

Abstract
As a behavioural addiction, gambling disorder (GD) provides an opportunity to characterize addictive processes without the potentially confounding effects of chronic excessive drug and alcohol exposure. Impulsivity is an established precursor to such addictive behaviours, and GD is associated with greater impulsivity. There is also evidence of GABAergic dysregulation in substance addiction and in impulsivity. This study therefore investigated GABAA receptor availability in 15 individuals with GD and 19 healthy volunteers (HV) using [11C]Ro15‐4513, a relatively selective α5 benzodiazepine receptor PET tracer and its relationship with impulsivity. We found significantly higher [11C]Ro15‐4513 total distribution volume (VT) in the right hippocampus in the GD group compared with HV. We found higher levels of the ‘Negative Urgency’ construct of impulsivity in GD, and these were positively associated with higher [11C]Ro15‐4513 VT in the amygdala in the GD group; no such significant correlations were evident in the HV group. These results contrast with reduced binding of GABAergic PET ligands described previously in alcohol and opiate addiction and add to growing evidence for distinctions in the neuropharmacology between substance and behavioural addictions. These results provide the first characterization of GABAA receptors in GD with [11C]Ro15‐4513 PET and show greater α5 receptor availability and positive correlations with trait impulsivity. This GABAergic dysregulation is potential target for treatment.
Keywords: GABA system, gambling disorder, [11C]Ro15‐4513 PET
Introduction
There is considerable overlap between gambling disorder (GD) (previously termed pathological gambling) with drug and alcohol addiction with regard to clinical phenomenology and treatment, comorbidity, heritability and neurobiological profile (Clark 2014; Clark and Limbrick‐Oldfield 2013). Predicated on such evidence, this condition was recently reclassified from an ‘impulse control disorder’ in DSM‐IV to the ‘substance‐related and addictive disorders’ in DSM‐5 (Clark 2014). As a behavioural addiction, GD provides an opportunity to characterize addictive processes without the potentially confounding effects of chronic excessive drug/alcohol exposure. Compared with drug or alcohol addiction, little is known about the neuropharmacology of GD, and a better characterization of its neurobiology will inform developments in the prevention and treatment of both substance and behavioural addictions.
Recent studies using positron emission tomography (PET) to directly assess the dopamine and opioid systems in GD have demonstrated some unexpected differences to established findings in substance addictions. For instance, in contrast to studies in cocaine and alcohol addiction, [11C]raclopride and [11C]‐(+)‐PHNO PET revealed no differences in dopamine receptor DRD2/3 availability and greater stimulant‐induced dopamine release in individuals with GD compared with healthy volunteers (HV; Albein‐Urios et al. 2012a,2012b; Boileau et al. 2013; Boileau et al. 2014). A key modulator of the dopaminergic system is the mu opioid receptor (MOR), and opiate receptor antagonists have some efficacy in treating GD. Higher MOR availability has been reported in studies by using [11C]carfentanil or [11C]diprenorphine PET in cocaine, opiate and alcohol addiction (Gorelick et al. 2005; Heinz et al. 2005; Williams et al. 2007; Williams et al. 2009). However, we recently reported no difference in [11C]carfentanil binding in individuals with GD (Mick et al. 2015). Thus, in vivo PET imaging studies of neuropharmacology suggest that GD shows important differences to substance addiction.
The GABAergic system is another key modulator of mesolimbic dopaminergic processes but has received scant attention in the context of addictions (Hayes et al. 2014). With the [11C]Ro15‐4513 PET radiotracer, which is relatively selective for the α5 subtype of the benzodiazepine receptor, we observed in alcohol and in heroin addiction lower levels of limbic [11C]Ro15 4513 binding, particularly in the nucleus accumbens (NAc) and right hippocampus (Lingford‐Hughes et al. 2016; Lingford‐Hughes et al. 2012a). We have also shown that [11C]Ro15 4513 binding was higher in participants with a history of tobacco smoking (Stokes et al. 2013) .
With regard to gambling, Nussbaum et al. (2011) proposed that GABAergic modulation of opioid release in the brain reward system may be important to the ‘rush’ experienced by gamblers in response to major jackpots and that differences in this modulation could distinguish individuals with GD from non‐dependent gamblers. There is limited further evidence regarding the GABAergic system in individuals with GD, with some inconsistent evidence of greater GABA concentrations measured in the CSF in GD (Nordin and Sjodin 2007; Roy et al. 1989). These observations, as well as the growing use of GABAergic medication such as baclofen and topiramate to treat addiction, indicate that it is timely and important to characterize the GABAA receptor system in GD (Lingford‐Hughes et al. 2012b).
A major feature common to gambling and substance‐related disorders is impaired impulse control, which may be fundamental to both the development and maintenance of addictive disorders (Bechara 2005; Lawrence et al. 2009; Verdejo‐Garcia et al. 2008). We have previously shown that Negative Urgency (NU) derived from the UPPS‐P impulsivity scale is related to dopamine D2/3 receptor availability in GD (Clark et al. 2012) and also to MOR availability (Mick et al. 2015). As the main inhibitory neurotransmitter in the human brain, the role of GABAergic functioning in impulsivity is attracting attention with growing preclinical evidence supporting its involvement (Hayes et al. 2014). For instance, a GABA agonist and antagonist in the prefrontal cortex of rats increased and reduced, respectively, impulsive responses in the 5‐choice serial reaction time task (Murphy et al. 2012; Paine et al. 2011). GABAA receptor binding is lower in the anterior cingulate cortex of high‐impulsive rats compared with low‐impulsive rats and is inversely correlated with impulsive responding (Jupp et al. 2013). Caprioli et al. reported increased impulsive behaviour in rats following a reduction in glutamate decarboxylase 65/67, an enzyme responsible for the decarboxylation of glutamate to GABA, gene expression in the NAc core (Caprioli et al. 2014).
In this study, we therefore used [11C]Ro15‐4513 PET to measure GABAA receptor availability in GD and its relationship with impulsivity. Based on our previous studies showing lower [11C]Ro15‐4513 binding in the NAc in both alcohol and in opiate addiction, we hypothesized that GD would similarly be associated with lower levels in the NAc. In addition, based on the preclinical evidence, we hypothesized that impulsivity, specifically the NU trait, in GD would be associated with higher [11C]Ro15‐4513 binding; we have shown that [11C]Ro15‐4513 is sensitive to GABA and low levels of GABA are associated with higher [11C]Ro15‐4513 binding (Stokes et al. 2014).
