

Abstract
Lipids and lipid metabolites play important roles in plant-microbe interactions. Despite the extensive studies of lipases in lipid homeostasis and seed oil biosynthesis, the involvement of lipases in plant immunity remains largely unknown. In particular, GDSL esterases/lipases, characterized by the conserved GDSL motif, are a subfamily of lipolytic enzymes with broad substrate specificity. Here, we functionally identified two GDSL lipases, OsGLIP1 and OsGLIP2, in rice immune responses. Expression of OsGLIP1 and OsGLIP2 was suppressed by pathogen infection and salicylic acid (SA) treatment. OsGLIP1 was mainly expressed in leaf and leaf sheath, while OsGLIP2 showed high expression in elongating internodes. Biochemical assay demonstrated that OsGLIP1 and OsGLIP2 are functional lipases that could hydrolyze lipid substrates. Simultaneous down-regulation of OsGLIP1 and OsGLIP2 increased plant resistance to both bacterial and fungal pathogens, whereas disease resistance in OsGLIP1 and OsGLIP2 overexpression plants was significantly compromised, suggesting that both genes act as negative regulators of disease resistance. OsGLIP1 and OsGLIP2 proteins mainly localize to lipid droplets and the endoplasmic reticulum (ER) membrane. The proper cellular localization of OsGLIP proteins is indispensable for their functions in immunity. Comprehensive lipid profiling analysis indicated that the alteration of OsGLIP gene expression was associated with substantial changes of the levels of lipid species including monogalactosyldiacylglycerol (MGDG) and digalactosyldiacylglycerol (DGDG). We show that MGDG and DGDG feeding could attenuate disease resistance. Taken together, our study indicates that OsGLIP1 and OsGLIP2 negatively regulate rice defense by modulating lipid metabolism, thus providing new insights into the function of lipids in plant immunity.
Author summary
Lipases are a large family of enzymes conferring lipid metabolism. Lipids and their metabolites play diverse roles in plant growth as well as response to environmental stimuli. Accumulating evidence implicates lipids as signaling molecules mediating plant immunity. Therefore, lipases are presumed to be actively involved in plant defense responses. Based on gene expression profiling, we have identified two functional GDSL lipases, encoded by OsGLIP1 and OsGLIP2, whose expression was suppressed by pathogen infection in the model cereal rice. Both OsGLIP1 and OsGLIP2 proteins localize to lipid droplets and the endoplasmic reticulum (ER) membrane, and they likely coordinate lipid metabolism with differential but complementary expression patterns in tissues and developmental stages. Consequently, alteration of OsGLIP gene expression was associated with substantial changes of lipid abundance and plant disease resistance. Our work identifies and characterizes two lipases that function as negative regulators of plant immune responses, strengthening the understanding of lipid metabolism in plant-microbe interactions.
Introduction
Plants are continuously challenged by invading microorganisms, some of which are pathogens that threaten plant survival and cause a big loss of production in crops. During evolution, multiple layers of defense systems have been adopted by plant hosts in the fight against pathogen attack, such as physical barriers including cell wall and epidermal cuticles [1]. More importantly, plants have developed two elaborate immune systems, pathogen-associated molecular pattern (PAMP)-triggered immunity (PTI) and effector-triggered immunity (ETI). Perception of PAMPs conserved among different microorganisms by plant pattern recognition receptors (PRRs) triggers PTI, which constitutes the basal layer of plant defense. ETI is specifically induced when pathogen secreted effectors are recognized by host resistance (R) proteins, often leading to hypersensitive response (HR), which is a more robust defense response usually associated with reactive oxygen species (ROS) burst and programmed cell death [2].
Accumulating evidence has suggested lipids as important regulators of plant defense [3,4]. Cutin, a polymer matrix composed of short-chain fatty acids (FA), and wax, a mixture of very-long-chain fatty acids, form a physical barrier preventing pathogen infection [5]. Lipids and their derivatives have also been implicated as signaling molecules that could modulate plant immunity [6]. For instance, the well-known plant defense hormone, jasmonic acid (JA), is a lipid-derived molecule [7]. Oleic acid (depicted as 18:1), an endogenous unsaturated fatty acid, has been shown to suppress defense response [8,9]. Recent studies have also emphasized the important roles of lipids in the induction of systemic acquired resistance (SAR), a defense mechanism occurring in the systemic organs to protect plants against subsequent attack [10,11]. The identification of DIR1 (DEFECTIVE IN INDUCED RESISTANCE 1) as a potential lipid transfer protein provided strong evidence that a lipid-derived molecule was involved in long distance defense signaling [12]. Further studies characterized azelaic acid (AzA), a C9 dicarboxylic acid derived from C18 FAs and glycerol-3-phosphate (G3P, the precursor for all plant glycerolipids), as critical inducers of plant SAR [13,14]. G3P and AzA cooperate with DIR1 and another predicted lipid transfer protein AZELAIC ACID INDUCED 1 (AZI1) to form a feedback regulatory loop that regulates systemic immunity in plants [15]. Further investigation of DIR1 and AZI1 lipid transfer activities and their target lipid molecules would provide invaluable insights into the roles of lipids in plant immunity.
Lipid metabolism is mainly catalyzed by lipases. GDSL lipases are a newly discovered subclass of lipolytic enzymes characterized by a GDSL motif and are further classified as SGNH hydrolase due to the highly conserved residues Ser-Gly-Asn-His [16]. Despite the extensive studies in bacteria, relatively little is known about plant GDSL lipases although they have been identified in various plant species [3]. Interestingly, the Arabidopsis GDSL lipase AtGLIP1 has been shown to possess anti-microbial activity and modulates resistance to Alternaria brassicicola in association with ethylene signaling [17,18], while its closest homolog AtGLIP2 plays a role in defense by suppressing auxin response [19]. In hot pepper, CaGLIP1 is involved in pathogen and wound defense [20]. Moreover, TcGLIP from Tanacetum cinerariifolium confers acyltransferase activity responsible for the biosynthesis of natural insecticide pyrethrin [21].
Rice (Oryza sativa L.) serves as the major food crop supporting nearly half of the world population. The production and quality of rice are severely challenged by a variety of pathogens. Bacterial blight caused by Xanthomonas oryzae pv. oryzae (Xoo) and rice blast caused by Magnaporthe oryzae (M. oryzae) are major diseases that affect rice production and quality. Bioinformatics analysis identified a large family of GDSL lipases (~114 members) in the rice genome, but little is known about their functions [22,23]. Here we report the identification and functional characterization of two GDSL lipases, OsGLIP1 and OsGLIP2, and we show that the two lipases are involved in rice disease resistance. With comprehensive genetic, biochemical, and lipid profiling assays, our work indicated that OsGLIP1 and OsGLIP2 are functional lipases involved in lipid metabolism and negatively modulate rice immune responses.
Results
Characterization of two functional lipases OsGLIP1 and OsGLIP2 in rice
In order to identify new components involved in rice disease resistance, we had previously performed whole genome transcriptome analysis of rice genes in responses to Xoo and M. oryzae infection [24]. We identified a GDSL lipase gene, Os06g0129600 (OsGLIP1), the expression of which was suppressed after Xoo inoculation (fold change = 3.3). The down-regulation of OsGLIP1 by Xoo was confirmed by qRT-PCR, which was down-regulated and decreased to the lowest level at 24h post-inoculation (hpi) (Fig 1A). The suppression of gene expression following pathogen infection was also observed for the closest homolog of OsGLIP1, Os06g0156700, which we designated as OsGLIP2 (Fig 1A). Since SA is the major defense signaling molecule, we further examined the expression of OsGLIP1 and OsGLIP2 after SA treatment. The amount of OsGLIP1 and OsGLIP2 transcripts started to decrease as soon as 3h after SA treatment and reached to the lowest level in 6h to 9h, causing a faster expression inhibition in comparison with pathogen inoculation (Fig 1B). Similarly, treatment with BTH, a functional analog of SA, resulted in similar inhibition of OsGLIP1 and OsGLIP2 expression (Fig 1C). The results suggest that OsGLIP1 and OsGLIP2 might function in rice immune responses.
Fig 1. Expression of OsGLIP1 and OsGLIP2 was suppressed in responses to pathogen infection and chemical treatments.

(A) Down-regulation of OsGLIP1 and OsGLIP2 expression in eight-week-old plants infected with Xoo (strain PXO99A) in a time course of 48 hours. (B, C) Down-regulation of OsGLIP1 and OsGLIP2 expression in two-week-old seedlings sprayed with 1 mM SA (B) or 300 μM BTH (C). All treatments were repeated for three times with similar results (A-C). The rice Actin1 gene was used as an internal control. Data are shown as means ± SD from three biological replicates (Student’s t-test, *P < 0.05, **P < 0.01).
The highly conserved amino acid sequences of GDSL lipases among different organisms indicate that they might exhibit similar catalytic activities [25]. Phylogenetic analysis of rice GDSL lipases revealed that OsGLIP1 and OsGLIP2 group into the same clade, with 73% identity in amino acid sequences (S1 Fig). Further comparison of the OsGLIP1 and OsGLIP2 protein sequences with orthologues from other plant species revealed a high degree of similarity, especially for the GDSL/V motif in the N-terminal and the four invariant key catalytic residues Ser, Gly, Asn and His in the functional blocks (Fig 2A). To test the lipase activity of OsGLIP1 and OsGLIP2, we attempted to express GST-tagged OsGLIP1 and OsGLIP2 recombinant proteins in E. coli. However, we failed to express the full-length fusion proteins probably due to the signal peptides as predicted by SignalP (http://www.cbs.dtu.dk/services/SignalP) [26]. Instead, truncated OsGLIP proteins without the signal peptides (OsGLIP1Δ29 and OsGLIP2Δ35) could be produced successfully (Fig 2B). The recombinant proteins were then purified and incubated with p-nitrophenyl acetate and p-nitrophenyl butyrate, two synthetic substrates generally used for lipase activity assay [17,19]. Compared to the empty control and GST alone, both OsGLIP1Δ29 and OsGLIP2Δ35 hydrolyzed the lipid substrates with high efficiency (Fig 2C and 2D), demonstrating that OsGLIP1 and OsGLIP2 are functional lipases.
Fig 2. OsGLIP1 and OsGLIP2 exhibit lipase activities.

