

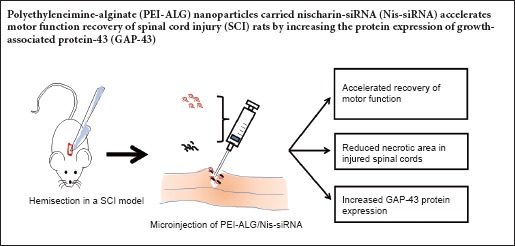
Keywords: nerve regeneration, spinal cord injury, polyethylenimine, alginate, nanoparticles, nischarin, small interfering RNA, necrotic area, growth-associated protein-43, motor function, neural regeneration
Abstract
A previous study by our group found that inhibition of nischarin promotes neurite outgrowth and neuronal regeneration in Neuro-2a cells and primary cortical neurons. In recent years, more and more studies have shown that nanomaterials have good prospects in treatment of spinal cord injury. We proposed that small interfering RNA targeting nischarin (Nis-siRNA) delivered by polyethyleneimine-alginate (PEI-ALG) nanoparticles promoted motor function recovery in rats with spinal cord injury. Direct microinjection of 5 μL PEI-ALG/Nis-siRNA into the spinal cord lesion area of spinal cord injury rats was performed. From day 7 after surgery, Basso, Beattie and Bresnahan score was significantly higher in rats from the PEI-ALG/Nis-siRNA group compared with the spinal cord injury group and PEI-ALG/Control-siRNA group. On day 21 after injection, hematoxylin-eosin staining showed that the necrotic area was reduced in the PEI-ALG/Nis-siRNA group. Immunohistochemistry and western blot assay results confirmed successful inhibition of nischarin expression and increased protein expression of growth-associated protein-43 in the PEI-ALG/Nis-siRNA group. These findings suggest that a complex of PEI-ALG nanoparticles and Nis-siRNA effectively suppresses nischarin expression, induces expression of growth-associated protein-43, and accelerates motor function recovery after spinal cord injury.
Introduction
Spinal cord injury (SCI) often leads to severe disability. Although great progress has been made in the laboratory, so far this has not led to any clinical success (Baptiste et al., 2009; Blesch and Tuszynski, 2009; Ahuja and Fehlings, 2016; Kim et al., 2017). In recent years, with the rapid development of nanotechnology, nanoparticles are being increasingly used as vectors for delivery of therapeutics in vivo. Numerous nanoparticles are currently being tested for therapeutic delivery potential (Bin et al., 2017; Gao et al., 2017; Nejati-Koshki et al., 2017; Song et al., 2017). One of the most extensively studied materials is cationic polymer polyethylenimine (PEI) (Patnaik et al., 2006, 2010; Patnaik and Gupta, 2013). Furthermore, alginate (ALG) is reported to be a good nanomaterial for treating SCI, as it promotes structural repair and functional regeneration following SCI (Grulova et al., 2015; Gunther et al., 2015; Hosseini et al., 2016; Shahriari et al., 2016).
Nischarin is a novel protein discovered in 2000 by Alahari et al (2000). A previous study by our group found that nischarin inhibits neurite outgrowth and regeneration in neuronal cells (Ding et al., 2015). With RNA interference (RNAi) technology, small interfering RNAs (siRNA), small hairpin RNAs (shRNA), or microRNAs (miRNA) can act on target cells to knockdown expression of specific genes (Good et al., 2016; Koenig et al., 2017; Mohammadzadeh et al., 2017). Recently, PEI-ALG self-assembling peptide nanoparticles were shown to carry siRNAs and successfully mediate siRNA gene silencing in vitro (Sharma et al., 2009; Becker and McDonald, 2012; Blesch et al., 2012; Li et al., 2014). However, whether PEI-ALG nanoparticles can deliver nischarin-targeted siRNA (Nis-siRNA) to the spinal cord and promote motor function recovery after SCI in animals is not known.
Using RNAi technology and novel nanoparticles in the present study, we aimed to prepare self-assembling nanocomplexes of PEI-ALG and Nis-siRNA for treating SCI, to preliminarily investigate novel methods for treatment of SCI.
Materials and Methods
Preparation of PEI-ALG nanoparticles
Branched PEI (25 kDa) and sodium alginate were purchased from Sigma-Aldrich (Merck-Millipore, Darmstadt, Germany). PEI-ALG nanoparticles were prepared according to a previously published method, but with certain modifications (Patnaik et al., 2006, 2010). Initially, 5 mg branched PEI (25 kDa) was dissolved in 500 mL distilled water. In a separate vessel, 1 mg alginate was dissolved in 100 mL distilled water under constant stirring at 90°C over 4 hours (Patnaik et al., 2010). Subsequently, the solutions were filtered through 0.22-μm membranes. The alginate solution was then slowly added to the PEI solution while stirring, and then vacuum concentrated in a rotary evaporator followed by lyophilizationon to a final concentration of 0.1 μg/μL. The mixture was incubated at room temperature for 30 minutes with continuous stirring. The final product of PEI-alginate nanoparticles was stored at 4°C until further use.
Preparation of PEI-ALG/siRNA complexes
A commercially available siRNA, containing a pool of three target-specific nischarin siRNA sequences (catalogue No. SC-108099; Santa Cruz Biotechnology, Santa Cruz, CA, USA), was used for preparation of PEI-ALG/siRNA complexes for gene silencing studies. Scrambled siRNA was used as a non-targeting control siRNA (Ctl-siRNA; catalogue No. SC-370007; Santa Cruz Biotechnology). To prepare PEI-AlG/siRNA complexes, Nis-siRNA and Ctl-siRNA were diluted in RNase-free diethylpyrocarbonate-treated water to produce a 1 μg/μL solution. Next, 44 μL PEI-alginate nanoparticles (0.1 μg/μL) were mixed thoroughly with 6 μL siRNA solution. The ratio of polymer amine groups to siRNA phosphate groups in the complex was 10. Solutions were incubated at room temperature for 30 minutes to obtain PEI-ALG/Nis-siRNA or PEI-ALG/Ctl-siRNA complexes, which were subjected to size and zeta potential measurement using a Zetasizer Nano-ZS90 (Malvern Instruments, Malvern, UK) with the following settings: 14 measurements per sample; viscosity for water, 0.89 cP; temperature, 25°C.
