

Abstract
Convergence is the tendency of independent species to evolve similarly when subjected to the same environmental conditions. The primitive blueprint for the circulatory system emerged around 700–600 Mya and exhibits diverse physiological adaptations across the radiations of vertebrates (Subphylum Vertebrata, Phylum Chordata). It has evolved from the early chordate circulatory system with a single layered tube in the tunicate (Subphylum Urchordata) or an amphioxus (Subphylum Cephalochordata), to a vertebrate circulatory system with a two‐chambered heart made up of one atrium and one ventricle in gnathostome fish (Infraphylum Gnathostomata), to a system with a three‐chambered heart made up of two atria which maybe partially divided or completely separated in amphibian tetrapods (Class Amphibia). Subsequent tetrapods, including crocodiles and alligators (Order Crocodylia, Subclass Crocodylomorpha, Class Reptilia), birds (Subclass Aves, Class Reptilia) and mammals (Class Mammalia) evolved a four‐chambered heart. The structure and function of the circulatory system of each individual holds a vital role which benefits each species specifically. The special characteristics of the four‐chamber mammalian heart are highlighted by the peculiar structure of the myocardial muscle.
Keywords: circulatory system, comparative anatomy, evolution, heart, vertebrate
Introduction
The heart is the centre of the circulatory system whose primary role is to pump blood through the pulmonary and systemic systems of the body network (Holmes, 1975; Standring, 2015). The circulatory system is a body‐wide network of vessels that transports nutrients, respiratory gases, metabolic waste and hormones, distributes and dissipates heat and assists in defending the body against disease. The first heart‐like organ appeared in our biological history over 500 million years ago (Mya) and has undergone many changes and adaptations during its evolution from a single‐layered tube with own contractility supporting an open circulatory system, to a powerful four‐chambered muscular pump devoted to loading and unloading a large amount of blood around a closed, valved circuit circulatory system. The vertebrate heart is biologically specific to a species and is the product of millions of years of fine tuning (Olson, 2006).
Multicellular life
The first multicellular organisms originated 1.5 Bya (for a detailed timeline and updated phylogenetic classification see Lecointre & Le Guyader, 2007 and revised higher order classifications by Ruggiero et al. 2015) and over time became more specialised and diverse, giving rise to the many plants, animals and fungi we see today (Hofkin, 2010). About 750 million years (My) after the appearance of multicellular life we saw the arrival of the Phylum Porifera (sponges), the most ancient metazoan (Chang et al. 2017). The body plans of the earliest multicellular organisms are asymmetric and do not have a head or tail. Sponges are diploblastic, meaning they only have two embryonic layers, the endoderm and ectoderm (Bishopric, 2005). Sponges are sedentary filter‐feeding organisms that have no need for a circulatory system and use the water currents to obtain oxygen and nutrients that filter through the multiple pores in its ectoderm (osculi; Hoffmann et al. 2005). As biology became more organised, radial symmetry became evident in the Phylum Cnidaria (600 Mya), such as the jellyfish (Subphylum Medusozoa; Swire, 2011; Bettex et al. 2014). Like sponges, jellyfish are also diploblastic, with inner and outer cell layers divided by a gelatinous matrix (Bishopric, 2005). Jellyfish also have no need for a circulatory system as all their cells are in contact with the water in their gastrovascular cavity. Once the prey is ingested through their mouth, seawater is circulated by the cilia inside the jellyfish cavity and oxygen, nutrients and waste simply diffuse directly into and out of each individual cell (Karleskint, 2012).
Bilaterian animals
Biology became more complex 700–600 Mya with the emergence of bilateral symmetry and the introduction of a third germ layer in the bilateralians (Subkingdom Bilateria sensu, Ruggiero et al. 2015; Bishopric, 2005). Bilateral symmetry is accomplished by the orthogonal juncture of an anterior‐posterior (A‐P) axis and an additional dorsal‐ventral (D‐V) axis allowing for a top, bottom, left and right. It is the evolutionary descent that includes more than 1.5 million species comprising flyers, swimmers, runners and burrowers (Finnerty et al. 2004). One major difference that is apparent in the bilaterian and absent in the cnidarians is the presence of a third germ layer called mesoderm, of which the heart and circulatory system is one of its greatest achievements (Gilbert, 2000). Yet, as a jellyfish enters its reproductive stage, nerve cells, sensory cells, as well as specialised non‐myoepithelial cells including striated cells can be found in a layer (Bishopric, 2005) called entocodon that separates away from the ectoderm. Entocodon of the cnidarians is comparable to the mesodermal germ layer found in bilaterian animals. Specific genes have been isolated in the jellyfish species Podocoryne carnea including Brachyury, Mef2, and Snail which have been shown to play a role in myogenesis (Spring et al. 2002), leading to the conclusion that these different genes may be the primordial beginnings of the heart and circulatory system that we see in later bilaterian species.
Primitive heart tube
The primordial gene patterns involved in the organisation of the A‐P and D‐V axis seen in bilaterian development are similar in the invertebrate insects and vertebrates, indicative of a common ancestor prior to the divergence some 600 Mya of the deuterostomes (Superphylum Deuterostomia; alt. Infrakingdom Deuterostomia sensu Ruggiero et al. 2015) and the polyphyletic protostomes (Infrakingdom Protostomia; which include arthropods and other invertebrates, annelids and worms, molluscs and cephalopods; Sedmera & Wang, 2012; Bettex et al. 2014).
Replacing the gastrovascular cavity of the earlier metazoans, the deuterostome body design had a throughgut with an opening at the head and again at the rear. The appearance of the mouth meant separation of gas exchange from digestive function. Another critical beginning in the deuterostome heart evolution approximately 570–540 Mya is the multiplication and radiation of specific genes that encoded for contractile proteins such as actin, myosin and troponin (Oota & Saitou, 1999).
The glimpse of the first circulatory primordium most likely occurred in an ancestral bilaterian (Bishopric, 2005; Olson, 2006) approximately 500 Mya (Olson, 2006). This system most likely resembled that of the most primitive urchordates (e.g. tunicate) or cephalochordate (e.g. amphioxus); the former are considered the closest living relatives of the invertebrate progenitor of vertebrates (Yu & Holland, 2009). The circulatory system of both amphioxi and tunicate species did not have a definitive heart, but included a single‐layered tube with pulsatile contractility, in support of an open circulatory system (Fig. 1; Olson, 2006). An open system involved high output of fluid but under low pressure, which performs well over short distances (Bettex et al. 2014). This system allowed fluid to be forced from the ventral to dorsal aorta through paired aortic arches (Fig. 1; Holmes, 1975), delivering oxygen and nutrients to the cells. However, without any defining chambers, septa or valves these simple early heart tubes did not support a one‐way flow (Bishopric, 2005; Olson, 2006).
Figure 1.