(A) Alignment of OsGLIP1 and OsGLIP2 amino acid sequences with functionally known homologues from Arabidopsis and Tanacetum cinerariifolium. Sequences were aligned using Genedoc. (B) Expression and purification of recombinant OsGLIP1-GST and OsGLIP2-GST proteins in E. coli. (C, D) Lipase activities of OsGLIP1 and OsGLIP2. OsGLIP1 and OsGLIP2 were incubated with p-nitrophenyl acetate (C) and p-nitrophenyl butyrate (D) at 30°C. The absorbance readings were collected every 5 minutes in a time course of 60 min or 120 min. The substrates were incubated with either GST or no protein as controls. Data are shown as means ± SD (n = 3).
OsGLIP1 and OsGLIP2 modulate plant lipid homeostasis
To further investigate the roles of OsGLIP1 and OsGLIP2 in lipid metabolism in plants, we performed lipid profiling assay in transgenic plants with altered OsGLIP1 and OsGLIP2 expression. The maize Ubiquitin1 promoter (Ubi1) was used to drive OsGLIP1 and OsGLIP2 overexpression in wild-type plants. For reduced expression, a conserved fragment from the OsGLIP1 and OsGLIP2 coding regions was selected to generate an RNA interference (RNAi) construct that targets both genes simultaneously. qRT-PCR analysis revealed that the transcript levels of both genes was increased or decreased in independent transgenic lines (S2A and S2B Fig).
Lipids were extracted from the leaves of grouped OsGLIP1 and OsGLIP2 overexpression and silencing plants and analysed by reverse phase high-performance liquid chromatography/electrospray ionization tandem mass spectrometry (RP-HPLC/ESI/MS/MS). As shown in Fig 3A, although no significant difference was detected for the amount of phospholipids PC (phosphatidylcholine), PE (phosphatidylethanolamine), PG (phosphatidylglycerol) and PI (phosphatidylinositol) in the RNAi and overexpression transgenic plants, the abundance of other lipids were substantially changed. For instance, total TAG was significantly increased in OsGLIP1/2-RNAi and decreased in OsGLIP1-OE plants. By contrast, total MGDG levels increased in OsGLIP1 overexpression plants. Enhanced expression of OsGLIP1 also led to reduction in DAG levels while OsGLIP2 overexpression decreased PA levels. We further compared the individual lipid molecular species (Fig 3B–3F and S3 Fig), showing that TAG molecules such as TAG50, TAG52 and TAG54 containing polyunsaturated FAs either markedly increased in OsGLIP1/2-RNAi plants or decreased in OsGLIP1-OE plants, respectively (Fig 3B). While in OsGLIP2-OE plants, many PA species decreased significantly (Fig 3C), and some TAG species accumulated (Fig 3B). We also observed significant increase in levels of MGDG and DGDG molecules in OsGLIP1-OE and OsGLIP2-OE plants (Fig 3D and 3F). These results, taken together, suggest that OsGLIP1 and OsGLIP2 function in lipid metabolism in planta and these two lipases exhibit overlapping as well as distinct properties in lipid metabolism.
Fig 3. Lipidomic profiling of OsGLIP1/2-RNAi and overexpression plants.

Leaves from six individual plants (eight-week-old) were mixed as one sample from three representative transgenic lines of each transgene were used to normalize samples. Five leaf samples each genetic background were statistically analysed. (A) Total lipid composition in leaves of eight-week-old plants. (B-F) Abundance of individual lipid species, TAG (B), PA (C), MGDG (D), DAG (E) and DGDG (F). The lipid structures are presented as the number of carbon atoms: total double bonds in the fatty acyl groups. Data are shown as means ± SD (n = 5) of mixed leaf samples from three representative transgenic lines. *P < 0.05 or **P < 0.01, by Student’s t-test and Bonferroni correction for multiple (three comparisons) tests.
OsGLIP1 and OsGLIP2 affect rice resistance to bacterial pathogen
Due to the important roles of lipids in plant immunity [3,4,6], we postulated that OsGLIP1 and OsGLIP2 might respond to pathogen infection and modulate lipid homeostasis to regulate rice immune responses. To explore the roles of OsGLIP1 and OsGLIP2 in rice immunity, OsGLIP1 and OsGLIP2 overexpression and RNAi plants were infected with bacterial blight pathogen Xoo (Fig 4A–4C). Compared to the wild type, the lesion was significantly longer in OsGLIP1-OE and OsGLIP2-OE but shorter in OsGLIP1/2-RNAi plants (Fig 4D–4F). We then chose representative lines from OsGLIP1-OE, OsGLIP2-OE and OsGLIP1/2-RNAi transgenic plants to further analyze the bacterial lesion development and growth in leaves after infection. The results showed that disease lesions expanded more quickly and bacteria grew faster in both OsGLIP1 and OsGLIP2 overexpression plants; while reduction of OsGLIP1 and OsGLIP2 expression significantly inhibited Xoo growth in comparison with the wild-type control, indicating marked increase of resistance against bacterial blight (Fig 4G and 4H). Taken together, pathogen inoculation assays indicated that OsGLIP1 and OsGLIP2 play a negative role in rice defense.
Fig 4. Disease resistance to bacterial blight in OsGLIP1-OE, OsGLIP2-OE and OsGLIP1/2-RNAi plants.

(A-F) Lesions and statistical analysis of lesion lengths of representative OsGLIP1-OE (A and D), OsGLIP2-OE (B and E) and OsGLIP1/2-RNAi (C and F) lines (eight-week-old) inoculated with bacterial pathogen Xoo at 14 dpi, with the wild type (TP309, WT) as control. Arrows indicate the bottoms of lesions. Data are shown as means ± SD (n > 10). Asterisks indicate significant difference in comparison with the wild-type control (Student’s t-test, *P < 0.05; ** P < 0.01). (G) Disease development during 12 days of inoculation in the representative lines of OsGLIP1-OE, OsGLIP2-OE and OsGLIP1/2-RNAi, compared with the wild type. Data are shown as means ± SD (n > 10). Asterisks indicate significant difference in comparison with the wild-type control (Student’s t-test, ** P < 0.01). (H) Bacterial growth during 12 days of inoculation in the representative lines of OsGLIP1-OE, OsGLIP2-OE and OsGLIP1/2-RNAi, compared with the wild type. Data are shown as means ± SD (n = 3). Asterisks indicate significant difference in comparison with the wild-type control (Student’s t-test, *P < 0.05; ** P < 0.01).
Activation of plant defense responses after pathogen infection is accompanied by up-regulation of pathogenesis-related (PR) genes [27]. We monitored the expression of PR genes in response to pathogen attack in the OsGLIP1 and OsGLIP2 transgenic lines. The leaves of transgenic as well as wild-type plants were inoculated with Xoo and samples were collected at different time points. We observed that the expression of PR1a, PR1b, PR5 and PR10 were usually induced after Xoo inoculation in the wild-type plants. Consistent with the enhanced disease resistance, PR genes showed much higher induction in OsGLIP1/2-RNAi plants, whereas their induction was attenuated in the OsGLIP1-OE and OsGLIP2-OE overexpression plants (Fig 5A–5D). It has been shown that PR1a and PR1b are the marker genes indicative of SA signaling activation while PR5 and PR10 are involved in both SA and JA signaling [28,29], the altered expression of the PR genes implied that the OsGLIP genes might affect rice immunity through indirectly modifying the defense hormone signaling pathways.
Fig 5. Altered expression of pathogenesis-related (PR) genes in OsGLIP1 transgenic plants.

Eight-week-old transgenic and wild-type plants were inoculated with Xoo (strain PXO99A). The induction of the PR genes, PR1a (A), PR1b (B), PR5 (C) and PR10 (D), in response to pathogen infection was compromised in OsGLIP1-OE plants, while silencing of both OsGLIP1 and OsGLIP2 significantly promoted the induction of the PR genes. Data shown are means ± SD from three biological replicates. Asterisks indicate significant difference in comparison with the wild-type control (Student’s t-test, *P < 0.05, **P < 0.01).
OsGLIP1 and OsGLIP2 affect rice resistance to fungal pathogen M. oryzae
We further investigated the roles of OsGLIP1 and OsGLIP2 in disease resistance against fungal blast (M. oryzae). OsGLIP1 and OsGLIP2 transgenic plants (three independent transgenic lines for each transgene) were inoculated with M. oryzae. Compared to the wild type, rice blast infection was strongly inhibited in OsGLIP1/2-RNAi plants, while the OsGLIP1-OE and OsGLIP2-OE plants exhibited more severe symptoms than the wild type (Fig 6A). We further compared the disease lesion sizes. Overexpression of OsGLIP1 and OsGLIP2 increased the lesion size to approximately 7 and 3 times, respectively, of the wild type. In contrast, the lesion area on OsGLIP1/2-RNAi leaves reduced to ~30% of wild-type plants (Fig 6B). Therefore, these pathogen inoculation assays again indicate that OsGLIP1 and OsGLIP2 negatively affect rice resistance to various pathogens.
Fig 6. OsGLIP genes negatively affect resistance to rice blast.
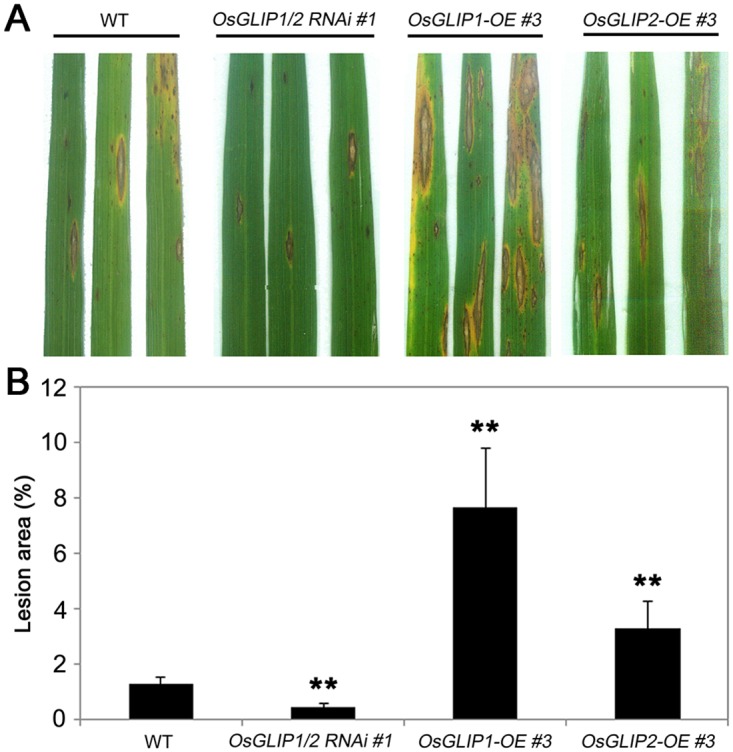
(A) Eight-week-old plants were inoculated with fungal pathogen M. oryzae by injection. Three leaves from the wild type and a representative line of OsGLIP1-OE, OsGLIP2-OE and OsGLIP1/2-RNAi transgenic plants were shown at 7 dpi. (B) Disease index of M. oryzae in the infected leaves of the wild-type and transgenic plants. Data are shown as relative lesion area compared to the whole leaf. Student’s t-test, ** P < 0.01.