Animals
A total of 45 adult female Sprague-Dawley rats weighing 200–250 g and aged 7–8 weeks old were obtained from the Experimental Animal Center of Zhejiang University of China (License No. SCXK (Zhe) 2014-0001). Rats were housed (four per cage) in a temperature (21 ± 1°C) and humidity (55 ± 5%) controlled environment under a 12-hour light/dark cycle with free access to standard food and water.
The study protocol was approved by the Animal Ethics Committee of Zhejiang University of China (ZJU2015-429-01). The experimental procedure followed the United States National Institutes of Health (NIH) Guide for the Care and Use of Laboratory Animals (NIH Publication No. 85-23, revised 1986).
Rat models of SCI
Rats were randomly divided into: SCI group, PEI-ALG group, PEI-ALG/control (Ctl)-siRNA group, and PEI-ALG/Nis-siRNA group (n = 10 in each group), as well as a sham group (n = 5).
Rats were anesthetized by intraperitoneal injection of sodium pentobarbital (50 mg/kg body weight; Sigma-Aldrich, St. Louis, MO, USA). The vertebral column was exposed and T10 laminectomy was performed. Subsequently, the right half of the spinal cord was sectioned with a razor blade (Tachi et al., 2015). Rats in treatment groups received microinjections of 5 μL PEI-ALG, PEI-ALG/Ctl-siRNA, or PEI-ALG/Nis-siRNA into the central lesion site. Rats in the SCI group with no subsequent treatment were used as controls. Sham rats received laminectomy alone. After surgery, rats were treated with normal saline by subcutaneous injection into the fat layer of the back to replace lost fluids. To prevent infection, penicillin (100 mg/d) was administered twice daily for 3 days post-injury.
Right hindlimb movement was assessed by Basso, Beattie and Bresnahan (BBB) locomotor rating scale scores after the animals woke up from anesthesia. Rats that exhibited little or no hindlimb movement were considered successful models (n = 8 in each group), while those with incomplete loss of motor function with BBB scores > 8 were excluded (Umezawa et al., 2017). After assessing BBB locomotor rating scale, three rats in each group were sacrificed for western blot assays, with the remaining five used for morphological studies.
Motor function testing
Scoring according to BBB locomotor rating scale was performed by observers blinded to treatment status. To confirm successful hemisection, BBB scoring was performed on each animal prior to SCI and on the first day after SCI (Basso, 2004). Subsequently, the scoring was repeated on days 3, 7, 14, and 21 after SCI based on ipsilateral hindlimb performance.
Tissue processing
At the end of 21 days post-injury, five rats were randomly selected from each group and sacrificed with an overdose of sodium pentobarbital (100 mg/kg), then transcardially perfused with 0.1 M phosphate buffer, followed by ice-cold 4% paraformaldehyde (catalogue No. P6148, Sigma-Aldrich). Spinal cord tissue was harvested, fixed in 4% paraformaldehyde, and soaked overnight in a sequence of sucrose solutions (10%, 20%, and 30%). Tissue surrounding the lesion site was embedded in optimal cutting temperature compound (Sakura Finetek Japan Co., Ltd., Tokyo, Japan), fast frozen with dry ice, and stored at −80°C until processing. Spinal cord tissue blocks were cut into 30-μm serial longitudinal sections. Every fifth section was mounted on glass slides and stored at −20°C until analyzed.
Hematoxylin-eosin (HE) staining
For general histological examination, spinal cord sections (10 sections per rat) were stained using a HE staining kit (Beyotime Institute of Biotechnology, Inc., Haimen, China), according to the manufacturer's instructions. In brief, sections were rinsed in distilled water and stained with hematoxylin for 5 minutes. After washing, sections were differentiated in 1% acid-alcohol for 30 seconds and incubated with eosin for 30 seconds. After dehydration with ethanol, sections were covered with xylene-based mounting medium and imaged using an Eclipse 50i microscope (Nikon Corporation, Tokyo, Japan). ImageJ software version 1.37 (NIH, Bethesda, MD, USA) was used to measure necrotic areas of spinal tissue in 3-mm segments. In each section, the necrotic area was determined and amount of each section summed per rat to produce total necrotic area per 3-mm segment.
Immunofluorescence staining
Spinal cord sections were subjected to immunostaining, as previously described (Ding et al., 2013). In brief, slices were dried at room temperature, rinsed with 0.01 M phosphate-buffered saline (PBS) for 30 minutes, and blocked in 10% normal goat serum (Gibco, New York, NY, USA) in PBS at room temperature for 2 hours to prevent nonspecific staining. Subsequently, sections were incubated with mouse anti-nischarin monoclonal antibody (1:500 dilution; catalogue No. 558262; BD Biosciences, San Jose, CA, USA) or rabbit anti-growth-associated protein (GAP)-43 polyclonal antibody (1:1,000 dilution; catalogue No. 8945; Cell Signaling Technology, Danvers, MA, USA) overnight at 4°C. Sections were then washed three times with PBS and incubated with fluorescein isothiocyanate (FITC)-labeled goat anti-mouse immunoglobulin (Ig)G (1:200 dilution; catalogue No. 115-095-003; Jackson ImmunoResearch Laboratories, West Grove, PA, USA) or Cy3-labeled goat anti-rabbit IgG (1:200 dilution; catalogue No. 111-165-003; Jackson ImmunoResearch Laboratories) at room temperature for 2 hours, followed by a PBS wash. Specimens were mounted onto gelatin-coated glass microscope slides, air-dried, mounted in glycerol-based mounting medium and observed under a confocal microscope (FV1000; Olympus, Tokyo, Japan). Relative intensity of FITC or Cy3 associated with nischarin or GAP-43 protein content, respectively, was measured using ImageJ software version 1.37 (NIH).
