

New hemophilia treatments aim at releasing the inhibition of tissue factor pathway inhibitor (TFPI) on the extrinsic tenase. Our team has demonstrated that Gla-domainless factor Xa (GD-FXa) can restore coagulation in FVIII- or FIX-deficient plasmas and is unable to trigger coagulation per se. In parallel, a catalytically inactive version of GD-FXa, Andexanet alfa, was proposed as antidote to direct oral anticoagulants targeting FXa. Here we show by molecular modeling that GD-FXa and inactive S195A GD-FXa have similar binding properties to TFPI Kunitz 2 domain. The binding parameters between TFPI and inactive S195A GD-FXa, measured using surface plasmon resonance, were similar to those of plasma-derived GD-FXa. As previously found for GD-FXa, inactive GD-FXa was able to restore thrombin generation in FVIII- and FIX-deficient plasmas as well as in plasmas from hemophiliacs. We, therefore, propose to use catalytically inactive GD-FXa in hemophilia.
The problem of inhibitors to substitution factors used to treat hemophilia makes the search for new treatments highly desirable.1 Releasing TFPI inhibition towards the extrinsic tenase is a strategy of choice, and several approaches have been adopted to achieve this and are in clinical trials.2,3 Indeed, when the intrinsic tenase pathway leading to factor Xa (FXa) activation is blocked due to FVIII or FIX deficiency, a small amount of FXa can be generated by the extrinsic tenase (FVIIa/tissue factor), but it is immediately neutralized by binding of TFPI through its Kunitz 2 (K2) domain while the Kunitz 1 (K1) domain of TFPI subsequently binds to FVIIa, inhibiting the tenase.4,5 Our team has postulated an original anti-TFPI strategy, Gla-domainless factor Xa (GD-FXa), deprived of its membrane binding domain, and we have validated the proof of concept of this approach. Indeed, GD-FXa reconstituted thrombin generation in plasmas from hemophiliacs.6,7 During its production as recombinant protein, GD-FXa was very unstable due to auto-proteolysis.8 Simultaneously, a catalytically inactive version of GD-FXa, Andexanet alfa, was developed and tested as antidote to direct oral anticoagulants targeting FXa.9 This S195A GD-FXa molecule has now passed the phase III procedure and is being evaluated for approval as a new drug by the US Food & Drug Administration. During the phase III study, thrombotic complications were observed and the hypothesis of an interaction with TFPI was raised.10,11 Because of stability problems encountered with wild-type GD-FXa, the inactive S195A GD-FXa mutant appeared easier to handle for biological experiments. We, therefore, decided to study the binding properties of inactive GD-Xa to TFPI both in silico and ex vivo.
We used molecular modeling and molecular dynamics to characterize the binding interface between TFPI K2 domain and either the wild-type or S195A catalytic domains of FXa. Both models were built and submitted to molecular dynamics (see Online Supplementary Appendix). The simulations showed that the S195A mutation did not significantly modify the molecular interface with the K2 domain either structurally or energetically.
For recombinant protein expression, a drosophila S2 expression system was chosen and GD-FXa was produced as a single polypeptide in which the heavy and the light chains were separated by a double furin cleavage site.12 Wild-type GD-FXa could not be purified due to auto-proteolysis. For control experiments, we used plasma-derived GD-FXa that was more stable. In contrast, the S195A mutant was expressed at a higher level in the supernatant of S2 cells and was purified to near homogeneity. SDS-PAGE analysis showed that the protein was 95% pure, and mass spectrometry showed that the protein was properly processed at the furin cleavage site (see Online Supplementary Appendix). Recombinant TFPI alpha was produced in the same system.
Direct binding of S195A GD-FXa and plasma-derived GD-FXa to immobilized TFPI was measured by surface plasmon resonance spectroscopy (SPR). S195A GD-FXa displayed association and dissociation curves that could be fitted to a 1:1 Langmuir binding model (Figure 1). The KD was 1.49±0.09 nM for plasma-derived GD-FXa and 1.99±0.23 nM for S195A GD-FXa. Thus, both proteins were able to bind similarly with high affinity to immobilized TFPI.
Figure 1.

Representative kinetic analysis of the interaction of S195A GD-FXa with immobilized tissue factor pathway inhibitor (TFPI). The GD-FXa samples were injected at the indicated concentrations over immobilized TFPI (1300 RU) in 18 mM Hepes, 135 mM NaCl, 2.5 mM CaCl2, 0.005% surfactant P20, pH 7.35. Binding is expressed in resonance units (RU). Fits are shown as red lines and were obtained by global fitting of the data using a 1:1 Langmuir binding model. The KD was calculated from 3 independent experiments.
