

Abstract
Human saphenous vein (HSV) is harvested and prepared prior to implantation as an arterial bypass graft. Injury and the response to injury from surgical harvest and preparation trigger cascades of molecular events and contribute to graft remodeling and intimal hyperplasia. Apoptosis is an early response after implantation that contributes the development of neointimal lesions. Here, we showed that surgical harvest and preparation of HSV leads to vasomotor dysfunction, increased apoptosis and downregulation of the phosphorylation of the anti-apoptotic protein, Niban. A model of subfailure overstretch injury in rat aorta (RA) was used to demonstrate impaired vasomotor function, increased extracellular ATP (eATP) release, and increased apoptosis following pathological vascular injury. The subfailure overstretch injury was associated with activation of p38 MAPK stress pathway and decreases in the phosphorylation of the anti-apoptotic protein Niban. Treatment of RA after overstretch injury with antagonists to purinergic P2X7 receptor (P2X7R) antagonists or P2X7R/pannexin (PanX1) complex, but not PanX1 alone, restored vasomotor function. Inhibitors to P2X7R and PanX1 reduced stretch-induced eATP release. P2X7R/PanX1 antagonism led to decrease in p38 MAPK phosphorylation, restoration of Niban phosphorylation and increases in the phosphorylation of the anti-apoptotic protein Akt in RA and reduced TNFα-stimulated caspase 3/7 activity in cultured rat vascular smooth muscle cells. In conclusion, inhibition of P2X7R after overstretch injury restored vasomotor function and inhibited apoptosis. Treatment with P2X7R/PanX1 complex inhibitors after harvest and preparation injury of blood vessels used for bypass conduits may prevent the subsequent response to injury that lead to apoptosis and represents a novel therapeutic approach to prevent graft failure.
Keywords: P2X7 receptor, Subfailure overstretch, Saphenous vein graft, Vascular injury, ATP, Apoptosis, Purinergic signaling
Introduction
Autologous human saphenous vein grafts (HSV) are commonly used for infrainguinal and coronary revascularization procedures. Injury during graft harvest and preparation contribute to the development of intimal hyperplasia, the primary cause of vein graft failure. The outcome from these procedures remains limited by high rates of vein graft failure, 39% at 12–18 months after peripheral vascular bypass [1] and 45% at 12–18 months after coronary artery bypass grafting (CABG) [2]. Various methods have been developed for the surgical harvest of saphenous veins for CABG with endoscopic harvest remains most commonly performed which leads to traction on the vein and injury to the endothelium and media [3–5]. After harvest, the veins are further “prepared” prior to implantation by pressure distension [6], off-label marking with a surgical skin marker [7], and storing in a non-buffered solution [8], resulting in physiologic dysfunction, increased oxidative stress [9] and increased expression of genes that involved in inflammation and apoptosis [10–12]. Thus, minimizing operative injury to the conduit at the time of harvest and preparation improves conduit function [13] and may prevent vein graft failure.
Our previous studies have shown that brilliant blue FCF (BB FCF), a P2X7R antagonists, ameliorated injury in HSV and porcine saphenous veins (PSV), suggesting an involvement of P2X7R in vascular injury [7, 14]. We have recently developed a rat aorta (RA) model of subfailure overstretch injury [15]. This stretch injury recapitulates the haptic endpoint of traction during endoscopic harvest of HSV. Subfailure overstretch injury to RA leads to impaired physiologic function that can be partially restored with A438079 and oxidized ATP, inhibitors to the purinergic P2X7 receptor (P2X7R), further implicating a role of P2X7R signaling in iatrogenic stretch injury to the conduits [15].
Pathophysiologic ATP release by damaged cells has been demonstrated during astrocytes swelling and stretching [16], urinary bladder distention [17], alveolar cell stretching [18], and T cells hypertonic challenge [19]. This release is mediated by the P2X7R and, depending on cell types, the pannexin channel [20, 21]. Once released, high extracellular ATP (eATP) further amplifies P2X7R activation and ATP release in neighboring cells. In addition, elevation of P2X7R expression has been linked to injury [22, 23]. The functional sequelae of P2X7R activation includes formation of large membrane pores, influx of calcium and activation of caspases, and ultimately apoptosis [24]. Antagonists to the P2X7R have been shown to ameliorates stretch injury of the spinal cord [25] and modulate responses in murine models of various inflammatory and neurological disorders [26].
In this study, we hypothesized that overstretch injury during harvest and preparation of HSV leads to release of ATP and activation of P2X7R, which potentiates the injury response. We first determined the impact of surgical harvest and preparation on vasomotor function and apoptosis in HSV, and then confirmed the role of ATP release, P2X7R activation in stretch-induced apoptosis in the RA model of subfailure overstretch injury. Our data provide new mechanistic insights into the purinergic signaling that regulates apoptotic responses to vascular overstretch injury.
Materials and methods
All chemicals were purchased from Sigma Chemical Co. (St. Louis, MO) unless otherwise specified.
Procurement of human saphenous vein
HSV was obtained after approval from the Institutional Review Boards of Vanderbilt University Medical Center from patients undergoing coronary artery bypass grafting procedures. Method of vein harvest and graft preparation [including hydrostatic distention, marking with a surgical skin marker, and placement in storage solution (normal saline or PlasmaLyte)] was at the discretion of the surgical team. For each HSV (n = 18), paired segments were collected (1) immediately following surgical harvest (“unprepared” vein samples, UP) and (2) after graft preparation (“after preparation” vein samples, AP) from the same patient. Each vein segment was transported immediately to laboratory in heparinized PlasmaLyte (HP; 10 unit heparin/mL PlasmaLyte) for experimentation, formalin fixation, or frozen within 30 min of collection. Areas showing visible signs of injury were not used in experimentation.
Procurement of rat aortae
Aorta (RA) was collected from female, 250-300 g, Sprague Dawley rats. Animal procedures followed study protocols approved by the Vanderbilt Institutional Animal Care and Use Committee and adhered to National Institute of Health guidelines for care and use of laboratory animals. Immediately after euthanasia, the abdominal RA was isolated via an incision along the mid-abdomen, placed in HP and transported to the laboratory for immediate testing.
Mechanical stretch injury and treatment of RA
RA was dissected free of fat and connective tissue. A segment was reserved as non-stretched control. The remaining RA was stretched to 200% the resting length for 10 s and repeated twice [15]. Stretched RA was then cut in segments and incubated for 1 h at room temperature in heparinized PlasmaLyte (HP) with or without inhibitors to P2XR or pannexin 1 (PanX1) [BB FCF (50 and 100 μM), A740003 (100 μM), the pannexin inhibitory peptide 10PanX1 peptide (200 μM; EZBiolab, IN), or carbenoxolone (100 μM)].
