

Abstract
Vascular endothelial growth factor A (VEGFA) is a key factor in the regulation of angiogenesis in adipose tissue. Poor vascularization during adipose tissue proliferation causes fibrosis and local inflammation, and is associated with insulin resistance. It is known that 17-beta estradiol (E2) regulates adipose tissue function and VEGFA expression in other tissues; however, the ability of E2 to regulate VEGFA in adipose tissue is currently unknown. In this study, we showed that, in 3T3-L1 cells, E2 and the estrogen receptor 1 (ESR1) agonist PPT induced VEGFA expression, while ESR1 antagonist (MPP), and selective knockdown of ESR1 using siRNA decreased VEGFA and prevented the ability of E2 to modulate its expression. Additionally, we found that E2 and PPT induced the binding of hypoxia inducible factor 1 alpha subunit (HIF1A) in the VEGFA gene promoter. We further found that VEGFA expression was lower in inguinal and gonadal white adipose tissues of ESR1 total body knockout female mice compared to wild type mice. In conclusion, our data provide evidence of an important role for E2/ESR1 in modulating adipose tissue VEGFA, which is potentially important to enhance angiogenesis, reduce inflammation and improve adipose tissue function.
Introduction
Adipose tissue is a highly vascularized tissue and there are data to support ‘healthy’ expansion of adipose tissue occurs when adipose tissue is fully vascularized1,2. Importantly, ‘unhealthy’ expansion of adipose tissue, which occurs in obesity, is associated with relatively nonvascularized adipose tissue leading to inflammation, ectopic fat deposition, and insulin resistance3,4. Vascular Endothelial Growth Factor A (VEGFA) is an angiogenic factor in adipose tissue5; however, the regulation of VEGFA in adipose tissue remains unclear. Interesrtingly, while VEGFA is important for the development of new vessels it is also considered a pro-inflammatory cytokine2,6, potentially indicating a differential effect of VEGFA. When VEGFA is overexpressed in adipose tissue, it results in increased blood vessel number and size, protection against high-fat diet–induced hypoxia and obesity, and improves whole-body insulin sensitivity and glucose tolerance2. On the other hand, there are data to suggest that blocking neovascularization in adipose tissue prevents the development of obesity7,8, further suggesting that the impact of VEGFA on adipose tissue function has yet to be fully understood.
17 beta-estradiol (E2) is a hormone that is critically involved in reproduction as well as energy homeostasis9,10. E2 has been demonstrated to regulate VEGFA in a variety of tissues11, but it is unclear if E2 modulates VEGFA in adipose tissue. Within adipose tissue, E2 can suppress lipid accumulation, adipocyte size, fatty acid uptake, lipogenesis, inflammation and fibrosis12–14. Reductions in circulating E2, as seen in ovariectomized mice and rats, induce insulin resistance and increase susceptibility to high-fat diet induced-metabolic disorders15. Moreover, E2 is hypothesized to be a key feature in the sexual dimorphisms observed in body fat accumulation and adipose tissue expandability. Males accumulate visceral fat, which has been highly correlated to increased cardiovascular risk and tends to be more fibrotic and less vascularized; while females accumulate fat in the subcutaneous depot prior to menopause, and subcutaneous adipose tissue is highly vascularized and less fibrotic. It is further predicted that subcutaneous accumulation of adipose tissue is one of the features that protects pre-menopausal women from diseases associated with obesity and the metabolic syndrome16,17.
E2 binds to its classical receptors- estrogen receptor 1 (ESR1) and estrogen receptor 2 (ESR2) and triggers genomic and non-genomic actions18–20. E2 can also bind to a membrane-bound G-protein-coupled estrogen receptor (GPER)21. The expression and function of estrogen receptors (ERs) are tissue specific18,20,22,23. The classical mechanisms of E2/ERs interactions include ligand binding, receptor dimerization and activation of estrogen response elements (ERES) in target genes leading to activation or repression of genes24,25. In addition, E2/ERs can act rapidly via extracellular and membrane located receptors26. Furthermore, E2 can modulate gene expression by interacting with its receptors, which in turn activate other transcription factors that bind to DNA26,27. As an example, hypoxia-inducible factor 1 alpha subunit (HIF1A) is a transcription factor proposed as a candidate for E2-induced regulation of the VEGFA gene in non-adipose tissues28.
Human subcutaneous and visceral adipose tissue expresses both receptors, ESR1 and ESR229,30. In 3T3-L1 cells and in mouse adipocytes, ESR1 is predominant, but both isoforms are co-expressed in the nucleus of adipocytes30–32. ESR1 is thought to be the predominate ER involved in the regulation of adipose tissue function13. Total body deletion of ESR1 as well as tissue-specific knockdown of ESR1 in male and female mice promotes increased adiposity, fibrosis, insulin resistance and glucose intolerance33–35. On the other hand, total body ESR2 knockout mice do not present with alterations in body composition and glucose homeostasis36,37, suggesting a more predominate role for ESR1 in regulating adipose tissue function, yet the mechanisms by which it does so are currently unknown.
As previously mentioned, there are data to suggest that the VEGFA gene is transcriptionally regulated by direct E2-dependent ESR1 binding and activation38,39; however, it is not known if this regulation occurs in adipose tissue. Therefore, we hypothesized that VEGFA gene is directly regulated by E2-dependent ESR1 binding and activation in adipose tissue, which would in turn improve adipose tissue angiogenesis and promote metabolically healthy adipose tissue. To probe for the involvement of E2/ESR1 in the activation of VEGFA in adipose tissue, we explored the mechanisms by which E2 interacts with ESR1 to regulate VEGFA.
Results
E2 induces VEGFA mRNA and protein in 3T3-L1 cells
We first investigated the impact of E2 across a time-course on VEGFA protein, and to do so we used fully differentiated 3T3-L1 adipocytes which were treated with 0.1, 10 and 100 nM of E2 for 2, 6, 12 and 24 hours. We found that treatments with 10 and 100 nM of E2 for 24 hours were capable of increasing VEGFA protein when compared to control treated cells (Fig. 1a and b). Consistently we also found that Vegfa mRNA was increased by 10 and 100 nM E2 within 24h when compared to control treated cells (Fig. 1c). As both doses of E2 (10 and 100 nM) were effective at increasing VEGFA gene expression and protein, the following experiments were performed using the lower dose, 10 nM for 24 hours, which is a physiological concentration of E2 in vivo 40.
Figure 1.

E2 effects on VEGFA mRNA and protein expression: 3T3-L1 adipocytes were treated for 2, 6, 12 and 24 hours with 0.1, 10 nM and 100 nM of E2. Cells treated only with vehicle were used as control (0E2). VEGFA protein (a and b) and mRNA (c) were analyzed by Western blotting and qPCR, respectively. In panel b, are shown the graph with the results referent to the treatments with 10 and 100nM of E2 for 24 hours with a representative autoradiogram. Full-length blots of the cropped image are presented in Supplementary Figure S1. Data on the graphs represent mean ± SEM of 5 to 8 samples. *P < 0.05 vs 0E2, #P < 0.05 vs 0.1E2, &P < 0.05 vs 0.1 E2 in the same time.