Materials and Methods
Participants
Study participants comprised of 15 male treatment‐seeking individuals with GD (DSM‐IV) (34.8 ± 7.5 years, four current and two ex‐smokers (mean ± standard deviation (SD)) and 19 male age‐matched HV (31.7 ± 7.5 years, six current and three ex‐smokers). Individuals with GD were recruited from the National Problem Gambling Clinic, Central North West London NHS Foundation Trust, UK, and participated in the study either before (n = 7) or during (n = 8) their 8‐week programme of cognitive–behavioural therapy. HV were recruited by advertisements in daily newspapers or from our database. Written informed consent was obtained before participation in the study, which was approved by West London Research Ethics Committee and the Administration of Radioactive Substances Advisory Committee, UK.
Following a detailed telephone eligibility interview, participants attended a screening visit to assess their current and previous medical and psychiatric health. Disordered gambling was assessed with the Massachusetts Gambling Screen (6.7 ± 1.9; a score of 5 or more indicates GD) and the Problem Gambling Severity Index (16.9 ± 3.5; a score of 8 or above indicates disordered gambling). All individuals with GD had a recent history of active gambling with length of abstinence ranging from 3 to 158 days (63 ± 49 days). HV had no current or past history of psychiatric disorders (ICD‐10 or DSM‐IV Axis I diagnostic criteria). As depression and anxiety disorders are common comorbidities in GD, a history of these was permitted, but not a current depression or anxiety disorder. Past recreational drug use was allowed (>10 times in lifetime but never met criteria for abuse or dependence: seven HV: five cannabis, one cannabis and stimulants and one cannabis, hallucinogens and sedatives; four GD: two cannabis and two cannabis, stimulants and cocaine), but abstinence from illicit drugs 2 weeks before and during study participation was required. Current or past diagnosis of substance dependence, except nicotine, was an exclusion criterion for both groups. Participants were asked not to drink more than 21 UK units of alcohol (166 g) per week 2 weeks before and during the study. Urine drug screens (for cocaine, amphetamine, methamphetamine, morphine, methadone, benzodiazepines and tetrahydrocannabinol) and alcohol breath analyses were performed at screening and PET scanning days. Cigarette smoking was not allowed 1 hour prior to each magnetic resonance (MR) or PET scan. All participants had laboratory (haematology, clinical chemistry and clotting parameters) within normal range. None of the participants took regular medication.
Depressive symptoms were measured by using the Beck Depression Inventory and anxiety with the Spielberger State/Trait Inventory. Tobacco use was evaluated in the smokers with the Fagerstrom Test for Nicotine Dependence. To assess impulsivity, participants completed the UPPS‐P Impulsive Behavior Scale (Cyders et al. 2007), a 59 item self‐report questionnaire with five subscales: Negative Urgency (NU), Positive Urgency, Lack of Planning (LoP), Lack of Perseverance and Sensation Seeking.
MR and PET imaging
On the screening day, a structural magnetic resonance imaging (MRI) was performed on a 3 T MR scanner (Magnetom Trio Syngo MR B13 Siemens 3 T; Siemens AG, Medical Solutions), including a volumetric T1‐weighted magnetization‐prepared rapid acquisition gradient‐echo sequence. All structural images were reviewed by an experienced neuroradiologist for unexpected findings of clinical significance and structural variation that might affect quantitative image analysis, and none were observed. Participants also completed a functional MRI battery (Paterson et al. 2015) and performed a neurocognitive assessment whose results will be described elsewhere.
For the [11C]Ro15‐4513 PET scan, we followed our previous protocol (Lingford‐Hughes et al. 2012a). Briefly, each participant underwent a 90‐minute [11C]Ro15‐4513 PET acquisition. PET data were reconstructed into 23 frames (4 × 15, 4 × 60, 2 × 150, 10 × 300 and 3 × 600 seconds) and corrected for head motion by using normalized mutual information rigid‐body registration of each frame realignment to frame 16. A hierarchical probabilistic atlas of 119 regions (Tziortzi et al. 2014) was non‐rigidly registered to PET space by using reverse deformation parameters derived from the normalization of the coregistered structural MRI to a standard template. Atlas fits were checked visually, and PET dynamic data were sampled in each region of interest (ROI) to generate time‐activity curves, combining left and right ROIs to generate single time‐activity curves where laterality was not hypothesized, i.e. in amygdala and orbitofrontal cortex (OFC). A parent plasma input function was derived by calibrating the continuous online blood counts to discrete samples and correcting for plasma fraction and radiolabelled metabolites collected at intervals throughout the scan. The [11C]Ro15‐4513 VT in each ROI could then be quantified by using a 2‐tissue‐compartmental model. This was shown, by parsimony criteria, to describe appropriately the ligand kinetics in the range of ROIs considered (Myers et al. 2016a). In this dataset, spectral analysis to partition this signal was not applied since we have shown the total VT to represent robustly α5 subtype binding (Myers et al. 2016a; Myers et al. 2012). All image and kinetic analysis procedures were carried out by using MIAKATTM (Imanova Ltd, UK).
Statistical analysis
Demographic differences between groups, and injected mass/activity, were analysed by using independent‐samples t‐tests (two‐tailed). An omnibus mixed‐model ANCOVA tested [11C]Ro15‐4513 VT as a function of ROI (four levels) and Group (HV and GD) including age and smoking as covariates. Results are presented as mean + SD. Correlations between VT values and clinical variables were performed by using Spearman's rank correlation coefficients.
Based upon our previous findings in alcohol and opiate dependence for the [11C]Ro15‐4513 PET analysis, we selected right hippocampus and NAc as a priori ROIs, and we chose two additional a priori ROIs based upon their established role in impulse control: OFC and amygdala (Goldstein and Volkow 2002; Jentsch and Taylor 1999; Ko et al. 2015; Nikolova et al. 2016). As previous work (Clark et al. 2012; Michalczuk et al. 2011) has shown that NU, as a mood‐related subgroup of impulsivity, is most strongly associated with GD, we chose to test for correlations between UPPS‐P NU and a priori selected brain regions. Correlations were considered significant at p < 0.05, and for the group comparisons, if ROIs are to be treated as independent, multiple comparison correction adjusts the significance value to p < 0.0125.