Western blot assay
The remaining animals from each group were anesthetized by intraperitoneal injection of sodium pentobarbital (50 mg/kg) and sacrificed by decapitation on day 21 after hemisection. Next, 10-mm spinal cord segments centered around the injury site were carefully resected and homogenized in 200 μl radioimmunoprecipitation lysis buffer (Beyotime Institute of Biotechnology), supplemented with protease inhibitor cocktail (Sigma-Aldrich, Merck-Millipore). After centrifugation (15,000 × g, 30 minutes, 4°C), supernatants were obtained and protein content estimated using the bicinchoninic acid assay. Total protein (25 μg in each well) was separated by 8–10% (8% gels for nischarin and 10% gels for others) sodium dodecyl sulfate polyacrylamide gel electrophoresis gels. Gels were transferred onto polyvinylidene difluoride membrane (Millipore, Billerica, MA, USA). After blocking with 5% non-fat dry milk (Bio-Rad Laboratories, Inc., Hercules, CA, USA) in tris-buffered saline with Tween-20, membranes were incubated at 4°C overnight with antibodies against nischarin, GAP-43, or β-actin (1:1,000). After incubation with horseradish peroxidase-labeled secondary antibody at room temperature for 2 hours, blots were developed using enhanced chemiluminescence reagents (catalogue No. RPN2235; GE Healthcare Life Sciences, Chalfont, UK) and exposed to X-ray films (Eastman-Kodak, Rochester, NY, USA). Films were scanned and images obtained. The optical density of each band was quantified using Image J software, version 1.37 (NIH, Bethesda, MD, USA) with normalization to β-actin.
Statistical analysis
Data are expressed as the mean ± SEM, and processed with SPSS 12.0 software (SPSS, Chicago, IL, USA). Comparisons between groups were analyzed by two-way analysis of variance followed by Bonferroni's multiple-comparison test or one-way analysis of variance followed by Scheffe's post hoc test. P < 0.05 indicated statistically significant differences between values.
Results
Characterization of nanocomplexes and silencing efficiency of siRNA
Mean size and surface charge of PEI-ALG nanoparticles were 206.9 ± 36.3 nm and 10.1 ± 1.2 mV, respectively. After loading with Nis-siRNA, size and surface charge of the nanoparticles were 243.8 ± 43.6 nm and 8.9 ± 1.7 mV, respectively. Nanoparticles were injected into the injury site of spinal cord hemisection rat models. To measure gene silencing efficiency of Nis-siRNA delivered by nanocomplexes, nischarin expression levels in the injured region of the treated spinal cord were determined by western blot assay after 3 weeks. Compared with the sham group, nischarin expression was upregulated in SCI groups (P < 0.05). Expression of nischarin protein in the PEI-ALG/Nis-siRNA group was significantly decreased compared with the SCI group (P < 0.05), PEI-ALG group (P < 0.05), and PEI-ALG/Ctl-siRNA group (P < 0.05). However, there was no difference among the SCI, PEI-ALG, and PEI-ALG/Ctl-siRNA groups Figure 1A and B). These findings were confirmed by immunofluorescence staining. Compared with the PEI-ALG/Ctl-siRNA group, nischarin expression in the PEI-ALG/Nis-siRNA group was significantly suppressed in the center of the lesion site (injection spot) (P < 0.05), but not the region 1 cm caudal to the injury epicenter (Figure 1C–G).
Figure 1.

Nischarin protein expression in the injured spinal cord of rats.
(A, B) Western blot assay shows markedly upregulated nischarin protein expression in SCI, PEI-ALG, and PEI-ALG/Ctl-siRNA groups compared with the sham group (§P < 0.05). However, nischarin expression was significantly decreased in the PEI-ALG/Nis-siRNA group compared with the SCI group (*P < 0.05), SCI + PEI-ALG group (#P < 0.05), and SCI + PEI-ALG/Ctl-siRNA group (†P < 0.05). (C–G) Immunofluorescence staining to examine nischarin expression. (C, D) Immunofluorescence intensity of nischarin in the lesion site center. The dotted white lines indicate the edges of cavities. (E, F) Immunofluorescence intensity of nischarin in the region 1 cm distal to the lesion site. Scale bar: 100 μm. Data are expressed as the mean ± SEM (n = 3, one-way analysis of variance followed by Scheffe's post hoc test). SCI: Spinal cord injury; PEI-ALG: polyethylenimine-alginate; Nis-siRNA: small interfering RNA targeting nischarin; Ctl: control.
PEI-ALG/Nis-siRNA improved motor function of SCI rats
Within 21 days after SCI, BBB scores in the right hindlimb of SCI rats increased gradually in each group. Compared with the SCI group without any treatment, BBB score in only the PEI-ALG group was significantly increased from day 21 after operation (P < 0.05), while in the PEI-ALG/Ctl-siRNA group, it significantly increased from day 14 (P < 0.05). BBB score in the PEI-ALG/Nis-siRNA group was significantly higher than those in the SCI group (P < 0.01), PEI-ALG group (P < 0.05), and PEI-ALG/Ctl-siRNA group (P < 0.05) from day 7 after operation. These results suggest that PEI-ALG nanoparticles and Nis-siRNA may be jointly used to treat SCI and significantly accelerate hind limb motor function of rats (Figure 2).
Figure 2.
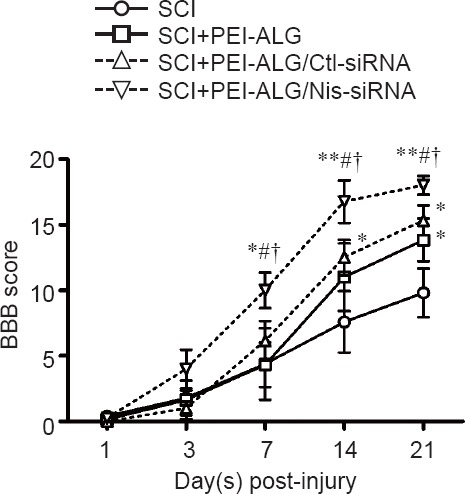
BBB score in rats with SCI at different time points.