We then assessed the effect of inactive GD-FXa in thrombin generation assays in Factor VIII immuno-depleted plasma using Hemker’s method.13 Thrombin generation induced by S195A GD-FXa on factor VIII-deficient plasma showed a dose-dependent effect (Figure 2) that was comparable to that of previously studied plasma-derived GD-FXa.6 The same result was found in FIX immuno-depleted plasma. In addition, we tested plasmas from 4 severe hemophilia A patients and 4 severe hemophilia B patients. We checked that the same results could be obtained with different batches of S195A GD-FXa. Parameters of thrombin generation are summarized in Table 1. In all cases, the effect of S195A GD-FXa did not increase at concentrations above 40 nM and the values were comparable to the relevant positive controls. In the case of hemophilia A with inhibitors, as expected, Factor VIII did not restore thrombin generation unless in 10-fold excess, but S195A GD-FXa did restore thrombin generation perfectly well.
Figure 2.
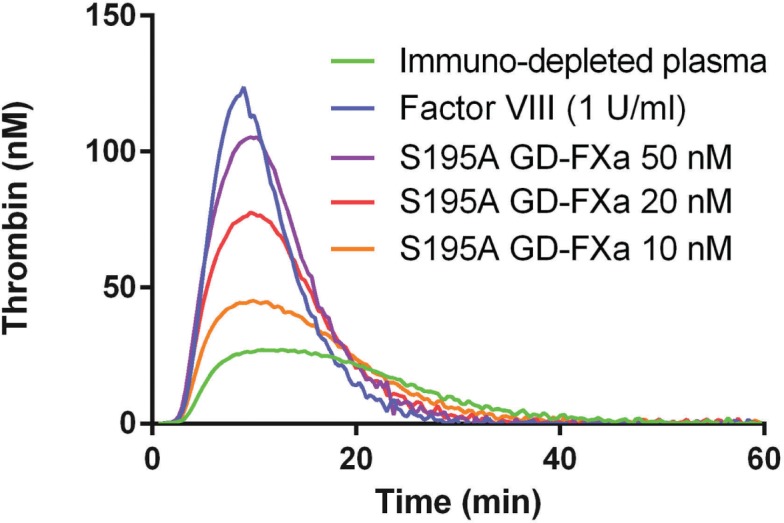
Correction of thrombin generation by S195A GD-FXa in FVIII-immuno-depleted plasma. FVIII-immuno-depleted plasma (Stago) was spiked with S195A GD-FXa at low tissue factor concentration: 1 pM tissue factor and 4 μM phospholipids [PPP-reagent low, (Stago), final concentration]. The positive control was made with recombinant FVIII (Octocog alpha, Bayer).
Table 1.
Parameters of thrombin generation assay after supplementation of hemophilia A and B plasmas and immuno-depleted plasmas by S195A GD-FXa.

We checked that the protein could not replace FXa in FX-deficient plasma and that S195A GD-FXa-induced thrombin generation was dependent on tissue factor. Thus, inactive GD-FXa behaves like wild-type GD-FXa and could be suitable for treatment of hemophilia A and B even in the presence of inhibitors.
We had previously proposed that GD-FXa was a bait to TFPI and provided the proof of concept of this approach using plasma-derived GD-FXa protein.6 Production of GD-FXa as recombinant protein was not successful. In contrast, the catalytically inactive mutant was successfully produced and used as antidote to direct oral anticoagulants,8 but the general view was that a functional active site was required for TFPI binding. Indeed, Broze had shown that diisopropylfluorophosphate-treated FXa lost its ability to bind to TFPI.14 However, the reported procoagulant effects of Andexanet alfa during its phase III trial suggested some TFPI binding capacity.10 Although a catalytically inactive molecule was expected to be totally safe, neutralization of TFPI by Andexanet alfa may have contributed to some of the reported thrombotic events.
We performed molecular dynamics to show that the S195A mutation does not significantly modify the binding interface of the serine protease domain with the K2 domain of TFPI, at both structural and energetic levels, and that the theoretical affinity is unchanged. We then set up expression systems and produced the protein S195A GD-FXa as a single chain in the drosophila S2 cell line with the activation peptide replaced by a double furin cleavage site. The S195A GD-FXa was properly processed by furin, efficiently secreted in the cell culture medium, and could be easily purified. Moreover, SPR experiments showed that inactive GD-FXa was able to bind to TFPI with affinity constants similar to those of the plasma-derived GD-FXa protein, and that it could reconstitute thrombin generation in factor VIII- and IX-deficient plasmas, and also in plasmas from hemophiliacs.
We, thus, propose a new application for a known product, Andexanet alpha or S195A GD-FXa, as a novel anti-hemophilia treatment (MC Dagher et al., 2017, Patent Pending. FR 17 53060). Having no catalytic activity, this protein cannot substitute for FXa even at high doses, and is, therefore, perfectly secure in the context of hemophilia. This potential new hemophilia treatment now requires testing in animal models. The choice of the animal model is crucial as the mouse TFPI sequence has an insertion of six amino acids in the basic C-terminal region involved in prothrombinase inhibition and thus could behave differently from human TFPI.4 A rabbit model of hemophilia would be ideal to prove the efficiency and safety of S195A GD-FXa as the rabbit TFPI sequence differs from that of human TFPI by only one amino acid in the C-terminal basic region.
