human Brain Mapping. 2010;31(10):1459–1468. doi: 10.1002/hbm.20950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang J, Baucom LB, Shinkareva SV. Decoding abstract and concrete concept representations based on single-trial fMRI data. Human Brain Mapping. 2013;34(5):1133–1147. doi: 10.1002/hbm.21498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weiser M, Shertz J. Programming problem representation in novice and expert programmers. International Journal of Man-Machine Studies. 1983;19:391–398. [Google Scholar]
- Wisniewski EJ, Murphy GL. Superordinate and basic category names in discourse: A textual analysis. Discourse Processes. 1989;12:245–261. [Google Scholar]
- Wu L, Barsalou LW. Perceptual simulation in conceptual combination: evidence from property generation. Acta Psychologica. 2009;132(2):173–189. doi: 10.1016/j.actpsy.2009.02.002. [DOI] [PubMed] [Google Scholar]