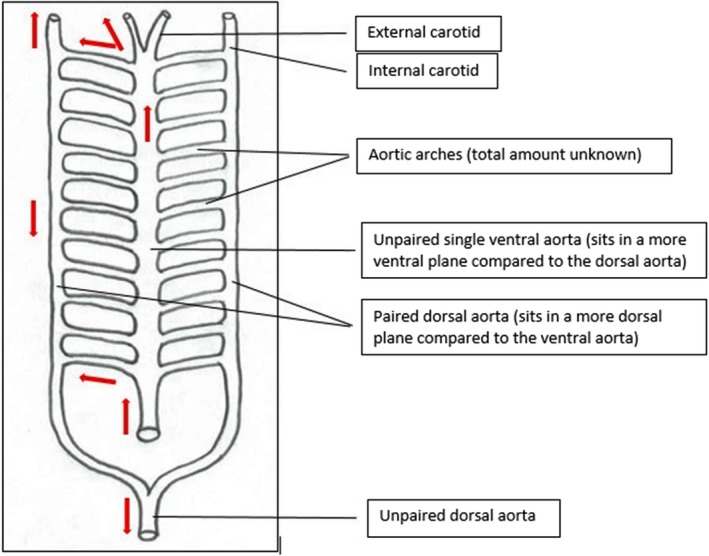
Ventral schematic of prevertebral tubular heart. Blood flow is indicated by red arrows (adapted from Holmes, 1975).
Vertebrate heart
All chordates possess a dorsal nerve cord and a notochord which is supported by surrounding muscle (Bishopric, 2005). The three major subphyla of the Phylum Chordata are the urchordates (Subphylum Urchordata), cephalochordates (Subphylum Cephalochordata) and vertebrates (Subphylum Vertebrata). Vertebrates are distinguished by a multi‐chambered heart and a closed vascular system with capillaries lined with endothelial cells (Bettex et al. 2014).
Whereas the tunicates and amphioxi species possess tubular hearts that contract peristaltically (Bishopric, 2005), evolutionary gene mutations ensured that the anatomy and physiology of the vertebral heart became more complex (Fishman & Olson, 1997). The vertebrate heart is formed via the folding of the lateral cardiogenic mesodermal layer of the embryonic disk to create the tubular heart. The heart section of the tube then loops around and produces a protruding ventricle and atrium, leaving the remaining inner tubular system with its original tubular appearance (Jensen et al. 2013).
Early vertebrate heart
Jawless fish (Infraphylum Agnatha)
Among the most primitive vertebrates in evolutionary history are the agnathans, which include the living hagfish and lampreys (Yasuhiro et al. 2012). The oldest fossil of the hagfish was found in Pennsylvania and dates back to around 300 My (Bardack, 1991); however, agnathans are believed to have evolved over 450 My (Diogo, 2010). Living agnathans lack defining characteristics seen in other vertebrates, such as the absence of a calcified jaw (Alexander, 1986) as well as only a partly calcified skull with a cartilaginous vertebral column (Bishopric, 2005).
Living agnathans have a circulatory system which consists of the main ‘systemic (or brachial)’ heart and three accessory hearts (Fig. 2). The ‘portal’ heart is used to pump blood from the intestines to the liver, the ‘cardinal’ heart pumps blood from the head to the body and ‘caudal’ heart pumps blood from the trunk and kidneys to the rest of the body. The circulatory system is similar to that of a worm and shares with it both open and closed blood vessels (Jorgensen et al. 1998). In comparison with other vertebrates, agnathans also have a different sinus venosus (SV), which makes studying the agnathan circulatory system essential when investigating cardiac evolution. The SV is attached to the left side of the atrium, which has a collagenous wall. A layer of collagen and muscle marks the boundary between the SV and the atrium. Underneath the atrium is a single ventricle, which is attached via an elongated atrioventricular (AV) canal. Small amounts of myocardium can be noted inside the canal, which incorporates a two‐leafed AV valve without any defining papillary muscles (Icardo et al. 2016).
Figure 2.
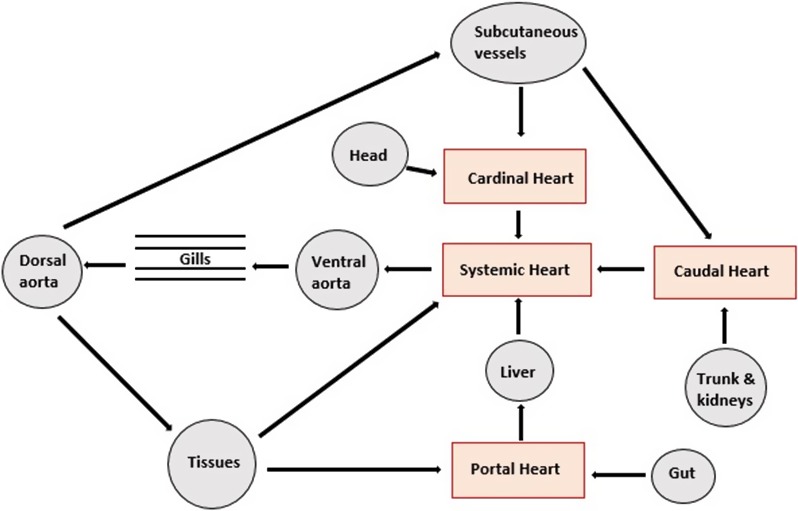
Simplified view of the relationship between the main ‘systemic (or brachial)’ heart and the three accessory hearts of the hagfish (adapted from Jensen, 1965 and Jorgensen et al. 1998).
The circulatory system of the hagfish has not changed in millions of years, with majority of species showing great diversity in habitats, behaviour and environments (Murphy, 1967). As living agnathans such as the hagfish are demersal (e.g. ‘bottom‐dwellers’) they have adapted to an environment that requires low arterial blood pressure along with low cardiac power. A few characteristics specific to this demersal adaptations include hypoxia‐tolerant heart muscle, an extended action potential, as well as the widely spaced intervals in the contractile tissue of the myocardium along with an accessory portal heart supplying the liver.
Lobe‐finned fish (Superclass Sarcopterygii)
Due to oxygen deficiency and the high metabolic cost of obtaining oxygen from water, vertebrates were forced to make their way out of the ocean and on to land 350–400 Mya. This transition gave the heart a more complex role in terms of blood convection and gas transport. The ability to obtain oxygen straight out of the air called for a redesign of the gas chamber, as the small capillaries inside the gills could no longer function efficiently (Jorgensen, 2010).
Anatomical, physiological, genetic and fossil studies have provided evidence that the sacropterygians are a probable vertebrate ancestor to the tetrapods (or sister group; alt. Superclass Tetrapoda sensu Ruggiero et al. 2015; Bassi et al. 2010) and appear in the evolutionary timeline in the Late Devonian period (385–355 Mya; Prothero, 2015). The most studied sarcopterygian circulatory system is that of lungfish (Class Dipnoi), which have two main sites of blood oxygenation. Lungfish primarily use gills to breathe when aquatic, but as they inhabit stagnant ponds and swampland, which are often threatened with drought, lungfish adopted a vascularised lung supplied by two pulmonary arteries that enables them to breathe outside aquatic environments in times of hypoxia (Alexander, 1986; Bettex et al. 2014). With very few exceptions, and in comparison with exclusively aquatic fish, oxygenated blood flow moving out of pulmonary circulation in extant lungfish species does not directly continue into systemic circulation. Instead, it moves back into the heart, where it is then pumped into systemic circulation, ultimately creating dual circulation, the first ever seen in vertebrate evolutionary history. The reason dual circulation was required was because the vascular resistance needed in pulmonary circulation was so high as to dissipate the majority of the kinetic energy conveyed to the blood by the heart (Jorgensen, 2010). As a result of this adjustment, the heart of the lungfish is highly specialised in order to preserve separation of oxygenated and deoxygenated blood through its body cavities (Icardo et al. 2005).