Supplementary Material
Acknowledgments
The simulations were performed using the Froggy platform of the CIMENT infrastructure, which is supported by the Rhône-Alpes region (GRANT CPER07_13 CIRA) and the Equip@Meso project (reference ANR-10-EQPX-29-01) of the program Investissements d’Avenir supervised by the Agence Nationale pour la Recherche. The authors would like to thank Pierre Girard and Antoine Fortuné for technical support. This work used the platforms of the Grenoble Instruct-ERIC Centre (ISBG; UMS 3518 CNRS-CEA-UJF-EMBL) with support from FRISBI (ANR-10-INSB-05-02) and GRAL (ANR-10-LABX-49-01) within the Grenoble Partnership for Structural Biology (PSB). We thank Isabelle Bally and Jean-Baptiste Reiser for assistance and access to the SPR facility. We thank Elisabetta Boeri Erba and Luca Signor for the technical support to acquire MS spectra. We are indebted to Xavier Brazzoloto who provided the drosophila expression system. We gratefully acknowledge our colleagues of LFB SA Toufik Abache, Alexandre Fontayne and Jean-Luc Plantier for fruitful discussions.
Footnotes
Funding: the project was funded by the Agence Nationale pour la Recherche 13-RPIB-0011
Information on authorship, contributions, and financial & other disclosures was provided by the authors and is available with the online version of this article at www.haematologica.org.
References
- 1.Hartmann J, Croteau SE. 2017 Clinical trials update: Innovations in hemophilia therapy. Am J Hematol. 2016;91(12):1252–1260. [DOI] [PubMed] [Google Scholar]
- 2.Dockal M, Hartmann R, Fries M, et al. Small peptides blocking inhibition of factor Xa and tissue factor-factor VIIa by tissue factor pathway inhibitor (TFPI). J Biol Chem. 2014;289(3):1732–1741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hilden I, Lauritzen B, Sorensen BB, et al. Hemostatic effect of a monoclonal antibody mAb 2021 blocking the interaction between FXa and TFPI in a rabbit hemophilia model. Blood. 2012;119(24):5871–5878. [DOI] [PubMed] [Google Scholar]
- 4.Wood JP, Ellery PE, Maroney SA, Mast AE. Biology of tissue factor pathway inhibitor. Blood. 2014;123(19):2934–2943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Girard TJ, Warren LA, Novotny WF, et al. Functional significance of the Kunitz-type inhibitory domains of lipoprotein-associated coagulation inhibitor. Nature. 1989;338(6215):518–520. [DOI] [PubMed] [Google Scholar]
- 6.Marlu R, Polack B. Gla-domainless factor Xa: molecular bait to bypass a blocked tenase complex. Haematologica. 2012;97(8):1165–1172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Thomas A, Polack B. A new procoagulant molecular decoy for treatment of hemophilia A or B with or without inhibitor. 2014. February 20 US Patent N. 20140050716. [Google Scholar]
- 8.Mertens K, Bertina RM. Pathways in the activation of human coagulation factor X. Biochem J. 1980;185(3):647–658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lu G, DeGuzman FR, Hollenbach SJ, et al. A specific antidote for reversal of anticoagulation by direct and indirect inhibitors of coagulation factor Xa. Nat Med. 2013;19(4):446–451. [DOI] [PubMed] [Google Scholar]
- 10.Connolly SJ, Milling TJ, Jr, Eikelboom JW, et al. Andexanet Alfa for Acute Major Bleeding Associated with Factor Xa Inhibitors. N Engl J Med. 2016;375(12):1131–1141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Mar F, Crowther M, Gold A, et al. 176 Andexanet Alfa: An Investigational Universal Antidote for Reversal of Anticoagulation of Factor Xa Inhibitors in Healthy Human Volunteers. Neurosurgery. 2016;63(Suppl 1):170. [Google Scholar]
- 12.Thomas A, Dagher M-C, Marlu R, Polack B, Plantier J-L, Fontayne A, Abache T. Gla-domainless factor X. 2016. June 23 US Patent N. 20160177286. [Google Scholar]
- 13.Hemker HC, Al Dieri R, De Smedt E, Beguin S. Thrombin generation, a function test of the haemostatic-thrombotic system. Thromb Haemost. 2006;96(5):553–561. [PubMed] [Google Scholar]
- 14.Broze GJ., Jr The rediscovery and isolation of TFPI. J Thromb Haemost. 2003;1(8):1671–1675. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.