E2 treatment increases ESR1 mRNA expression in 3T3-L1 cells
The mRNA expression of Esr1 and Esr2 was analyzed after treatment with 10 nM of E2 for 24 hours. E2 increased Esr1 expression, but did not change ESR1 protein (Fig. 2a and b), while Esr2 mRNA expression was not affected by the E2 treatment. Interestingly, we determined that 3T3-L1 cells have more Esr1 than Esr2 transcripts (Fig. 2a).
Figure 2.

(a) mRNA expression of Esr1 and Esr2 after treatment with 10 nM of E2 for 24 hours in 3T3-L1 cells. (b) ESR1 total protein expression after treatment with 10 nM of E2 for 24 hours in 3T3-L1 cells. Full-length blots of the cropped image are presented in Supplementary Figure S1. Control cells were treated with vehicle (water). Data on the graphs are mean ± SEM of 3 to 5 samples. &P < 0.05 vs control.
ESR1 agonist PPT enhances VEGFA expression while ESR2 agonist DPN represses it in 3T3-L1 cells
To determine the role of ERs to regulate VEGFA, we first probed the activity of ESR1 and ESR2 using their agonists. ESR1 agonist PPT, independently of E2, induced VEGFA mRNA and protein expressions (Fig. 3a and b). Interestingly, when ESR2 agonist DPN was used, VEGFA mRNA and protein were decreased (Fig. 3c and d), demonstrating an opposite effect of ERs on the regulation of VEGFA in adipocytes.
Figure 3.

Effects of ESR1 agonist (PPT) and ESR2 agonist (DPN) on VEGFA mRNA and protein: 3T3-L1 adipocytes were treated for 24 hours with 10 nM of PPT (a,b) or 10 µmol/L of DPN (c,d). Full-length blots of these cropped images are presented in Supplementary Figure S1. Cells treated with vehicle (0.1% DMSO) were used as control. The representative autoradiograms are shown on top. Data are shown as the mean ± SEM of 6 samples. &P < 0.05 vs control.
To test whether blocking ESR1 activity specifically reduces VEGFA expression in adipocytes, we used the ESR1 antagonist MPP and found that it decreased VEGFA mRNA and protein and prevented the ability of E2 to induce VEGFA expression (Figs 4a and b). On the other hand, the ESR2 antagonist PHTPP did not affect VEGFA, but in the presence of 10 nM of E2, VEGFA protein was increased (Fig. 4c and d). Additionally, we used an siRNA for ESR1 and further demonstrated a reduction in ESR1 mRNA and protein, as well as a reduction in VEFGA mRNA (Fig. 5a,b). Also, we demonstrate following reductions in ESR1 with the siRNA, adding back E2 does not restore VEGFA expression (Fig. 5b).
Figure 4.

Effects of ESR1 antagonist (MPP) and ESR2 antagonist (PHTPP) on VEGFA mRNA and protein: 3T3-L1 adipocytes were treated for 24 hours with 10 µmol/L of MPP (a,b) or 100 nmol/L of PHTPP (c,d), in the absence or presence of 10 nM of E2. Full-length blots of these cropped images are presented in Supplementary Figure S1. Cells treated with vehicle (0.1% DMSO) were used as control. The representative autoradiograms are shown on top. Data are shown as the mean ± SEM of 6 samples. &P < 0.05 vs control.
Figure 5.

Effects of Esr1 silencing on VEGFA mRNA expression. Esr1 gene was knocked-down using siRNA, after 48 hours cells were treated with 10 nM of E2 and collected after 24 hours. Data were presented in mean ± SEM values. &p < 0.05 vs control; $p < 0.05 vs E2 (n = 3 independent rounds of cells).
E2 induces the binding of HIF1A in the VEGFA promoter in 3T3-L1 cells
First, we tested if treatment with E2 induced direct binding of ERs in the ERE sites in the VEGFA promoter, using an Electrophoretic mobility shift assay (EMSA). We observed two DNA/protein complexes corresponding to the binding of ERs in the VEGFA promoter and both receptors, ESR1 and ESR2, are present in these complexes, suggesting ESR1 and ESR2 directly interact with the VEGFA gene promoter in adipocytes (Figure S2). Furthermore, we did not observe an enhancement of the effect following E2 exposure in the intensity of the complex 1 and 2 (Fig. 6a), suggesting that the treatment with E2 did not increase the direct binding of ERs in the VEGFA gene.
Figure 6.
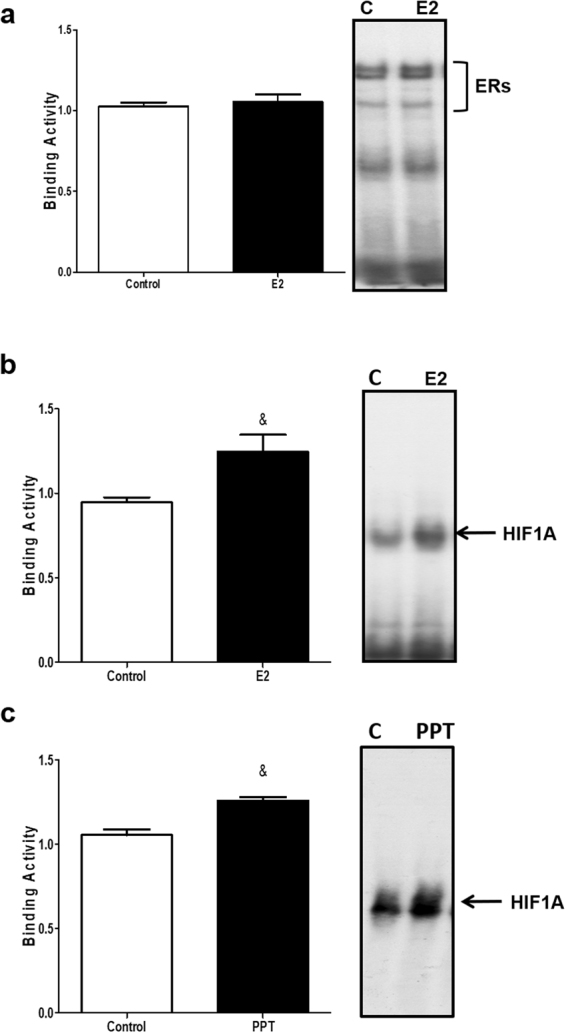
Effect of E2 on ERs and HIF1A binding activity to the Vegfa gene promoter. 3T3-L1 adipocytes were treated with vehicle (0.01% DMSO; Control); 10 nM of E2 or 10 nmol/L of PPT for 24 hours. EMSA was performed as described in Materials and Methods. (a) ERs, biding in VEGFA promoter after 24 hours of E2 treatment. (b) HIF1A biding in VEGFA promoter after 24 hours of E2 treatment; (c) HIF1A binding in VEGFA gene promoter after 24 hours of PPT treatment. Representative images of the experiments are in the right. Results represent three independent experiments. Data are shown as the mean ± SEM of 4 to 6 samples. &P < 0.05 vs control.
We next tested whether HIF1A may be involved in VEGFA upregulation by E2. To do this, we analyzed the binding of HIF1A to the VEGFA promoter (Figure S3). Treatment with 10 nM of E2 for 24 hours increased the DNA–protein complex signal when compared to the control cells (Fig. 6b). In addition, treatment with the ESR1 agonist PPT also increased the DNA-protein complex intensity (Fig. 6c). These results reveal that HIF1A participates in the regulation of VEGFA mediated by E2/ESR1 in adipocytes.