Measurement of physiologic responses
HSV or RA rings (1–2 mm) were suspended in a muscle bath containing a bicarbonate buffer (120 mM sodium chloride, 4.7 mM potassium chloride, 1.0 mM magnesium sulfate, 1.0 mM monosodium phosphate, 10 mM glucose, 1.5 mM calcium chloride, and 25 mM sodium bicarbonate, pH 7.4) equilibrated with 95% O2/5% CO2 at 37 °C for 1 h at a resting tension of 1 g, manually stretched to three times the resting tension, and maintained at resting tension for an additional 1 h. This produced the maximal force tension relationship as previously described [9]. Force measurements were obtained using the Radnoti force transducer (model 159901A) interfaced with a PowerLab data acquisition system and Chart software (AD Instruments). After equilibration, the rings were primed with 110 mM potassium chloride (with equimolar replacement of sodium chloride in bicarbonate buffer) to determine functional viability. Viable rings were then tested for contractile response to a dose of phenylephrine (PE) to yield submaximal contraction (approximately 60–70% of maximum KCl; 5 × 10−6 M for HSV and 1-5 × 10−7 M for RA). Contractile responses were defined by stress, calculated using force generated by tissues as follows: stress (×105 N/m2) = force (g) × 0.0987/area, where area = wet weight (mg)/ at maximal length (mm)]/1.055. Each data point was averaged from at least two rings from the same specimen.
Measurement of apoptosis by terminal deoxynucleotidyl transferase dUTP nick-end labeling
Immediately after collection, HSV (n = 11) were formalin-fixed, dehydrated with ethanol, embedded in paraffin. For RA (n = 5), after stretch injury and BB FCF treatment for 1 h, segments were cultured in RPMI 1640 media with 30% FBS at 37 °C in humidified incubator with 5% CO2 for 24 h prior to formalin fixation. Apoptosis were detected using DeadEndTM Flourometric TUNEL system (Promega, CA) per manufacturer’s instructions. Sections were then stained with anti-Von Willebrand Factor (Dako, CA) followed by Alex Fluor® 568-labeled secondary antibody (Invitrogen, CA) and mounted with SlowFade® Gold antifade reagent with DAPI (Life Technologies, CA). Images were acquired on Zeiss Axiovert 200 Fluorescence microscope at ×40 magnification. Terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) signals were quantified by Image J with the plugin “color_pixel_counter.class” with minimum intensity value set at 100. Extent of apoptosis (apoptosis index) was represented as percent green color pixels (TUNEL-positive cells) of the blue color pixels (DAPI-stainednuclei) of the entire vein section. In addition, serial sections of HSV were stained with Verhoeff Van Gieson stain to visualize elastic lamina.
Immunohistochemistry of P2X7R in RA
Freshly isolated, non-stretched RA (n = 4) was formalin-fixed and paraffin-embedded. Antigen retrieval of sections was performed with citrate buffer (pH 6) at 95°C for 5 min. After preincubation with 5% goat serum to block nonspecific sites, sections were incubated with primary antibodies against P2X7R (Alomone Labs, Jerusalem, Israel) overnight at 4°C. The sections were then incubated with Alexa 568-tagged anti-rabbit antibodies (Invitrogen) for 1 h. Controls were performed by preabsorbing the primary antibody with the immunogen peptide. Whole slide imaging was performed in the Digital Histology Shared Resource at Vanderbilt University Medical Center (www.mc.vanderbilt.edu/dhsr) on an Aperio Versa 200 automated slide scanner (Leica Biosystems). Tissue sections were imaged at ×40 magnification to a resolution of 0.162 μm/pixel.
Measurement of ATP released from RA
After stretch injury, RA (n = 23) were immediately cut into segments and placed in 60 μl of HP supplemented with the ectonuclease inhibitor ARL67156 (50 μM) and EDTA (2 mM) in the presence of P2X7R or PanX1 antagonists [oATP (100 μM), BB FCF (50 μM), or A740003 (100uM), 10PanX1 peptide (200 μM), or carbenoxolone (100 μM)]. ATP released was collected for 10 min, boiled for 5 min and stored at −80°C. ATP concentration in the perfusates was determined using the ATP Bioluminescent Assay Kit (FL-AAM; Sigma) and normalized to tissue weight.
Measurement of caspase activity in vascular smooth muscle cells
Rat aortic smooth muscle cells (A7r5; American Type Culture Collection, VA) were maintained in growth medium per suppliers’ recommendation and cultured at 37 °C in humidified incubator with 5% CO2. Cells were seeded in 96-well plates and pretreated with either BB FCF (100 μM) or medium for 1 h prior to stimulation with tumor necrosis factor (TNFα; 10 ng/ml; Life Technologies, CA) for 24 h. Caspase activity was determined using the Caspase-Glo 3/7 Assay Systems. Each data point was averaged from eight technical replicates from each independent experiments (n = 5).
Immunoblotting
Tissues were snap-frozen in liquid nitrogen immediately after collection, stretch injury or BB FCF treatment. Frozen vein segments were pulverized, and proteins were extracted in modified RIPA buffer (50 mM Tris-Cl, 150 mM NaCl, 1% NP40, 0.5% deoxycholic acid, 1 mM EDTA, 1 mM EDTA) supplement with protease inhibitor cocktail and phosphatase inhibitor cocktail 2. Total protein (30 μg) were subjected to SDS-PAGE and transferred to nitrocellulose membrane followed by immunoblotting with the following primary antibodies against phospho-p38 MAPK-Thr180/Tyr182, p38 MAPK, phospho-Akt-Ser473, Akt (Cell Signaling Technology, CA), phospho-Niban-Ser602 (Signalway, TX), Niban (Santa Cruz, CA), and GAPDH (Millipore, MA). The blots were then incubated with IRDye-labeled secondary antibodies (LI-COR Biosciences, NE). The protein-antibodies complexes were visualized and quantified using the on the Odyssey Infrared Imaging System. Phosphorylation was calculated as a ratio of the phosphorylated protein to total protein and was then normalized to the unstimulated control with the control value set as 1.0.
Statistical analysis
Data were reported as mean responses ± standard error of the mean and presented as scattered plots with mean and 95% confidence interval. Outliers, normality, and statistical significance (p value), and achieved power of each experiment was determined using GraphPad Prizm version 5.0 and G*Power version 3.1.9.2 (www.gpower.hhu.de/en.html). Differences among groups were determined by paired t test for experiments with dependent (matched) pairs. One-way ANOVA with post hoc test were used to determine differences among multiple, dependent samples from the same animal or multiple treatments in cells. A p value < 0.05 was considered statistically significant.
Results
Harvest and graft preparation impairs physiologic function of HSV
Paired, freshly harvested, unprepared (UP) and after typical intraoperative graft preparation (AP) HSV were collected at the time of CABG procedures. The average duration of intraoperative vein storage was 2.8 ± 1.5 h (data not shown). Rings were suspended in the muscle bath and contractile responses to depolarizing KCl (Fig. 1a) and the contractile agonist PE (Fig. 1b) were determined as previously described [9]. Vasomotor function was reduced in AP-HSV when compare to the cognate UP-HSV (Fig. 1).
Fig. 1.