Differential expression patterns of OsGLIP1 and OsGLIP2 in plants
Given the involvement of OsGLIP1 and OsGLIP2 in lipid metabolism and rice disease resistance, we were then interested in their spatiotemporal expression patterns in plants. Quantitative RT-PCR analysis showed that both OsGLIP1 and OsGLIP2 transcripts are detected in various rice tissues. OsGLIP1 expressed highly in leaves and sheaths, while OsGLIP2 mainly expressed in nodes, internodes and leaves (Fig 7A). To further determine their expression patterns, the promoters of OsGLIP1 and OsGLIP2 were used to drive β-glucuronidase (GUS) expression. Consistent with the qRT-PCR assay, GUS staining of transgenic plants expressing pOsGLIP1::GUS and pOsGLIP2::GUS revealed that OsGLIP1 is expressed in leaf, leaf sheath and young lemma (Fig 7B), while OsGLIP2 is expressed in node and internode (Fig 7C), demonstrating that OsGLIP1 and OsGLIP2 exhibited differential but complementary expression patterns.
Fig 7. Expression patterns of OsGLIP1 and OsGLIP2.

(A) Tissue-specific expression of OsGLIP1 and OsGLIP2 in different tissues detected by qRT-PCR. The expression level of OsGLIP1 was normalized to that of young seedling and OsGLIP2 to panicle. Data shown are means ±SD from three biological replicates. (B, C) GUS staining of pOsGLIP1::GUS and pOsGLIP2::GUS transgenic plants revealed OsGLIP1 and OsGLIP2 expression in panicle, leaf, leaf sheath, node and internode of heading plants. Scale bars = 0.5 cm.
Subcellular localization of OsGLIP1 and OsGLIP2
Arabidopsis GLIP1 and Tanacetum TcGLIP have been shown to be secreted into the intercellular space when transiently expressed in onion epidermal [17,21]. However, the localization of most lipases in plant cells still remains elusive. To investigate the subcellular localization of OsGLIP1 and OsGLIP2, both proteins were fused with green fluorescent protein (GFP) and expressed under the control of the maize Ubi1 promoter in stable transgenic rice. Xoo inoculation analysis indicated that OsGLIP1-GFP and OsGLIP2-GFP transgenic plants were more susceptible than the wild type, suggesting that the fusion proteins are functional equivalents to the wild-type proteins (S4 Fig). Localization of the fusion proteins were then visualized using confocal microscopy, and showed similar localization patterns. Both OsGLIP1-GFP and OsGLIP2-GFP proteins were observed principally in vesicle-like structures throughout the cytoplasm (Fig 8A). After plasmolysis, the OsGLIP-containing vesicles were mainly detected adjacent to the plasma membrane (PM) but some fluorescence signals remained in the apoplast (S5A and S5B Fig). To determine the nature of the apoplastic signals, high- or super- resolution microscopy would be required to test if OsGLIP1-GFP and OsGLIP2-GFP are normally secreted.
Fig 8. Subcellular localization of OsGLIP1 and OsGLIP2.

(A) Localization of OsGLIP1-GFP and OsGLIP2-GFP in the root cells of transgenic plants. OsGLIP1 and OsGLIP2 were fused with GFP and expressed under the control of the maize Ubiquitin (Ubi1) promoter in the stable transgenic rice plants. Scale bars = 20 μm. (B) A 3D projection of OsGLIP1-GFP fluorescence signals. Scale bar = 20 μm. (C) Images of OsGLIP1-GFP labelled vesicle-like structures (Type I and Type II). Scale bar = 2 μm. (D) Size distribution of OsGLIP1-GFP labelled vesicles. The size of vesicles is represented by the diameter. (E) The localization of OsGLIP1-GFP in response to BFA treatment, in comparison with mock treatment. Scale bar = 20 μm. (F) Co-localization of OsGLIP1-GFP with Nile Red that labels lipid droplets. Scale bars = 20 μm (left) or 5 μm (right). (G) Distribution of OsGLIP1-GFP and OsGLIP2-GFP proteins in fractionated membranes of two-week-old rice seedlings. Microsomal membranes were fractionated on linear 20% to 55% (w/v) sucrose gradients. Equal volumes of protein samples were separated on SDS-PAGE gel and analysed by immunoblot using antibodies specific for GFP (GLIP1/2-GFP), BiP2 (ER), HSP70 (cytoplasm) and ACTIN. Note that OsGLIP1 and OsGLIP2 mainly localize to the lipid bodies (cytoplasm) and the ER.
A 3D projection of OsGLIP1-GFP fluorescence signals indicated that the distribution of these OsGLIP-containing vesicles were not discrete but connected with each other by endomembrane systems (Fig 8B and S5C Fig). The size and morphology of these vesicles were found to be variable. Most of them are small and highly mobile (Type I) (Fig 8C, S1 Movie). There are also few much larger vesicles that are relatively static (Type II) (Fig 8C). Measurement of the vesicle size revealed that the diameter could be up to 10 μm but the majority of them were < 2 μm (Type I) (Fig 8D). The continuous trafficking of these small OsGLIP vesicles in the cytoplasm led us to examine their response to the treatment of brefeldin A (BFA), a specific vesicle trafficking inhibitor. We found BFA incubation resulted in aggregation of OsGLIP1-GFP fluorescence signals into big BFA compartments (Fig 8E). As BFA specifically blocks exocytosis but does not affect endocytosis, the sensitivity of OsGLIP vesicles to BFA treatment suggest that OsGLIP proteins might undergo active recycling.
The molecular nature of OsGLIP1 and OsGLIP2 as lipases and their vesicle localization pattern are reminiscent of lipid droplets. To directly verify this hypothesis, root cells expressing OsGLIP1-GFP were stained with Nile Red, a dye that specifically labels lipid bodies. The lipid staining assay revealed that in both small and large OsGLIP vesicles, GFP signals could overlap very well with the red signals from Nile Red (Fig 8F). The co-localization of OsGLIP1-GFP and OsGLIP2-GFP with Nile Red staining was further confirmed in rice protoplasts (S6 Fig). To further investigate the intracellular distribution of OsGLIP proteins, we carried out sucrose density gradient centrifugation assay. Microsomal membranes were prepared from OsGLIP1-GFP and OsGLIP2-GFP seedlings, fractionated on sucrose gradient (20% to 55%), and detected by Western blot analysis. We found that while both proteins are present in all membrane fractions, a majority of OsGLIP1-GFP and OsGLIP2-GFP proteins co-fractionate with the ER marker, BiP2 (Fig 8G). Taken together, these results suggest that OsGLIP proteins localize to endomembrane trafficking system as well as lipid droplets.
Proper intracellular localization is a prerequisite for OsGLIP1 function in immune inhibition
Proteins function with special subcellular localization, and it has been shown that the signal peptides of GDSL lipases are crucial for lipase localization [17,21]. Both OsGLIP1 and OsGLIP2 contain a typical signal peptide at the N-terminus with 29 and 35 aa, respectively. To evaluate the roles of the signal peptides in OsGLIP protein localization as well as biological functions, we generated two constructs, OsGLIP1ΔSP-GFP (without the signal peptide) and SP-GFP (the signal peptide alone), and transformed them into wild-type plants (Fig 9A). Confocal microscopy analysis revealed that the deletion of the OsGLIP1 signal peptide completely abolished its ER and lipid body localization, leading to OsGLIP1ΔSP-GFP localized ubiquitously in the cell including cytoplasm and nucleus (Fig 9B and 9C). By contrast, the signal peptide alone showed subcellular localization similar to the full length OsGLIP1 protein (Fig 9D), indicating that the signal peptide is necessary and sufficient for ER and lipid body targeting. Intriguingly, in contrast to the transgenic plants expressing full length OsGLIP1 protein compromising disease resistance, the transgenic plants expressing the truncated protein without signal peptide did not decrease disease resistance (Fig 9E and 9F), suggesting that the proper localization is critical for OsGLIP1 function in inhibiting plant immunity.
Fig 9. Proper intracellular localization is essential for OsGLIP1 function in rice defense responses.

(A) Schematic diagram shows full-length OsGLIP1 with its signal peptide (SP) and the truncated protein without SP (OsGLIP1ΔSP-GFP) or SP alone (OsGLIP1SP-GFP) fused with GFP. (B-D) Subcellular localization of OsGLIP1 -GFP (B) OsGLIP1ΔSP-GFP (C) and OsGLIP1SP-GFP (D) proteins in root cells of transgenic plants. Note that removing of the signal peptide (SP) abolished OsGLIP1-GFP ER and lipid body targeting, while the SP alone was sufficient for the subcellular compartment targeting. Scale bars = 20 μm. (E, F) Deletion of the signal peptide attenuated OsGLIP1 action in suppressing plant immunity. Lesions (E) and lesion lengths (F) of representative OsGLIP1-GFP and OsGLIP1ΔSP-GFP transgenic plants inoculated with Xoo. Note that the OsGLIP1-GFP fusion protein also suppressed rice defense, while OsGLIP1ΔSP-GFP lost its immune inhibition capacity. Arrows indicate bottoms of lesions. Data are shown as means ± SD (n > 10). Scale bar in (E) = 1cm. Student’s t-test, **P < 0.01.
MGDG and DGDG are involved in the inhibition of rice disease resistance
MGDG and DGDG are abundant galactolipids present in thylakoid membranes and photosynthesis system I and II, which is crucial for photosynthetic efficiency and plant development [30,31]. Interestingly, recent reports also showed that both MGDG and DGDG are required for the induction of SAR non-redundantly in Arabidopsis. DGDG contributes to NO and SA biosynthesis involved in defense responses, while MGDG promotes the biosynthesis of AzA and G3P that function downstream of NO [11,32]. In our current work, we found that the levels of both MGDG and DGDG were increased in OsGLIP1 and OsGLIP2 overexpression plants (Fig 3D and 3F). To investigate the roles of MGDG and DGDG in rice defense, we performed feeding assays. Exogenous application of MGDG and DGDG increased the levels of both lipids in the leaves fed (S7 Fig), and importantly they also facilitated the growth of Xoo in rice leaves in dependent inoculation experiments (Fig 10A and 10B). In addition, the basal level of PR gene expression as well as pathogen-induced PR gene upregulation were both compromised in MGDG and DGDG treated plants (Fig 10C–10E), suggesting that MGDG and DGDG likely play a negative role in rice immunity.