The lack of ESR1 reduces VEGFA expression in different fat depots of female mice
To further explore the interaction between ESR1 and VEGFA expression in vivo, we investigated their expression in inguinal (iWAT) and gonadal (gWAT) adipose tissues from wild-type (WT) and total body ESR1 knockout (ERKO) mice.
Vegfa mRNA expression was lower in the iWAT and gWAT adipose tissues of ERKO compared to WT mice (Fig. 7a). Moreover, iWAT adipose tissue has more Vegfa and Esr1 mRNA expression than gWAT (Fig. 7a and b). We also analyzed the protein by immunofluorescence and quantified fluorescence intensity. In the images, VEGFA is stained green and membrane marker is stained red. VEGFA-positive staining is observed in vessels and adipocytes (Fig. 8a and d). The intensity of VEGFA-positive staining was reduced in iWAT and gWAT adipose tissues from ERKO compared to WT mice (Fig. 8a,b,d and e). Thus, it appears that ESR1 is important in the regulation of VEGFA in both adipose tissue depots.
Figure 7.
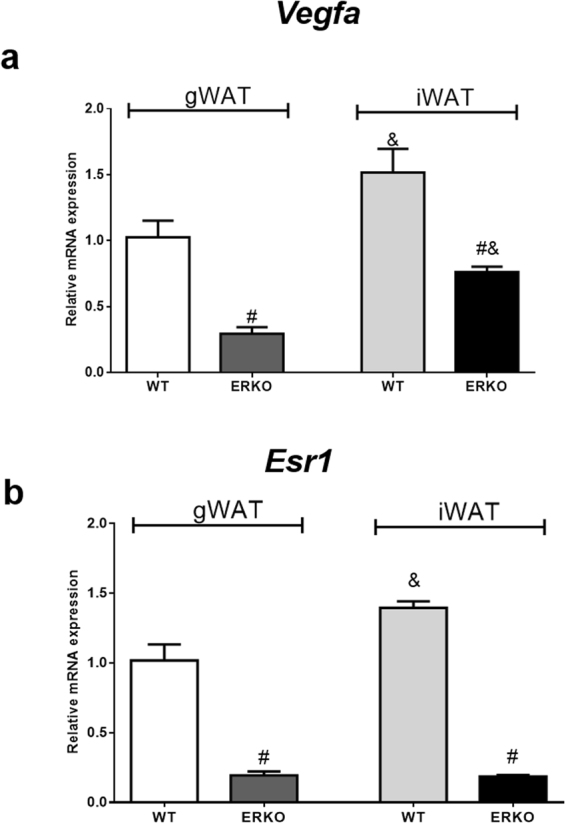
Vegfa (a) and Esr1 (b) mRNA expression in gonadal (gWAT) and in inguinal (iWAT) adipose tissues from WT and ERKO female mice. Data on the graphs are means ± SEM of 4–7 samples, #p < 0.05 vs WT; &p < 0.05 vs gWAT WT.
Figure 8.

VEGFA protein expression in gonadal (gWAT) and inguinal (iWAT) adipose tissue from WT and ERKO female mice. Immunofluorescence staining of VEGFA in gWAT (a) and iWAT (d). Nucleus in blue (DAPI), VEGFA in green, membrane marker in red, and composite in yellow/orange; arrows indicate VEGFA expression in adipocytes and arrowheads in vessels; scale 50 μm; Fluorescence intensity quantification in gWAT (b) iWAT (e). Adipocyte size (surface area- μm2) in gWAT (c) iWAT (f). Data on the graphs are means ± SEM of 4–7 samples, #p < 0.05 vs WT. NC = negative control.
In addition, measurement of adipocyte size was performed to demonstrate the role of ESR1 in adipose tissue morphology. Adipocyte hypertrophy has been demonstrated to impair adipose tissue function by inducing local inflammation, mechanical stress, alterations in metabolism, and insulin resistance41. Consistent with previous findings, adipocytes from ERKO were larger than those from WT mice in iWAT and gWAT (Fig. 8c and f) further supporting the notion that lacking ESR1 induces adipocyte hypertrophy.
To further explore the role of ESR1 and its impact on adipose tissue function, mRNA expression of genes related to VEGFA signaling and markers of inflammation were analyzed. VEGFA binds to and activates VEGFR1 (Flt1) and VEGFR2 (KDR/Flk1). The major pro-angiogenic signal is generated from the ligand-activated KDR, and KDR can be considered a vascular marker. In iWAT, we found neither Flt1 nor Kdr mRNA expression was significantly different in ERKO when compared to WT mice (Fig. 9b and d). However, both Flt1 and Kdr genes were downregulated in gWAT of ERKO mice (Fig. 9a and c).
Figure 9.

Relative mRNA expression of Vegf mRNA receptors, and inflammatory markers in iWAT and gWAT from WT and ERKO female mice. (a and b) Flt1 in gWAT and iWAT respectively; (c and d) Kdr in gWAT and iWAT respectively; (e and f) Tnfa in gWAT and iWAT respectively; (g and h) Il6 in gWAT and iWAT respectively. Data on the graphs are means ± SEM of 3–7 samples, #p < 0.05 vs WT.
We further found that in gWAT, the proinflammatory marker, tumor necrosis factor alpha (Tnfa) mRNA expression was higher in ERKO when compared to WT mice (Fig. 9e); however, in iWAT, Tnfa mRNA expression was lower in ERKO mice compared to WT mice (Fig. 9f). It is important to highlight that the expressions of Tnfa and VEGFA were opposite in the gWAT. No significant differences were observed for Il-6 expression in either iWAT or gWAT (Fig. 9g and h).
Discussion
Reductions in circulating E2 levels are associated with weight gain, increased visceral adiposity, an increased prevalence of obesity, insulin resistance and type 2 Diabetes12,42. It is known that E2 deficiency leads to increased expression and secretion of proinflammatory cytokines12,43. There are data to suggest that VEGFA is a proinflammatory cytokine, and in some tissues, that VEGFA is regulated transcriptionally by E211, but what is not known is the interaction between E2 and VEGFA in adipose tissue. VEGFA is also an important angiogenic factor in adipose tissue and regulates the vascular infrastructure required for healthy expansion of adipose tissue1,2,5. In addition, angiogenesis plays a crucial role in the regulation of adipogenesis44. Here we demonstrate VEGFA is regulated by E2 in 3T3-L1 cells and adipose tissues, and this regulation appears to be through its interaction with ESR1.
In 3T3-L1 cells exposed to E2, both ESR1 and VEGFA were upregulated compared to control cells, and the activation of ESR1 per se by PPT also increased VEGFA expression. On the other hand, the inhibition of ESR1 or the activation of ESR2 decreased VEGFA expression. These data suggest, that the ERs regulate VEGFA in an opposite way. It is known that the decline of E2 levels in postmenopausal women is associated with reductions of ESR1 expression, as well as there is a change in ESR1/ESR2 ratio such that there is more ESR2 relative to ESR145. In addition, the postmenopausal period is characterized by increased visceral adiposity and enhanced metabolic risks associated with obesity46, which may reflect either the reductions in E2 and/or ESR1 expression, and their relative impact on VEGFA.