Within 3 weeks after spinal cord hemisection, BBB scores increased gradually in rats from different groups, indicating gradual recovery of hindlimb locomotor function. From 7 days after injury, BBB scores were significantly greater in the PEI-ALG/Nis-siRNA group compared with the SCI group (*P < 0.05, **P < 0.01), SCI + PEI-ALG group (#P < 0.05), and SCI + PEI-ALG/Ctl-siRNA group (†P < 0.05). Data are expressed as the mean ± SEM (n = 6–8, two-way analysis of variance followed by Bonferroni multiple-comparison test). SCI: Spinal cord injury; PEI-ALG: polyethylenimine-alginate; Nis-siRNA: small interfering RNA targeting nischarin; Ctl: control; BBB: Basso, Beattie and Bresnahan locomotor rating.
PEI-ALG/Nis-siRNA reduced necrotic area of injured spinal cord
To investigate the influence of PEI-ALG/Nis-siRNA on morphological changes and necrotic area in spinal cord tissue of rats in different groups, necrotic tissue in 3-mm cord segments were stained with HE and observed under light microscopy. The results showed that in the SCI group without treatment, spinal cord tissue structure was destroyed with massive infiltration of inflammatory cells and cavities. Similarly, in the PEI-ALG and PEI-ALG/Ctl-siRNA groups, inflammatory cell infiltration and cavities were also identified in spinal cord tissue. However, in the PEI-ALG/Nis-siRNA group, inflammatory cell infiltration at the injury site was not obvious with smaller cavities (Figure 3A). Quantification of necrotic area in different groups revealed that total necrotic area was reduced in the PEI-ALG/Nis-siRNA group compared with the SCI, PEI-ALG, and PEI-ALG/Ctl-siRNA groups, (P < 0.05; Figure 3B). Consequently, PEI-ALG nanomaterial and Nis-siRNA may be used jointly for treating SCI, protecting injured cells, and reducing inflammatory cell infiltration and necrotic area.
Figure 3.

Hematoxylin-eosin staining of damaged areas in the spinal cord of rats on day 21 after SCI.
(A) Representative SCI images in the four groups (scale bar, 100 μm). Necrotic cells (arrowhead) and infiltrated inflammatory cells (arrows) were observed in necrotic areas. (B) Quantification of necrotic areas in the injured spinal cord region. Necrosis areas were significantly reduced in the PEI-ALG/Nis-siRNA group compared with the SCI group (*P < 0.05), SCI + PEI-ALG group (#P < 0.05), and SCI + PEI-ALG/Ctl-siRNA group (†P < 0.05). Data are expressed as the mean ± SEM (n = 5; one-way analysis of variance followed by Scheffe's post hoc test). SCI: Spinal cord injury; PEI-ALG: polyethylenimine-alginate; Nis-siRNA: small interfering RNA targeting nischarin; Ctl: control.
PEI-ALG/Nis-siRNA promoted GAP-43 protein expression in injured spinal cord
GAP-43 is a neuronal specific protein involved in outgrowth and regeneration of neuronal cells that is highly expressed during axonal regeneration. To clarify the mechanism through which PEI-ALG/Nis-siRNA promotes motor function recovery after SCI, GAP-43 was detected by immunofluorescence staining on longitudinal spinal cord sections of rats from different groups. Compared with the other three groups, GAP-43 expression was increased around the injured area in the PEI-ALG/Nis-siRNA group, with a long filamentous, complete structure present. In contrast, it was less evident in the other three groups (Figure 4A). Quantification of immunofluorescence intensity revealed significantly higher GAP-43 protein expression in the PEI-ALG/Nis-siRNA group compared with the SCI, PEI-ALG, and PEI-ALG/Ctl-siRNA groups (P < 0.01; Figure 4B).
Figure 4.

Effect of Nis-siRNA delivered by PEI-ALG nanoparticles on GAP-43 expression in the injured spinal cord of rats on day 21 after SCI.
(A) At 21 days after SCI, immunofluorescence staining was performed to examine GAP-43 expression. Scale bars: 100 μm. GAP-43-positive structures (arrows) were observed in the injured spinal cord. (B) Quantification of GAP-43 relative fluorescence intensity. GAP-43 expression was increased in the PEI-ALG/Nis-siRNA group compared with the SCI group (**P < 0.01), SCI+PEI-ALG group (##P < 0.01), and SCI + PEI-ALG/Ctl-siRNA group (††P < 0.01). Data are expressed as the mean ± SEM (n = 3; one-way analysis of variance followed by post hoc test). SCI: Spinal cord injury; PEI-ALG: polyethylenimine-alginate; Nis-siRNA: small interfering RNA targeting nischarin; Ctl: control; GAP-43: growth-associated protein-43.
Western blot assay was further applied to detect GAP-43 protein content in injured spinal cord of rats. The results showed that relative expression of GAP-43 in the SCI, PEI-ALG, and PEI-ALG/Ctl-siRNA groups was significantly decreased compared with baseline expression in the sham group (P < 0.05). However, GAP-43 expression in the PEI-ALG/Nis-siRNA group was higher compared with the SCI group (P < 0.05), PEI-ALG group (P < 0.01), and PEI-ALG/Ctl-siRNA group (P < 0.05; Figure 5).
Figure 5.