Fig 10. Exogenous feeding of MGDG and DGDG impairs rice disease resistance.
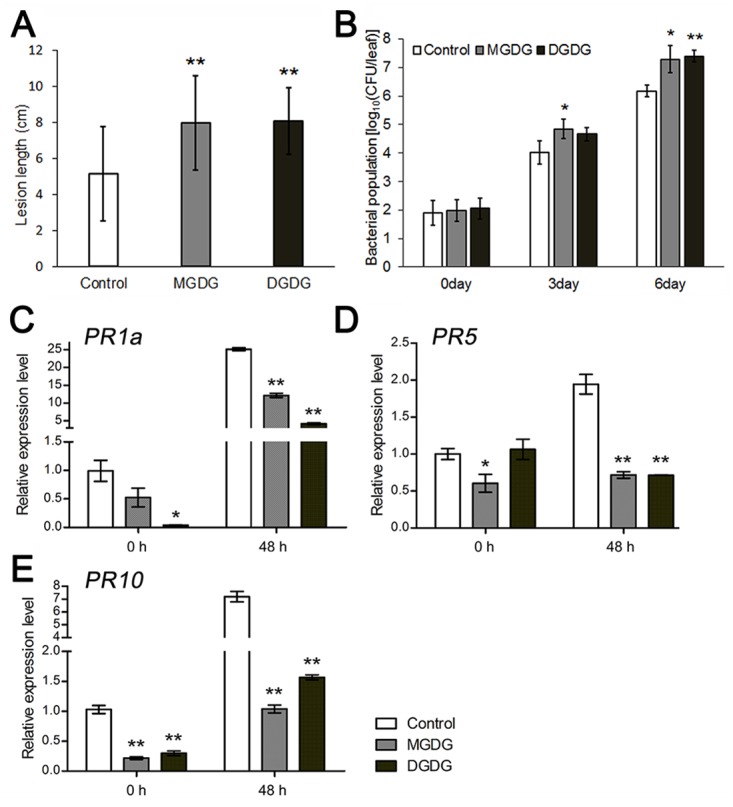
(A) Two-week-old seedling leaves were cultured in liquid medium supplemented with MGDG or DGDG (100 μM with 0.1% Tween-20) for 24 hours. The treated plants were extensively washed and then inoculated with Xoo (strain PXO99A). Lesion lengths of MGDG and DGDG-fed plants were measured at 10 dpi with three biological replicates (> 10 plants each replicate). Data are shown as means ± SD from three biological replicates. (B) Bacterial growth in the MGDG and DGDG-fed plants at 0, 3 and 6 dpi, with mock treatment as control. Data are shown as means ± SD from three biological replicates. (C-E) Relative expression levels of PR genes PR1a (C), PR5 (D) and PR10 (E) in MGDG/DGDG treated leaves at 0 and 48 hpi. Data are shown as means ± SD from three biological replicates (n = 3). Student’s t-test, *P<0.05, **P < 0.01 (A to E).
Discussion
Plant immune response is an energetically costly process that involves a rapid burst of ROS and programmed cell death [33]. To reduce the trade-off cost of defense over growth, a number of immune suppressors have been deployed such as rice NRR (an NPR1 interacting protein), WRKY transcription factors and SPL11 and EBR1 encoded E3 ubiquitin ligases, to compromise defense response thus maximizing fitness [34–38]. In this work, we have shown that two close GDSL lipases, OsGLIP1 and OsGLIP2, negatively affect rice immunity by modulation of lipid homeostasis, adding to the regulatory network of host-pathogen interaction and plant defense activation.
OsGLIP1 and OsGLIP2 modulates lipid metabolism
GDSL lipases belong to a newly classified lipase family [25]. Although there are more than 100 GDSL lipases predicted in the rice genome [22,23], only two of them, WDL1 (WILTED DWARF AND LETHAL 1) and GER1 (GDSL CONTAINING ENZYME RICE 1), have been functionally studied, which regulate cuticle formation and coleoptile elongation, respectively [39,40]. In this study, we showed that both OsGLIP1 and OsGLIP2 exhibited lipase activities that could hydrolyze common lipid substrates (Fig 2), as reported for other GDSL lipases in diverse plant species [17,19]. Plant GDSL lipases have been implicated in both developmental and physiological processes; however, only TcGLIP from Tanacetum cinerariifolium has been shown to catalyze the ester-forming reaction for pyrethrin synthesis [21]. While the in vivo substrates and products of most GDSL lipases still remain enigmatic.
Lipid profiling has enabled us to compare the composition and abundance of individual lipid species in OsGLIP transgenic plants with the established methods [41,42]. We found that the overexpression of OsGLIP1 and OsGLIP2 increased the levels of MGDG and DGDG, two abundant galactolipids present in plant cells and crucial for photosynthetic efficiency and plant development [30,31]. MGDG and DGDG are produced by the galactosylation of diacylglycerol (DAG) [43]. In eukaryotic cells, DAG is mainly derived either from PA through the action of phospholipases [44], or from TAG by TAG lipases [45]. Although DAG levels only showed slight difference in OsGLIP1 and OsGLIP2 transgenic plants, the amount of its precursors TAG and PA exhibited significant changes. In OsGLIP1-OE plants, the amount of TAG was reduced; while in OsGLIP2-OE plants, the PA level was strongly decreased, suggesting that OsGLIP1 might act on TAG while OsGLIP2 is probably involved in PA hydrolysis. Consistent with this hypothesis, simultaneous down-regulation of both genes led to increasing of both TAG and PA levels (Fig 9). In plant cells, the production of DAG from PA mainly takes place in the ER, and hydrolysis of TAG into DAG occurs in lipid droplets [44,46]. DAG is then rapidly imported into plastid membranes for MGDG and DGDG synthesis [46,47]. Subcellular localization of both OsGLIP1 and OsGLIP2 into lipid bodies as well as the ER correlates with their potential roles in lipid metabolism. Further biochemical assays to dissect the catalytic functions of recombinant OsGLIP proteins on various lipid substrates would provide invaluable insights into the activity of the lipases.
OsGLIP1 and OsGLIP2 are negative regulators of rice immunity
Recent studies are beginning to reveal roles of GDSL lipases in plant defense responses [17–21]. For instance, Arabidopsis GDSL lipases GLIP1 and GLIP2 exhibit antimicrobial activities and play positive roles in defense [17,19]. By contrast, virus-induced silencing of CaGLIP1 in pepper resulted in enhanced resistance to bacterial pathogen Xanthomonas campestris pv. vesicatoria (Xcv), while overexpression of this gene in Arabidopsis increased plant susceptibility to both bacterial and fungal pathogens, indicating that CaGLIP1 is an immune suppressor [20]. We proposed that OsGLIP1 and OsGLIP2 are suppressors of rice immunity based on both genetic and molecular evidence. Firstly, overexpression of OsGLIP1 and OsGLIP2 compromised plant resistance to both bacterial and fungal pathogens. Secondly, down-regulation of both genes significantly increased disease resistance. Finally, the up-regulation of PR genes in response to pathogen infection was suppressed in overexpression plants but substantially enhanced in OsGLIP1/2-RNAi plants compared to wild-type plants. With this scenario, OsGLIP1 and OsGLIP2 were down-regulated in response to pathogen infection and SA/BTH treatment. The dual roles of GDSL lipases as either positive or negative regulators of plant defense in different species reflect their broadly diverse catalytic activities in lipid metabolism, and substrates and/ or products of GDSL lipases likely function differentially in diverse plants [16], given that structural and biochemical analysis of bacteria GDSL lipases has revealed a flexible active site compatible with different substrates [48].
The altered disease resistance in OsGLIP transgenic plants was associated with significant changes of TAG, PA, MGDG and DGDG. Although the role of TAG in plant immunity remains to be elucidated, increasing evidence has implicated PA as an important regulator of plant immune responses. It has been shown that PA is rapidly produced upon pathogen infection/elicitor treatments. The elevated levels of PA subsequently induced ROS production, SA accumulation and defense activation [49–51]. Given the positive role of PA in plant immune responses, the increased/ decreased level of PA in OsGLIP1/2-RNAi/OsGLIP-OE plants might contribute to enhanced/compromised basal resistance to pathogens. More interestingly, exogenous application of MGDG and DGDG compromised rice disease resistance (Fig 10). Similarly, resistance was also compromised in OsGLIP1-OE and OsGLIP2-OE plants in which the abundance of MGDG and DGDG was increased. Consistent with these observations, both MGDG and DGDG contents have been shown to decrease in rice leaves infected by M. oryzae [52]. These findings together suggest that MGDG and DGDG most likely function as suppressor of rice immune responses. This is in contrast to the positive roles of MGDG and DGDG in Arabidopsis disease resistance via the establishment of SAR [11,32]. However, the SAR signaling pathway is not well documented in monocot plants such as rice [53]. Compared to the basal levels of SA in Arabidopsis, rice contains much higher levels of endogenous SA even in absence of pathogen infection [53,54]. Therefore, MGDG and DGDG, as well as GDSL lipases, would presumably play divergent roles in plant defense across different species.
Sub-functionalization of OsGLIP1 and OsGLIP2
Rice GDSL lipases constitute a large gene family which can be divided into four clades and twelve subclades, with each subclade consisting of sister gene pairs. Phylogenic analysis suggested that gene duplication plays a major role in the expansion of these GDSL genes [22]. Duplicated genes usually undergo sub-functionalization in protein activities, cellular localizations and/ or gene expression patterns [55]. For example, duplicated Populus Class III peroxidases exhibit divergent subcellular localization in either the cell wall or vacuole [56]. Rice cell-wall invertase genes GIF1 and OsCIN1 are a pair of duplicate genes required for seed development as well as disease resistance, GIF1 is mainly expressed in the ovular vascular tissue while OsCIN1 transcript is detected in pericarp and endosperm [57]. Similarly, we found that OsGLIP1 and OsGLIP2 also exhibited divergent expression patterns. The differential but also complementary expression patterns of OsGLIP1 and OsGLIP2 likely help to establish an elaborate metabolic cascade to regulate plant immunity throughout different tissues and development stages. Biochemical and transgenic analysis indicated that OsGLIP1 and OsGLIP2 proteins display similar catalytic activities and biological functions. However, comparison of lipid profiles revealed that the two lipases might have subtle difference in substrate preference in planta. Therefore, OsGLIP1 and OsGLIP2 might constitute a pair of genes with divergent expression patterns and different catalytic properties, and function synergistically to regulate rice immunity via modulation of lipid homeostasis in a temporal and spatial context.