Previously, it has been demonstrated that VEGFA is regulated positively by both ERs11. In addition, there are data to suggest that in cancer cells, there is an inhibition of VEGFA following treatment with E2 that occurs when both ERs are present, but there is an induction of VEGFA when ERs are expressed alone47. E2 has also been shown to stimulate angiogenesis primarily via ESR1, but again this was not known for adipose tissues48. In our animal model, there was a lower expression of VEGFA in ERKO compared to WT mice in gWAT and iWAT. In addition, we demonstrated a positive correlation between the amount of ESR1 and VEGFA, which differs by adipose tissue depots. Specifically, we found that iWAT expressed more ESR1 and VEGFA mRNA than gWAT. These findings indicate that ESR1 might play an important role in the regulation of VEGFA in white adipose tissues. It has been shown that in obese humans, VEGFA expression and angiogenic potential are similar between subcutaneous and visceral adipose tissues49. Moreover, others showed that in individuals with abdominal obesity, stimulation of subcutaneous adipose tissue angiogenesis promoted adipocytes hyperplasia, thus enhancing the storage capability of adipose tissue, which, in turn, is a protective effect against metabolic disorders3.
Our data further suggests that E2 does not regulate VEGFA in 3T3-L1 cells via direct binding of ERs to the VEGFA gene, which is not consistent with other cells11. However, there are data demonstrating that E2 is also able to augment gene expression through its interaction with ERs, which may in turn bind to other transcriptional factors23, such as HIF1A, to regulate VEGFA48,50,51. Adipose tissue HIF1A activity is influenced by multiple signals, including adipogenesis, insulin, hypoxia, and obesity, but the induction of HIF1A by E2 can induce VEGFA and thereafter promote angiogenesis, a process that is required for adipocyte differentiation and adipose tissue growth52,53. HIF1A is the mediator of physiological and pathophysiological responses to hypoxia54; however, many studies have demonstrated that HIF1A regulates VEGFA expression in various cell types also in situations of normoxia28. There are data to suggest that HIF1A is a direct transcriptional target of ESR1 and that the HIF1A gene bears a canonical ER-binding element that responds to estrogen signaling, and these data demonstrate a direct regulatory link between the ESR1 and HIF1A pathways, but this has been demonstrated in breast cancer55. HIF1A is a labile protein that is rapidly degraded by many different factors, and we have previously reported that indeed E2/ESR1 transcriptionally regulates PHD3 which targets HIF1A for ubiquination and degradation within adipose tissue56. We speculate that E2/ESR1 binds to the ER-binding element within the HIF1A gene to facilitate angiogenesis by then inducing VEGFA in conditions of normoxia; whereas, in conditions of hypoxia, E2/ESR1 binds to the promotor of PHD3 and targets HIF1A for degradation.
Here we observed that E2 through ESR1 activation induced HIF1A binding on the VEGFA promoter. However, the regulation regarding HIF1A in adipose tissue is controversial. It has been demonstrated that in adipose tissue HIF1A promotes fibrosis and inflammation, and E2/ESR1 can negatively regulate HIF1A transcription and degradation56. On the other hand, inhibition of HIF1A in adipose tissue induces obesity and glucose intolerance, due to the impairment of energy expenditure in mice57. Moreover, in the uterus, it has been demonstrated that E2 induced VEGFA expression via membrane ER and activation of PI3K/Akt pathway, which is required for the recruitment of HIF1 to the VEGFA gene promoter28. Indeed, our group has previously observed that treatments with E2 and ER-agonist PPT induced AKT phosphorylation in adipocytes58. Together, these data support the notion that E2/ESR1 can activate HIF1A in adipocytes to induce VEGFA expression. Acute induction of HIF1A binding to VEGFA can increase the vascularization in adipose tissue that in turn prevents the chronic hypoxia that is directly linked to metabolic dysfunction in adipose tissue59.
VEGFA binds to and activates its receptors, Flt1 and KDR. The major pro-angiogenic signal is generated from the ligand-activated KDR that is also considered a marker of vascular density60. The inhibition of VEGFA, which is induced by the inactivation of KDR, occurs during the early phase of high fat diet exposure and leads to weight gain and systemic insulin resistance2. Our findings indicate that Kdr mRNA expression was lower in gWAT from ERKO mice, which could impair VEGFA signaling, leading to reduced formation of new vessels and increased inflammatory markers, as demonstrated by a higher Tnf expression in gWAT of ERKO mice. There is a depot difference in the effect of ESR1 to regulate VEGFA receptors and inflammatory markers, and this could be due to the depot difference in the amount of ESR1. We observed that gWAT has less ESR1 mRNA expression than in iWAT and the gonadal depot was the most affected by inflammation. The health expandability of white adipose tissue has been linked to sexual dimorphisms of adipose tissues and gonadal adipose from female rats is reported to have a higher capacity to expand and has a lower level of inflammation when compared to male adipose tissue. Our data further suggest this may be due to the higher presence of E2 and ESR1 in female gonadal adipose tissue13,17.
VEGFA triggers angiogenesis in adipose tissue, as reflected by enhanced vascular density and improvements in systemic metabolism61. Moreover, it has been demonstrated that VEGFA has a role in the activation and expansion of BAT and “brown fat-like” adipocytes in the subcutaneous adipose tissue a process termed ‘beiging’ and this has been linked to improvements in energy homeostasis and of systemic metabolism2,62. We observed in BAT, Vegfa and Kdr mRNA expression was downregulated in ERKO mice compared to WT (Figure S4), suggesting that E2 may facilitate activation of BAT or beiging of white adipose tissue through ESR1/VEGFA induction.
Our data also demonstrated that visceral and subcutaneous adipocytes from ERKO had a larger cell surface area when compared to fat cells from WT mice. These results indicate that total body loss of ESR1 induces adipocyte hypertrophy, in agreement with results previously published33,63. Adipocyte hypertrophy is linked to inflammation and insulin resistance. Indeed, the larger adipocyte size was associated with a decrease in VEGFA expression, perhaps suggesting a reduction in vascularization in those tissues, and further supporting an important role for ESR1 in mediating vascularity of adipose tissue through increasing VEGFA to facilitate adipose tissue expansion7. Finally, there are data to suggest that ablation of VEGFA in adipose tissue produces higher levels of inflammatory markers and this was accomplished by adipocyte death and reductions in glucose homeostasis and insulin sensitivity64.
In conclusion, our in vitro and in vivo data suggest that E2 regulates VEGFA via ESR1/HIF1A interaction in adipocytes and adipose tissue, which may lead to enhanced angiogenesis and improvements in adipose tissue function.