Results
Participants' characteristics
Demographic, clinical, and impulsivity data are summarized in Table 1. There were no differences in age, intelligence quotient, smoking status or amount of alcohol consumed per week. Individuals with GD had greater levels of depression (Beck Depression Inventory: p = 0.013) and anxiety [Spielberger Trait Anxiety Inventory (STAI): p = 0.001; Spielberger State Anxiety Inventory (SSAI): p = 0.014] and Alcohol Use Disorders Identification Test (AUDIT) score (p = 0.037) than HV, although none of the participants reached a clinically significant threshold in either group.
Table 1.
Participants' characteristics, clinical and impulsivity measures, mean ± SD.
| HV n = 19 | GD n = 15 | Significance (two‐tailed) | |
|---|---|---|---|
| Age | 31.7 ± 7.5 | 34.8 ± 7.5 | 0.245 |
| IQ | 114.0 ± 13.5 | 116.5 ± 12.2 | 0.502 |
| CPGI | 0.2 ± 0.5 | 16.9 ± 3.5 | 0.001* |
| Gambling abstinence (days) | ‐ | 63.3 ± 50.0 | ‐ |
| AUDIT | 4.2 ± 2.8 | 6.5 ± 3.5 | 0.037* |
| Alcohol (units/ week) | 9.0 ± 8.0 | 12.5 ± 7.4 | 0.182 |
| Current smoking status (smoker/ex‐smoker) | 6/3 | 4/2 | 0.957 |
| FTND | 1.0 ± 1.8 | 2.4 ± 2.3 | 0.266 |
| Cigarettes (per day) | 6.0 ± 5.8 | 5.0 ± 7.1 | 0.793 |
| BDI | 1.3 ± 2.8 | 4.9 ± 5.0 | 0.013* |
| STAI | 31.0 ± 9.0 | 42.5 ± 9.9 | 0.001* |
| SSAI | 26.0 ± 5.1 | 36.3 ± 13.8 | 0.014* |
| UPPS‐P NU | 21.7 ± 5.8 | 30.7 ± 6.2 | 0.001* |
| UPPS‐P PU | 20.7 ± 6.9 | 25.0 ± 8.0 | 0.130 |
| UPPS‐P LoP | 20.2 ± 4.7 | 24.3 ± 5.6 | 0.028* |
| UPPS‐P LoPe | 18.2 ± 4.4 | 21.1 ± 4.8 | 0.076 |
| UPPS‐P SS | 34.3 ± 8.1 | 33.3 ± 7.7 | 0.722 |
GD, individuals with gambling disorder; HV, healthy volunteers; AUDIT, Alcohol Use Disorders Identification Test; BDI, Beck Depression Inventory; CPGI, Canadian Problem Gambling Inventory; FTND, Fagerstrom Test for Nicotine Dependence; GUQ, Gambling Urges Questionnaire; NART, National Adult Reading Test; SSAI, Spielberger State Anxiety Inventory; STAI, Spielberger Trait Anxiety Inventory.
Significant difference between groups.
PET measures
There were no significant differences (p > 0.05) between the groups for injected [11C]Ro15‐4513 mass HV: 2.98 ± 1.06 µg versus GD: 3.19 ± 1.24 µg nor for injected radioactivity HV: 352 ± 77 MBq versus GD 356 ± 90 MBq.
The omnibus ANCOVA including age and smoking (Fagerstrom Test for Nicotine Dependence) for the a priori regions revealed a significant main effect of Group (F 1,28 = 6.1, p = 0.020), with greater mean VT in individuals with GD compared with HV. This was driven by significant group differences in the [11C]Ro15‐4513 total distribution volume (VT) in the right hippocampus [t(32) = 2.7; p = 0.011], which survives Bonferroni correction for multiple comparisons (Table 2). We did not show any significant difference in the NAc, OFC or amygdala (p > 0.05). There was also a significant main effect of smoking (F 1,28 = 4.8, p = 0.037), such that Fagerstrom scores correlated negatively with mean VT across the four ROIs (rs = −0.36; p = 0.041). In both groups, there was no significant correlation between Alcohol Use Disorders Identification Test scores and [11C]Ro15‐4513 VT. There was also no significant correlation between ‘length of abstinence’ and [11C]Ro15‐4513 VT. There was no significant difference between those that had started and not started their cognitive–behavioural treatment (p > 0.05) in [11C]Ro15‐4513 VT in any of the a priori ROIs.
Table 2.
Comparison between groups of [11C]Ro15‐4513 VT in a priori defined brain regions, mean ± SD.
| HV n = 19 | GD n = 15 | ||||||
|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Significance (two‐tailed) | |||
| Orbitofrontal cortex | 6.99 | ± | 0.75 | 7.18 | ± | 0.87 | 0.563 |
| Amygdala | 7.44 | ± | 0.65 | 7.97 | ± | 0.87 | 0.058 |
| Hippocampus R | 7.44 | ± | 0.72 | 8.13 | ± | 0.74 | 0.011* |
| Nucleus accumbens | 9.76 | ± | 0.72 | 9.95 | ± | 1.04 | 0.544 |
GD, individuals with gambling disorder; HV, healthy volunteers; L, left hemisphere; R, right hemisphere. HV: Amygdala n = 18; GD: N. Accumbens n = 14.
Significant at p < 0.05.
Relationship between PET measures, impulsivity and anxiety
Individuals with GD showed significantly higher impulsivity scores in UPPS‐P NU (p < 0.001) and LoP subscales (p = 0.028) compared with HV (Table 1). Non‐parametric correlations were run between [11C]Ro15‐4513 VT and UPPS‐P NU and revealed in the GD group, significant positive correlations in the amygdala (rs = 0.67; p = 0.006), right hippocampus (rs = 0.53; p = 0.038) and NAc (rs = 0.57; p = 0.034; Table 3) (Fig. 1). There were no significant correlations in the HV group (p > 0.05). The differential relationship with NU in the two groups was confirmed statistically by using a generalized linear model, which showed a significant impulsivity × group interaction in the amygdala (p = 0.015). These interactions were non‐significant for right hippocampus and NAc (p > 0.05). There were no significant correlations between [11C]Ro15‐4513 VT in a priori ROIs and LoP in either of the groups.
Table 3.