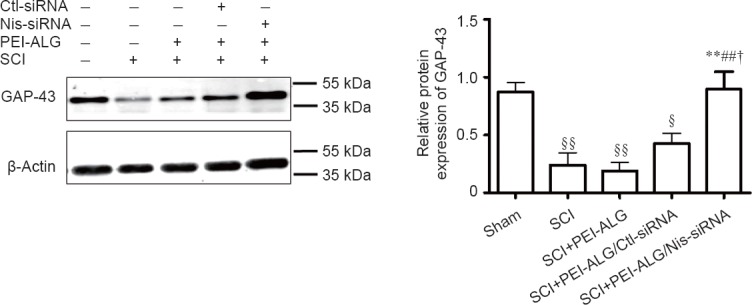
Effect of Nis-siRNA delivered by PEI-ALG nanoparticles on GAP-43 protein expression in the injured spinal cord of rats on day 21 after SCI.
Protein expression of GAP-43 was markedly decreased in the SCI, PEI-ALG, and PEI-ALG/Ctl-siRNA groups compared with the sham group (§P < 0.05, §§P < 0.01). However, GAP-43 protein expression was increased in the PEI-ALG/Nis-siRNA group compared with the SCI group (**P < 0.01), SCI + PEI-ALG group (##P < 0.01), and SCI + PEI-ALG/Ctl-siRNA group (†P < 0.05). Data are expressed as the mean ± SEM (n = 3, one-way analysis of variance followed by Scheffe's post hoc test). SCI: Spinal cord injury; PEI-ALG: polyethylenimine-alginate; Nis-siRNA: small interfering RNA targeting nischarin; Ctl: control; GAP-43: growth-associated protein-43.
Discussion
Due to its high specificity, efficiency and other advantages, RNAi has become a potential treatment for SCI. For example, Sun et al. (2012) used Nogo-shRNA to effectively inhibit protein expression of Nogo and promote SCI repair in rats. Qu et al. (2014) inhibited expression of the ephrin B3 gene, and consequently accelerated motor function recovery in SCI rats. Zukor et al. (2013) demonstrated that suppression of phosphatase and tensin homolog gene expression in mice promotes regeneration of corticospinal tract neurons. The novel protein nischarin, which acts on cytoskeletal actin through Rho-GTPase family members (Alahari et al., 2004; Reddig et al., 2005; Ding et al., 2008; Maziveyi and Alahari, 2015), is highly expressed in the nervous system and inhibits neuronal cell migration (Ding et al., 2013). A recent study by our group found that neurite outgrowth is significantly promoted if nischarin expression is inhibited by siRNAs in vitro (Ding et al., 2015). Thus, we speculated that Nis-siRNA may stimulate axonal regeneration after SCI in vivo.
Due to their good biodegradability, high flexibility, suitable mechanical properties, and absence of immunogenicity, nanoparticles have been studied in the field of biology and medicine, particularly in treatment of SCI, with promising results obtained (Kubinova and Sykova, 2010; Tyler et al., 2013; White-Schenk et al., 2015). For example, Kim et al., (2009) used poly(lactic-co-glycolic acid)-based nanoparticles to deliver the drug methylprednisolone in vitro as well as in a SCI model in vivo. The study reported that local, sustained delivery of methylprednisolone via nanoparticles is significantly more effective in SCI treatment than systemic delivery. Tsai et al. (2006) produced a gel tunnel support with a complex of polyhydroxyethylmethacrylate and methyl methacrylate, filled it with methyl cellulose matrix and then implanted it into a rat with a completely transected T8 spinal cord. After 8 weeks, regeneration of vestibular and red nucleus neurons was significantly increased.
Simultaneously, investigation of the combination of nanoparticles and RNAi technology for clinical treatment is being pursued (Acharya et al., 2017; Gentile et al., 2017). Recent research has focused on SCI treatment using nano-controlled release systems as siRNA carriers to achieve regulation of specific genes at the site of SCI (Li et al., 2016; Gao and Li, 2017). A previous study by our group has shown that ALG gel activates bone marrow mesenchymal stem cells and has unique advantages as a good carrier for stem cell transplantation in SCI treatment (Shi et al., 2011). Cationic PEI has been extensively studied in recent years for its high efficiency of gene delivery in vivo, thereby showing its therapeutic potential in humans (Lai, 2011; Patnaik and Gupta, 2013; Xun et al., 2014; Dabbaghi et al., 2017; Nakamura et al., 2017). Studies have revealed that PEI-ALG nanoparticles effectively deliver siRNAs, prevent enzymatic hydrolysis, and facilitate entry into cells for gene therapy (Patnaik et al., 2006; Patnaik et al., 2010). However, whether PEI-ALG nanoparticles can successfully deliver Nis-siRNA into cells in vivo to effectively interfere with target genes and promote rehabilitation of rats with SCI has remained elusive.
In the present study, a rat hemisection model of SCI was established. PEI-ALG nanoparticles were injected into the injured area, and hind limb motor function and morphological changes of spinal cord tissue in rats from different groups examined. As expected, nischarin expression in the center of lesion sites was significantly decreased at 3 weeks after PEI-ALG/Nis-siRNA injection. Efficient delivery distance of PEI-ALG nanoparticles was limited to the injection site, since nischarin expression was not different from the Ctl-siRNA group in the region 1 cm distal to the lesion site. Compared with the SCI, PEI-ALG, and PEI-ALG/Ctl-siRNA groups, BBB score in rats of the PEI-ALG/Nis-siRNA group began to increase from day 7 following SCI, which indicates that suppression of nischarin expression by Nis-siRNA effectively accelerated motor function recovery of SCI rats. HE staining showed less inflammatory cell infiltration and smaller cavities around the SCI site in the PEI-ALG/Nis-siRNA group. Immunofluorescence staining indicated that PEI-ALG/Nis-siRNA treatment increased GAP-43 expression in the injured spinal cord, which was confirmed by western blot assay. GAP-43 is highly expressed in neuronal growth cones, and GAP-43 upregulation is a useful marker of neurite growth or axonal regeneration (Ondarza et al., 2003; Novotna et al., 2011). Thus, suppression of nischarin expression may promote axonal regeneration by upregulating GAP-43 protein expression, although the specific mechanisms require further study.