Methods
Cells 3T3-L1
Mouse 3T3-L1 preadipocytes were obtained from American Type Culture Collection, (ATCC® Number: CL-173TM), and their growth and differentiation were performed as previously described64,65. Briefly, two days after the cell´s confluency, they were induced to differentiation using DMEM high glucose, containing 10% FBS (FBS; Vitrocell Embriolife, Campinas, Brazil), 3-isobutyl- 1-methylxanthine (IBMX) 0.5 mM, dexamethasone (DEXA) 1μM and insulin 10 μg/ml (Sigma–Aldrich; St Louis, MO, USA). After differentiation, which lasted 10–12 days, fully-differentiated cells were ready to be used in the experiments. Twenty-four hours before the treatments, cells were exposed to DMEM without phenol-red and 10% of charcoal striped FBS (Sigma-Aldrich). Adipocytes were treated with different doses of E2 (0, 0.1, 10 and 100) nmol/L (nM) for 24 hours and/or agonist and antagonists of ESR1 and ESR2, followed by their collection and isolation for mRNA and protein analyses. The concentrations of E2, ESR1 and ESR2 agonists and antagonists were established according to previous reports25,32,66–68: we used 10 nM of the ESR1 agonist, PPT: 4,4′,4″-(4-Propyl-[1H]-pyrazole-1,3,5-triyl) trisphenol (Tocris, St. Louis, MO,USA,; ESR1 antagonist, MPP: 1,3-Bis(4-hydroxyphenyl)-4-methyl-5-[4-(piperidinylethoxy)phenol]-1H-pyrazol dihydrochloride (Tocris) (1µmol/L); ESR2 agonist, DPN: 2,3-bis(4-Hydroxyphenyl)-propionitrile) (Tocris) (100 nM); and 100 nM was used of the ESR2 antagonist: PHTPP, 4-[2-Phenyl-5,7-bis(trifluoromethyl) pyrazolo[1,5-a]pyrimidin-3-yl]phenol (Tocris).
RNAi in cell culture
In this series of experiments, we used siRNA to knockdown Esr1 in the presence and absence of estrogens to determine if Esr1 mediates estrogens’ effects on Vegfa. To do this, 3T3-L1 cells were transfected with siRNA targeting murine Esr1 (siGENOME Mouse siEsr1, GE Dharmacon, Lafayette, U.S.). As a control, cells were treated with an unrelated control siRNA (siGENOME non-targeting siRNA, GE Dharmacon), using Lipofectamine RNAi max transfection reagent (Thermo Fisher), according to the manufacturer’s instructions. 48h after siRNA transfection, cells were treated with estrogens and as indicated, and then lysed for RNA extraction.
Animals
Female total body ESR1 knockout (ERKO) and wide type (WT) mice were purchased from Jackson Laboratories (C57BL Maine, USA, Stock No. 004744), and these mice are a functional knockout of ESR1; however, they do express a low level of a truncated form of ESR1 which is the result of alternate splicing of the transcript. Animals were group housed by genotype in a temperature controlled environment on a 12-h light/12-h dark illumination schedule and fed a standard pelleted diet with water provided ad libitum. Nine-month-old mice were used in the experiments. Samples from inguinal and gonadal white adipose tissues were collected and frozen immediately for Real time qPCR, or fixed in 4% paraformaldehyde overnight, then embedded in paraffin wax for immunofluorescence. All mouse experiments were carried out as approved by AAALCA (Association for Assessment and Accreditation for Laboratory Care) at the Pennington Biomed Center at LSU, and tissues were sent to Cedars-Sinai Medical Center (Los Angeles, USA) for processing. All methods were performed in accordance with the relevant guidelines and regulations.
Real time qPCR
Cells or tissues were lysed in TRIzol® reagent (Thermo Fisher Scientific, Massachusetts, USA). RNA was extracted using phase separation reagent (BCP) (MRC®, Ohio, USA) and RNeasy kit (Qiagen®), according to the manufacturer´s instructions. The extracted RNA was measured by NanoDrop 2000c (Thermo Fisher Scientific). Total cDNA was obtained using the High Capacity cDNA Reverse Transcription kit (Thermo Fisher Scientific). Real-time qPCR was performed in duplicate by using Taqman Universal Mastermix II (Thermo Fisher Scientific). Taqman specific primers and QuantStudio 12K Flex Real-Time PCR System machine: Gapdh- Mm99999915_g1; Esr1- Mm00433149_m1; Esr2- Mm00599821_m1; Kdr- Mm01222421_m1; Ftl1- Mm00438980_m1; Tnf- Mm00443258_m1; Vegfa- 00437306_m1; Il6- Mm00446190_m1. Relative expression values were calculated from the threshold cycle (Ct) following the 2−∆∆CT method, using Gapdh as the reference gene.
Western blotting
Cells were homogenized in RIPA buffer. Protein concentration was determined by Bradford method (Bio-Rad Laboratories, CA). Proteins (25 μg) from cell lysates were separated by electrophoresis, and electrotransferred to nitrocellulose membrane Hybond-ECL (Amersham, Buckinghahmshire, UK). Later, membranes were stained with Ponceau S to be used as the loading control. Membranes were then blocked in TBS-T containing 5% non-fat powdered milk, followed by overnight incubation with primary antibodies (ESR1 Sc-542–1:1000; VEGFA Sc152–1:500; Santa Cruz, Dallas, TX, USA). Blot´s intensity was quantified by densitometry (ImageScanner III, GE Healthcare, Uppsala, Sweden). Results were expressed as arbitrary units in comparison to controls.
Electrophoretic Mobility Shift Assay (EMSA)
Nuclear protein extraction was conducted according to (Xing et al.)69. Briefly, cells were washed with Phosphate buffered saline (PBS; NaCl 137 mM; KCL 2.68 mm, KH2PO4 1.27 mm, Na2HPO4 8.06 mM) and centrifuged 2000 g and 4 °C, for 5 min. The pellet was resuspended in lyses buffer (HEPES-KOH pH 7.9, 10 mM; MgCl2 1.5 mM; KCl 10 mM; DTT 0.5 mM; PMSF 0.2 mM; leupeptin 5 µg/mL; aprotinin 15 µg/mL), incubated on ice for 10 min and 10% of Nonidet P-40 was added. After the centrifugation at 12000 g and 4 °C, for 20 min, the pellet was resuspended in 50 μl of extraction buffer (HEPES-KOH pH 7.9, 20 mM; MgCl2 1.5 mM; EDTA 0.5 mM; DTT 0.2 mM; NaCl 420 mM; Glicerol 25%; PMSF 0.2 mM; leupeptin 5 µg/ml; aprotinin 15 µg/ml) and incubated on ice for 20 min.
To determine ER binding in the VEGFA gene, we used a wild-type double-stranded oligonucleotide containing the consensus region for the receptors in the 3′UTR of VEGFA gene 5′TCTACAAAAGCACCCCGCCCCTCTGG 3′70. Moreover, we used two mutated probes: 5′-TCTACAAttGCACCCaGCCCCTCTGG-3′; 5′-TCTACAAAAGCACCtCGCttCTCTGG-3′. To determine HIF1a binding in the VEGFA gene, we used the probe 5′-TGCATACGTGGGTTTCCACAG-353.
Probes were end-labeled using T4Polynucleotide Kinase (Invitrogen Life Technologies, California, USA) and [γ-32P] ATP (PerkinElmer Life, Massachusetts, USA). The EMSA experiments were carried out according to those previously described35. The competitive binding assay was performed by adding unlabeled probes at 50-fold excess. To verify the presence of our protein of interest in the DNA-protein complex, specified antibodies were added in the final mixtures (Anti-HIF1A mouse monoclonal – Abcam 16066; Anti-ESR1 Rabbit polyclonal - Santa Cruz 542; Anti-ESR2 Goat polyclonal - Santa Cruz 6821 X; Supplementary Figures 2 and 3). The reaction mixture was then electrophoresed on a 4% nondenaturing polyacrylamide, after which gels were dried and exposed overnight to an X-ray film. The blots corresponded to each treatment were analyzed by scanner densitometry and the results of the binding activity were expressed as arbitrary units.