Materials and methods
Plant materials, growth conditions and chemical induction
All the experiments were performed with rice variety Taipei 309 (TP309). Plants were grown in the paddy field under natural growing conditions. For experiments with seedlings, such as chemical treatments, two-week-old plants were grown in a growth chamber under conditions of 12-h day, 28°C, 80% RH followed by 12-h night, 26°C, 60% RH. BTH and SA treatments were performed according to the previous study [58]. BTH (in 0.5% acetone and 0.05% Tween 20) and SA (in 0.01% Tween 20) were sprayed onto leaves (1 ml/plant). Mock treatments were done by spraying the solvents only.
Gene cloning and plasmid construction
OsGLIP1 (1197 bp) and OsGLIP2 (1212 bp) cDNA were isolated according to rice genome information. The cDNAs were amplified further using primers GLIP1-OE-F/R and GLIP2-OE-F/R and inserted into the binary vector pUN1301 to form the OsGLIP1-OE and OsGLIP2-OE overexpression constructs. For RNAi cloning, we chose the conserved sequence of OsGLIP1 and OsGLIP2 coding region and amplified a short fragment with primers GLIP1/2-CK303-F (with BamHI site) and GLIP1/2-CK303-R (KpnI and SpeI sites). The fragments were introduced into vector pTCK303 [59]. The transgenic constructs were transformed into TP309 by the Agrobacterium-mediated transformation method. More than 30 independent transgenic lines were obtained for each construct. The primers for cloning used in this study are detailed in S1 Table.
Phylogenetic analysis
Protein sequences of GDSL lipase family members in rice and other organisms were retrieved from NCBI (http://www.ncbi.nlm.nih.gov). Multiple sequence alignments of protein were done in Clustal X (1.83). A phylogenetic tree of aligned sequence was constructed by MEGA (version 4.0.2) using the neighbor-joining method with the following parameters: Poisson correction, complete deletion, and bootstrap (1000 replicates, random seed).
Pathogen inoculation and disease assay
For Xoo resistance assay, eight-week-old plants were inoculated with Philippine race P6 (PXO99A) by the leaf-clipping method as previously described [60,61]. Lesion length was recorded 14 days after inoculation. More than thirty leaves were used for statistical analysis. For Xoo growth curve, 20 cm of leaf tissue from the top was ground and resuspended in 10 ml sterile water to collect bacteria. The suspensions were diluted accordingly and plated on peptone sucrose agar (PSA) plates containing 15 mg/l cephalexin. Bacteria clones were counted after 3 days incubation at 28°C.
The injection-inoculation method was employed for M. oryzae infection assay as described in [62]. Each tiller was injected with 0.1 ml mixed blast spore suspensions. Lesions developed on leaves were recorded 7 days after inoculation. Disease index was calculated by measuring the percentage of the lesion area (Lesion area/ Leaf area).
Protein purification and lipase activity assay
The OsGLIP1 and OsGLIP2 coding regions without N-terminal signal peptide sequences were amplified by primer GLIP1Δ29-enzyme-F/R and GLIP2Δ35-enzyme-F/R, respectively (S1 Table). The fragments were subsequently cloned into the pGEX-4T-3 vector (GE Healthcare). Constructs were transformed into Escherichia coli BL21 (DE3) cells and the expression of the recombinant proteins was induced by the addition of 0.2 mM isopropylthio-β-galactoside. Cells were harvested 6 hours after induction, suspended in PBS buffer and sonicated. The supernatants (lysates) were then purified using GST-tag beads (GE Healthcare). The purified proteins were confirmed by SDS-PAGE gel and stored at –70°C.
Lipase activity was measured as described [63]. The enzyme reaction mixture contained 0.5 M HEPES, pH 6.5, 1 mM substrate (p-nitrophenyl acetate or p-nitrophenyl butyrate), and aliquots of recombinant proteins (2 to 4 μg) and was incubated for 60 min at 30°C. Absorbance was measured at 405 nm every 5 min for 60 min.
Western blot of the fusion proteins (OsGLIP1-GFP and OsGLIP2-GFP) was performed with an anti-GFP antibody, signals were visualized by using ECL systems, and images were captured using the Tanon-5200 Chemiluminescent imaging system (Tanon).
Tissue-specific expression of OsGLIP1 and OsGLIP2
A 3.3-kb or 2.9-kb promoter region of OsGLIP1 or OsGLIP2 was amplified using primers GLIP1-promoter-F/R and GLIP2-promoter-F/R (S1 Table), respectively, and then inserted into the expression vector 1300-GUS-Nos to generate pOsGLIP1::GUS and pOsGLIP2::GUS. The constructs were transformed into TP309 to generate fusion reporter gene transgenic plants. For GUS staining, various tissues of pOsGLIP1::GUS and pOsGLIP2::GUS transgenic plants were incubated in a solution containing 50 mM NaPO4 buffer (pH 7.0), 5 mM K3Fe(CN)6, 5 mM K4Fe(CN)6, 0.1% Triton X-100, and 1 mM X-Gluc at 37°C.
Subcellular localization of OsGLIP1 and OsGLIP2
The full-length coding regions of OsGLIP1 and OsGLIP2 were amplified using primers GLIP1-GFP-F/R and GLIP2-GFP-F/R (S1 Table) and inserted into pUN1301 with in-frame fusion with GFP. The resulting constructs were transformed into TP309 to produce GFP fusion plants. GFP signals in roots were visualized using confocal laser scanning microscope (Zeiss LSM510 and Leica TCS SP8). For observation of protein localization in protoplasts, rice protoplasts were isolated from leaf sheaths of the OsGLIP1-GFP and OsGLIP2-GFP transgenic plants according to a previously reported method [64].
Quantitative real time-PCR
Total RNA was extracted from different tissues using TRIzol reagent according to the manufacturer’s instructions (Invitrogen). For RT–PCR analysis, 2 μg RNA was reverse-transcribed into cDNA using oligo (dT) primer and SuperScript III reverse transcriptase (Invitrogen) and then used as templates for PCR with gene-specific primers. Quantitative RT–PCR analysis was performed using SYBR Premix Ex Taq (TaKaRa) and gene-specific primers (S2 Table).
Lipid measurements
Lipid extraction was carried using modified Bligh & Dye’s protocol as previously described [65]. Leaves from six individual plants (eight-week-old) were mixed as one sample and five samples (5 replicates) from three independent transgenic lines of each transgene were used to normalize samples. All analyses were conducted using an Agilent 1260 HPLC system coupled with a triple quadrupole/ion trap mass spectrometer (5500Qtrap; SCIEX) in the electrospray ionization (ESI) mode under the following conditions: curtain gas = 20, ion spray voltage = 5500 V, temperature = 400°C, ion source gas 1 = 35, and ion source gas 2 = 35.
For normal phase (NP) LC/MS, polar lipid analysis was conducted as previously described [65]. Briefly, individual classes of polar lipids were separated by NP-HPLC with the use of a Phenomenex Luna 3μm-silica column (internal diameter 150 × 2.0 mm) under the following conditions: mobile phase A (chloroform: methanol: ammonium hydroxide, 89.5: 10: 0.5) and mobile phase B (chloroform: methanol: ammonium hydroxide: water, 55: 39: 0.5: 5.5). The gradient began with 95% mobile phase A for 5 min, followed by linear reduction to 60% mobile phase A over 7 min. The gradient was held for 4 min, and mobile phase A was then further reduced to 30% and held for 15 min. The column was then reconditioned with the initial gradient for 5 min.
MRM transitions were constructed for comparative analysis of various polar lipids. Quantification of individual lipid species were carried out by referencing to spiked internal standards; namely PC-14:0/14:0, PC34:1-d31, PE-14:0/14:0, PE34:1-d31, PS-14:0/14:0, PA-17:0/17:0, PG-14:0/14:0, which were obtained from Avanti Polar Lipids (Alabaster, AL) and LIPID MAPS. Dioctanoyl phosphatidylinositol (PI) (16:0-PI) was purchased from Echelon Biosciences, Inc. (Salt Lake City, UT) and used together with PI34:1-d31 (LIPID MAPS) for PI quantitation. Qualitative deuterated lipid standards from LIPID MAPS were pre-corrected based on available quantitative lipid standards prior to their use for quantitation.
For reverse phase LC/MS, glycerol lipids [DAGs and TAGs] were analyzed using a modified version of reverse phase (RP)-HPLC/ESI/MS/MS as reported previously [65]. In brief, separation of the aforementioned lipids was carried out on a Phenomenex Kinetex 2.6μm-C18 column (internal diameter 4.6×100 mm) using an isocratic mobile phase chloroform:methanol:0.1M ammonium acetate (100:100:4) at a flow rate of 160 μl/min for 20 min. Based on neutral loss MS/MS techniques, the levels of TAGs were calculated relative to the intensity of spiked d5-TAG 48:0 internal standard (CDN Isotopes), while DAG species were quantified using 4ME 16:0 diether DG as an internal standard (Avanti Polar Lipids). Separation of galatolipids (MGDGs and DGDGs) were conducted using Phenomenex Kinetex 2.6μm-C18 column (internal diameter 4.6 × 100 mm) with an isocratic gradient of chloroform:methanol: 2% of 50 mmol/L sodium acetate (49: 49: 2) at a flow rate of 160 μl/min for 25 min as reported previously [42]. MRM (multiple reaction monitoring) and SIM (selective ion monitoring) transitions for lipids presented in this study were provided in S3 Table.
MGDG and DGDG feeding assays
MGDG and DGDG (Avanti Polar Lipids, Inc) were dissolved in acetone respectively, and then diluted with water to 100 μM with 0.1% Tween-20. Leaves of two-week-old seedlings (TP309) were soaked in the MGDG or DGDG solution for 24 hours, with mock (acetone diluted with a comparable amount of water with 0.1% Tween-20) solution as control. Leaves were immediately were washed extensively to remove residual solution. Treated seedlings were inoculated with Xoo (strain PXO99A) by the leaf-clipping method [60]. Xoo growth curve was assayed as described above.