Surgical harvest and graft preparation impairs physiologic function of human saphenous veins. Paired HSV collected immediately after harvest (unprepared, UP) and after typical graft preparation (AP) from CABG patients (n = 7) were suspended in the muscle bath. Contractile responses to (a) 110 mM KCl and (b) PE were measured. *p < 0.05
Harvest and graft preparation injury induces apoptosis and Niban dephosphorylation prior to implantation in HSV
Verhoeff Van Gieson staining of UP-HSV and AP-HSV preparations (Fig. 2a, d) revealed greater luminal distortion and disruption of lumen lining and medial layers in AP-but not UP-HSV. Detection of accumulated DNA degradation fragments using the TUNEL assay revealed considerably varied levels of TUNEL-positive nuclei among the UP- and AP-HSV segments examined (Fig. 2b, c, e, f); however, pair-wise comparison showed significant increases in the number of TUNEL signals in AP-HSV (Fig. 2g). There was a negative correlation between KCl-induced contractility and apoptotic index (Fig. 2h; r 2 = 0.213, p = 0.031). Taken together, these data suggest that surgical harvest and vein graft preparation lead to early activation of cellular apoptosis.
Fig. 2.
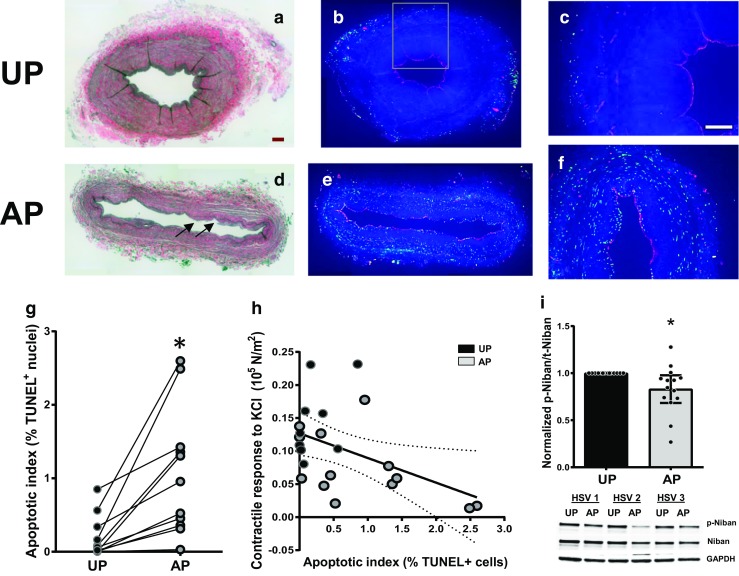
Vein graft preparation leads to apoptosis and modulates Niban phosphorylation in human saphenous veins. Paired HSV (UP and AP) from CABG patients were formalin-fixed and paraffin-embedded (a–h; n = 11) or snap-frozen for analysis (i; n = 14). a, d Representative images of Verhoeff Van Gieson staining showing elastic laminar and collagen. Note the lumen distortion and disruption of lumen lining and medial layers in the AP segment (d; arrows). b, e Representative images of TUNEL staining. Green, TUNEL-positive cells; Red, vWF-positive cells. c, f Boxed regions of b and e, respectively, showing TUNEL-positive cells in all three vessel layers. Scale bars = 200 μm. *p < 0.05. g Quantitative analysis of apoptotic index of the whole ring. h A linear regression of KCl-induced contractility as a function of apoptosis index yielding a r 2 = 0.213 and a negative correlation (p = 0.031). Black = UP; Gray = AP. Note that some of the data points overlaid each other. i Tissue protein lysates were immunoblotted for levels of phospho-Niban Ser602, total Niban and GAPDH. Top, quantitative analysis of Niban Ser602 phosphorylation levels normalized to UP tissues. Bottom, representative image of western blot analysis of paired HSVs from three different patients. *p < 0.05
In an untargeted, global phosphorylation profiling study comparing paired UP- and AP-HSV, we observed a modulation in Niban phosphorylation (Joyce Cheung-Flynn, unpublished data). Niban is highly expressed in tumors [27] and downregulation of phosphorylation is implicated in UV-induced cell death [28]. Western blot analysis revealed reduced levels of Niban phosphorylation in AP- while pan-Niban proteins shown no significant changes (Fig. 2d), indicating that Niban may be involved in the early, acute response to surgical harvest and preparation in HSV.
Subfailure overstretch injury impairs functional responses and causes release of extracellular ATP in RA
To examine mechanical injury to vein graft under controlled conditions, we previously developed a subfailure overstretch (longitudinal) injury model using isolated rat abdominal aorta (RA) [15]. Subfailure overstretch injury impaired the contractile response of isolated RA to depolarizing KCl (Fig. 3a, c) and PE (Fig. 3b, d) in the muscle bath, similar to those observed with HSV (Fig. 1).
Fig. 3.

P2X7R antagonism after stretch injury restores contractility in a subfailure overstretch model. Isolated rat abdominal aorta (RA) were either left untreated (NS) or stretched to twice the resting length. Stretched segments were then cut into 2 mm segments and incubated for 60 min in heparinized PlasmaLyte in the absence of presence of P2X7R or PanX1 inhibitors: BB FCF (50 and 100 μM), A740003 (A74; 100 μM), the pannexin inhibitory peptide 10PanX1 (200 μM), or carbenoxolone (CBX; 100 μM). Contractile response to (a, c) 110 mM KCl and (b, d) PE were measured in the muscle bath (n = 8–45). *p < 0.05. e Representative images showing P2X7R expression in untreated (NS) RA by immunohistochemistry. Red, P2X7R; blue, nuclei. Arrows indicate P2X7R expression in the endothelial (open arrow) and medial layers (closed arrow). Bottom panels show enlarged images of the boxed regions. Scale bar = 100 μm (top panels) and 10 μm (bottom panels)
Mechanical stretching leads to release of eATP [17, 29, 30]. To determine whether subfailure overstretch of vascular tissues triggers ATP release, RA were stretch-injured and ATP released was measured in the tissue perfusates (Fig. 4). eATP increased by 2.3 ± 0.4 μmole/mg acutely at 10 min following stretch injury. This increase trended to sustain for up to 1 h (data not shown).
Fig. 4.

P2X7R and PanX1 antagonism after stretch injury reduces stretch-mediated extracellular ATP release in a subfailure overstretch model. Isolated rat abdominal aorta (RA) were either left untreated (NS) or stretched to twice the resting length. Stretched segments were then cut into 1–2 mm segments and incubated in heparinized PlasmaLyte for 10 min in the absence or presence of inhibitors to P2X7R/PanX1 complex: a BB FCF (50 μM), oATP (100 μM), or A740003 (100 μM) and b the pannexin inhibitory peptide 10PanX1 (200 μM), or carbenoxolone (CBX; 100 μM). ATP released were collected and concentration in the perfusates were measured (n = 6–25)
P2X7R antagonism restores stretch-induced loss of contractile functions and prevents eATP release in RA
We previously showed that P2X7R is activated by subfailure overstretch in the RA model [15]. Immunohistochemical staining of RA showed that P2X7R is expressed in the medial and endothelial layers of RA (Fig. 3e). Treatment with pharmacological inhibitors of P2X7R, A740003 (A74; n = 13) [31], after subfailure overstretch injury restored contractile function impairment in RA (Fig. 3a, b). Treatment with BB FCF, a non-toxic dye that restored function in HSV [14], also restored functional impairment in stretched RA, in a dose-dependent manner (Fig. 3a, b; n = 6–13). Treatment with BB FCF (100 μM; n = 13) did not alter vasomotor function in non-injured RA. eATP release after subfailure overstretch was also blocked by A740003, oATP, and BB FCF (Fig. 4a). The PanX1 channel is functionally associated with P2X7R and is implicated in eATP release [32, 33]. Treatment with the PanX1 inhibitory peptide 10PanX1 and carbenoxolone (CBX) blocked eATP release in stretched RA (Fig. 4b; n = 8–15). However, treatment with these PanX1 inhibitors did not restore contractility in stretched RA (Fig. 3c, d; n = 12).