Generally, sarcopterygians breathe by pulling in water through their mouth and out through their gills. A capillary network on each side of their pharynx (throat) allows the fish to pull in oxygen from the water and into their circulatory system, which is then pumped to individual cells (Park et al. 2014). Generally, bony fish have a two‐chambered heart (one atrium and one ventricle) which makes up 0.2% of their mass (Bettex et al. 2014); however, these are subdivided into a linear series of a total of four different compartments: a SV, atrium, ventricle and conus arteriosus (CA; Holmes, 1975).
Oxygen enters the capillaries through the gills and is transported in the blood via the aortic arches, down through the pulmonary arteries and into the lungs, where it is further oxygenated (Icardo et al. 2005; Bettex et al. 2014). In contrast to other osteichthyan (bony) fishes (e.g. Superclass Actinopterygii), the ventral aortic arches are shortened in a lungfish so that they arise closer to the heart (Holmes, 1975). Afferent brachial arteries include the aortic arches between the ventral aorta and the gill capillaries, and the efferent brachial aorta between the gill capillaries and the dorsal aorta. The oxygenated blood then travels through a set of pulmonary veins which unite into a single vein. The now single pulmonary vein empties into the left side of the atria. Blood returning from circulation also enters the atria but via the SV, which has shifted to the right side of the heart. It is here that the oxygenated blood is partially mixed with returning deoxygenated blood from systemic circulation (Icardo et al. 2005; Bettex et al. 2014). The CA has become subdivided into two compartments by an intricate spiral fold (Holmes, 1975). The atria and ventricle are separated by the atrioventricular plug, or cushion. Lungfish have the beginning of a separation in the atria by a pulmonalis fold, and also the separation of the ventricle know as a vertical septum. Therefore, only partial mixing of the blood takes place as oxygenated blood is diverted into the aorta, whereas the deoxygenated blood is diverted back towards the gills and the lungs (Icardo et al. 2005; Bettex et al. 2014). In a hypoxic environment where the lungfish must rely primarily on obtaining oxygen from the air, the capillaries in the gills partially vasoconstrict, but the pulmonary vessels remain open (Bettex et al. 2014).
The Australian lungfish (Neoceratodus forsteri; Order Ceradontiformes) is considered the most primitive of the dipnoids in that it primarily relies on its gills for respiration. The primary cardiovascular anatomical difference relative to other exclusively aquatic bony fish is the pulmonary artery, which arises from both of the six efferent brachial arteries. The African lungfish (Propterus sp.; Order Lepidosireniformes) relies more on its lungs than its gills for respiration, and instead of breaking up into a network of capillaries, two of the aortic arches are carried directly to the dorsal aorta. The South American lungfish (Lepidosiren paradoxa; Order Lepidosireniformes) is the most advanced, as it obtains the majority of its oxygen from the air. The efferent and afferent brachial arteries are in constant communication so that most of the blood flow bypasses the gills (which are still used for carbon dioxide excretion; Holmes, 1975).
The amphibian heart
Amphibians (Class Amphibia, Superclass Tetrapoda) evolved significantly during the Carboniferous period around 350 Mya (Bridgewater, 2012) and show many similarities in heart structure with lungfish. In a more primitive amphibian such as the lungless salamander (e.g. Family Plethodontidae, Order Caudata) the atrium is partially separated; however, in frogs and toads (Order Anura) the atrium is completely separated, resulting in the evolution of a three‐chambered heart (two atria and one ventricle; Fig. 3 for a comparison in the evolutionary tree). Like the lungfish, the SV, CA and pulmonary veins are the same, but there is no ventricular septum (Holmes, 1975). The single ventricle is made of trabeculated myocardium (Bettex et al. 2014) with internal ridges. These ridges decrease the mixing of the oxygenated and deoxygenated blood and divert the blood flow into the appropriate vessels (and may form a partial interventricular septum as in the salamander genus Siren; Family Sirenidae). In their adult form, amphibians do not have gills, therefore the aortic arches do not break up into capillaries for gas exchange (Holmes, 1975).
Figure 3.
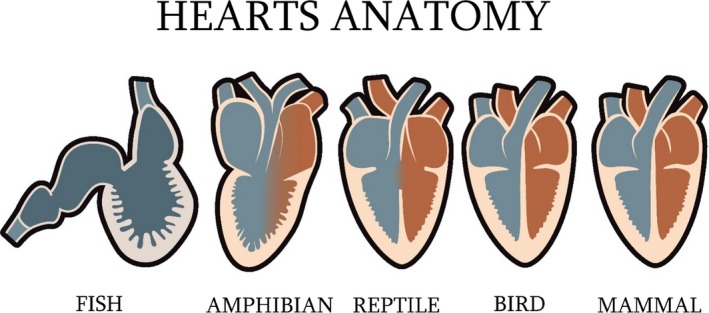
Simplified evolution of the heart shape and chambers.
From the ventricle, the deoxygenated blood is drawn up into pulmonary circulation via the pulmonary artery and the oxygenated blood is pumped into systemic circulation via the aorta (Kisia, 2010; Bettex et al. 2014). The ventral aorta is internally divided into two pathways. One pathway channels blood from the pulmonary artery, which continues to arise from the sixth aortic arch, and all of the systemic blood is diverted into the other arches. Other arches have been lost and modified through evolution to result in the joining of the ventral and dorsal aorta by the fourth aortic arch, which is now known as the arch of aorta (Holmes, 1975).
The lungs of amphibians cannot be relied on for all of their air supply, and other pathways are therefore necessary (Holmes, 1975). The pulmonary arteries of anurans, for example, divert 25% of cardiac output to the skin via cutaneous capillaries. This blood, along with pulmonary return, is 96% saturated with oxygen. Although anurans have the ability to breathe through the skin, the majority of the respiration occurs in the lungs. Physiological adaptations occur when anurans dive underwater or hibernate, such as vasoconstriction of the pulmonary arteries via the vagal stimulation and acetylcholine secretions and vasodilated capillaries in the skin (Bettex et al. 2014). During hibernation, cryoprotectants are secreted from the liver. This stops anurans from freezing when their heart rate and breathing stop (Costanzo & Lee, 2005). Oxygen consumption via the capillaries during skin respiration is enough to maintain life throughout hibernation (Zug et al. 2001; Bettex et al. 2014).
The reptile heart
The earliest known reptiles (Class Reptilia, Superclass Tetrapoda) lived on Earth about 327 Mya (Dixon et al. 2001). The Class is extremely taxonomically diverse and includes lizards and snakes (Subclass Squamata), turtles (Subclass Testudinata), crocodiles (Order Crocodylia, Subclass Crocodylomorpha) as well as the radiation of Dinosauria that includes extant birds (Subclass Aves) and the extinct dinosaurians (Ruggiero et al. 2015; Baron et al. 2017). The anatomy and physiology of the heart across reptiles are therefore very diverse, but in terms of cardiac evolution they can be divided into two groups: the crocodiles and all other non‐crocodile reptile subclasses. In non‐crocodiles such as turtles, a three‐chambered heart with deviating blood flow (shunting) and low pressures can be observed. Anatomically separated compartments with high arterial pressures can be found in some snakes (Order Serpentes, Subclass Squamata), such as species of python (Family Pythoidae), and in the crocodile a highly evolved four‐chambered heart can be found (Bettex et al. 2014). With both three‐ and four‐chambered hearts in this class, assessment of the non‐crocodile reptile heart holds a unique place of importance in cardiac evolution (Kazuko et al. 2009).