Spearman's correlations between UPPS‐P NU and [11C]Ro15‐4513 VT in the GD and HV groups.
| HV n = 19 | GD n = 15 | |||
|---|---|---|---|---|
| Correlation coefficient | Significance (two‐tailed) | Correlation coefficient | Significance (two‐tailed) | |
| Orbitofrontal cortex | 0.08 | 0.731 | 0.51 | 0.052 |
| Amygdala | 0.00 | 0.990 | 0.67 | 0.006** |
| Hippocampus R | −0.10 | 0.680 | 0.53 | 0.043* |
| Nucleus accumbens | 0.46 | 0.050 | 0.57 | 0.034* |
HV, healthy volunteers; GD, individuals with gambling disorder; L, left hemisphere; R, right hemisphere. HV: Amygdala n = 18; GD: nucleus accumbens n = 14.
Significant at p < 0.05.
Significant at p < 0.01.
Figure 1.
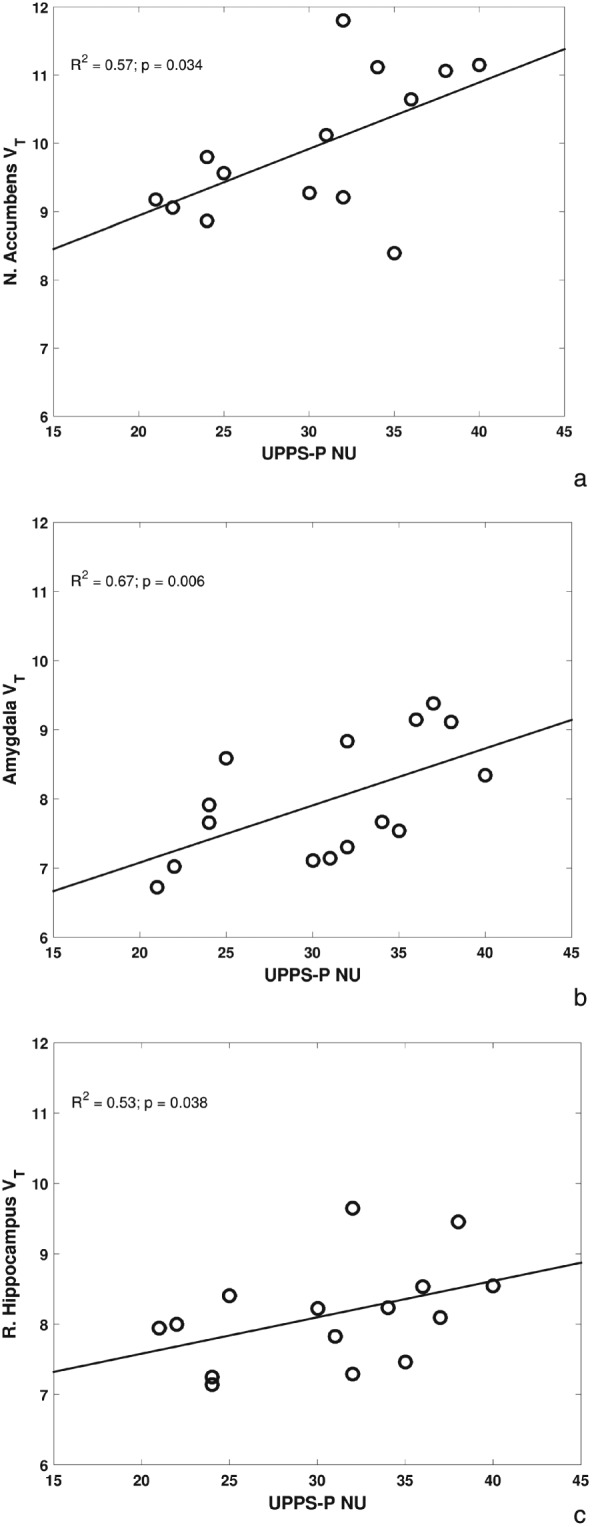
Significantly positive correlations between Negative Urgency impulsivity and [11C]Ro15‐4513 VT in individuals with gambling disorder
As anxiety scores were significantly different in HV and individuals with GD (STAI; p = 0.001 and SSAI; p = 0.014) and anxiety is related to UPPS‐P NU, we also explored correlations between trait anxiety (STAI), state anxiety (SSAI), NU and [11C]Ro15‐4513 VT in the 4 ROIs. There was a significant positive correlation between STAI and [11C]Ro15‐4513 VT in the amygdala (rs = 0.51; p = 0.05) in the GD group, which was the only one that survived Bonferroni correction. After controlling for anxiety, the association between UPPS‐P NU and the amygdala [11C]Ro15‐4513 VT remained significant (rs = 0.63; p = 0.016). There were no significant correlations with trait anxiety in the HV group, and state anxiety was not significantly correlated with [11C]Ro15‐4513 VT in either group.
Discussion
We report here the first study investigating GABAA receptor availability in GD and show significantly greater [11C]Ro15‐4513 VT in the right hippocampus of individuals with GD and a positive relationship between [11C]Ro15‐4513 VT and impulsivity. Notably, our finding of higher [11C]Ro15‐4513 VT is in contrast to our previous finding of lower levels of [11C]Ro15‐4513 binding in the NAc in alcohol and in opiate dependence and lower levels in the hippocampus in alcohol dependence (Lingford‐Hughes et al. 2016; Lingford‐Hughes et al. 2012a). Higher [11C]Ro15‐4513 binding reflects greater GABAA receptor availability, which may be due to increased receptor expression or, as a consequence of Ro15‐4513 being an inverse agonist, lower endogenous GABA levels (Stokes et al. 2014). We are unable to separate these two mechanisms within our study. Nevertheless, our [11C]Ro15‐4513 PET studies show clear differences in GABAA receptor availability between substance addictions and a behavioural addiction with lower levels of [11C]Ro15‐4513 binding in both alcohol and opiate addiction not apparent in GD (Lingford‐Hughes et al. 2016; Lingford‐Hughes et al. 2012a).