To measure the content of MGDG and DGDG in fed leaves, leaves after feeding were extensively washed to remove residual solution. Lipid samples were prepared according to the methods described in [52]. In detail, fresh seedling leaves (0.15g) were powdered with liquid N2. A solution (1.5mL) of methanol: chloroform: water (1: 1: 0.74, v/v) containing 200 μM L-1 of butylated hydroxytoluene was then added to extract total lipids. Chloroform layer was then concentrated with an Eppendorf concentrator. Extracts were kept at -20°C in tubes filled with gaseous N2 to avoid oxidation and hydrogenation of the galactolipids. Extracts were dissolved in 200 μL acetonitrile: water: isopropanol (3: 2: 5, v/v, 5mM ammonium formate), then diluted by 20 times for measuring.
Liquid chromatography–high resolution mass spectrometry (LC–HRMS) was applied to determine the contents of MGDG and DGDG. HRMS data was obtained from a Q-Exactive Quadrupole Orbitrap mass spectrometer (Thermo Fisher Scientific, Bremen, Germany) coupled to an Acquity Ultra Performance LC system (Waters). An Acquity UPLC BEH C18 column (2.1×50mm, 1.7 μm) was used at 40°C. The lipid extract of 1 μl was injected at 12°C. Acetonitrile:water (60/40, v/v, 10mM ammonium formate) and isopropanol:acetonitrile (90/10, v/v) were used as mobile phases A and B, respectively. The flow rate was 0.3 mL/min. The elution was performed with a 14 min gradient with 30% B at the beginning, then linearly increased from 30% B to 95% B in 8 min. After washing the column for 2 min with 95% B, the buffer was decreased to 30% B immediately and the column was reconditioned with initial gradient for 4 min. The MS was operated with HESI ion source in positive mode. The following conditions were used for HESI source: capillary voltage, 4.0 KV; capillary temperature, 320°C; sheath gas, 35.00 units; auxiliary gas, 5.00 units; probe heater temperature, 300°C; S-Lens RF level, 60.00. The mass spectrometer was run in full MS-ddMS2 mode. The full MS scan used the following settings: resolution, 70,000; AGC target, 3e6; scan range, 500–1200 m/z. The ms2 scan parameters: Loop count, 5; resolution, 17,500; AGC target, 1e5; max IT, 50 ms; isolation window, 1.0 m/z; normalized HCD collision energy, 15 eV and 35 eV. Standard DGDG and MGDG (Avanti Polar Lipids) were used to make standard curves. The resulting linear equation in our experiments for MGDG was Y = -3.34662 × 107 + 4.12273 × 106X, R2 = 0.9967; and for DGDG, Y = -1.39113 × 107 + 5.6299×106X, R = 0.9985, with X as amounts of MGDG or DGDG (μg), and Y as the integrated area.
Sucrose density-gradient centrifugation
Two-week-old plants were used for membrane fractionation. Membrane fractionation was carried out by a protocol described previously [66,67] with some modifications. In brief, tissues were homogenized in cold buffer [30mM Tris, pH7.5, 150mM NaCl, 10mM EDTA, 20% glycerol with proteins inhibitor cocktail (Roche)], and were filtered through two layers of Miracloth (Merck) and then centrifuged at 10,000g for 10 min at 4°C. Supernatants were centrifuged again at 100,000g for 50 min at 4°C to pellet the membrane fraction. The resulting microsome pellets were resuspended in 0.5 mL of resuspension buffer [10mM Tris, pH7.5, 10mM EDTA, 10% sucrose with 1mM DTT and proteins inhibitor cocktail (Roche)]. Serial 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55% (w/v) sucrose solutions were prepared with 20 mM HEPES (pH7.5), 1 mM EDTA, and were then added to ultracentrifuge tubes (Beckman SW14) one by one to form continuous gradients. Lipid samples (0.5 ml) were added and centrifuged at 100,000g for 12 h with an ultracentrifuge (Optima L-90K, Beckman). Fractions (1 ml) were collected from top to bottom and transferred to new tubes for protein assay.
Accession numbers
Sequence data from this article can be found in the GenBank/EMBL database under the following accession numbers: KU350624 (OsGLIP1), NM_001063391 (OsGLIP2).
Supporting information
(DOCX)
(DOCX)
(DOCX)
(A) Gene structure of OsGLIP1 and OsGLIP2. (B) A phylogenetic tree of GDSL lipases in rice. Protein sequences were aligned with CLUSTAL W, and the phylogenetic tree was constructed with MEGA4.
(PDF)
The expression of OsGLIP1 and OsGLIP2 in 9 independent OsGLIP1-OE (A) and OsGLIP2-OE (B) lines and 8 independent OsGLIP1/2-RNAi lines (C) was compared to the wild-type control with normalization to rice Actin1 gene. Error bars, ± SD (n = 3).
(PDF)
The lipids, PC (A), PI (B), PE (C) and PG (D), are presented as the number of carbon atoms: total double bonds in the fatty acyl groups. Data are shown as means ± SD (n = 5) of mixed leaf samples from three representative transgenic lines. *P < 0.05 or **P < 0.01, by Student’s t-test and Bonferroni correction for multiple (three comparisons) tests.
(PDF)
(A) Lesion lengths in the leaves of wild-type and independent OsGLIP1-GFP and OsGLIP2-GFP transgenic plants. Plants at booting stage were infected with Xoo. Lesion length was calculated at 14 dpi. Data are shown as means ± SD (n > 10). Student’s t-test, **P < 0.01. (B) Western blot analysis with GFP antibodies confirmed the accumulation of the fusion proteins of OsGLIP1-GFP and OsGLIP2-GFP in the transgenic plants. Rubisco staining was used as loading control. (C) Bacterial growth during 8 days of inoculation in the representative transgenic lines. Data are shown as means ± SD (n = 3). Asterisks indicate significant difference in comparison with the wild-type control (Student’s t-test, *P < 0.05; ** P < 0.01).
(PDF)
The root tips of transgenic plants were incubated in 25% sucrose to induce cell plasmolysis and then observed under confocal microscopy. (A, B) Confocal images of OsGLIP1–GFP (A) and OsGLIP2–GFP (B) after cell plasmolysis. Scale bars = 20 μm. (C) OsGLIP1-GFP root cells without plasmolysis to show the endomembrane systems that link OsGLIP1-GFP labelled vesicles. Scale bar = 20 μm.
(PDF)
Protoplasts were generated from seedlings of transgenic plants expressing OsGLIP1-GFP and OsGLIP2-GFP fusion protein. Note the overlapping of GFP green signals and Nile Red stained red signals, while OsGLIP1-GFP and OsGLIP2-GFP were not associated with chloroplasts. Scale bars = 10 μm.
(PDF)
Two-week-old rice leaves were fed with 100μM MGDG (A) and DGDG (B) for 24 hours and subsequently used for lipid extraction and measurements. Data are shown as means ± SD of four biological replicates. Student’s t-test. *P < 0.05.
(PDF)
(AVI)
Acknowledgments
We thank Longjun Zeng, Jianjun Wang, Xiaoming Zhang, Junmin Wang and Shenghai Ye for help with rice growth, Genyun Chen for help with protein purification, Yuanhong Shan for DGDG and MGDG measurement. We also thank Hajk-Georg Drost for statistical analysis of lipid profiling data, and Raymond Wightman for careful reading of the manuscript.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by grants from National Natural Science Foundation of China (http://www.nsfc.gov.cn/) (31330061), the National GMO project (http://www.nmp.gov.cn/zxjs/200901/t20090113_2113.htm (2013ZX08009-003-001), and the Shanghai Committee of Science and Technology (http://www.stcsm.gov.cn/) (14JC1406700). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Pollard M, Beisson F, Li Y, Ohlrogge J (2008) Building lipid barriers: biosynthesis of cutin and suberin. Trends Plant Sci 13: 236–246. doi: 10.1016/j.tplants.2008.03.003 [DOI] [PubMed] [Google Scholar]
- 2.Jones JDG, Dangl JL (2006) The plant immune system. Nature 444: 323–329. doi: 10.1038/nature05286 [DOI] [PubMed] [Google Scholar]
- 3.Shah J (2005) Lipids, lipases, and lipid-modifying enzymes in plant disease resistance. Annu Rev Phytopathol 43: 229–260. doi: 10.1146/annurev.phyto.43.040204.135951 [DOI] [PubMed] [Google Scholar]
- 4.Reina-Pinto JJ, Yephremov A (2009) Surface lipids and plant defenses. Plant Physiol Biochem 47:540–549. doi: 10.1016/j.plaphy.2009.01.004 [DOI] [PubMed] [Google Scholar]
- 5.Hématy K, Cherk C, Somerville S (2009) Host-pathogen warfare at the plant cell wall. Curr Opin Plant Biol 12: 406–413. doi: 10.1016/j.pbi.2009.06.007 [DOI] [PubMed] [Google Scholar]
- 6.Kachroo A, Kachroo P (2009) Fatty acid-derived signals in plant defense. Annu Rev Phytopathol 47: 153–176. doi: 10.1146/annurev-phyto-080508-081820 [DOI] [PubMed] [Google Scholar]
- 7.Farmer EE, Almeras E, Krishnamurthy V (2003) Jasmonates and related oxylipins in plant responses to pathogenesis and herbivory. Curr Opin Plant Biol 6: 372–378. doi: 10.1016/S1369-5266(03)00045-1 [DOI] [PubMed] [Google Scholar]
- 8.Kachroo A, Venugopal SC, Lapchyk L, Falcone D, Hildebrand D, Kachroo P (2004) Oleic acid levels regulated by glycerolipid metabolism modulate defense gene expression in Arabidopsis. Proc Natl Acad Sci USA 101: 5152–5157. doi: 10.1073/pnas.0401315101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Chandra-Shekara AC, Venugopal SC, Barman SR, Kachroo A, Kachroo P (2007) Plastidial fatty acid levels regulate resistance gene-dependent defense signaling in Arabidopsis. Proc Natl Acad Sci USA 104: 7277–7282. doi: 10.1073/pnas.0609259104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Fu ZQ, Dong X (2013) Systemic acquired resistance: turning local infection into global defense. Annu Rev Plant Biol 64: 839–863. doi: 10.1146/annurev-arplant-042811-105606 [DOI] [PubMed] [Google Scholar]