P2X7R/PanX1 antagonism reduces injury-induced apoptosis by subfailure overstretch in RA
Apoptosis has been associated with neuronal injury-induced P2X7R activation [34]. We next determined whether subfailure overstretch injury induces apoptosis in RA. TUNEL assay revealed an increase in number of TUNEL-positive nuclei within stretched RA segments compared to the non-stretched RA segment (Fig. 5a). Apoptosis was detected in all three layers of the vessel: intima (opened arrows), media (closed arrows), and adventitia (asterisks) (Fig. 5b). Since BB FCF has been recently approved for vein graft marking [35] and restored stretch-induced injury and ATP release, we determined whether it attenuates stretch-induced apoptosis in RA. BB FCF treatment after overstretch injury reduced increase in TUNEL-positive cells, suggesting that P2X7R/PanX1 complex activation is involved in stretch-induced apoptosis (Fig. 5a, b).
Fig. 5.

P2X7R/PanX1 antagonism reduces overstretch injury-induced vascular apoptosis in rat aorta. a Quantitative analysis of TUNEL assay (n = 5). NS, non-stretched; S, stretched. b Representative images of TUNEL staining of rat aorta. Green, TUNEL-positive cells; red, vWF-positive cells. Boxed region enlarged to show TUNEL-positive endothelial (open arrows), medial cell (closed arrows), and adventitial (asterisk) nuclei. Scale bar = 200 μm. c Caspase activity in TNFα–treated A7r5 cells with or without BB FCF pretreatment (n = 5). *p < 0.05
Given that P2X7R is implicated in maturation of the NLRP3 inflammasome multiprotein complex and that its expression and functional responses can be enhanced by proinflammatory cytokines [36, 37], interfering of P2X7R may reduce cell death resulting from inflammatory signaling. Therefore, we examined whether cytokine-induced apoptosis can be prevented by P2X7R/PanX1 complex blockade in cultured rat aortic smooth muscle cells (A7r5; Fig. 5c). Pretreatment with BB FCF reduced TNFα-induced caspase activity after 24 h. BB FCF also reduced caspase activity in cells that were not exposed to TNFα suggesting that protected protective effects on cell death caused by serum deprivation (data not shown).
Signaling networks of eATP-P2X7R modulated by subfailure overstretch injury in RA
To define the molecular signaling downstream of eATP-P2X7R/PanX1 complex in pathologic stretch injury, total protein extracts were prepared from RA after subfailure overstretch injury and analyzed using immunoblotting.
p38 MAPK activation plays a role in the P2X7R signal transduction and has been implicated in to P2X7R-mediated cell death [34, 38]. Subfailure overstretch injury increased the p38 MAPK phosphorylation level in RA (Fig. 6a). BB FCF treatment following stretch injury attenuated p38 MAPK phosphorylation.
Fig. 6.

P2X7R/PanX1 antagonism attenuates stress pathway activation and restores anti-apoptotic signaling in subfailure overstretch injured rat aorta. a p38MAPK Thr180/Tyr182 phosphorylation. b Niban Ser602 phosphorylation. c Akt Ser473 phosphorylation in RA. Top, quantitative analyses; Bottom, representative images of western blot analyses. Data shown were relative phosphorylation to total protein level normalized to non-stretched tissues. NS, non-stretched; S, stretched; S + BB FCF, stretched and treated with BB FCF (100 μM). n = 5, * p < 0.05
We showed in this study that phosphorylation level of the anti-apoptotic protein Niban is reduced in surgically prepared HSV (Fig. 2). Stretch injury of RA also led to decreases in Niban phosphorylation that can be restored by BB FCF treatment (Fig. 6b). These data suggest that Niban participates in stretch-induced P2X7R-mediated apoptosis in vascular tissues.
ATP cytotoxicity is mediated via P2X7R coupling to the PI3K-Akt pathway in neuronal cells and many tumor cell types [39] and is dependent on Akt activation upon UV-induced stress [28]. To determine whether eATP-induced cell death and P2X7R signaling is modulated via the Akt pathway in RA, Akt phosphorylation was measured. Subfailure overstretch had no apparent effect on Akt-Ser473 phosphorylation levels (Fig. 6c). Interestingly, BB FCF treatment increased Akt-Ser473 phosphorylation levels in stretch-injured (Fig. 6c) and non-stretched (data not shown) tissues suggesting that anti-apoptotic effect of P2X7R antagonism may be mediated by promoting the pro-survival property of Akt.
Discussion
Injury to vein graft used for CABG occurs during surgical harvest and intraoperative preparation and handling, and resulted in decreased vasomotor function (Fig. 1). Decreased vasomotor function is associated with decreased viability [40]. Additionally, surgical harvest and preparation of HSV leads to apoptosis (Fig. 2).
Traction or longitudinal stretching during endoscopic harvest inflicts mechanical trauma to saphenous vein and reduces HSV graft patency (reviewed in [41]). To determine the molecular mechanisms by which injury leads to decreased function and viability of HSV, a model of traction injury using RA was developed where the haptic endpoint of stretch during vein harvest is simulated [15]. While studies using cultured vascular cells to modeled mechanical stretch offer insight into molecular signaling that determine cell fate (reviewed in [42, 43]), very few studied the effects of pathologic stretch in intact tissues [40, 44, 45]. The RA stretch injury model demonstrates a causal relationship between subfailure overstretch injury and loss of smooth muscle functional physiologic responses that can be restored by the antagonism to the P2X7R-PanX1 crosstalk after injury (Fig. 3a).