Immunofluorescence
In this series, we verified protein expression and localization of VEGFA in adipose tissues from ERKO and WT mice. Adipose tissue was collected from different depots and embedded in paraffin wax and sectioned. The sections were incubated with primary antibody anti-VEGFA (1:100- sc-152) overnight at 4 °C in a moist chamber. Next, sections were incubated in a secondary antibody (Goat to rabbit- FITC, Abcam® ab6717) for 1h. Following which, sections were incubated with the membrane marker Wheat Germ Agglutinin (Life Technologies) for 30 min and Dapi (1:200, Abcam). Slides were mounted with Vectashield (Vector®, H1000). Images were acquired using Keyence BZ-9000 microscope, and analyzed by BZ-9000 analyzed software. Three sections from each mouse were stained, five pictures of each section were taken, and the fluorescence intensity was quantified using the ImageJ software (National Institutes of Health Bethesda, MD USA). The measurement of adipose cell surface area was done in the same pictures using the plugin Adiposoft (ImageJ software). Results were expressed as arbitrary units in comparison to controls.
Data analysis
All data are presented as mean ± standard error of the mean (SEM). Data was tested for normality (Levene Statistic). Statistical significance between groups was determined by one-way ANOVA and Bonferroni as post-hoc-test. Two-tailed paired Student´s test was used as appropriated. P values < 0.05 were considered statistically significant. Statistical analyses and graphs were generated using GraphPad Prism 6 software (GraphPad software, San Diego, CA, USA).
Electronic supplementary material
Acknowledgements
This material was based upon work supported by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), under Grants: 15/13652-1, 13/03343-6, 12/04831-1.
Author Contributions
Experiments in vitro were performed by L.A.F., R.S.C. and H.S.F. Ex vivo experiments were performed by L.A.F., R.S.S. and A.P.F. Statistics were performed by L.A.F., L.A.F., R.S.C., R.S.S., A.P.F., U.F.M. and D.J.C. designed and discussed the experiments and results. L.A.F. and D.J.C. wrote the main manuscript tex. U.F.M. and D.J.C. revised the manuscript.
Competing Interests
The authors declare that they have no competing interests.
Footnotes
U. F. Machado and D. J. Clegg jointly supervised this work.
Electronic supplementary material
Supplementary information accompanies this paper at 10.1038/s41598-017-16686-7.
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Cao Y. Adipose tissue angiogenesis as a therapeutic target for obesity and metabolic diseases. Nat Rev Drug Discov. 2010;9:107–115. doi: 10.1038/nrd3055. [DOI] [PubMed] [Google Scholar]
- 2.Sun K, et al. Dichotomous effects of VEGF-A on adipose tissue dysfunction. Proc Natl Acad Sci USA. 2012;109:5874–5879. doi: 10.1073/pnas.1200447109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kim JY, et al. Obesity-associated improvements in metabolic profile through expansion of adipose tissue. J Clin Invest. 2007;117:2621–2637. doi: 10.1172/JCI31021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Gustafson B, Hedjazifar S, Gogg S, Hammarstedt A, Smith U. Insulin resistance and impaired adipogenesis. Trends Endocrinol Metab. 2015;26:193–200. doi: 10.1016/j.tem.2015.01.006. [DOI] [PubMed] [Google Scholar]
- 5.Zhang QX, et al. Vascular endothelial growth factor is the major angiogenic factor in omentum: mechanism of the omentum-mediated angiogenesis. J Surg Res. 1997;67:147–154. doi: 10.1006/jsre.1996.4983. [DOI] [PubMed] [Google Scholar]
- 6.Reinders ME, et al. Proinflammatory functions of vascular endothelial growth factor in alloimmunity. J Clin Invest. 2003;112:1655–1665. doi: 10.1172/JCI17712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Rupnick MA, et al. Adipose tissue mass can be regulated through the vasculature. Proc Natl Acad Sci USA. 2002;99:10730–10735. doi: 10.1073/pnas.162349799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Daquinag AC, Zhang Y, Kolonin MG. Vascular targeting of adipose tissue as an anti-obesity approach. Trends Pharmacol Sci. 2011;32(5):300–307. doi: 10.1016/j.tips.2011.01.004. [DOI] [PubMed] [Google Scholar]
- 9.Stubbins RE, Holcomb VB, Hong J, Núñez NP. Estrogen modulates abdominal adiposity and protects female mice from obesity and impaired glucose tolerance. Eur J Nutr. 2012;51:861–870. doi: 10.1007/s00394-011-0266-4. [DOI] [PubMed] [Google Scholar]
- 10.Tchernof A, et al. Ovarian hormone status and abdominal visceral adipose tissue metabolism. J Clin Endocrinol Metab. 2004;89:425–3430. doi: 10.1210/jc.2003-031561. [DOI] [PubMed] [Google Scholar]
- 11.Applanat MP, Buteau-Lozano H, Herve MA, Corpet A. Vascular endothelial growth factor is a target gene for estrogen receptor and contributes to breast cancer progression. Adv Exp Med Biol. 2008;617:437–444. doi: 10.1007/978-0-387-69080-3_42. [DOI] [PubMed] [Google Scholar]
- 12.Clegg DJ, Brown LM, Woods SC, Benoit SC. Gonadal hormones determine sensitivity to central leptin and insulin. Diabetes. 2006;55:978–987. doi: 10.2337/diabetes.55.04.06.db05-1339. [DOI] [PubMed] [Google Scholar]
- 13.Davis KE, et al. The sexually dimorphic role of adipose and adipocyte estrogen receptors in modulating adipose tissue expansion, inflammation, and fibrosis. Mol Metab. 2013;2:227–242. doi: 10.1016/j.molmet.2013.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ribas V, et al. Impaired oxidative metabolism and inflammation are associated with insulin resistance in ERalpha-deficient mice. Am J Physiol Endocrinol Metab. 2010;298:E304–319. doi: 10.1152/ajpendo.00504.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Campbell SE, Febbraio MA. Effect of the ovarian hormones on GLUT4 expression and contraction-stimulated glucose uptake. Am J Physiol Endocrinol Metab. 2002;282:E1139–1146. doi: 10.1152/ajpendo.00184.2001. [DOI] [PubMed] [Google Scholar]
- 16.Davis S, et al. Isolation of angiopoietin-1, a ligand for the TIE2 receptor, by secretion-trap expression cloning. Cell. 1996;87:1161–1169. doi: 10.1016/S0092-8674(00)81812-7. [DOI] [PubMed] [Google Scholar]
- 17.Palmer BF, Clegg DJ. The sexual dimorphism of obesity. Mol Cell Endocrinol. 2015;402:113–119. doi: 10.1016/j.mce.2014.11.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Grandien K. Determination of transcription start sites in the human estrogen receptor gene and identification of a novel, tissue-specific, estrogen receptor-mRNA isoform. Mol Cell Endocrinol. 1996;116:207–212. doi: 10.1016/0303-7207(95)03716-0. [DOI] [PubMed] [Google Scholar]