- 11.Gao QM, Zhu S, Kachroo P, Kachroo A (2015) Signal regulators of systemic acquired resistance. Front Plant Sci 6:228 doi: 10.3389/fpls.2015.00228 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Maldonado AM, Doerner P, Dixon RA, Lamb CJ and Cameron RK (2002) A putative lipid transfer protein involved in systemic resistance signalling in Arabidopsis. Nature 419: 399–403. doi: 10.1038/nature00962 [DOI] [PubMed] [Google Scholar]
- 13.Jung HW, Tschaplinski TJ, Wang L, Glazebrook J, Greenberg JT (2009) Priming in systemic plant immunity. Science 324: 89–91. doi: 10.1126/science.1170025 [DOI] [PubMed] [Google Scholar]
- 14.Chanda B, Xia Y, Mandal MK, Yu K, Sekine KT, Gao QM, Selote D, Hu Y, Stromberg A, Navarre D, et al. (2011) Glycerol-3-phosphate is a critical mobile inducer of systemic immunity in plants. Nat Genet 43: 421–427. doi: 10.1038/ng.798 [DOI] [PubMed] [Google Scholar]
- 15.Yu K, Soares JM, Mandal MK, Wang C, Chanda B, Gifford AN, Fowler JS, Navarre D, Kachroo A, Kachroo P (2013) A feedback regulatory loop between G3P and lipid transfer proteins DIR1 and AZI1 mediates azelaic-acid-induced systemic immunity. Cell Rep 3: 1266–1278. doi: 10.1016/j.celrep.2013.03.030 [DOI] [PubMed] [Google Scholar]
- 16.Akoh CC, Lee GC, Liaw YC, Huang TH, Shaw JF (2004) GDSL family of serine esterases/lipases. Prog Lipid Res 43: 534–552. doi: 10.1016/j.plipres.2004.09.002 [DOI] [PubMed] [Google Scholar]
- 17.Oh IS, Park AR, Bae MS, Kwon SJ, Kim YS, Lee JE, Kang NY, Lee SM, Cheong HS, Park OM (2005) Secretome analysis reveals an Arabidopsis lipase involved in defense against Alternaria brassicicola. Plant Cell 17: 2832–2847. doi: 10.1105/tpc.105.034819 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kwon SJ, Jin HC, Lee S, Nam MH, Chung JH, Kwon SI, Ryu CM, Park OK (2009) GDSL lipase-like 1 regulates systemic resistance associated with ethylene signaling in Arabidopsis. Plant J 58: 235–245. doi: 10.1111/j.1365-313X.2008.03772.x [DOI] [PubMed] [Google Scholar]
- 19.Lee DS, Kim BK, Kwon SJ, Jin HC, Park OK (2009) Arabidopsis GDSL lipase 2 plays a role in pathogen defense via negative regulation of auxin signaling. Biochem. Biophys. Res. Commun. 379: 1038–1042. doi: 10.1016/j.bbrc.2009.01.006 [DOI] [PubMed] [Google Scholar]
- 20.Hong JK, Choi HW, Hwang IS, Kim DS, Kim NH, Choi du S, Kim YJ, Hwang BK (2008) Function of a novel GDSL-type pepper lipase gene, CaGLIP1, in disease susceptibility and abiotic stress tolerance. Planta 227: 539–558. doi: 10.1007/s00425-007-0637-5 [DOI] [PubMed] [Google Scholar]
- 21.Kikuta Y, Ueda H, Takahashi M, Mitsumori T, Yamada G, Sakamori K, Takeda K, Furutani S, Nakayama K, Katsuda Y, et al. (2012) Identification and characterization of a GDSL lipase-like protein that catalyzes the ester-forming reaction for pyrethrin biosynthesis in Tanacetum cinerariifolium—A new target for plant protection. Plant J. 71: 183–193. doi: 10.1111/j.1365-313X.2012.04980.x [DOI] [PubMed] [Google Scholar]
- 22.Chepyshko H, Lai CP, Huang LM, Liu JH, Shaw JF (2012) Multifunctionality and diversity of GDSL esterase/lipase gene family in rice (Oryza sativa L. japonica) genome: New insights from bioinformatics analysis. BMC Genomics 13: 309 doi: 10.1186/1471-2164-13-309 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Jiang YY, Chen RJ, Dong JL, Xu ZJ, Gao XL (2012) Analysis of GDSL lipase (GLIP) family genes in rice (Oryza sativa). Plant Omics 5: 351–358. [Google Scholar]
- 24.Li Q, Chen F, Sun L, Zhang Z, Yang Y, He Z (2006) Expression profiling of rice genes in early defense responses to blast and bacterial blight pathogens using cDNA microarray. Physiol Mol Plant Pathol 68:51–60. doi: 10.1016/j.pmpp.2006.06.002 [Google Scholar]
- 25.Volokita M, Rosilio-Brami T, Rivkin N, Zik M (2011) Combining comparative sequence and genomic data to ascertain phylogenetic relationships and explore the evolution of the large GDSL-lipase family in land plants. Mol Biol Evol 28: 551–565. doi: 10.1093/molbev/msq226 [DOI] [PubMed] [Google Scholar]
- 26.Nielsen H, Engelbrecht J, Brunak S, and von Heijne G (1997) Identification of prokaryotic and eukaryotic signal peptides and prediction of their cleavage sites. Protein Eng 10: 1–6. [DOI] [PubMed] [Google Scholar]
- 27.van Loon LC, Rep M, Pieterse CM (2006) Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 44: 135–162. doi: 10.1146/annurev.phyto.44.070505.143425 [DOI] [PubMed] [Google Scholar]
- 28.Xiao W, Liu H, Li Y, Li X, Xu C, Long M, Wang S (2009) A rice gene of de novo origin negatively regulates pathogen-induced defense response. PLoS One 4: e4603 doi: 10.1371/journal.pone.0004603 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Deng H, Liu H, Li X, Xiao J, Wang S (2012) A CCCH-type zinc finger nucleic acid-binding protein quantitatively confers resistance against rice bacterial blight disease. Plant Physiol 158:876–889. doi: 10.1104/pp.111.191379 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Benning C, Ohta H (2005) Three enzyme systems for galactoglycerolipid biosynthesis are coordinately regulated in plants. J Biol Chem 280: 2397–2400. doi: 10.1074/jbc.R400032200 [DOI] [PubMed] [Google Scholar]
- 31.Kobayashi K, Kondo M, Fukuda H, Nishimura M, Ohta H (2007) Galactolipid synthesis in chloroplast inner envelope is essential for proper thylakoid biogenesis, photosynthesis, and embryogenesis. Proc Natl Acad Sci USA 104: 17216–17221. doi: 10.1073/pnas.0704680104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Gao QM, Yu K, Xia Y, Shine MB, Wang C, Navarre D, Kachroo A, Kachroo P (2014) Mono- and digalactosyldiacylglycerol lipids function nonredundantly to regulate systemic acquired resistance in plants. Cell Rep 9: 1681–1691. doi: 10.1016/j.celrep.2014.10.069 [DOI] [PubMed] [Google Scholar]
- 33.Huot B, Yao J, Montgomery BL, He SY (2014) Growth-defense tradeoffs in plants: a balancing act to optimize fitness. Molecular Plant 7: 1267–1287. doi: 10.1093/mp/ssu049 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Chern M, Canlas PE, Fitzgerald HA, Ronald PC (2005) Rice NRR, a negative regulator of disease resistance, interacts with Arabidopsis NPR1 and rice NH1. Plant J 43: 623–635. doi: 10.1111/j.1365-313X.2005.02485.x [DOI] [PubMed] [Google Scholar]
- 35.Peng Y, Bartley LE, Chen X, Dardick C, Chern M, Ruan R, Canlas PE, Ronald PC (2008) OsWRKY62 is a negative regulator of basal and Xa21-mediated defense against Xanthomonas oryzae pv. oryzae in rice. Mol Plant 1: 446–458. doi: 10.1093/mp/ssn024 [DOI] [PubMed] [Google Scholar]
- 36.Pandey SP, Somssich IE (2009) The role of WRKY transcription factors in plant immunity. Plant Physiol 150: 1648–1655. doi: 10.1104/pp.109.138990 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Zeng LR, Qu S, Bordeos A, Yang C, Baraoidan M, Yan H, Xie Q, Nahm BH, Leung H, Wang GL (2004) Spotted leaf11, a negative regulator of plant cell death and defense, encodes a U-box/armadillo repeat protein endowed with E3 ubiquitin ligase activity. Plant Cell 16: 2795–2808. doi: 10.1105/tpc.104.025171 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.You Q, Zhai K, Yang D, Yang W, Wu J, Liu J, Pan W, Wang J, Zhu X, Jian Y, Liu J, Zhang Y, Deng Y, Li Q, Lou Y, Xie Q, He Z (2016) An E3 ubiquitin ligase-BAG protein module controls plant innate immunity and broad-spectrum disease resistance. Cell Host Microbe. 20: 758–769. doi: 10.1016/j.chom.2016.10.023 [DOI] [PubMed] [Google Scholar]
- 39.Park JJ, Jin P, Yoon J, Yang JI, Jeong HJ, Ranathunge K, Schreiber L, Franke R, Lee IJ, An G (2010) Mutation in Wilted Dwarf and Lethal 1 (WDL1) causes abnormal cuticle formation and rapid water loss in rice. Plant Mol. Biol. 74: 91–103. doi: 10.1007/s11103-010-9656-x [DOI] [PubMed] [Google Scholar]
- 40.Riemann M, Gutjahr C, Korte A, Danger B, Muramatsu T, Bayer U, Waller F, Furuya M, Nick P (2007) GER1, a GDSL motif-encoding gene from rice is a novel early light- and jasmonate-induced gene. Plant Biol (Stuttg) 9: 32–40. doi: 10.1055/s-2006-924561 [DOI] [PubMed] [Google Scholar]
- 41.Welti R, Wang X (2004) Lipid species profiling: A high throughput approach to identify lipid compositional changes and determine the function of genes involved in lipid metabolism and signaling. Curr Opin Plant Biol 7: 337–344. doi: 10.1016/j.pbi.2004.03.011 [DOI] [PubMed] [Google Scholar]
- 42.Cheong WF, Wenk MR, Shui G (2014) Comprehensive analysis of lipid composition in crude palm oil using multiple lipidomic approaches. J Genet Genomics 41: 293–304. doi: 10.1016/j.jgg.2014.04.002 [DOI] [PubMed] [Google Scholar]
- 43.Rousseau B, Lopez R, Maréchal E (2011) Chemical inhibitors of monogalactosyldiacylglycerol synthases in Arabidopsis thaliana. Nature Chem Biol 7: 834–842. doi: 10.1038/nchembio.658 [DOI] [PubMed] [Google Scholar]