Of note, the present study also found higher BBB scores in the PEI-ALG and PEI-ALG/Ctl-siRNA groups than those in the SCI group, particularly on day 21. Based on a previous study by our group, the beneficial effect of PEI-ALG and PEI-ALG/Ctl-siRNA is likely due to the positive impact of ALG hydrogel (Shi et al., 2011). Furthermore, the in vivo therapeutic effect of ALG was recently reported in SCI rat models (Grulova et al., 2015; Tamosaityte et al., 2015).
In summary, PEI-ALG/Nis-siRNA nanoparticles efficiently inhibit in vivo expression of nischarin protein, upregulate expression of GAP-43, and have a beneficial role in motor function recovery after SCI.
Acknowledgments
We thank the staff from Department of Basic Medicine, College of Medicine, Zhejiang University, China, for their assistance in the preparation of PEI-ALG/siRNA complexes.
Footnotes
Conflicts of interest: None declared.
Research ethics: The study protocol was approved by the Animal Ethics Committee of the Zhejiang University (ZJU2015-429-01). The experimental procedure followed the United States National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publication No. 85-23, revised 1986).
Data sharing statement: Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check: Checked twice by iThenticate.
Peer review: Externally peer reviewed.
Funding: This work was supported by the Natural Science Foundation of Zhejiang Province of China, No. LY15H250001 and LY14H090002; the National Natural Science Foundation of China, No. 81000535 and 81402872; the Medical Science and Technology Project Foundation of Zhejiang Province of China, No. 2014KYA166; and the Science and Technology Innovation Talents Development Plan Foundation for High School Students in Zhejiang Province of China, No. 2014R401186.
Copyedited by James R, Norman C, Wang J, Li CH, Qiu Y, Song LP, Zhao M
References
- Acharya R, Saha S, Ray S, Hazra S, Mitra MK, Chakraborty J. siRNA-nanoparticle conjugate in gene silencing: a future cure to deadly diseases? Mater Sci Eng C Mater Biol Appl. 2017;76:1378–1400. doi: 10.1016/j.msec.2017.03.009. [DOI] [PubMed] [Google Scholar]
- Ahuja CS, Fehlings M. Concise review: bridging the gap: novel neuroregenerative and neuroprotective strategies in spinal cord injury. Stem Cells Transl Med. 2016;5:914–924. doi: 10.5966/sctm.2015-0381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alahari SK, Lee JW, Juliano RL. Nischarin, a novel protein that interacts with the integrin alpha5 subunit and inhibits cell migration. J Cell Biol. 2000;151:1141–1154. doi: 10.1083/jcb.151.6.1141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alahari SK, Reddig PJ, Juliano RL. The integrin-binding protein Nischarin regulates cell migration by inhibiting PAK. EMBO J. 2004;23:2777–2788. doi: 10.1038/sj.emboj.7600291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baptiste DC, Tighe A, Fehlings MG. Spinal cord injury and neural repair: focus on neuroregenerative approaches for spinal cord injury. Exp Opin Investig Drugs. 2009;18:663–673. doi: 10.1517/13543780902897623. [DOI] [PubMed] [Google Scholar]
- Basso DM. Behavioral testing after spinal cord injury: congruities, complexities, and controversies. J Neurotrauma. 2004;21:395–404. doi: 10.1089/089771504323004548. [DOI] [PubMed] [Google Scholar]
- Becker D, McDonald JW., 3rd Approaches to repairing the damaged spinal cord: overview. Handb Clin Neurol. 2012;109:445–461. doi: 10.1016/B978-0-444-52137-8.00028-0. [DOI] [PubMed] [Google Scholar]
- Bin S, Zhou N, Pan J, Pan F, Wu XF, Zhou ZH. Nano-carrier mediated co-delivery of methyl prednisolone and minocycline for improved post-traumatic spinal cord injury conditions in rats. Drug Dev Ind Pharm. 2017;43:1033–1041. doi: 10.1080/03639045.2017.1291669. [DOI] [PubMed] [Google Scholar]
- Blesch A, Tuszynski MH. Spinal cord injury: plasticity, regeneration and the challenge of translational drug development. Trends Neurosci. 2009;32:41–47. doi: 10.1016/j.tins.2008.09.008. [DOI] [PubMed] [Google Scholar]
- Blesch A, Fischer I, Tuszynski MH. Gene therapy, neurotrophic factors and spinal cord regeneration. Handb Clin Neurol. 2012;109:563–574. doi: 10.1016/B978-0-444-52137-8.00035-8. [DOI] [PubMed] [Google Scholar]
- Dabbaghi M, Kazemi Oskuee R, Hashemi K, Afkhami Goli A. Evaluating polyethyleneimine/DNA nanoparticles-mediated damage to cellular organelles using endoplasmic reticulum stress profile. Artif Cells Nanomed Biotechnol. 2017 doi: 10.1080/21691401.2017.1304406. doi: 10.1080/21691401.2017.1304406. [DOI] [PubMed] [Google Scholar]