There is increasing evidence for the role of extracellular ATP in pathological conditions [46]. A steep ATP concentration gradient between the cytoplasm (1–10 mM) [47] and the extracellular space (1–10 nM) [48] suggests that a rapid spike in ATP levels in the local extracellular milieu due to tissue stress or damage can readily elicit responses via P2X7R (EC50 of 300–800 μM) [49]. Subfailure overstretch injury of RA leads to release of ATP (Fig. 4), activation of P2X7R, and apoptosis (Fig. 5). Notably, treatment with P2X7R inhibitors after injury ameliorated the injurious responses and reduced eATP release. These findings suggest the existence of a positive feedback loop that potentiates the effect of eATP, amplifying the “death” signal (Fig. 7). It is widely accepted that PanX1 activity is tightly coupled to P2X7R activation and the two channel physically interact [21, 50]. Wang et al. previously reported that BB FCF inhibits PanX1 current but not P2X7R current [51]. In the same study, oATP, a P2X7R antagonist reported to have no significant inhibitory effect on PanX1 current [51], alleviated contractility loss and blocked eATP release in stretch-injured RA. Interestingly, inhibition of vascular PanX1with the 10PanX1 peptide and CBX blocked eATP release in RA (Fig. 4) but did not restore functional impairment after stretch (Fig. 3b, d). It is conceivable that while inhibition of eATP release is inhibited by interfering with either P2XR or PanX1 after stretch injury, P2X7R may be saturated with the initial burst of ATP release from stretch-injured cells. Additionally, activation of downstream signaling cascade, such as p38 MAPK activation (Fig. 6a) which inhibits vascular smooth muscle contraction (Colleen Brophy, unpublished observation), may precedes the amplification of ATP release. Without inhibiting P2X7R activation, blockade of further ATP release and its consequences may be too subtle to produce notable changes in acute physiologic response in the RA model. Nonetheless, amplification of ATP signal and downstream signaling of the eATP-P2X7R/PanX1 signalome overtime may contribute to other pathological manifestation of vascular injury. These data suggest that eATP and functional relationship between P2X7R and PanX1 contribute to subfailure overstretch injury and decrease vasomotor function in vascular tissues.
Fig. 7.

Putative molecular mechanism of P2X7R-induced apoptosis in vascular tissues during subfailure overstretch injury. Traction and stretch injury leads to release of extracellular ATP (1), activating P2X7R and the associated PanX1 (2). Activation of p38MAPK and dephosphorylation of Niban follows (3) and ultimately leads to caspase 3 cleavage and apoptosis ensues (4). P2X7R activation also results in the formation of large membrane pores, intracellular calcium flux, and cytolysis (5). ATP released from dying cells activates the P2X7R of neighboring cells, propagating the response to injury (6). By antagonizing the P2X7R during vein harvest and preparation (7), further release of ATP and prolonged P2X7R activation is prevented, thus alleviates stretch-induced apoptosis in part via the pro-survival activity of Akt and Niban and possibly crosstalk with other signaling mechanisms have yet to be determined
The use of BB FCF to reverse injury occur during endoscopic traction injury has clinically relevance as it is a non-toxic vein marking dye [35]. Treatment with BB FCF after subfailure overstretch injury reduced apoptosis in RA (Fig. 5), suggesting that P2X7R/PanX1 complex mediates cell death after stretch injury in vascular tissues. Apoptosis is one of the early events of injury response and is proposed to mediate subsequent proliferation of synthetic smooth muscle cells (SMC) that populate the neointima of vein grafts [52]. Ahmed et al. showed that within 30 min of surgical preparation, features of SMC cell division and apoptosis were observed histologically in conventionally prepared HSV compared to the “no touch” harvest confirming that harvest technique influences apoptosis [6]. In animal models of vein grafting, nuclear condensation and membrane blebbing in medial SMCs were evident as early as 1 h post-grafting [53], and apoptosis peaked within 8 h of grafting [54]. Mechanical stretch associated both physiological or pathologic conditions is an inducer of cytokine production in difference tissues [55, 56]. In addition to alleviating functional impairment, BB FCF also protects against cytokine-induced apoptosis of smooth muscle cells (Fig. 5c). It is conceivable that blocking P2X7R/PanX1 signaling during vein graft preparation may protect vascular cells from stretch-induced cell death and the secondary inflammatory response.
Activation of the p38 MAPK pathway is a downstream effector of P2X7R [38, 57–59]. Mechanical stretch-induced phosphorylation of p38 MAPK and apoptosis were reported in human saphenous vein under pressure distension [60] and in cyclically stretched cultured smooth muscle cells [61]. Subfailure overstretch-induced activation of P2X7R increased p38 MAPK phosphorylation in intact vascular tissues that was reversed by treatment with BB FCF after the injury (Fig. 6a). Pathological stretch injury also led to decreases in Niban phosphorylation in RA (Fig. 6b). Niban is overexpressed in several cancers and modulate cell death through regulation of its expression and phosphorylation [62–66]. The anti-apoptotic activity of Niban is governed by Akt-dependent phosphorylation as it disrupts p53-containing protein complex under UV-stress [28] and experimental overexpression of the protein alone failed to diminished apoptosis in tumor cells [67]. Depending on the cell type and the nature of the stimuli, cytotoxic concentration of ATP either inhibits [68, 69] or stimulates Akt phosphorylation [39, 70]. Similarly, variable results have been documented for stretch-induced Akt activation depending on the experimental settings. In the subfailure overstretch injury model, stretch did not modulate Akt S473 phosphorylation while P2X7R antagonism enhances it in both normal (data not shown) and injured tissues (Fig. 6c).
Taken together, our data suggest that subfailure overstretch injury during harvest leads to release of ATP and activation of P2X7R, which in turn further amplifies the release of eATP through the P2X7R/PanX1 complex (Fig. 7). This ATP-induced activation of P2X7R contributes to functional impairment, activates cellular signaling events, and leads to apoptosis. Treatment with P2X7R antagonists after traction injury abrogates this response to injury suggesting that P2X7R inhibition represents a viable therapeutic approach in that vein grafts could be treated ex vivo with P2X7R antagonists prior to implantation. In addition, PanX1 inhibitors can also be explored as additional therapeutics that prevents further release of ATP. Since response to injury initiates the processes that ultimately lead to intimal hyperplasia, P2X7R and/or Panx1 antagonism may represent a novel approach to preventing vein graft failure.
Conclusion
Vascular P2X7R plays a central role in the acute injury response to traction stretch injury in vein grafts (Fig. 7). A novel eATP-P2X7R model may explain some of the early molecular events that contribute to vein graft injury—mechanical stretch leads to release of eATP, which diffuses and influences adjacent cells, propagating deleterious eATP-mediated P2X7R positive feedback signaling that involves p38 MAPK, Akt and Niban-mediated regulation of apoptosis. In this context, eATP might serve as an important molecular mediator of vascular stretch injury response. Blockade of the vascular P2X7R after injury not only restored vasomotor function, it also inhibits detrimental functional sequelae of stretch injury by attenuating signaling of pro-apoptotic proteins while enhancing activity of pro-survival pathways. Moreover, the finding that inhibition of the PanX1 reduced eATP release underscores the potential role of the channel in P2X7R activation by vascular stretch injury. By preventing acute apoptotic response to preparation injury, it is plausible to alter the kinetics of graft adaptation that contributes to the progression of remodeling that leads to graft occlusion. P2X7R inhibition represents a potential strategy to improve vein graft quality used in bypass procedures.
Limitations
Human saphenous veins varied due to patient demographics, but effect of graft preparation on physiologic responses and apoptosis was adjusted for in pair-wise comparison of segments collected from the same patients. Isolated rat vessels were used in this study, which may present species differences in molecular response to injury. Compared to cultured cells, the use of intact tissue takes into account the natural tissue environment of the vessel which is three dimensional with interaction among multiple cell types; however, the cell type-specific responses were not delineated. Apoptosis of RA were detected after a 1-day organ culture which lacks the blood component and hemodynamics forces that influence early vein graft remodeling in vivo.
Acknowledgments
We would like to thank the Vanderbilt cardiac surgical teams for providing human specimens for this study.