- 19.Kuiper GG, Enmark E, Pelto-Huikko M, Nilsson S, Gustafsson JA. Cloning of a novel receptor expressed in rat prostate and ovary. Proc Natl Acad Sci USA. 1996;93:5925–5930. doi: 10.1073/pnas.93.12.5925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Saunders PT, Maguire SM, Gaughan J, Millar MR. Expression of oestrogen receptor beta (ER beta) in multiple rat tissues visualised by immunohistochemistry. J Endocrinol. 1997;154:R13–16. doi: 10.1677/joe.0.154R013. [DOI] [PubMed] [Google Scholar]
- 21.Revankar CM, Cimino DF, Sklar LA, Arterburn JB, Prossnitz ER. A transmembrane intracellular estrogen receptor mediates rapid cell signaling. Science. 2005;307:1625–1630. doi: 10.1126/science.1106943. [DOI] [PubMed] [Google Scholar]
- 22.Kuiper GG, et al. Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors alpha and beta. Endocrinology. 1997;138:863–870. doi: 10.1210/endo.138.3.4979. [DOI] [PubMed] [Google Scholar]
- 23.Nilsson S, et al. Mechanisms of estrogen action. Physiol Rev. 2001;81:1535–1565. doi: 10.1152/physrev.2001.81.4.1535. [DOI] [PubMed] [Google Scholar]
- 24.Mauvais-Jarvis F, Clegg DJ, Hevener AL. The role of estrogens in control of energy balance and glucose homeostasis. Endocr Rev. 2013;34:309–338. doi: 10.1210/er.2012-1055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Helguero LA, Faulds MH, Gustafsson JA, Haldosén LA. Estrogen receptors alfa (ERalpha) and beta (ERbeta) differentially regulate proliferation and apoptosis of the normal murine mammary epithelial cell line HC11. Oncogene. 2005;24:6605–6616. doi: 10.1038/sj.onc.1208807. [DOI] [PubMed] [Google Scholar]
- 26.Hammes SR, Levin ER. Extranuclear steroid receptors: nature and actions. Endocr Rev. 2007;28:726–741. doi: 10.1210/er.2007-0022. [DOI] [PubMed] [Google Scholar]
- 27.Nilsson S, Gustafsson JA. Estrogen receptor transcription and transactivation: Basic aspects of estrogen action. Breast Cancer Res. 2000;2:360–366. doi: 10.1186/bcr81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kazi AA, Koos RD. Estrogen-induced activation of hypoxia-inducible factor-1alpha, vascular endothelial growth factor expression, and edema in the uterus are mediated by the phosphatidylinositol 3-kinase/Akt pathway. Endocrinology. 2007;148:2363–2374. doi: 10.1210/en.2006-1394. [DOI] [PubMed] [Google Scholar]
- 29.Pedersen SB, et al. Identification of oestrogen receptors and oestrogen receptor mRNA in human adipose tissue. Eur J Clin Invest. 1996;26:262–269. doi: 10.1046/j.1365-2362.1996.145278.x. [DOI] [PubMed] [Google Scholar]
- 30.Dieudonné MN, Leneveu MC, Giudicelli Y, Pecquery R. Evidence for functional estrogen receptors alpha and beta in human adipose cells: regional specificities and regulation by estrogens. Am J Physiol Cell Physiol. 2004;286:C655–661. doi: 10.1152/ajpcell.00321.2003. [DOI] [PubMed] [Google Scholar]
- 31.Barros RP, Gabbi C, Morani A, Warner M, Gustafsson JA. Participation of ERalpha and ERbeta in glucose homeostasis in skeletal muscle and white adipose tissue. Am J Physiol Endocrinol Metab. 2009;297:E124–133. doi: 10.1152/ajpendo.00189.2009. [DOI] [PubMed] [Google Scholar]
- 32.Campello RS, et al. Estrogen Receptor 1 Agonist PPT Stimulates Slc2a4 Gene Expression and Improves Insulin-Induced Glucose Uptake in Adipocytes. Curr Top Med Chem. 2012;12:2059–2069. doi: 10.2174/156802612804910197. [DOI] [PubMed] [Google Scholar]
- 33.Heine PA, Taylor JA, Iwamoto GA, Lubahn DB, Cooke PS. Increased adipose tissue in male and female estrogen receptor-alpha knockout mice. Proc Natl Acad Sci USA. 2000;97:12729–12734. doi: 10.1073/pnas.97.23.12729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Barros RP, Machado UF, Warner M, Gustafsson JA. Muscle GLUT4 regulation by estrogen receptors ERbeta and ERalpha. Proc Natl Acad Sci USA. 2006;103:1605–1608. doi: 10.1073/pnas.0510391103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Takeda K, et al. Progressive development of insulin resistance phenotype in male mice with complete aromatase (CYP19) deficiency. J Endocrinol. 2003;176:237–246. doi: 10.1677/joe.0.1760237. [DOI] [PubMed] [Google Scholar]
- 36.Jones ME, et al. Aromatase-deficient (ArKO) mice have a phenotype of increased adiposity. Proc Natl Acad Sci USA. 2000;97:12735–12740. doi: 10.1073/pnas.97.23.12735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Barros RP, Machado UF, Gustafsson JA. Estrogen receptors: new players in diabetes mellitus. Trends Mol Med. 2006;12:425–431. doi: 10.1016/j.molmed.2006.07.004. [DOI] [PubMed] [Google Scholar]
- 38.Sengupta K, Banerjee S, Saxena N, Banerjee SK. Estradiol-induced vascular endothelial growth factor-A expression in breast tumor cells is biphasic and regulated by estrogen receptor-alpha dependent pathway. Int J Oncol. 2003;22:609–614. [PubMed] [Google Scholar]
- 39.Stoner M, et al. Estrogen regulation of vascular endothelial growth factor gene expression in ZR-75 breast cancer cells through interaction of estrogen receptor alpha and SP proteins. Oncogene. 2004;23:1052–1063. doi: 10.1038/sj.onc.1207201. [DOI] [PubMed] [Google Scholar]
- 40.Ma YL, et al. The effects of different doses of estradiol (E2) on cerebral ischemia in an in vitro model of oxygen and glucose deprivation and reperfusion and in a rat model of middle carotid artery occlusion. BMC Neurosci. 2013;14:118. doi: 10.1186/1471-2202-14-118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Henninger AM, Eliasson B, Jenndahl LE, Hammarstedt A. Adipocyte hypertrophy, inflammation and fibrosis characterize subcutaneous adipose tissue of healthy, non-obese subjects predisposed to type 2 diabetes. PLoS One. 2014;9:105262. doi: 10.1371/journal.pone.0105262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Barros RP, Gustafsson J. Estrogen receptors and the metabolic network. Cell Metab. 2011;14:289–299. doi: 10.1016/j.cmet.2011.08.005. [DOI] [PubMed] [Google Scholar]
- 43.Pfeilschifter J, Köditz R, Pfohl M, Schatz H. Changes in proinflammatory cytokine activity after menopause. Endocr Rev. 2002;23:90–119. doi: 10.1210/edrv.23.1.0456. [DOI] [PubMed] [Google Scholar]