In our studies in substance dependence, significantly lower [11C]Ro15‐4513 VT was found in the hippocampus in alcohol dependence but not in opiate dependence. In these studies, we used spectral analysis (Myers et al. 2012) to show that these findings in alcohol dependence were driven by the α5 subtype rather than α1; this is consistent with the high level of α5 subtype in the hippocampus and selectivity of [11C]Ro15‐4513 PET (Lingford‐Hughes et al. 2016). In the current study in GD, [11C]Ro15‐4513 VT in each ROI was quantified by using a 2‐tissue‐compartmental model, which we have shown to be suitable to describe the α5‐subtype‐specific signal of [11C]Ro15‐4513 in most ROIs (Myers et al. 2016a) though the contribution of other subtypes in regions with low GABAA α5 density, such as the cerebellum, should also be considered.
We have previously reported higher [11C]Ro15‐4513 VT in various brain regions including the hippocampus, in HV smokers, which was particularly evident in ex‐smokers with levels in current smokers similar to non‐smokers (Stokes et al. 2013). However, these individuals were not heavy smokers, and in the current study, the amount of smoking was even lower so the majority were not dependent smokers. Given there were only a few current smokers, we are unable to explore further the impact of smoking on [11C]Ro15‐4513 VT binding in GD.
Consistent with previous work, we found higher impulsivity scores in individuals with GD compared with HV (Clark et al. 2012; Michalczuk et al. 2011). Preclinical studies suggest that reductions in or lower levels of GABA are associated with greater impulsivity (Boy et al. 2011; Murphy et al. 2012; Paine et al. 2011). We have previously shown that [11C]Ro15‐4513 is sensitive to GABA levels, and since it is an inverse benzodiazepine agonist, lower levels of GABA would be associated with higher [11C]Ro15‐4513 binding. Consistent with our hypothesis that there would be a positive relationship between impulsivity and [11C]Ro15‐4513 VT, we found such a relationship in subcortical limbic regions in GD but not in HV. The relationship between [11C]Ro15‐4513 VT and ‘NU’ construct of impulsivity adds further support to the idea that mood‐related impulsivity is especially relevant to disordered gambling (Clark et al. 2012; Nussbaum et al. 2011). It is notable that we have found NU to be associated with our PET studies of dopamine D2, MOR and now GABA‐A receptor in those with GD but not healthy controls suggesting that dysregulation in these systems has particular relevance to impulsivity in gambling (Clark et al. 2012; Mick et al. 2015). Given the association with NU, further exploration revealed a positive relationship between trait anxiety and [11C]Ro15‐4513 VT in the amygdala in the GD group only. Notably, the positive correlation with [11C]Ro15‐4513 VT and NU in the amygdala remained significant after controlling for anxiety. The amygdala is a key region involved in a number of processes such as emotional processing, fear, reward valence and neuropsychiatric disorders including addiction and anxiety (Elman et al. 2012; Janak and Tye 2015; van Holst et al. 2012). However, evidence from neuroimaging studies using non‐specific benzodiazepine radiotracers [11C]flumazenil PET or [123I]iomazenil SPET about GABAA receptor availability within the amygdala in relation to these processes is limited and inconsistent. The most evidence relates to anxiety where no relationship or negative correlations have been reported in HV and both positive and negative correlations in patients with anxiety disorders (Abadie et al. 1999; Esterlis et al. 2009; Hasler et al. 2008; Lingford‐Hughes et al. 1998).
Our finding of an association between impulsivity and GABAA receptors in vivo in individuals with GD is consistent with evidence from pharmacological challenges with GABA modulating drugs such as benzodiazepines, which suggest that increased GABAergic neurotransmission is associated with impulsivity (Hayes et al. 2014). Other studies in man have explored the relationship between GABA levels assessed with magnetic resonance spectroscopy. Whilst GABA measured this way is predominantly metabolic in origin (Myers et al. 2014; Myers et al. 2016b), it is interesting that a lowered magnetic resonance spectroscopy GABA signal ([GABA+]) in HV has in some studies been associated with higher levels of impulsivity. For instance, it has been shown that [GABA+] levels in dorsolateral prefrontal regions negatively correlate with urgency impulsivity (Boy et al. 2011); low [GABA+] in perigenual anterior cingulate cortex was associated with greater delay aversion on Cambridge gambling task (Fujihara et al. 2015) and higher scores on self‐report Barratt Impulsiveness Scale‐11 in one study (Silveri et al. 2013) but not another (Janes et al. 2013). Lower [GABA+] in the striatum is associated with poorer impulse control in Go‐NoGo task (Quetscher et al. 2015). Such relationships add to the notion that modulating GABA is a potential target for treatment of GD.
We acknowledge that the number of patients included in this study is small. Based on our previous [11C]Ro15‐4513 studies in alcohol and opiate dependent participants, we expected to find lower VT in the NAc. A post hoc power calculation revealed that we would need more than 150 additional [11C]Ro15‐4513 PET scans in order to be able to detect a statistically significant group difference in the NAc. The power calculation was performed by using the mean and SD data collected in our study, with alpha = 0.05, to show a difference with 80 percent power. This number is beyond what is ethically acceptable and feasible for a PET study, and we therefore did not continue scanning.
In summary, we provide here the first evidence of GABAA dysregulation in individuals with GD with increased [11C]Ro15‐4513 VT in the right hippocampus. This is different to substance dependence where we have shown reduced [11C]Ro15‐4513 VT in the NAc (Lingford‐Hughes et al. 2016; Lingford‐Hughes et al. 2012a). As described, GD was categorized in DSM‐IV as an impulse control disorder but based on shared clinical and aetiological features is now classified as a behavioural addiction in DSM‐5. The current study adds to our and other previous studies failing to show similar findings in dopamine D2/3 receptor availability or mu opiate receptor availability between GD and substance dependence (Clark et al. 2012; Mick et al. 2015). Thus, our data suggest that the view that, as a behavioural addiction, the neuropharmacology of GD would be similar to substance addiction is now less certain. However, these differences may reflect the impact of the chronic excessive drug/alcohol exposure on the brain as opposed to the underlying addictive processes themselves. On the other hand, our PET studies and those of others all report a relationship between impulsivity in GD with dopamine D2/3, with MOR and with GABAA receptor availability but not control groups. These associations suggest that pharmacological modulation may result in reduced impulsivity as an approach to treatment. Future studies are necessary to further investigate the neurobiology of this behavioural addiction in order to clearly address similarities and distinctions given the critical role for GABAergic function in addictive processes and its role in neural regulation. The evidence, albeit limited, and our data suggest that modulation of the GABA receptor system is a potential target for the treatment of GD.