Funding Information
This work was supported by NIH grants R01HL70715-09 (to CB) R01HL105731-01 (to JC), a VA merit award I01BX002036 (to CB), and in part by CTSA award No. UL1TR000445 from the National Center for Advancing Translational Sciences (to JC). Its contents are solely the responsibility of the authors and do not necessarily represent official views of the National Center for Advancing Translational Sciences or the National Institutes of Health.
Compliance with ethical standards
Conflict of interest
Drs. Brophy and Cheung-Flynn are inventors on U.S. Patent 8691556 and disclose that they have financial relationships with VasoPrep Surgical, Inc. (Morristown, NJ).
Ethical approval
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards. All applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
References
- 1.Conte MS, Bandyk DF, Clowes AW, et al. Results of PREVENT III: a multicenter, randomized trial of edifoligide for the prevention of vein graft failure in lower extremity bypass surgery. J Vasc Surg. 2006;43:742–751. doi: 10.1016/j.jvs.2005.12.058. [DOI] [PubMed] [Google Scholar]
- 2.Alexander JH, Hafley G, Harrington RA, et al. Efficacy and safety of edifoligide, an E2F transcription factor decoy, for prevention of vein graft failure following coronary artery bypass graft surgery: PREVENT IV: a randomized controlled trial. JAMA. 2005;294:2446–2454. doi: 10.1001/jama.294.24.3108. [DOI] [PubMed] [Google Scholar]
- 3.Rousou LJ, Taylor KB, XG L, et al. Saphenous vein conduits harvested by endoscopic technique exhibit structural and functional damage. Ann Thorac Surg. 2009;87:62–70. doi: 10.1016/j.athoracsur.2008.08.049. [DOI] [PubMed] [Google Scholar]
- 4.Cook RC, Crowley CM, Hayden R, et al. Traction injury during minimally invasive harvesting of the saphenous vein is associated with impaired endothelial function. J Thorac Cardiovasc Surg. 2004;127:65–71. doi: 10.1016/S0022-5223(03)01024-9. [DOI] [PubMed] [Google Scholar]
- 5.Kiani S, Desai PH, Thirumvalavan N, et al. Endoscopic venous harvesting by inexperienced operators compromises venous graft remodeling. Ann Thorac Surg. 2012;93:11–17. doi: 10.1016/j.athoracsur.2011.06.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ahmed SR, Johansson BL, Karlsson MG, Souza DS, Dashwood MR, Loesch A. Human saphenous vein and coronary bypass surgery: ultrastructural aspects of conventional and “no-touch” vein graft preparations. Histol Histopathol. 2004;19:421–433. doi: 10.14670/HH-19.421. [DOI] [PubMed] [Google Scholar]
- 7.Hocking KM, Luo W, Li FD, Komalavilas P, Brophy C, Cheung-Flynn J. Brilliant blue FCF is a nontoxic dye for saphenous vein graft marking that abrogates response to injury. J Vasc Surg. 2016;64:210–218. doi: 10.1016/j.jvs.2014.12.059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wise ES, Hocking KM, Eagle S, et al. Preservation solution impacts physiologic function and cellular viability of human saphenous vein graft. Surgery. 2015;158:537–546. doi: 10.1016/j.surg.2015.03.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Osgood MJ, Hocking KM, Voskresensky IV, et al. Surgical vein graft preparation promotes cellular dysfunction, oxidative stress, and intimal hyperplasia in human saphenous vein. J Vasc Surg. 2014;60:202–211. doi: 10.1016/j.jvs.2013.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Galea J, Armstrong J, Francis SE, Cooper G, Crossman DC, Holt CM. Alterations in c-fos expression, cell proliferation and apoptosis in pressure distended human saphenous vein. Cardiovasc Res. 1999;44:436–448. doi: 10.1016/S0008-6363(99)00220-5. [DOI] [PubMed] [Google Scholar]
- 11.Moggio RA, Ding JZ, Smith CJ, Tota RR, Stemerman MB, Reed GE. Immediate-early gene expression in human saphenous veins harvested during coronary artery bypass graft operations. J Thorac Cardiovasc Surg. 1995;110:209–213. doi: 10.1016/S0022-5223(05)80027-3. [DOI] [PubMed] [Google Scholar]
- 12.Hinokiyama K, Valen G, Tokuno S, Vedin JB, Vaage J. Vein graft harvesting induces inflammation and impairs vessel reactivity. Ann Thorac Surg. 2006;82:1458–1464. doi: 10.1016/j.athoracsur.2006.05.038. [DOI] [PubMed] [Google Scholar]
- 13.Wise ES, Hocking KM, Feldman D, Komalavilas P, Cheung-Flynn J, Brophy CM. An optimized preparation technique for saphenous vein graft. Am Surg. 2015;81:E274–E276. [PMC free article] [PubMed] [Google Scholar]
- 14.Voskresensky IV, Wise ES, Hocking KM, et al. Brilliant blue FCF as an alternative dye for saphenous vein graft marking: effect on conduit function. JAMA Surg. 2014;149:1176–1181. doi: 10.1001/jamasurg.2014.2029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Luo W, Guth CM, Jolayemi O, Duvall CL, Brophy CM, Cheung-Flynn J. Subfailure overstretch injury leads to reversible functional impairment and purinergic P2X7 receptor activation in intact vascular tissue. Front Bioeng Biotechnol. 2016;4:75. doi: 10.3389/fbioe.2016.00075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Beckel JM, Argall AJ, Lim JC, et al. Mechanosensitive release of adenosine 5′-triphosphate through pannexin channels and mechanosensitive upregulation of pannexin channels in optic nerve head astrocytes: a mechanism for purinergic involvement in chronic strain. Glia. 2014;62:1486–1501. doi: 10.1002/glia.22695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Sadananda P, Shang F, Liu L, Mansfield KJ, Burcher E. Release of ATP from rat urinary bladder mucosa: role of acid, vanilloids and stretch. Br J Pharmacol. 2009;158:1655–1662. doi: 10.1111/j.1476-5381.2009.00431.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Grygorczyk R, Furuya K, Sokabe M. Imaging and characterization of stretch-induced ATP release from alveolar A549 cells. J Physiol. 2013;591:1195–1215. doi: 10.1113/jphysiol.2012.244145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Loomis WH, Namiki S, Ostrom RS, Insel PA, Junger WG. Hypertonic stress increases T cell interleukin-2 expression through a mechanism that involves ATP release, P2 receptor, and p38 MAPK activation. J Biol Chem. 2003;278:4590–4596. doi: 10.1074/jbc.M207868200. [DOI] [PubMed] [Google Scholar]