The non‐crocodile reptile heart
Extant non‐crocodile reptiles have a three‐chambered heart which accounts for 0.3–0.5% of their bodyweight. The SV is quite small and only a small remnant of the CA remains (Fig. 3, see Holmes, 1975 for further details). The two atria are separated by a complete interatrial septum, whereas a single ventricle has two incomplete septa which allow these reptiles to shunt blood. The major septa is a thick defining muscular ridge originating from the ventral ventricular wall, running from the apex to the base. The right side of the septum is fused with the right dorsolateral ventricle wall, and the anterior part of the septum is free and angled towards, but not attached to, the right wall of the ventricle. This major septa partially separates the ventricle into two cavities, the cavum pulmonale which is the smaller cavity located on the ventral lateral aspect of the ventricle, and a larger dorsolateral cavity. The dorsolateral cavity is further divided into two cavities: a smaller cavum venosum and a larger cavum arteriosum (Hicks, 2002). Most of the ventricle is made up of trabecular myocardium; however, compact myocardium can also be found in the cavum arteriosum (Bettex et al. 2014). The ventral aorta derives from the right side of the ventricle and divides into three major vessels, the pulmonary artery, and left and right aortic arch (Holmes, 1975). These vessels unite distally into a single descending aorta and due to the partially separated ventricle, carry a mixture of oxygenated and deoxygenated blood (Bettex et al. 2014).
During diastole, the regular pathway of blood flow through the non‐crocodile reptile heart starts with oxygenated blood moving from the lungs to the left atrium and into the cavum arteriosum in the left ventricle (Fig. 4). At the same time, deoxygenated blood crosses the SV and right atrium and empties into the cavum venosum and pulmonale. A muscular ridge, vertical septum, atrioventricular valves and laminar blood flow all work together to keep the mixing of oxygenated and deoxygenated blood at a minimal before ventricular ejection (systole). Upon ventricular contraction, deoxygenated blood from the venous cavity enters the pulmonary cavity and is then ejected into the pulmonary artery; oxygenated blood from the cavum arteriosum is directed into the left and right aortas (Fig. 4; Holmes, 1975).
Figure 4.
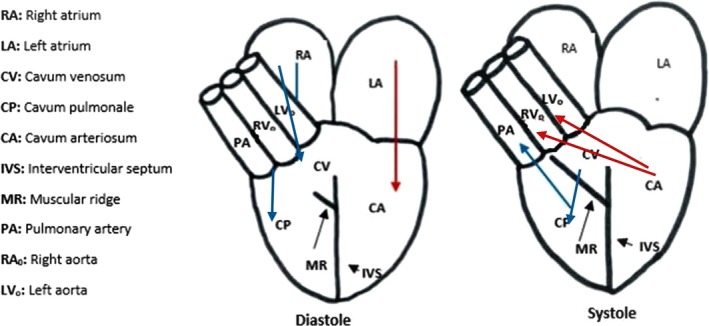
Diastole and systole in a non‐reptile heart. Red arrows indicate oxygen‐rich blood, blue arrows indicate oxygen‐poor blood. Abbreviations explained in the key (adapted from Holmes, 1975 and Bettex et al. 2014).
Non‐crocodile left‐, right and right–left blood shunting
All ectothermic non‐crocodile reptiles have the ability to shunt their own blood through cardiac circulation. The mechanism of shunting is dependent on pulmonary resistance. A left‐to‐right (L‐R) shunt (oxygen‐rich) is caused by a large decrease in pulmonary resistance and a substantial amount of oxygenated blood from the left side of the heart, and is forced into the pulmonary artery (Fig. 5). A right‐to‐left (R‐L) shunt (oxygen‐poor) is caused by a large rise in pulmonary resistance and a substantial amount of deoxygenated blood from the left side of the heart is forced into the two aortas. In many of these non‐crocodile reptiles, lung ventilation is intermittent and an R‐L shunt can be associated with quiet ventilation or diving. During this decrease or cessation of ventilation, the increase in parasympathetic innervation induces bradycardia, which decreases pulmonary blood flow, resulting in an increase in pulmonary resistance. In contrast, an L‐R shunt can be seen in times of increased ventilation which increases sympathetic innervation, inducing tachycardia, increasing pulmonary blood flow and resulting in a decrease in pulmonary resistance (Holmes, 1975; Hicks, 2002; Bettex et al. 2014).
Figure 5.
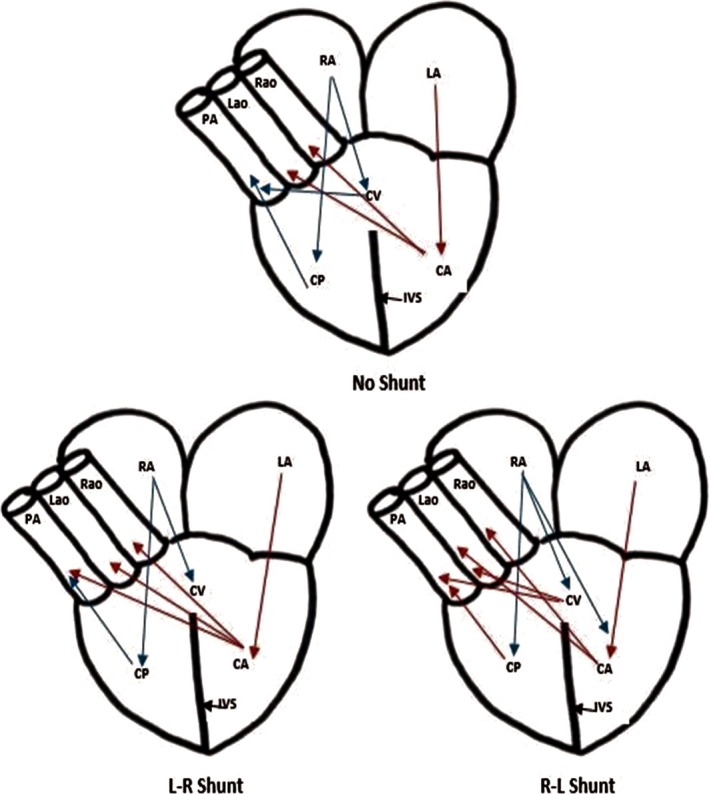
Blood shunting in a non‐crocodilian reptile heart. Red arrows indicate oxygen‐rich blood, blue arrows indicate oxygen‐poor blood. Muscular ridge and valves not shown. Abbreviations explained in the key (adapted from Bettex et al. 2014 and Holmes, 1975).
The crocodile heart
The cardiac anatomy and physiology is different in crocodiles than in other reptiles. Extant crocodiles have retained the dual aortic arches; however, the ventricle is separated by an interventricular septum which is continuous with the interatrial septum (Figs 3, 4, 5). These septa completely divide the heart into four separate chambers with two ventricles and two atria (Holmes, 1975; Hicks, 2002); like the non‐crocodiles, the heart accounts for 0.3–0.5% of bodyweight (Bettex et al. 2014). The interventricular septum also inhibits the crocodiles from achieving an L‐R shunt (oxygen‐rich). Unlike the non‐crocodiles, where all three major vessels (pulmonary artery and left and right aortic arches) arise from the right side of the ventricle, in crocodiles the pulmonary artery and left aortic arch arise from their right ventricle, and the right aortic arch from their left ventricle (Holmes, 1975).