- Ding Y, Milosavljevic T, Alahari SK. Nischarin inhibits LIM kinase to regulate cofilin phosphorylation and cell invasion. Mol Cell Biol. 2008;28:3742–3756. doi: 10.1128/MCB.01832-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ding Y, Li Y, Lu L, Zhang R, Zeng L, Wang L, Zhang X. Inhibition of nischarin expression promotes neurite outgrowth through regulation of pak activity. PLoS One. 2015;10:e0144948. doi: 10.1371/journal.pone.0144948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ding Y, Zhang R, Zhang K, Lv X, Chen Y, Li A, Wang L, Zhang X, Xia Q. Nischarin is differentially expressed in rat brain and regulates neuronal migration. PLoS One. 2013;8:e54563. doi: 10.1371/journal.pone.0054563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gao SJ, Liu Y, Wang HJ, Ban DX, Cheng SZ, Ning GZ, Wang LL, Chang J, Feng SQ. New approach to treating spinal cord injury using PEG-TAT-modified, cyclosporine-A-loaded PLGA/polymeric liposomes. J Drug Target. 2017;25:75–82. doi: 10.1080/1061186X.2016.1191082. [DOI] [PubMed] [Google Scholar]
- Gao W, Li J. Targeted siRNA delivery reduces nitric oxide mediated cell death after spinal cord injury. J Nanobiotechnol. 2017;15:38. doi: 10.1186/s12951-017-0272-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gentile E, Oba T, Lin J, Shao R, Meng F, Cao X, Lin HY, Mourad M, Pataer A, Baladandayuthapani V, Cai D, Roth JA, Ji L. Cationic liquid crystalline nanoparticles for the delivery of synthetic RNAibased therapeutics. Oncotarget. 2017 doi: 10.18632/oncotarget.18421. doi: 10.18632/oncotarget.18421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Good ME, Begandt D, DeLalio LJ, Johnstone SR, Isakson BE. Small interfering RNA-mediated connexin gene knockdown in vascular endothelial and smooth muscle cells. Methods Mol Biol. 2016;1437:71–82. doi: 10.1007/978-1-4939-3664-9_5. [DOI] [PubMed] [Google Scholar]
- Grulova I, Slovinska L, Blasko J, Devaux S, Wisztorski M, Salzet M, Fournier I, Kryukov O, Cohen S, Cizkova D. Delivery of alginate scaffold releasing two trophic factors for spinal cord injury repair. Sci Rep. 2015;5:13702. doi: 10.1038/srep13702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gunther MI, Weidner N, Muller R, Blesch A. Cell-seeded alginate hydrogel scaffolds promote directed linear axonal regeneration in the injured rat spinal cord. Acta Biomater. 2015;27:140–150. doi: 10.1016/j.actbio.2015.09.001. [DOI] [PubMed] [Google Scholar]
- Hosseini SM, Sharafkhah A, Koohi-Hosseinabadi O, Semsar-Kazerooni M. Transplantation of neural stem cells cultured in alginate scaffold for spinal cord injury in rats. Asian Spine J. 2016;10:611–618. doi: 10.4184/asj.2016.10.4.611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim YH, Ha KY, Kim SI. Spinal cord injury and related clinical trials. Clin Orthop Surg. 2017;9:1–9. doi: 10.4055/cios.2017.9.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim YT, Caldwell JM, Bellamkonda RV. Nanoparticle-mediated local delivery of Methylprednisolone after spinal cord injury. Biomaterials. 2009;30:2582–2590. doi: 10.1016/j.biomaterials.2008.12.077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koenig O, Nothdurft D, Perle N, Neumann B, Behring A, Degenkolbe I, Walker T, Schlensak C, Wendel HP, Nolte A. An Atelocollagen coating for efficient local gene silencing by using small interfering RNA. Mol Ther Nucleic Acids. 2017;6:290–301. doi: 10.1016/j.omtn.2017.01.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kubinova S, Sykova E. Nanotechnology for treatment of stroke and spinal cord injury. Nanomedicine (Lond) 2010;5:99–108. doi: 10.2217/nnm.09.93. [DOI] [PubMed] [Google Scholar]
- Lai WF. In vivo nucleic acid delivery with PEI and its derivatives: current status and perspectives. Expert Rev Med Devices. 2011;8:173–185. doi: 10.1586/erd.10.83. [DOI] [PubMed] [Google Scholar]
- Li J, Liu Y, Xu H, Fu Q. Nanoparticle-delivered IRF5 siRNA facilitates M1 to M2 transition, reduces demyelination and neurofilament loss, and promotes functional recovery after spinal cord injury in mice. Inflammation. 2016;39:1704–1717. doi: 10.1007/s10753-016-0405-4. [DOI] [PubMed] [Google Scholar]
- Li T, Wang GD, Tan YZ, Wang HJ. Inhibition of lymphangiogenesis of endothelial progenitor cells with VEGFR-3 siRNA delivered with PEI-alginate nanoparticles. Int J Biol Sci. 2014;10:160–170. doi: 10.7150/ijbs.6719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maziveyi M, Alahari SK. Breast cancer tumor suppressors: a special emphasis on novel protein nischarin. Cancer Res. 2015;75:4252–4259. doi: 10.1158/0008-5472.CAN-15-1395. [DOI] [PubMed] [Google Scholar]
- Mohammadzadeh R, Saeid Harouyan M, Ale Taha SM. Silencing of bach1 gene by small interfering RNA-mediation regulates invasive and expression level of miR-203, miR-145, matrix metalloproteinase-9, and CXCR4 receptor in MDA-MB-468 breast cancer cells. Tumour Biol. 2017 doi: 10.1177/1010428317695925. doi:10.1177/1010428317695925. [DOI] [PubMed] [Google Scholar]
- Nakamura Y, Sato H, Nobori T, Matsumoto H, Toyama S, Shuno T, Kishimura A, Mori T, Katayama Y. Modification of ligands for serum albumin on polyethyleneimine to stabilize polyplexes in gene delivery. J Biomater Sci Polym Ed. 2017 doi: 10.1080/09205063.2017.1328730. doi:10.1080/09205063.2017.1328730. [DOI] [PubMed] [Google Scholar]
- Nejati-Koshki K, Mortazavi Y, Pilehvar-Soltanahmadi Y, Sheoran S, Zarghami N. An update on application of nanotechnology and stem cells in spinal cord injury regeneration. Biomed Pharmacother. 2017;90:85–92. doi: 10.1016/j.biopha.2017.03.035. [DOI] [PubMed] [Google Scholar]