Acknowledgements
This study was funded by the Medical Research Council—MRC G1002226. Anna Ramos has received financial support with a scholarship from CAPES (process number: PDSE 99999.014476/2013‐04).
The authors wish to thank the study participants and the clinical team at Imanova Ltd., Centre for Imaging Sciences. The research was supported by the National Institute for Health Research Imperial Biomedical Research Centre. The views expressed are those of the author(s) and not necessarily those of the NHS, the National Institute for Health Research or the Department of Health.
Authors Contribution
ARLH, DJN, LC, HBJ, EAR, RNG, DE, PARS and IM were responsible for the study concept and design. MAM, DE and IM contributed to the acquisition of data. JM performed the PET analysis. ARLH, DJN, LC, HBJ, ADW, JCFG, GES, EAR, SC, PRAS, JM, ACR and IM assisted with data analysis and interpretation of findings. ACR and IM drafted the manuscript. ARLH, DJN and LC provided critical revision of the manuscript for important intellectual content. All authors critically reviewed the content and approved the final version for publication.
Mick, I. , Ramos, A. C. , Myers, J. , Stokes, P. R. , Chandrasekera, S. , Erritzoe, D. , Mendez, M. A. , Gunn, R. N. , Rabiner, E. A. , Searle, G. E. , Galduróz, J. C. F. , Waldman, A. D. , Bowden‐Jones, H. , Clark, L. , Nutt, D. J. , and Lingford‐Hughes, A. R. (2017) Evidence for GABA‐A receptor dysregulation in gambling disorder: correlation with impulsivity. Addiction Biology, 22: 1601–1609. doi: 10.1111/adb.12457.
References
- Abadie P, Boulenger JP, Benali K, Barre L, Zarifian E, Baron JC (1999) Relationships between trait and state anxiety and the central benzodiazepine receptor: a PET study. Eur J Neurosci 11:1470–1478. [DOI] [PubMed] [Google Scholar]
- Albein‐Urios N, Martinez‐Gonzalez JM, Lozano O, Clark L, Verdejo‐Garcia A (2012a) Comparison of impulsivity and working memory in cocaine addiction and pathological gambling: implications for cocaine‐induced neurotoxicity. Drug Alcohol Depend 126:1–6. [DOI] [PubMed] [Google Scholar]
- Albein‐Urios N, Martinez‐Gonzalez JM, Lozano O, Clark L, Verdejo‐Garcia A (2012b) Comparison of impulsivity and working memory in cocaine addiction and pathological gambling: implications for cocaine‐induced neurotoxicity. Drug Alcohol Depend 126:1–6. [DOI] [PubMed] [Google Scholar]
- Bechara A (2005) Decision making, impulse control and loss of willpower to resist drugs: a neurocognitive perspective. Nat Neurosci 8:1458–1463. [DOI] [PubMed] [Google Scholar]
- Boileau I, Payer D, Chugani B, Lobo D, Behzadi A, Rusjan PM, Houle S, Wilson AA, Warsh J, Kish SJ, Zack M (2013) The D2/3 dopamine receptor in pathological gambling: a positron emission tomography study with [11C]‐(+)‐propyl‐hexahydro‐naphtho‐oxazin and [11C]raclopride. Addiction (Abingdon, England) 108:953–963. [DOI] [PubMed] [Google Scholar]
- Boileau I, Payer D, Chugani B, Lobo DS, Houle S, Wilson AA, Warsh J, Kish SJ, Zack M (2014) In vivo evidence for greater amphetamine‐induced dopamine release in pathological gambling: a positron emission tomography study with [C]‐(+)‐PHNO. Mol Psychiatry 19:1305–1313. [DOI] [PubMed] [Google Scholar]
- Boy F, Evans CJ, Edden RA, Lawrence AD, Singh KD, Husain M, Sumner P (2011) Dorsolateral prefrontal gamma‐aminobutyric acid in men predicts individual differences in rash impulsivity. Biol Psychiatry 70:866–872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caprioli D, Sawiak SJ, Merlo E, Theobald DE, Spoelder M, Jupp B, Voon V, Carpenter TA, Everitt BJ, Robbins TW, Dalley JW (2014) Gamma aminobutyric acidergic and neuronal structural markers in the nucleus accumbens core underlie trait‐like impulsive behavior. Biol Psychiatry 75:115–123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clark L (2014) Disordered gambling: the evolving concept of behavioral addiction. Ann N Y Acad Sci 1327:46–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clark L, Limbrick‐Oldfield EH (2013) Disordered gambling: a behavioral addiction. Curr Opin Neurobiol 23:655–659. [DOI] [PubMed] [Google Scholar]
- Clark L, Stokes PR, Wu K, Michalczuk R, Benecke A, Watson BJ, Egerton A, Piccini P, Nutt DJ, Bowden‐Jones H, Lingford‐Hughes AR (2012) Striatal dopamine D(2)/D(3) receptor binding in pathological gambling is correlated with mood‐related impulsivity. Neuroimage 63:40–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cyders MA, Smith GT, Spillane NS, Fischer S, Annus AM, Peterson C (2007) Integration of impulsivity and positive mood to predict risky behavior: development and validation of a measure of positive urgency. Psychol Assess 19:107–118. [DOI] [PubMed] [Google Scholar]
- Elman I, Becerra L, Tschibelu E, Yamamoto R, George E, Borsook D (2012) Yohimbine‐induced amygdala activation in pathological gamblers: a pilot study. PLoS One 7:e31118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Esterlis I, Cosgrove KP, Batis JC, Bois F, Kloczynski TA, Stiklus SM, Perry E, Tamagnan GD, Seibyl JP, Makuch R, Krishnan‐Sarin S, O'Malley S, Staley JK (2009) GABAA‐benzodiazepine receptor availability in smokers and nonsmokers: relationship to subsyndromal anxiety and depression. Synapse (New York, NY) 63:1089–1099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fujihara K, Narita K, Suzuki Y, Takei Y, Suda M, Tagawa M, Ujita K, Sakai Y, Narumoto J, Near J, Fukuda M (2015) Relationship of gamma‐aminobutyric acid and glutamate + glutamine concentrations in the perigenual anterior cingulate cortex with performance of Cambridge Gambling Task. Neuroimage 109:102–108. [DOI] [PubMed] [Google Scholar]