- 44.Xue Y, et al. Hypoxia-independent angiogenesis in adipose tissues during cold acclimation. Cell Metab. 2009;9:99–109. doi: 10.1016/j.cmet.2008.11.009. [DOI] [PubMed] [Google Scholar]
- 45.Park YM, Erickson C, Bessesen D, Van Pelt RE, Cox-York K. Age- and menopause-related differences in subcutaneous adipose tissue estrogen receptor mRNA expression. Steroids. 2017;121:17–21. doi: 10.1016/j.steroids.2017.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Broekmans FJ, Soules MR, Fauser BC. Ovarian aging: mechanisms and clinical consequences. Endocr Rev. 2009;30:465–493. doi: 10.1210/er.2009-0006. [DOI] [PubMed] [Google Scholar]
- 47.Buteau-Lozano H, Ancelin M, Lardeux B, Milanini J, Perrot-Applanat M. Transcriptional regulation of vascular endothelial growth factor by estradiol and tamoxifen in breast cancer cells: a complex interplay between estrogen receptors alpha and beta. Cancer Res. 2002;62:4977–4984. [PubMed] [Google Scholar]
- 48.Kazi AA, Jones JM, Koos RD. Chromatin immunoprecipitation analysis of gene expression in the rat uterus in vivo: estrogen-induced recruitment of both estrogen receptor alpha and hypoxia-inducible factor 1 to the vascular endothelial growth factor promoter. Mol Endocrinol. 2005;19:2006–2019. doi: 10.1210/me.2004-0388. [DOI] [PubMed] [Google Scholar]
- 49.Ledoux. S, et al. Angiogenesis associated with visceral and subcutaneous adipose tissue in severe human obesity. Diabetes. 2008;57:3247–3257. doi: 10.2337/db07-1812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Forsythe JA, et al. Activation of vascular endothelial growth factor gene transcription by hypoxia-inducible factor 1. Mol Cell Biol. 1996;16:4604–4613. doi: 10.1128/MCB.16.9.4604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.George AL, Rajoria S, Suriano R, Mittleman A, Tiwari RK. Hypoxia and estrogen are functionally equivalent in breast cancer-endothelial cell interdependence. Mol Cancer. 2012;11:80. doi: 10.1186/1476-4598-11-80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Cao Y. Angiogenesis modulates adipogenesis and obesity. J Clin Invest. 2007;117:2362–2368. doi: 10.1172/JCI32239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Hausman GJ, Richardson RL. Adipose tissue angiogenesis. J Anim Sci. 2004;82:925–934. doi: 10.2527/2004.823925x. [DOI] [PubMed] [Google Scholar]
- 54.Schofield CJ, Ratcliffe P. J. Oxygen sensing by HIF hydroxylases. Nat Rev Mol Cell Biol. 2004;5:343–354. doi: 10.1038/nrm1366. [DOI] [PubMed] [Google Scholar]
- 55.Yang J, et al. Estrogen receptor-α directly regulates the hypoxia-inducible fator 1 pathway associated with antiestrogen response in breast cancer. Proc Natl Acad Sci USA. 2015;112:15172–7. doi: 10.1073/pnas.1422015112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Kim M, et al. ERα upregulates Phd3 to ameliorate HIF-1 induced fibrosis and inflammation in adipose tissue. Mol Metab. 2014;3:642–651. doi: 10.1016/j.molmet.2014.05.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Zhang X, et al. Adipose tissue-specific inhibition of hypoxia-inducible factor 1{alpha} induces obesity and glucose intolerance by impeding energy expenditure in mice. J Biol Chem. 2010;285:32869–32877. doi: 10.1074/jbc.M110.135509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Campello R, et al. Estradiol-induced regulation of GLUT4 in 3T3-L1 cells: involvement of ESR1 and AKT activation. J Mol Endocrinol. 2017;59:257–268. doi: 10.1530/JME-17-0041. [DOI] [PubMed] [Google Scholar]
- 59.Corvera S, Gealekman O. Adipose tissue angiogenesis: impact on obesity and type-2 diabetes. Biochim Biophys Acta. 2014;1842:463–472. doi: 10.1016/j.bbadis.2013.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Terman BI, et al. Identification of the KDR tyrosine kinase as a receptor for vascular endothelial cell growth factor. Biochem Biophys Res Commun. 1992;187:1579–1586. doi: 10.1016/0006-291X(92)90483-2. [DOI] [PubMed] [Google Scholar]
- 61.Elias I, et al. Adipose tissue overexpression of vascular endothelial growth factor protects against diet-induced obesity and insulin resistance. Diabetes. 2012;61:1801–1813. doi: 10.2337/db11-0832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Sun K, et al. Brown adipose tissue derived VEGF-A modulates cold tolerance and energy expenditure. Mol Metab. 2014;3:474–483. doi: 10.1016/j.molmet.2014.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Pedram A, Razandi M, Blumberg B, Levin ER. Membrane and nuclear estrogen receptor α collaborate to suppress adipogenesis but not triglyceride content. FASEB J. 2016;30:230–240. doi: 10.1096/fj.15-274878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Sung HK, et al. Adipose Vascular Endothelial Growth Factor Regulates Metabolic Homeostasis through Angiogenesis. Cell Metab. 2013;17:61–72. doi: 10.1016/j.cmet.2012.12.010. [DOI] [PubMed] [Google Scholar]
- 65.Chamberlain LH. Inhibition of isoprenoid biosynthesis causes insulin resistance in 3T3-L1 adipocytes. FEBS Lett. 2001;507:357–361. doi: 10.1016/S0014-5793(01)03007-1. [DOI] [PubMed] [Google Scholar]
- 66.Furuya DT, Poletto AC, Freitas HS, Machado UF. Inhibition of cannabinoid CB1 receptor upregulates Slc2a4 expression via nuclear factor-κB and sterol regulatory element-binding protein-1 in adipocytes. J Mol Endocrinol. 2012;49:97–106. doi: 10.1530/JME-12-0037. [DOI] [PubMed] [Google Scholar]
- 67.Dos Santos EG, et al. Rapid nongenomic E2 effects on p42/p44 MAPK, activator protein-1, and cAMP response element binding protein in rat white adipocytes. Endocrinology. 2002;143:930–940. doi: 10.1210/endo.143.3.8678. [DOI] [PubMed] [Google Scholar]
- 68.Muraki K, Okuya S, Tanizawa Y. Estrogen receptor alpha regulates insulin sensitivity through IRS-1 tyrosine phosphorylation in mature 3T3-L1 adipocytes. Endocr J. 2006;53:841–851. doi: 10.1507/endocrj.K06-005. [DOI] [PubMed] [Google Scholar]
- 69.Xing H, et al. TNF alpha-mediated inhibition and reversal of adipocyte differentiation is accompanied by suppressed expression of PPARgamma without effects on Pref-1 expression. Endocrinology. 1997;138(7):2776–2783. doi: 10.1210/endo.138.7.5242. [DOI] [PubMed] [Google Scholar]
- 70.Hyder SM, Nawaz Z, Chiappetta C, Stancel GM. Identification of functional estrogen response elements in the gene coding for the potent angiogenic factor vascular endothelial growth factor. Cancer Res. 2000;60:3183–3190. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.