At the base of the aortas is an anatomical structure called the foramen of Panizza. It is this feature that sets crocodiles apart from other non‐crocodile reptiles (Axelsson & Franklin, 1997). It allows the two aortic arches to communicate with each other. During systole, the valves are pushed anteriorly, forcing the foramen of Panizza to close, and therefore blood can enter the aortas (Holmes, 1975). In contrast, during diastole, blood is pushed posteriorly and forces the aortas to close and the foramen of Panizza to open. Interestingly, the left aortic valve remains closed during normal ventricular contraction and blood enters the left aorta through the foramen of Panizza (Axelsson & Franklin, 1997).
Crocodile right–left blood shunting
Due to the interventricular septum of the crocodile heart, two different interventricular blood pressures are permitted. In a non‐shunting situation, the pressure in the right ventricle and pulmonary artery is lower than the blood pressure in the left ventricle and aortas. Blood from the right ventricle enters the pulmonary artery, whereas blood from the left ventricle enters right aorta. The left aorta remains closed (Holmes, 1975). As long as systemic blood flow exceeds the pressure in the right ventricle and pulmonary artery, no R‐L shunt will occur and blood will continue into the pulmonary artery to the lungs to become oxygenated (Axelsson & Franklin, 1997). When a crocodile dives under water, the valve located on the outflow tract of the right ventricular tract closes (cog‐toothed valve) and blood flow to pulmonary circulation is decreased. The increase in right ventricular blood pressure forces the left aortic valve to open, originating an R‐L shunt (Bettex et al. 2014). Physiological and environmental conditions allowing the crocodile to achieve the pulmonary bypass R‐L shunt have been well established (Eme et al. 2010). The R‐L shunt is beneficial to the crocodile as it allows the animal to conserve a large amount of otherwise wasted energy while underwater (Holmes, 1975; Hicks & Wang, 1996).
The avian heart
Extant birds (Subclass Aves, Class Reptilia) are generally regarded as the closest living relatives of the crocodiles. Research into their genome has placed the divergence of crocodile and avian lineages at around 240 Mya (Chatterjee, 2015). The high demand for tissue oxygenation placed on the extant avian body by their adaptations for flight is made possible by a highly efficient four‐chambered heart with separate pulmonary and systemic blood flow. The explanation of the special metabolic demands of avians is a biophysical need in which the ratio of heat loss (a function of body surface area) to heat production (a function of body mass) increases as body size is reduced. Prevention of a fall in body temperature in homoeotherms necessitates an increased metabolic rate, which in turn is correlated with, and is perhaps responsible for, an increased heart rate (Levine 1997). Probably in birds the intense metabolic demand related to their habits reflects their particular high heart rate.
Furthermore, unlike their crocodile relatives, birds are not ectothermic and constantly need to maintain their core temperature at 37 °C. This is achieved by an increase in basal metabolic rate, cardiac output and blood pressure (Bettex et al. 2014). Additionally, with the larger heart and increase in cardiac output, the avian heart and cardiovascular system provide enough oxygen to body tissues to enable and maintain flight (Rosenthal & Harvey, 2010).
The avian heart (Fig. 3) is encased in a fibrous fluid‐filled pericardial sac located in the thoraco‐abdominal cavity (Whitlow, 2000). Blood enters the right atrium, which is larger than the left; however, both are thin‐walled and have no inflow valves. Blood then flows into the right ventricle. The right atrium and right ventricle are separated by an atrioventricular valve which consists of spiral plane of myocardium. The right ventricle is smaller than the left as it does not require much force to eject the blood through an outflow valve, into the pulmonary artery and into pulmonary circulation. Once oxygenated, blood re‐enters the left atrium via the pulmonary veins. Blood flows from the left atrium, through the left atrioventricular valve and into the left ventricle. The left ventricle is two to three times more muscular than the right ventricle, as it needs excessive power to eject blood into the aortic arch and to the rest of the body. This larger left ventricle extends to the apex of the heart. It is separated from the aortic arch by an aortic outflow valve (Rosenthal & Harvey, 2010). The origin of the aortic arch is associated with the right fourth aortic arch (Holmes, 1975). The SV is lost in the majority of avian species, but remnants can be seen in the extant ostrich (Struthio camelus; Order Struthioformes, Infraclass Paleognathae) and the chicken (Gallus gallus domesticus; Order Galliformes, Infraclass Neognathae; Bettex et al. 2014).
The evolution of the avian heart also resulted in the loss of cardiac blood shunting, in favour of complete division of oxygenated and deoxygenated blood with dual circulation. Anatomically speaking, this occurred with the loss of an aorta, leaving a single aorta behind (Holmes, 1975). The average avian systemic blood pressure is 150/70, whereas the pulmonary blood pressure is 20/10; therefore the two atria and two ventricles sit next to each other to equalise pressure as blood flows throughout the heart (Bettex et al. 2014). The avian heart has specific mechanisms which are unique to its own specific function (Rosenthal & Harvey, 2010). Compared with other vertebrates, the avian heart is significantly heavier in proportion to the body and accounts for 0.8–1.2% of a bird's bodyweight (Bettex et al. 2014). In comparison with a similar sized tetrapod mammal (Class Mammalia), although similarities can be seen in the size of the stroke volume, cardiac output is six times higher. Birds also have a high heart rate. A hummingbird (Family Trochilidae, Order Apodiformes, Infraclass Neoaves) heart can beat 520–1260 times per minute (Rosenthal & Harvey, 2010; Bettex et al. 2014). Interestingly, heart rate is a marker of longevity, being inversely related to the life span among homoeothermic mammals and within individual species (Poirier, 2014; Levine, 1997). Of note, mammals that have a slower average heart rate tend to live much longer than those with a faster heart rate (Levine, 1997, Gent et al. 2015; Zhang & Zhang, 2009).
The mammalian heart
The earliest known reptiles lived on Earth about 210 Mya (Dixon et al. 2001). As with the extant avians within the larger extinct dinosaur radiation, the mammal tetrapod radiation also evolved separate circulation. The complete interventricular septum completes a four‐chambered heart (Fig. 3), accounting for 0.6% of a mammal's bodyweight (Bettex et al. 2014). The SV has evolved to become the sino‐atrial node in the atrial wall. Unlike avians, the origin of the aorta leaving the heart is associated with the left fourth aortic arch. Evolutionary anatomical changes are associated which this fourth paired aortic arch. Whereas the right paired dorsal aorta is lost, the left becomes the arch of aorta. Blood flow through the heart is similar to that in avians. Deoxygenated blood turns into the right atrium via the superior and inferior vena cava. It then passes through the tricuspid valve and enters the right ventricle. From here, blood is ejected through the pulmonary valve and into pulmonary circulation, where it is oxygenated by the lungs. Oxygenated blood returns into the left atrium via the pulmonary veins. It goes through the mitral valve into the left ventricle. The left ventricle ejects the blood by force through the aortic valve, into the aortic arch and to the rest of the body (Holmes, 1975). In humans, the average system blood pressure is 120/60 mmHg, whereas pulmonary blood pressure is 25/15 mmHg. The high pressure in systemic circulation allows for fast transfer of oxygen and nutrients, which could have been beneficial during times of hunting when a high metabolism was crucial. In contrast, the low pressure of pulmonary circulation facilitates oxygen loading in the lungs (Bettex et al. 2014).