- Goldstein RZ, Volkow ND (2002) Drug addiction and its underlying neurobiological basis: neuroimaging evidence for the involvement of the frontal cortex. Am J Psychiatry 159:1642–1652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gorelick DA, Kim YK, Bencherif B, Boyd SJ, Nelson R, Copersino M, Endres CJ, Dannals RF, Frost JJ (2005) Imaging brain mu‐opioid receptors in abstinent cocaine users: time course and relation to cocaine craving. Biol Psychiatry 57:1573–1582. [DOI] [PubMed] [Google Scholar]
- Hasler G, Nugent AC, Carlson PJ, Carson RE, Geraci M, Drevets WC (2008) Altered cerebral gamma‐aminobutyric acid type A‐benzodiazepine receptor binding in panic disorder determined by [11C]flumazenil positron emission tomography. Arch Gen Psychiatry 65:1166–1175. [DOI] [PubMed] [Google Scholar]
- Hayes DJ, Jupp B, Sawiak SJ, Merlo E, Caprioli D, Dalley JW (2014) Brain gamma‐aminobutyric acid: a neglected role in impulsivity. Eur J Neurosci 39:1921–1932. [DOI] [PubMed] [Google Scholar]
- Heinz A, Reimold M, Wrase J, Hermann D, Croissant B, Mundle G, Dohmen BM, Braus DF, Schumann G, Machulla HJ, Bares R, Mann K (2005) Correlation of stable elevations in striatal mu‐opioid receptor availability in detoxified alcoholic patients with alcohol craving: a positron emission tomography study using carbon 11‐labeled carfentanil. Arch Gen Psychiatry 62:57–64. [DOI] [PubMed] [Google Scholar]
- Janak PH, Tye KM (2015) From circuits to behaviour in the amygdala. Nature 517:284–292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janes AC, Jensen JE, Farmer SL, Frederick BD, Pizzagalli DA, Lukas SE (2013) GABA levels in the dorsal anterior cingulate cortex associated with difficulty ignoring smoking‐related cues in tobacco‐dependent volunteers. Neuropsychopharmacology : Official Publication of the American College of Neuropsychopharmacology 38:1113–1120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jentsch JD, Taylor JR (1999) Impulsivity resulting from frontostriatal dysfunction in drug abuse: implications for the control of behavior by reward‐related stimuli. Psychopharmacology (Berl) 146:373–390. [DOI] [PubMed] [Google Scholar]
- Jupp B, Caprioli D, Saigal N, Reverte I, Shrestha S, Cumming P, Everitt BJ, Robbins TW, Dalley JW (2013) Dopaminergic and GABA‐ergic markers of impulsivity in rats: evidence for anatomical localisation in ventral striatum and prefrontal cortex. Eur J Neurosci 37:1519–1528. [DOI] [PubMed] [Google Scholar]
- Ko CH, Hsieh TJ, Wang PW, Lin WC, Yen CF, Chen CS, Yen JY (2015) Altered gray matter density and disrupted functional connectivity of the amygdala in adults with Internet gaming disorder. Prog Neuropsychopharmacol Biol Psychiatry 57:185–192. [DOI] [PubMed] [Google Scholar]
- Lawrence AJ, Luty J, Bogdan NA, Sahakian BJ, Clark L (2009) Impulsivity and response inhibition in alcohol dependence and problem gambling. Psychopharmacology (Berl) 207:163–172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lingford‐Hughes A, Myers J, Watson B, Reid AG, Kalk N, Feeney A, Hammers A, Riano‐Barros DA, McGinnity CJ, Taylor LG, Rosso L, Brooks DJ, Turkheimer F, Nutt DJ (2016) Using [C]Ro15 4513 PET to characterise GABA‐benzodiazepine receptors in opiate addiction: similarities and differences with alcoholism. Neuroimage 132:1–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lingford‐Hughes A, Reid AG, Myers J, Feeney A, Hammers A, Taylor LG, Rosso L, Turkheimer F, Brooks DJ, Grasby P, Nutt DJ (2012a) A [11C]Ro15 4513 PET study suggests that alcohol dependence in man is associated with reduced alpha5 benzodiazepine receptors in limbic regions. J Psychopharmacol(Oxford, England) 26:273–281. [DOI] [PubMed] [Google Scholar]
- Lingford‐Hughes AR, Acton PD, Gacinovic S, Suckling J, Busatto GF, Boddington SJ, Bullmore E, Woodruff PW, Costa DC, Pilowsky LS, Ell PJ, Marshall EJ, Kerwin RW (1998) Reduced levels of GABA‐benzodiazepine receptor in alcohol dependency in the absence of grey matter atrophy. Br J Psychiatry 173:116–122. [DOI] [PubMed] [Google Scholar]
- Lingford‐Hughes AR, Welch S, Peters L, Nutt DJ (2012b) BAP updated guidelines: evidence‐based guidelines for the pharmacological management of substance abuse, harmful use, addiction and comorbidity: recommendations from BAP. J Psychopharmacol(Oxford, England) 26:899–952. [DOI] [PubMed] [Google Scholar]
- Michalczuk R, Bowden‐Jones H, Verdejo‐Garcia A, Clark L (2011) Impulsivity and cognitive distortions in pathological gamblers attending the UK National Problem Gambling Clinic: a preliminary report. Psychol Med 1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mick I, Myers J, Ramos AC, Stokes PR, Erritzoe D, Colasanti A, Gunn RN, Rabiner EA, Searle GE, Waldman AD, Parkin MC, Brailsford AD, Galduroz JC, Bowden‐Jones H, Clark L, Nutt DJ, Lingford‐Hughes AR (2015) Blunted endogenous opioid release following an oral amphetamine challenge in pathological gamblers. Neuropsychopharmacology: official publication of the American College of Neuropsychopharmacology . [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murphy ER, Fernando AB, Urcelay GP, Robinson ES, Mar AC, Theobald DE, Dalley JW, Robbins TW (2012) Impulsive behaviour induced by both NMDA receptor antagonism and GABAA receptor activation in rat ventromedial prefrontal cortex. Psychopharmacology (Berl) 219:401–410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Myers JF, Evans CJ, Kalk NJ, Edden RA, Lingford‐Hughes AR (2014) Measurement of GABA using J‐difference edited 1H‐MRS following modulation of synaptic GABA concentration with tiagabine. Synapse 68:355–362. [DOI] [PubMed] [Google Scholar]