- 20.Vandenbeuch A, Anderson CB, Kinnamon SC. Mice lacking Pannexin 1 release ATP and respond normally to all taste qualities. Chem Senses. 2015;40:461–467. doi: 10.1093/chemse/bjv034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Dahl G (2015) ATP Release through pannexon channels. Philos Trans R Soc Lond Ser B Biol Sci 370:20140191 [DOI] [PMC free article] [PubMed]
- 22.Song XM, XH X, Zhu J, et al. Up-regulation of P2X7 receptors mediating proliferation of Schwann cells after sciatic nerve injury. Purinergic Signal. 2015;11:203–213. doi: 10.1007/s11302-015-9445-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Feng L, Chen Y, Ding R, et al. P2X7R blockade prevents NLRP3 inflammasome activation and brain injury in a rat model of intracerebral hemorrhage: involvement of peroxynitrite. J Neuroinflammation. 2015;12:190. doi: 10.1186/s12974-015-0409-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Burnstock G, Kennedy C. P2X receptors in health and disease. Adv Pharmacol. 2011;61:333–372. doi: 10.1016/B978-0-12-385526-8.00011-4. [DOI] [PubMed] [Google Scholar]
- 25.Peng W, Cotrina ML, Han X, et al. Systemic administration of an antagonist of the ATP-sensitive receptor P2X7 improves recovery after spinal cord injury. Proc Natl Acad Sci U S A. 2009;106:12489–12493. doi: 10.1073/pnas.0902531106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Bartlett R, Stokes L, Sluyter R. The P2X7 receptor channel: recent developments and the use of P2X7 antagonists in models of disease. Pharmacol Rev. 2014;66:638–675. doi: 10.1124/pr.113.008003. [DOI] [PubMed] [Google Scholar]
- 27.Majima S, Kajino K, Fukuda T, Otsuka F, Hino OA. Novel gene “Niban” upregulated in renal carcinogenesis: cloning by the cDNA-amplified fragment length polymorphism approach. Jpn J Cancer Res. 2000;91:869–874. doi: 10.1111/j.1349-7006.2000.tb01027.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ji H, Ding Z, Hawke D, et al. AKT-dependent phosphorylation of Niban regulates nucleophosmin- and MDM2-mediated p53 stability and cell apoptosis. EMBO Rep. 2012;13:554–560. doi: 10.1038/embor.2012.53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Neary JT, Kang Y, Willoughby KA, Ellis EF. Activation of extracellular signal-regulated kinase by stretch-induced injury in astrocytes involves extracellular ATP and P2 purinergic receptors. J Neurosci. 2003;23:2348–2356. doi: 10.1523/JNEUROSCI.23-06-02348.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Takahara N, Ito S, Furuya K, et al. Real-time imaging of ATP release induced by mechanical stretch in human airway smooth muscle cells. Am J Respir Cell Mol Biol. 2014;51:772–782. doi: 10.1165/rcmb.2014-0008OC. [DOI] [PubMed] [Google Scholar]
- 31.Hong Y, Wang G, Del Arroyo AG, Hernandez J, Skene C, Erusalimsky JD. Comparison between anagrelide and hydroxycarbamide in their activities against haematopoietic progenitor cell growth and differentiation: selectivity of anagrelide for the megakaryocytic lineage. Leukemia. 2006;20:1117–1122. doi: 10.1038/sj.leu.2404180. [DOI] [PubMed] [Google Scholar]
- 32.Bao L, Locovei S, Dahl G. Pannexin membrane channels are mechanosensitive conduits for ATP. FEBS Lett. 2004;572:65–68. doi: 10.1016/j.febslet.2004.07.009. [DOI] [PubMed] [Google Scholar]
- 33.Godecke S, Roderigo C, Rose CR, Rauch BH, Godecke A, Schrader J, Thrombin-induced ATP. Release from human umbilical vein endothelial cells. Am J Physiol Cell Physiol. 2012;302:C915–C923. doi: 10.1152/ajpcell.00283.2010. [DOI] [PubMed] [Google Scholar]
- 34.Gandelman M, Peluffo H, Beckman JS, Cassina P, Barbeito L, Extracellular ATP. The P2X7 receptor in astrocyte-mediated motor neuron death: implications for amyotrophic lateral sclerosis. J Neuroinflammation. 2010;7:33. doi: 10.1186/1742-2094-7-33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Brophy C, and Lander C (2015) De novo request from VasoPrep Surgical (formerly Moerae Matrix, Inc). Fed Regist 80:46485-46486
- 36.Humphreys BD, Dubyak GR. Modulation of P2X7 nucleotide receptor expression by pro- and anti-inflammatory stimuli in THP-1 monocytes. J Leukoc Biol. 1998;64:265–273. doi: 10.1002/jlb.64.2.265. [DOI] [PubMed] [Google Scholar]
- 37.Franceschini A, Capece M, Chiozzi P, et al. The P2X7 receptor directly interacts with the NLRP3 inflammasome scaffold protein. FASEB J. 2015;29:2450–2461. doi: 10.1096/fj.14-268714. [DOI] [PubMed] [Google Scholar]
- 38.Donnelly-Roberts DL, Namovic MT, Faltynek CR, Jarvis MF. Mitogen-activated protein kinase and caspase signaling pathways are required for P2X7 receptor (P2X7R)-induced pore formation in human THP-1 cells. J Pharmacol Exp Ther. 2004;308:1053–1061. doi: 10.1124/jpet.103.059600. [DOI] [PubMed] [Google Scholar]
- 39.Jacques-Silva MC, Rodnight R, Lenz G, et al. P2X7 receptors stimulate AKT phosphorylation in astrocytes. Br J Pharmacol. 2004;141:1106–1117. doi: 10.1038/sj.bjp.0705685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hocking KM, Brophy C, Rizvi SZ, et al. Detrimental effects of mechanical stretch on smooth muscle function in saphenous veins. J Vasc Surg. 2011;53:454–460. doi: 10.1016/j.jvs.2010.09.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Raja SG, Sarang Z. Endoscopic vein harvesting: technique, outcomes, concerns & controversies. J Thorac Dis. 2013;5(Suppl 6):S630–S637. doi: 10.3978/j.issn.2072-1439.2013.10.01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Jufri NF, Mohamedali A, Avolio A, Baker MS. Mechanical stretch: physiological and pathological implications for human vascular endothelial cells. Vasc Cell. 2015;7:8. doi: 10.1186/s13221-015-0033-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Mantella LE, Quan A, Verma S. Variability in vascular smooth muscle cell stretch-induced responses in 2D culture. Vasc Cell. 2015;7:7. doi: 10.1186/s13221-015-0032-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Zhang K, Cao J, Dong R, Du J. Early growth response protein 1 promotes restenosis by upregulating intercellular adhesion molecule-1 in vein graft. Oxidative Med Cell Longev. 2013;2013:432409. doi: 10.1155/2013/432409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Rodriguez AI, Csanyi G, Ranayhossaini DJ, et al. MEF2B-Nox1 signaling is critical for stretch-induced phenotypic modulation of vascular smooth muscle cells. Arterioscler Thromb Vasc Biol. 2015;35:430–438. doi: 10.1161/ATVBAHA.114.304936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Khakh BS, North RA. P2X receptors as cell-surface ATP sensors in health and disease. Nature. 2006;442:527–532. doi: 10.1038/nature04886. [DOI] [PubMed] [Google Scholar]