In regard to the particular characteristics and organisation structure of the mammalian heart, a major advance has been the proposal of the myocardial structure as a helical shape by Torrent‐Guasp and Buckberg (Torrent‐Guasp et al. 2001; Kocica et al. 2006; Buckberg, 2001; Buckberg et al. 2008, 2015; Coghlan & Coghlan, 2001). The proposed helicoidal structure is a product of an embryologic imprinting and was found across major mammalian radiations (e.g. Orders Primates, Carnivora, Cetartiodactyla, Perissodactyla, Lagomorpha; Buckberg, 2001, 2015; Coghlan & Coghlan, 2001; Torrent‐Guasp et al. 2001; Kocica et al. 2006; Buckberg et al. 2008, 2015).
The description of the helical structure identifies the ventricular myocardial band as a functional unit of this complex tissue organisation (Torrent‐Guasp et al. 2001; Kocica et al. 2006; Buckberg, 2015).
In mammals, the cardiac helix includes two simple loops that start at the pulmonary artery and end in the aorta. The ‘contractile band’ is responsible for a spiral horizontal inferior loop that surrounds the two ventricles, with a change in direction originating the double helical structure of the ventricular muscle mass. This peculiar ‘rope structure’ explains why cardiac contraction is more similar to a mop torsion than to a rugby balloon contraction (Torrent‐Guasp et al. 2001; Kocica et al. 2006).
The torsion of the ventricles during cardiac activity both improves ejection efficiency and allows active suction during cardiac filling (this main property of heart mechanics can be explained by such a morphological feature and nicely fits it). The heart evolves indeed from the singular tube (as seen above) in the chordate ancestor, into a dual pumping chamber with separate right and left sides. The complex structure of the heart (a triple figure‐of‐eight spiral band with three S‐shaped helixes, see Torrent‐Guasp et al. 2001; Kocica et al. 2006; Buckberg, 2001, 2015; Buckberg et al. 2008, 2015 for images and details) correlates with the conventional embryologic development.
This complex shape is a crucial evolutionary factor of cardiac functioning and in a clinical setting is a major prognosis determinant with ejection fraction (EF). The mop structure increases the EF by around 50% in a dilated heart and is definitely one the most important consequences of the evolutionary process. Recent echocardiographic evidence of the helical ventricular myocardial band (HVMB; Buckberg, 2015; Hayabuchi et al. 2015) in modern human children supports this relevant aspect of cardiac functional anatomy and sheds further light on the link between heart shape/structure and function.
Although the HVMB model was not accepted by some (Anderson et al. 2005; Lunkenheimer et al. 2006; Gilbert et al. 2007), further imaging and magnetic resonance imaging (MRI) have supported the original proposal by Torrent‐Guasp et al. (Helm et al. 2005; Torrent‐Guasp et al. 2005; Nasiraei‐Moghaddam & Gharib, 2009; Sosnovik et al. 2009; Buckberg, 2015; Hayabuchi et al. 2015).
An explanation from an evolutionary point of view of this special shape can be inferred by mechano‐transmission (Takahashi et al. 2013; Swynghedauw, 2016). Specifically, mechano‐transmission generates protein folding that adapts to physical forces influencing and modifying transcriptional networks and protein expression in the myocardial tissue. Each structure that forms the heart senses mechanical stimuli, including the extracellular matrix, focal adhesion complexes, lipid bilayers and cellular orientation, and each of these structures plays a role in mechano‐transduction. In addition, numerous proteins are involved in mechano‐transduction, including integrins, Rho kinase, PI3K, integrin‐linked kinase, focal adhesion kinase, extracellular signal‐regulated kinase, MAP kinase, eNOS and others. These proteins are involved in cellular mechano‐transduction pathways that elicit several heart responses, including arrhythmias, hypertrophy and ischaemic heart disease (Takahashi et al. 2013; Swynghedauw, 2016). During development, cells, tissues and organs assume their characteristic shapes by sensing mechanical stimuli and responding to them. The heart is an organ that first starts functioning in vertebrate embryos. Appropriate elasticity is required for calcium excitation and contraction of the cardiomyocytes (Majkut & Discher, 2012). In fact, substrate stiffness influences the heart rate contraction forces, the cytoskeletal structure and intracellular calcium levels in cardiomyocytes (Bajaj et al. 2010; Rodriguez et al. 2012).
Another key factor involved in cardiogenesis and heart chamber shape is the rhythmic and sustained cyclic stretching of cardiomyocytes, which is caused by pulsatile changes in cardiac internal pressure. Ott et al. (2008) performed interesting experiments in which cardiac cells were reseeded onto a de‐cellularised heart matrix. When pulsatile perfusion was applied, thick viable cardiac muscles were obtained, whereas in a non‐perfusion environment, thin, weak muscles were obtained. Interestingly, cyclic stretching induces orientation of cardiomyocytes that is transverse to the stretch axis (Salameh et al. 2010). Thus, mechanical forces can affect intercellular communications via gap junction channels in the heart. Changes in blood flow patterns can impair cardiac septation and valve formation (Culver & Dickinson, 2010; Takahashi et al. 2013).
Conclusion
Convergence is the tendency of independent species to evolve similarly when subjected to the same environmental conditions (Pontarotti, 2016). The primitive blueprint for the heart and circulatory system emerged with the arrival of the third mesodermal germ layer in bilaterians. Since then, they have evolved from a single layered tube to a multiple chambered heart some 500 My later. Although the heart is physiologically specific to the anatomy of each individual species, it has evolved along the same directional path. Of note, recent contributions are making a robust case that the hearts of the amniotic vertebrates, and mammals in particular, birds and reptiles share a common building plan during their development, when evolutionarily maintained transcription factors master the growth of heart chambers and related structures (Olson, 2006; Fishman & Olson, 1997). Very recent and careful segmental study of the crocodilian heart highlighted common features between crocodilian and human heart as well as substantial differences, underscoring at the same time a common aspect likely derived from evolutionarily maintained morphology programmes (Cook et al. 2017). The physiological adaptations of the avian heart include the larger sized heart in comparison with the rest of its body, an increased heart rate, and six times the cardiac output volume when compared with other vertebrates. Ultimately, the peculiar shape of the myocardial muscle in mammals is important to understand human heart function in various pathophysiological settings.
Author contributions
AS: Literature research and review, assembly and draft of the manuscript and figures, final approval. JA: Evolutionary biology and cladistics expertise, critical review of the related terminology, final revision of the manuscript. MV: Idealisation and basic concepts, literature research and review, critical revision of the manuscript and final review, final approval.
Conflict of interest
None reported.