- Novotna I, Slovinska L, Vanicky I, Cizek M, Radonak J, Cizkova D. IT delivery of ChABC modulates NG2 and promotes GAP-43 axonal regrowth after spinal cord injury. Cell Mol Neurobiol. 2011;31:1129–1139. doi: 10.1007/s10571-011-9714-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ondarza AB, Ye Z, Hulsebosch CE. Direct evidence of primary afferent sprouting in distant segments following spinal cord injury in the rat: colocalization of GAP-43 and CGRP. Exp Neurol. 2003;184:373–380. doi: 10.1016/j.expneurol.2003.07.002. [DOI] [PubMed] [Google Scholar]
- Patnaik S, Gupta KC. Novel polyethylenimine-derived nanoparticles for in vivo gene delivery. Expert Opin Drug Deliv. 2013;10:215–228. doi: 10.1517/17425247.2013.744964. [DOI] [PubMed] [Google Scholar]
- Patnaik S, Arif M, Pathak A, Singh N, Gupta KC. PEI-alginate nanocomposites: efficient non-viral vectors for nucleic acids. Int J Pharm. 2010;385:194–202. doi: 10.1016/j.ijpharm.2009.10.041. [DOI] [PubMed] [Google Scholar]
- Patnaik S, Aggarwal A, Nimesh S, Goel A, Ganguli M, Saini N, Singh Y, Gupta KC. PEI-alginate nanocomposites as efficient in vitro gene transfection agents. J Control Release. 2006;114:398–409. doi: 10.1016/j.jconrel.2006.06.025. [DOI] [PubMed] [Google Scholar]
- Qu Y, Zhao J, Wang Y, Gao Z. Silencing ephrinB3 improves functional recovery following spinal cord injury. Mol Med Rep. 2014;9:1761–1766. doi: 10.3892/mmr.2014.2019. [DOI] [PubMed] [Google Scholar]
- Reddig PJ, Xu D, Juliano RL. Regulation of p21-activated kinase-independent Rac1 signal transduction by nischarin. J Biol Chem. 2005;280:30994–31002. doi: 10.1074/jbc.M502546200. [DOI] [PubMed] [Google Scholar]
- Shahriari D, Koffler J, Lynam DA, Tuszynski MH, Sakamoto JS. Characterizing the degradation of alginate hydrogel for use in multilumen scaffolds for spinal cord repair. J Biomed Mater Res A. 2016;104:611–619. doi: 10.1002/jbm.a.35600. [DOI] [PubMed] [Google Scholar]
- Sharma HS, Ali S, Tian ZR, Patnaik R, Patnaik S, Lek P, Sharma A, Lundstedt T. Nano-drug delivery and neuroprotection in spinal cord injury. J Nanosci Nanotechnol. 2009;9:5014–5037. doi: 10.1166/jnn.2009.gr04. [DOI] [PubMed] [Google Scholar]
- Shi CY, Ruan LQ, Feng YH, Fang JL, Song CJ, Yuan ZG, Ding YM. Marrow mesenchymal stem cell transplantation with sodium alginate gel for repair of spinal cord injury in mice. Zhejiang Da Xue Xue Bao Yi Xue Ban. 2011;40:354–359. [PubMed] [Google Scholar]
- Song Z, Wang Z, Shen J, Xu S, Hu Z. Nerve growth factor delivery by ultrasound-mediated nanobubble destruction as a treatment for acute spinal cord injury in rats. Int J Nanomed. 2017;12:1717–1729. doi: 10.2147/IJN.S128848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun HH, Gao F, Liu B, Yu HT, Kong N, Liu GM. Inhibition of Nogo expression to promote repair after spinal cord injury. Chin Med J (Engl) 2012;125:4044–4048. [PubMed] [Google Scholar]
- Tachi Y, Okuda T, Kawahara N, Kato N, Ishigaki Y, Matsumoto T. Expression of hyaluronidase-4 in a rat spinal cord hemisection model. Asian Spine J. 2015;9:7–13. doi: 10.4184/asj.2015.9.1.7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tamosaityte S, Galli R, Uckermann O, Sitoci-Ficici KH, Later R, Beiermeister R, Doberenz F, Gelinsky M, Leipnitz E, Schackert G, Koch E, Sablinskas V, Steiner G, Kirsch M. Biochemical monitoring of spinal cord injury by FT-IR spectroscopy-effects of therapeutic alginate implant in rat models. PLoS One. 2015;10:e0142660. doi: 10.1371/journal.pone.0142660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsai EC, Dalton PD, Shoichet MS, Tator CH. Matrix inclusion within synthetic hydrogel guidance channels improves specific supraspinal and local axonal regeneration after complete spinal cord transection. Biomaterials. 2006;27:519–533. doi: 10.1016/j.biomaterials.2005.07.025. [DOI] [PubMed] [Google Scholar]
- Tyler JY, Xu XM, Cheng JX. Nanomedicine for treating spinal cord injury. Nanoscale. 2013;5:8821–8836. doi: 10.1039/c3nr00957b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Umezawa H, Naito Y, Tanaka K, Yoshioka K, Suzuki K, Sudo T, Hagihara M, Hatano M, Tatsumi K, Kasuya Y. Genetic and pharmacological inhibition of p38alpha improves locomotor recovery after spinal cord injury. Front Pharmacol. 2017;8:72. doi: 10.3389/fphar.2017.00072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- White-Schenk D, Shi R, Leary JF. Nanomedicine strategies for treatment of secondary spinal cord injury. Int J Nanomed. 2015;10:923–938. doi: 10.2147/IJN.S75686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xun MM, Liu YH, Guo Q, Zhang J, Zhang QF, Wu WX, Yu XQ. Low molecular weight PEI-appended polyesters as non-viral gene delivery vectors. Eur J Med Chem. 2014;78C:118–125. doi: 10.1016/j.ejmech.2014.03.050. [DOI] [PubMed] [Google Scholar]
- Zukor K, Belin S, Wang C, Keelan N, Wang X, He Z. Short hairpin RNA against PTEN enhances regenerative growth of corticospinal tract axons after spinal cord injury. J Neurosci. 2013;33:15350–15361. doi: 10.1523/JNEUROSCI.2510-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]