- 47.Zamaraeva MV, Sabirov RZ, Maeno E, Ando-Akatsuka Y, Bessonova SV, Okada Y. Cells die with increased cytosolic ATP during apoptosis: a bioluminescence study with intracellular luciferase. Cell Death Differ. 2005;12:1390–1397. doi: 10.1038/sj.cdd.4401661. [DOI] [PubMed] [Google Scholar]
- 48.Fitz JG. Regulation of cellular ATP release. Trans Am Clin Climatol Assoc. 2007;118:199–208. [PMC free article] [PubMed] [Google Scholar]
- 49.North RA, Surprenant A. Pharmacology of cloned P2X receptors. Annu Rev Pharmacol Toxicol. 2000;40:563–580. doi: 10.1146/annurev.pharmtox.40.1.563. [DOI] [PubMed] [Google Scholar]
- 50.Silverman WR, de Rivero Vaccari JP, Locovei S, et al. The pannexin 1 channel activates the inflammasome in neurons and astrocytes. J Biol Chem. 2009;284:18143–18151. doi: 10.1074/jbc.M109.004804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Wang J, Jackson DG, Dahl G. The food dye FD&C Blue no. 1 is a selective inhibitor of the ATP release channel Panx1. The Journal of general physiology. 2013;141:649–656. doi: 10.1085/jgp.201310966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Liu SQ, Ruan YY, Tang D, Li YC, Goldman J, Zhong LA. Possible role of initial cell death due to mechanical stretch in the regulation of subsequent cell proliferation in experimental vein grafts. Biomech Model Mechanobiol. 2002;1:17–27. doi: 10.1007/s10237-002-0003-2. [DOI] [PubMed] [Google Scholar]
- 53.Stark VK, Warner TF, Hoch JR. An ultrastructural study of progressive intimal hyperplasia in rat vein grafts. J Vasc Surg. 1997;26:94–103. doi: 10.1016/S0741-5214(97)70152-6. [DOI] [PubMed] [Google Scholar]
- 54.Rodriguez E, Lambert EH, Magno MG, Mannion JD. Contractile smooth muscle cell apoptosis early after saphenous vein grafting. Ann Thorac Surg. 2000;70:1145–1153. doi: 10.1016/S0003-4975(00)01768-9. [DOI] [PubMed] [Google Scholar]
- 55.Lee YH, Shynlova O, Lye SJ. Stretch-induced human myometrial cytokines enhance immune cell recruitment via endothelial activation. Cell Mol Immunol. 2015;12:231–242. doi: 10.1038/cmi.2014.39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Gruden G, Setti G, Hayward A, et al. Mechanical stretch induces monocyte chemoattractant activity via an NF-kappaB-dependent monocyte chemoattractant protein-1-mediated pathway in human mesangial cells: inhibition by rosiglitazone. J Am Soc Nephrol. 2005;16:688–696. doi: 10.1681/ASN.2004030251. [DOI] [PubMed] [Google Scholar]
- 57.Panenka W, Jijon H, Herx LM, et al. P2X7-like receptor activation in astrocytes increases chemokine monocyte chemoattractant protein-1 expression via mitogen-activated protein kinase. J Neurosci. 2001;21:7135–7142. doi: 10.1523/JNEUROSCI.21-18-07135.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Noguchi T, Ishii K, Fukutomi H, et al. Requirement of reactive oxygen species-dependent activation of ASK1-p38 MAPK pathway for extracellular ATP-induced apoptosis in macrophage. J Biol Chem. 2008;283:7657–7665. doi: 10.1074/jbc.M708402200. [DOI] [PubMed] [Google Scholar]
- 59.Chen S, Ma Q, Krafft PR, et al. P2X7 receptor antagonism inhibits p38 mitogen-activated protein kinase activation and ameliorates neuronal apoptosis after subarachnoid hemorrhage in rats. Crit Care Med. 2013;41:e466–e474. doi: 10.1097/CCM.0b013e31829a8246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Cornelissen J, Armstrong J, Holt CM. Mechanical stretch induces phosphorylation of p38-MAPK and apoptosis in human saphenous vein. Arterioscler Thromb Vasc Biol. 2004;24:451–456. doi: 10.1161/01.ATV.0000116690.17017.8b. [DOI] [PubMed] [Google Scholar]
- 61.Mayr M, Li C, Zou Y, Huemer U, Hu Y, Biomechanical XQ. Stress-induced apoptosis in vein grafts involves p38 mitogen-activated protein kinases. FASEB J. 2000;14:261–270. doi: 10.1096/fasebj.14.2.261. [DOI] [PubMed] [Google Scholar]
- 62.Matsumoto F, Fujii H, Abe M, et al. A novel tumor marker, Niban, is expressed in subsets of thyroid tumors and Hashimoto’s thyroiditis. Hum Pathol. 2006;37:1592–1600. doi: 10.1016/j.humpath.2006.06.022. [DOI] [PubMed] [Google Scholar]
- 63.Ito S, Fujii H, Matsumoto T, Abe M, Ikeda K, Hino O. Frequent expression of Niban in head and neck squamous cell carcinoma and squamous dysplasia. Head Neck. 2010;32:96–103. doi: 10.1002/hed.21153. [DOI] [PubMed] [Google Scholar]
- 64.Kannangai R, Diehl AM, Sicklick J, Rojkind M, Thomas D, Torbenson M. Hepatic angiomyolipoma and hepatic stellate cells share a similar gene expression profile. Hum Pathol. 2005;36:341–347. doi: 10.1016/j.humpath.2005.01.002. [DOI] [PubMed] [Google Scholar]
- 65.Adachi H, Majima S, Kon S, et al. Niban gene is commonly expressed in the renal tumors: a new candidate marker for renal carcinogenesis. Oncogene. 2004;23:3495–3500. doi: 10.1038/sj.onc.1207468. [DOI] [PubMed] [Google Scholar]
- 66.Sun GD, Kobayashi T, Abe M, et al. The endoplasmic reticulum stress-inducible protein Niban regulates eIF2alpha and S6K1/4E-BP1 phosphorylation. Biochem Biophys Res Commun. 2007;360:181–187. doi: 10.1016/j.bbrc.2007.06.021. [DOI] [PubMed] [Google Scholar]
- 67.Liu J, Qin J, Mei W, et al. Expression of Niban in renal interstitial fibrosis. Nephrology (Carlton) 2014;19:479–489. doi: 10.1111/nep.12266. [DOI] [PubMed] [Google Scholar]
- 68.Bian S, Sun X, Bai A, et al. P2X7 integrates PI3K/AKT and AMPK-PRAS40-mTOR signaling pathways to mediate tumor cell death. PLoS One. 2013;8:e60184. doi: 10.1371/journal.pone.0060184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Mistafa O, Ghalali A, Kadekar S, Hogberg J, Stenius U. Purinergic receptor-mediated rapid depletion of nuclear phosphorylated Akt depends on pleckstrin homology domain leucine-rich repeat phosphatase, calcineurin, protein phosphatase 2A, and PTEN phosphatases. J Biol Chem. 2010;285:27900–27910. doi: 10.1074/jbc.M110.117093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Amoroso F, Capece M, Rotondo A et al (2015) The P2X7 receptor is a key modulator of the PI3K/GSK3beta/VEGF signaling network: evidence in experimental neuroblastoma. Oncogene [DOI] [PubMed]