References
- Alexander RM. (ed.) 1986) The Collins Encyclopedia of Animal Biology. London: Collins. [Google Scholar]
- Anderson RH, Ho SY, Redmann K, et al. (2005) The anatomical arrangement of the myocardial cells making up the ventricular mass. Eur J Cardiothorac Surg 28, 517–525. [DOI] [PubMed] [Google Scholar]
- Axelsson M, Franklin CE (1997) From anatomy to angioscopy: 164 Years of crocodilian cardiovascular research, recent advances, and speculations. Comp Biochem Physiol A Physiol 118, 51–62. [Google Scholar]
- Bajaj P, Tang X, Saif TA, et al. (2010) Stiffness of the substrate influences the phenotype of embryonic chicken cardiac myocytes. J Biomed Mater Res A 95, 1261–1269. [DOI] [PubMed] [Google Scholar]
- Bardack D (1991) First fossil hagfish (myxinoidea), a record from the Pennsylvanian of Illinois. Science 254, 701–703. [DOI] [PubMed] [Google Scholar]
- Baron MG, Norman DB, Barrett PM (2017) A new hypothesis of dinosaur relationships and early dinosaur evolution. Nature 543, 501–506. [DOI] [PubMed] [Google Scholar]
- Bassi M, Giusti H, Da Silva GS, et al. (2010) Blood gases and cardiovascular shunt in the South American lungfish (Lepidosiren paradoxa) during normoxia and hyperoxia. Respir Physiol Neurobiol 173, 47–50. [DOI] [PubMed] [Google Scholar]
- Bettex DA, Prêtre R, Chassot PG (2014) Is our heart a well‐designed pump? The heart along animal evolution. Eur Heart J 35, 2322–2332. [DOI] [PubMed] [Google Scholar]
- Bishopric NH (2005) Evolution of the heart from bacteria to man. Ann N Y Acad Sci 1047, 13–29. [DOI] [PubMed] [Google Scholar]
- Bridgewater S (2012) A Natural History of Belize: Inside the Maya Forest. Austin: University of Texas Press. [Google Scholar]
- Buckberg GD (2001) The structure and function of the helical heart and its buttress wrapping. II. Interface between unfolded myocardial band and evolution of primitive heart. Semin Thorac Cardiovasc Surg 13, 320–332. [DOI] [PubMed] [Google Scholar]
- Buckberg GD (2015) The helical ventricular myocardial band during standard echocardiography: a structure‐function relationship. Echocardiography 32, 199–204. [DOI] [PubMed] [Google Scholar]
- Buckberg G, Hoffman JI, Mahajan A, et al. (2008) Cardiac mechanics revisited. The relationship of cardiac architecture to ventricular function. Circulation 118, 2571–2587. [DOI] [PubMed] [Google Scholar]
- Buckberg GD, Hoffman JIE, Coghlan HC, et al. (2015) Ventricular structure–function relations in health and disease: Part I. The normal heart. Eur J Cardiothorac Surg 47, 587–601. [DOI] [PubMed] [Google Scholar]
- Chang S, Feng Q, Clausen S, et al. (2017) Sponge spicules from the Lower Cambrian in the Yanjiahe Formation, South China: the earliest biomineralizing sponge record. Palaeogeogr Palaeoclimatol Palaeoecol 474, 36–44. [Google Scholar]
- Chatterjee S (2015) The Rise of Birds: 225 Million Years of Evolution, 2nd edn Baltimore: Johns Hopkins University Press. [Google Scholar]
- Coghlan HC, Coghlan L (2001) Cardiac architecture: Gothic versus Romanesque. A cardiologist's view. Semin Thorac Cardiovasc Surg 13, 417–430. [DOI] [PubMed] [Google Scholar]
- Costanzo JP, Lee RE Jr (2005) Cryoprotection by urea in a terrestrially hibernating frog. J Exp Biol 208, 4079–4089. [DOI] [PubMed] [Google Scholar]
- Cook AC, Tran V‐H, Spicer DE, et al. (2017) Sequential segmental analysis of the crocodilian heart. J Anat, 1–16. doi.org/10.1111/joa.12661, in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Culver JC, Dickinson ME (2010) The effects of hemodynamic force on embryonic development. Microcirculation 17, 164–178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diogo R (2010) Muscles of Vertebrates: Comparative Anatomy, Evolution, Homologies and Development. Hoboken: Taylor and Francis. [Google Scholar]
- Dixon D, et al. (2001) Cassell's Atlas of Evolution: The Earth, Its Landscape and Life Forms. London: Orion Publishing Co. [Google Scholar]
- Eme J, Gwalthney J, Owerkowicz T, et al. (2010) Turning crocodilian hearts into bird hearts: growth rates are similar for alligators with and without right‐to‐left cardiac shunt. J Exp Biol 213(Pt 15), 2673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finnerty JR, Pang K, Burton P, et al. (2004) Origins of bilateral symmetry: Hox and dpp expression in a sea anemone. Science 304, 1335. [DOI] [PubMed] [Google Scholar]
- Fishman MC, Olson EN (1997) Parsing the heart: genetic modules for organ assembly. Cell 91, 153–156. [DOI] [PubMed] [Google Scholar]
- Gent S, Kleinbongard P, Dammann P, et al. (2015) Heart rate reduction and longevity in mice. Basic Res Cardiol 110, 2–9. [DOI] [PubMed] [Google Scholar]
- Gilbert S (2000) Development Biology, 6th edn Sunderland: Sinauer Associates. [Google Scholar]
- Gilbert SH, Benson AP, Li P, et al. (2007) Regional localisation of left ventricular sheet structure: Integration with current models of cardiac fibre, sheet and band structure. Eur J Cardiothorac Surg 32, 231–249. [DOI] [PubMed] [Google Scholar]
- Hayabuchi Y, Sakata M, Kagami S (2015) Assessment of the helical ventricular myocardial band using standard echocardiography. Echocardiography 32, 310‐308. doi:10.1111/echo.12624. [DOI] [PubMed] [Google Scholar]
- Helm P, Beg MF, Miller MI, et al. (2005) Measuring and mapping cardiac fiber and laminar architecture using diffusion tensor MR imaging. Ann N Y Acad Sci 1047, 296–307. [DOI] [PubMed] [Google Scholar]
- Hicks JW (2002) The physiological and evolutionary significance of cardiovascular shunting patterns in reptiles. News Physiol Sci, 17, 241–245. [DOI] [PubMed] [Google Scholar]
- Hicks J, Wang T (1996) Functional role of cardiac shunts in reptiles. J Exp Zool 275, 204–216. [Google Scholar]
- Hoffmann F, Larsen O, Tore Rapp H, et al. (2005) Oxygen dynamics in choanosomal sponge explants. Mar Biol Res 1, 160–163. [Google Scholar]
- Hofkin B (2010) Living in a Microbial World. New York: Garland Science, Taylor & Francis Group LLC. [Google Scholar]
- Holmes EB (1975) Manual of Comparative Anatomy: A Laboratory Guide and Brief Text. New York: Macmillan. [Google Scholar]
- Icardo JM, Ojeda JL, Colvee E, et al. (2005) Heart inflow tract of the African lungfish Protopterus dolloi . J Morphol, 263, 30–38. [DOI] [PubMed] [Google Scholar]
- Icardo JM, Colvee E, Schorno S, et al. (2016) Morphological analysis of the hagfish heart. II. The venous pole and the pericardium. J Morphol 277, 853–865. [DOI] [PubMed] [Google Scholar]
- Jensen B, Wang T, Christoffels V, et al. (2013) Evolution and development of the building plan of the vertebrate heart. Biochim Biophys Acta 1833, 783–794. [DOI] [PubMed] [Google Scholar]