- 44.Bates PD, Browse J (2012) The significance of different diacylgycerol synthesis pathways on plant oil composition and bioengineering. Front Plant Sci 3: 147 doi: 10.3389/fpls.2012.00147 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Watt MJ, Steinberg GR (2008) Regulation and function of triacylglycerol lipases in cellular metabolism. Biochem J 414: 313–325. doi: 10.1042/BJ20080305 [DOI] [PubMed] [Google Scholar]
- 46.Awai K, Xu C, Lu B, Benning C (2006) Lipid trafficking between the endoplasmic reticulum and the chloroplast. Biochem Soc Trans 34: 395–398. doi: 10.1042/BST0340395 [DOI] [PubMed] [Google Scholar]
- 47.Xu C, Fan J, Cornish AJ, Benning C (2008) Lipid trafficking between the endoplasmic reticulum and the plastid in Arabidopsis requires the extraplastidic TGD4 protein. Plant Cell 20: 2190–2204. doi: 10.1105/tpc.108.061176 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Lo YC, Lin SC, Shaw JF, Liaw YC (2003) Crystal structure of Escherichia coli thioesterase I/protease I/lysophospholipase L1: consensus sequence blocks constitute the catalytic center of SGNH-hydrolases through a conserved hydrogen bond network. J Mol Biol, 330: 539–551. doi: 10.1016/S0022-2836(03)00637-5 [DOI] [PubMed] [Google Scholar]
- 49.De Jong CF, Laxalt AM, Bargmann BO, DeWit PJ, Joosten MH, Munnik T (2004) Phosphatidic acid accumulation is an early response in the Cf-4/Avr4 interaction. Plant J 39: 1–12. doi: 10.1111/j.1365-313X.2004.02110.x [DOI] [PubMed] [Google Scholar]
- 50.Zhang Y, Zhu H, Zhang Q, Li M, Yan M, Wang R, Wang L, Welti R, Zhang W, Wang X (2009) Phospholipase Dα1 and phosphatidic acid regulate NADPH oxidase activity and production of reactive oxygen species in ABA-mediated stomatal closure in Arabidopsis. Plant Cell 21: 2357–2377. doi: 10.1105/tpc.108.062992 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Zhang Q, Xiao S (2015) Lipids in salicylic acid- mediated defense in plants: focusing on the roles of phosphatidic acid and phosphatidylinositol 4-phosphate. Front Plant Sci 6:387 doi: 10.3389/fpls.2015.00387 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Zhou L, Y. Zhang Y, Rao L, Cao Y and Peng Y (2008) Galactolipid depletion in blast fungus-infected rice leaves. Ann Appl Biol 153: 395–400. doi: 10.1111/j.1744-7348.2008.00266.x [Google Scholar]
- 53.Yuan YX, Zhong SH, Li Q, Zhu ZR, Lou YG, Wang LY, Wang JJ, Wang MY, Li QL, Yang DL, He ZH (2007) Functional analysis of rice NPR1-like genes reveals that OsNPR1/NH1 is the rice orthologue conferring disease resistance with enhanced herbivore susceptibility. Plant Biotech J 5: 313–324. doi: 10.1111/j.1467-7652.2007.00243.x [DOI] [PubMed] [Google Scholar]
- 54.Silverman P, Seskar M, Kanter D, Schweizer P, Me´traux JP, and Raskin I (1995) Salicylic acid in rice (biosynthesis, conjugation, and possible role). Plant Physiol 108: 633–639. doi: 10.1104/pp.108.2.633 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Prince VE, Pickett FB (2002) Splitting pairs: the diverging fates of duplicated genes. Nat Rev Genet 3: 827–837. doi: 10.1038/nrg928 [DOI] [PubMed] [Google Scholar]
- 56.Ren LL, Liu YJ, Liu HJ, Qian TT, Qi LW, Wang XR, Zeng QY (2014) Subcellular relocalization and positive selection play key roles in the retention of duplicate genes of Populus class III peroxidase family. Plant Cell 26: 2404–2419. doi: 10.1105/tpc.114.124750 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Wang E, Xu X, Zhang L, Zhang H, Lin L, Wang Q, Li Q, Ge S, Lu BR, Wang W, He Z (2010) Duplication and independent selection of cell-wall invertase genes GIF1 and OsCIN1 during rice evolution and domestication. BMC Evol Biol 10: 108 doi: 10.1186/1471-2148-10-108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Shimono M, Sugano S, Nakayama A, Jiang CJ, Ono K, Toki S, Takatsuji H (2007) Rice WRKY45 plays a crucial role in benzothiadiazole-inducible blast resistance. Plant Cell 19: 2064–2076. doi: 10.1105/tpc.106.046250 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Wang Z, Chen C, Xu Y, Jiang R, Han Y, Xu Z, Chong K (2004) A practical vector for efficient knockdown of gene expression in rice (Oryza sativa L.). Plant Mol Biol Rep 22: 409–417. doi: 10.1007/BF02772683 [Google Scholar]
- 60.Yang D, Li Q, Deng Y, Lou Y, Wang M, Zhou G, Zhang Y, He Z (2008) Altered disease development in the eui mutants and Eui overexpressors indicates that gibberellins negatively regulate rice basal disease resistance. Mol Plant 1: 528–537. doi: 10.1093/mp/ssn021 [DOI] [PubMed] [Google Scholar]
- 61.Li X, Yang DL, Sun L, Li Q, Mao B, He Z (2016) The Systemic Acquired Resistance Regulator OsNPR1 Attenuates Growth by Repressing Auxin Signaling through Promoting IAA-Amido Synthase Expression. Plant Physiol. 172:546–558. doi: 10.1104/pp.16.00129 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Deng Y, Zhai K, Xie Z, Yang D, Zhu X, et al. (2017) Epigenetic regulation of antagonistic receptors confers rice blast resistance with yield balance. Science 355: 962–965. doi: 10.1126/science.aai8898 [DOI] [PubMed] [Google Scholar]
- 63.Baudouin E, Charpenteau M, Roby D, Marco Y, Ranjeva R, Ranty B (1997) Functional expression of a tobacco gene related to the serine hydrolase family—Esterase activity towards short-chain dinitrophenyl acylesters. Eur J Biochem 248: 700–706. doi: 10.1111/j.1432-1033.1997.t01-1-00700.x [DOI] [PubMed] [Google Scholar]
- 64.Yang W, Gao M, Yin X, Liu J, Xu Y, Zeng L, Li Q, Zhang S, Wang J, Zhang X, He Z (2013) Control of rice embryo development, shoot apical meristem maintenance, and grain yield by a novel cytochrome p450. Mol Plant 6: 1945–1960. doi: 10.1093/mp/sst107 [DOI] [PubMed] [Google Scholar]
- 65.Lam SM, Tong L, Duan X, Petznick A, Wenk MR, Shui G (2014) Extensive characterization of human tear fluid collected using different techniques unravels the presence of novel lipid amphiphiles. J Lipid Res 55: 289–298. doi: 10.1194/jlr.M044826 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Qiao H, Shen Z, Huang SS, Schmitz RJ, Urich MA, Briggs SP, Ecker JR (2012) Processing and subcellular trafficking of ER-tethered EIN2 control response to ethylene gas. Science 338: 390–393. doi: 10.1126/science.1225974 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Wulfetange K, Lomin SN, Romanov GA, Stolz A, Heyl A, Schmulling T (2011) The cytokinin receptors of Arabidopsis are located mainly to the endoplasmic reticulum. Plant Physiol 156: 1808–1818. doi: 10.1104/pp.111.180539 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOCX)
(DOCX)
(DOCX)
(A) Gene structure of OsGLIP1 and OsGLIP2. (B) A phylogenetic tree of GDSL lipases in rice. Protein sequences were aligned with CLUSTAL W, and the phylogenetic tree was constructed with MEGA4.
(PDF)
The expression of OsGLIP1 and OsGLIP2 in 9 independent OsGLIP1-OE (A) and OsGLIP2-OE (B) lines and 8 independent OsGLIP1/2-RNAi lines (C) was compared to the wild-type control with normalization to rice Actin1 gene. Error bars, ± SD (n = 3).
(PDF)
The lipids, PC (A), PI (B), PE (C) and PG (D), are presented as the number of carbon atoms: total double bonds in the fatty acyl groups. Data are shown as means ± SD (n = 5) of mixed leaf samples from three representative transgenic lines. *P < 0.05 or **P < 0.01, by Student’s t-test and Bonferroni correction for multiple (three comparisons) tests.
(PDF)
(A) Lesion lengths in the leaves of wild-type and independent OsGLIP1-GFP and OsGLIP2-GFP transgenic plants. Plants at booting stage were infected with Xoo. Lesion length was calculated at 14 dpi. Data are shown as means ± SD (n > 10). Student’s t-test, **P < 0.01. (B) Western blot analysis with GFP antibodies confirmed the accumulation of the fusion proteins of OsGLIP1-GFP and OsGLIP2-GFP in the transgenic plants. Rubisco staining was used as loading control. (C) Bacterial growth during 8 days of inoculation in the representative transgenic lines. Data are shown as means ± SD (n = 3). Asterisks indicate significant difference in comparison with the wild-type control (Student’s t-test, *P < 0.05; ** P < 0.01).
(PDF)
The root tips of transgenic plants were incubated in 25% sucrose to induce cell plasmolysis and then observed under confocal microscopy. (A, B) Confocal images of OsGLIP1–GFP (A) and OsGLIP2–GFP (B) after cell plasmolysis. Scale bars = 20 μm. (C) OsGLIP1-GFP root cells without plasmolysis to show the endomembrane systems that link OsGLIP1-GFP labelled vesicles. Scale bar = 20 μm.
(PDF)
Protoplasts were generated from seedlings of transgenic plants expressing OsGLIP1-GFP and OsGLIP2-GFP fusion protein. Note the overlapping of GFP green signals and Nile Red stained red signals, while OsGLIP1-GFP and OsGLIP2-GFP were not associated with chloroplasts. Scale bars = 10 μm.
(PDF)
Two-week-old rice leaves were fed with 100μM MGDG (A) and DGDG (B) for 24 hours and subsequently used for lipid extraction and measurements. Data are shown as means ± SD of four biological replicates. Student’s t-test. *P < 0.05.
(PDF)
(AVI)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.