- Myers JFM, Comley RA, Gunn RN (2016a) Quantification of [11C]Ro15–4513 GABAAα5 specific binding and regional selectivity in humans. J Cereb Blood Flow Metab. [Epub ahead of print] DOI: 10.1177/0271678X16661339 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Myers JF, Nutt DJ, Lingford‐Hughes AR (2016b) Gamma‐aminobutyric acid as a metabolite: interpreting magnetic resonance spectroscopy experiments. J Psychopharmacol (Oxford, England) 30:422–427. [DOI] [PubMed] [Google Scholar]
- Myers JF, Rosso L, Watson BJ, Wilson SJ, Kalk NJ, Clementi N, Brooks DJ, Nutt DJ, Turkheimer FE, Lingford‐Hughes AR (2012) Characterisation of the contribution of the GABA‐benzodiazepine alpha1 receptor subtype to [(11)C]Ro15‐4513 PET images. J Cereb Blood Flow Metab 32:731–744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nikolova YS, Knodt AR, Radtke SR, Hariri AR (2016) Divergent responses of the amygdala and ventral striatum predict stress‐related problem drinking in young adults: possible differential markers of affective and impulsive pathways of risk for alcohol use disorder. Mol Psychiatry 21:348–356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nordin C, Sjodin I (2007) CSF cholecystokinin, gamma‐aminobutyric acid and neuropeptide Y in pathological gamblers and healthy controls. J Neural Transm(Vienna, Austria : 1996) 114:499–503. [DOI] [PubMed] [Google Scholar]
- Nussbaum D, Honarmand K, Govoni R, Kalahani‐Bargis M, Bass S, Ni X, Laforge K, Burden A, Romero K, Basarke S, Courbasson C, Deamond W (2011) An eight component decision‐making model for problem gambling: a systems approach to stimulate integrative research. J Gambl Stud 27:523–563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paine TA, Slipp LE, Carlezon WA Jr (2011) Schizophrenia‐like attentional deficits following blockade of prefrontal cortex GABAA receptors. Neuropsychopharmacology 36:1703–1713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paterson LM, Flechais RS, Murphy A, Reed LJ, Abbott S, Boyapati V, Elliott R, Erritzoe D, Ersche KD, Faluyi Y, Faravelli L, Fernandez‐Egea E, Kalk NJ, Kuchibatla SS, McGonigle J, Metastasio A, Mick I, Nestor L, Orban C, Passetti F, Rabiner EA, Smith DG, Suckling J, Tait R, Taylor EM, Waldman AD, Robbins TW, Deakin JW, Nutt DJ, Lingford‐Hughes AR (2015) The Imperial College Cambridge Manchester (ICCAM) platform study: an experimental medicine platform for evaluating new drugs for relapse prevention in addiction. Part A: study description. J Psychopharmacol (Oxford, England) 29:943–960. [DOI] [PubMed] [Google Scholar]
- Quetscher C, Yildiz A, Dharmadhikari S, Glaubitz B, Schmidt‐Wilcke T, Dydak U, Beste C (2015) Striatal GABA‐MRS predicts response inhibition performance and its cortical electrophysiological correlates. Brain Struct Funct 220:3555–3564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roy A, DeJong J, Ferraro T, Adinoff B, Gold P, Rubinow D, Linnoila M (1989) CSF GABA and neuropeptides in pathological gamblers and normal controls. Psychiatry Res 30:137–144. [DOI] [PubMed] [Google Scholar]
- Silveri MM, Sneider JT, Crowley DJ, Covell MJ, Acharya D, Rosso IM, Jensen JE (2013) Frontal lobe gamma‐aminobutyric acid levels during adolescence: associations with impulsivity and response inhibition. Biol Psychiatry 74:296–304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stokes PR, Benecke A, Myers J, Erritzoe D, Watson BJ, Kalk N, Barros DR, Hammers A, Nutt DJ, Lingford‐Hughes AR (2013) History of cigarette smoking is associated with higher limbic GABAA receptor availability. Neuroimage 69:70–77. [DOI] [PubMed] [Google Scholar]
- Stokes PR, Myers JF, Kalk NJ, Watson BJ, Erritzoe D, Wilson SJ, Cunningham VJ, Riano Barros D, Hammers A, Turkheimer FE, Nutt DJ, Lingford‐Hughes AR (2014) Acute increases in synaptic GABA detectable in the living human brain: a [(1)(1)C]Ro15‐4513 PET study. Neuroimage 99:158–165. [DOI] [PubMed] [Google Scholar]
- Tziortzi AC, Haber SN, Searle GE, Tsoumpas C, Long CJ, Shotbolt P, Douaud G, Jbabdi S, Behrens TE, Rabiner EA, Jenkinson M, Gunn RN (2014) Connectivity‐based functional analysis of dopamine release in the striatum using diffusion‐weighted MRI and positron emission tomography. Cereb Cortex (New York, NY : 1991) 24:1165–1177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Holst RJ, Veltman DJ, Buchel C, van den Brink W, Goudriaan AE (2012) Distorted expectancy coding in problem gambling: is the addictive in the anticipation? Biol Psychiatry 71:741–748. [DOI] [PubMed] [Google Scholar]
- Verdejo‐Garcia A, Lawrence AJ, Clark L (2008) Impulsivity as a vulnerability marker for substance‐use disorders: review of findings from high‐risk research, problem gamblers and genetic association studies. Neurosci Biobehav Rev 32:777–810. [DOI] [PubMed] [Google Scholar]
- Williams TM, Daglish MR, Lingford‐Hughes A, Taylor LG, Hammers A, Brooks DJ, Grasby P, Myles JS, Nutt DJ (2007) Brain opioid receptor binding in early abstinence from opioid dependence: positron emission tomography study. Br J Psychiatry 191:63–69. [DOI] [PubMed] [Google Scholar]
- Williams TM, Davies SJ, Taylor LG, Daglish MR, Hammers A, Brooks DJ, Nutt DJ, Lingford‐Hughes A (2009) Brain opioid receptor binding in early abstinence from alcohol dependence and relationship to craving: an [11C]diprenorphine PET study. Eur Neuropsychopharmacol 19:740–748. [DOI] [PubMed] [Google Scholar]