- Jensen D (1965) The aneural heart of the hagfish. Annals of the New York Academy of Sciences 127, 443‐458. doi:10.1111/j.1749‐6632.1965.tb49418.x. [DOI] [PubMed] [Google Scholar]
- Jorgensen JM (2010) The Biology of Lungfishes. Hoboken: Taylor and Francis. [Google Scholar]
- Jorgensen JM, Lomholt JP, Weber RE, et al. (1998) The Biology of Hagfishes. London: Chapman and Hall. [Google Scholar]
- Karleskint G (2012) Introduction to Marine Biology, 4th edn, Vol. 27. Portland: Ringgold Inc. [Google Scholar]
- Kazuko KT, Alessandro DM, Bogac LK, et al. (2009) Reptilian heart development and the molecular basis of cardiac chamber evolution. Nature 461, 95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kisia SM (2010) Vertebrates: Structures and Functions. Parkway, NW: CRC Press. [Google Scholar]
- Kocica MJ, Corno AF, Carreras‐Costa F, et al. (2006) The helical ventricular myocardial band: global, three dimensional, functional architecture of the ventricular myocardium. Eur J Cardiothorac Surg 29S, S21–S40. [DOI] [PubMed] [Google Scholar]
- Lecointre G, Le Guyader H (2007) The Tree of Life: A Phylogenetic Classification, 1st edn Cambridge, MA: Belknap Press Harvard University Press Series. [Google Scholar]
- Levine HJ (1997) Rest heart rate and life expectancy. J Am Coll Cardiol 30, 1104–1106. [DOI] [PubMed] [Google Scholar]
- Lunkenheimer PP, Redmann K, Westermann P, et al. (2006) The myocardium and its fibrous matrix working in concert as a spatially netted mesh: A critical review of the purported tertiary structure of the ventricular mass. Eur J Cardiothorac Surg 29, S41–S49. [DOI] [PubMed] [Google Scholar]
- Majkut SF, Discher DE (2012) Cardiomyocytes from late embryos and neonates do optimal work and striate best on substrates with tissue‐level elasticity: metrics and mathematics. Biomech Model Mechanobiol 11, 1219–1225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murphy RC (1967) Larousse Encyclopedia of Animal Life. London: Paul Hamlyn Ltd. [Google Scholar]
- Nasiraei‐Moghaddam A, Gharib M (2009) Evidence for the existence of a functional helical myocardial band. Am J Physiol Heart Circ Physiol 296, H127–H131. [DOI] [PubMed] [Google Scholar]
- Olson EN (2006) Gene regulatory networks in the evolution and development of the heart. Science 313, 1922–1927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oota S, Saitou N (1999) Phylogenetic relationship of muscle tissues deduced from superimposition of gene trees. J Mol Biol Evol 16, 856–867. [DOI] [PubMed] [Google Scholar]
- Ott HC, Matthiesen TS, Goh SK, et al. (2008) Perfusion‐decellularized matrix: using nature's platform to engineer a bioartificial heart. Nat Med 14, 213–221. [DOI] [PubMed] [Google Scholar]
- Park K, Kim W, Kim HY (2014) Optimal lamellar arrangement in fish gills. Proc Natl Acad Sci U S A 111, 8067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poirier P (2014) Exercise, heart rate variability, and longevity: the cocoon mystery? Circulation 129, 2085‐2087. doi:10.1161/CIRCULATIONAHA.114.009778. [DOI] [PubMed] [Google Scholar]
- Pontarotti P. (ed.) (2016) Evolutionary Biology 2016: Convergent Evolution, Evolution of Complex Traits, Concepts and Methods. Cham: Springer International. [Google Scholar]
- Prothero DR (2015) The Story of Life in 25 Fossils: The Tales of Intrepid Fossil Hunters and the Wonders of Evolution. New York: Columbia University Press. [Google Scholar]
- Rodriguez AG, Han SJ, Regnier M, et al. (2012) Substrate stiffness increases twitch power of neonatal cardiomyocytes in correlation with changes in myofibril structure and intracellular calcium. Biophys J 101, 2455–2464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenthal N, Harvey RP. (eds) (2010) Heart Development and Regeneration. London: Elsevier Inc. [Google Scholar]
- Ruggiero MA, Gordon DP, Orrell TM, et al. (2015) A higher level classification of all living organisms. PLoS One 10, e0119248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salameh A, Wustmann A, Karl S, et al. (2010) Cyclic mechanical stretch induces cardiomyocyte orientation and polarization of the gap junction protein connexin43. Circ Res 106, 1592–1602. [DOI] [PubMed] [Google Scholar]
- Sedmera D, Wang T (2012) Ontogeny and Phylogeny of the Vertebrate Heart. New York: Springer New York. [Google Scholar]
- Sosnovik DE, Wang R, Dai G, et al. (2009) Diffusion MR tractography of the heart. J Cardiovasc Magn Reson 11, 47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spring J, Yanze N, Jösch C, et al. (2002) Conservation of Brachyury, Mef2, and Snail in the myogenic lineage of jellyfish: a connection to the mesoderm of bilateria. Dev Biol 244, 372–384. [DOI] [PubMed] [Google Scholar]
- Standring S (2015) Gray's Anatomy. The Anatomical Basis of Clinical Practice, 41st edn London: Elsevier Health Science. [Google Scholar]
- Swire C (2011) The endosymbiotic theory. Young Sci J 4, 61–64. [Google Scholar]
- Swynghedauw B (2016) Darwinian evolution and cardiovascular remodeling. Heart Fail Rev 21, 795–802. [DOI] [PubMed] [Google Scholar]
- Takahashi K, Kakimoto Y, Toda K, et al. (2013) Mechanobiology in cardiac physiology and diseases. J Cell Mol Med 17, 225–232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torrent‐Guasp F, Buckberg GD, Clemente C, et al. (2001) The structure and function of the helical heart and its buttress wrapping. I. The normal macroscopic structure of the heart. Semin Thorac Cardiovasc Surg 13, 301–319. [DOI] [PubMed] [Google Scholar]
- Torrent‐Guasp F, Kocica MJ, Corno AF, et al. (2005) Towards new understanding of the heart structure and function. Eur J Cardiothorac Surg 27, 191–201. [DOI] [PubMed] [Google Scholar]
- Whitlow G (2000) Sturkies Avian Physiology. San Diego: Academic Press. [Google Scholar]
- Yasuhiro O, Kinya GO, Shigehiro K, et al. (2012) Craniofacial development of hagfishes and the evolution of vertebrates. Nature 493, 175. [DOI] [PubMed] [Google Scholar]
- Yu JK, Holland LZ (2009) Cephalochordates (amphioxus or lancelets): a model for understanding the evolution of chordate characters. Cold Spring Harb Protoc, 2009(9), 1084–1092. doi:10.1101/pdb.emo130 [DOI] [PubMed] [Google Scholar]
- Zhang GQ, Zhang W (2009) Heart rate, lifespan and mortality risk. Ageing Res Rev 8, 52–60. [DOI] [PubMed] [Google Scholar]
- Zug GR, Vitt LJ, Caldwell JP (2001) Herpetology: An Introductory Biology of Amphibians and Reptiles, 2nd edn San Diego: Academic Press. [Google Scholar]