

Summary
The intestinal microbiota influence neurodevelopment, modulate behavior, and contribute to neurological disorders. However, a functional link between gut bacteria and neurodegenerative diseases remains unexplored. Synucleinopathies are characterized by aggregation of the protein α-synuclein (αSyn), often resulting in motor dysfunction as exemplified by Parkinson's disease (PD). Using mice that overexpress αSyn, we report herein that gut microbiota are required for motor deficits, microglia activation, and αSyn pathology. Antibiotic treatment ameliorates, while microbial re-colonization promotes, pathophysiology in adult animals, suggesting postnatal signaling between the gut and the brain modulates disease. Indeed, oral administration of specific microbial metabolites to germ-free mice promotes neuroinflammation and motor symptoms. Remarkably, colonization of αSyn-overexpressing mice with microbiota from PD patients enhances physical impairments compared to microbiota transplants from healthy human donors. These findings reveal that gut bacteria regulate movement disorders in mice, and suggest that alterations in the human microbiome represent a risk factor for PD.
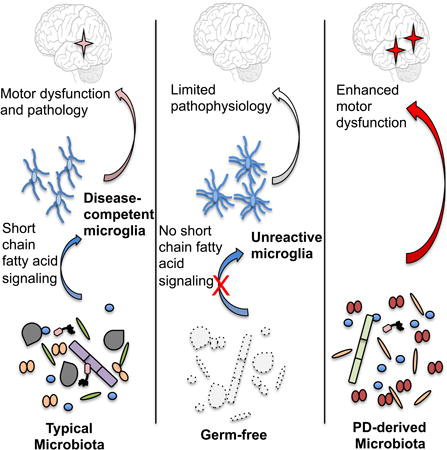
Graphical abstract
Signals from gut microbes are required for the neuroinflammatory responses as well as hallmark gastrointestinal and α-synuclein-dependent motor deficits in a model of Parkinson's disease.
Introduction
Neurological dysfunction is the basis of numerous human diseases. Behavioral, psychiatric and neurodegenerative disorders often display hallmark neuropathologies within the central nervous system (CNS). One neuropathology, amyloidosis, results from aberrant aggregation of specific neuronal proteins that disrupt many cellular functions. Affected tissues often contain insoluble aggregates of proteins that display altered conformations, a feature believed to contribute to an estimated 50 distinct human diseases (Sacchettini and Kelly, 2002). Neurodegenerative amyloid disorders, including Alzheimer's, Huntington's, and Parkinson's diseases (PD), are each associated with a distinct amyloid protein (Brettschneider et al., 2015). PD is the second most common neurodegenerative disease in the United States, affecting an estimated 1 million people and 1% of the US population over 60 years of age (Nalls et al., 2014). Worldwide, about 3 million patients and caregivers suffer from the often-debilitating symptoms of PD, which involve motor deficits including tremors, muscle rigidity, bradykinesia, and impaired gait. It is a multifactorial disorder that has a strong environmental component, as less than 10% of cases are hereditary (Nalls et al., 2014). Aggregation of α-synuclein (αSyn) is thought to be pathogenic in a family of diseases termed synucleinopathies, which includes PD, multiple system atrophy and Lewy body disease (Brettschneider et al., 2015; Luk et al., 2012; Prusiner et al., 2015). αSyn aggregation is a stepwise process, leading to oligomeric species and intransient fibrils that accumulate within neurons. Dopaminergic neurons of the substantia nigra pars compacta (SNpc) appear particularly vulnerable to effects of αSyn aggregates. Dopamine modulators are a first line therapeutic in PD; however treatments can carry serious side effects and often lose effectiveness (Jenner, 2008). Discovery of safe and effective therapeutics are needed to address the increasing burden of PD in an ever-aging population, a paradoxical consequence of mankind's achievements in increased lifespan.
Although neurological diseases have been historically studied within the CNS, peripheral influences have been implicated in the onset and/or progression of diseases that impact the brain (Dinan and Cryan, 2015). Indeed, emerging data suggest bidirectional communication between the gut and the brain in anxiety, depression, nociception and autism spectrum disorder (ASD), among others (Mayer et al., 2014; Schroeder and Backhed, 2016; Sharon et al., 2016) Gastrointestinal (GI) physiology and motility are influenced by signals arising both locally within the gut and from the CNS. Neurotransmitters, immune signaling, hormones and neuropeptides produced within the gut may, in turn, impact the brain (Selkrig et al., 2014; Wall et al., 2014). Research into how the gut-brain axis influences neurological conditions may reveal insights into disease etiology.
The human body is permanently colonized by microbes on virtually all environmentally exposed surfaces, the majority of which reside within the GI tract (Ley et al., 2006). Increasingly, research is beginning to uncover the profound impacts that the microbiota can have on neurodevelopment and the CNS (Sharon et al., 2016). Germ-free (GF) mice, and antibiotic treated specific pathogen free (SPF) mice, are altered in hippocampal neurogenesis, resulting in impaired spatial and object recognition (Mohle et al., 2016). The microbiota regulate expression of the 5-hydroxytryptamine receptor (5-HT1A), brain-derived neurotropic factor (BDNF), and NMDA receptor subunit 2 (NR2A) (Bercik et al., 2011; Diaz Heijtz et al., 2011; Sudo et al., 2004). GF mice have altered cortical myelination and impaired blood-brain barrier function (Braniste et al., 2014; Hoban et al., 2016). Additionally, the microbiota promotes enteric and circulating serotonin production in mice (Yano et al., 2015), and affects anxiety, hyperactivity and cognition (Clarke et al., 2013; Diaz Heijtz et al., 2011; Neufeld et al., 2011; Selkrig et al., 2014). To augment mouse models, dysbiosis (alterations to the microbial composition) of the human microbiome has been reported in subjects diagnosed with several neurological diseases (Schroeder and Backhed, 2016). For example, fecal and mucosa-associated gut microbes are different between individuals with PD and healthy controls (Hasegawa et al., 2015; Keshavarzian et al., 2015; Scheperjans et al., 2015; Unger et al., 2016). Yet, how dysbiosis arises and whether this feature contributes to PD pathogenesis remains unknown.
Gut bacteria control the differentiation and function of immune cells in the intestine, periphery and brain (Erny et al., 2015; Matcovitch-Natan et al., 2016; Rooks and Garrett, 2016). Intriguingly, subjects with PD exhibit intestinal inflammation (Devos et al., 2013), and GI abnormalities such as constipation often precede motor defects by many years (Braak et al., 2003; Verbaan et al., 2007). Braak's hypothesis posits that aberrant αSyn accumulation initiates in the gut and propagates via the vagus nerve to the brain in a prion-like fashion (Del Tredici and Braak, 2008). This notion is supported by pathophysiologic evidence: αSyn inclusions appear early in the enteric nervous system (ENS) and the glossopharyngeal and vagal nerves (Braak et al., 2003; Shannon et al., 2012), while vagotomized individuals are at reduced risk for PD (Svensson et al., 2015). Further, injection of αSyn fibrils into the gut tissue of healthy rodents is sufficient to induce pathology within the vagus nerve and brainstem (Holmqvist et al., 2014). However, the notion that αSyn aggregation initiates in the ENS and spreads to the CNS via retrograde transmission remains controversial (Burke et al., 2008), and experimental support for a gut microbial connection to PD is lacking.
Based on the common occurrence of GI symptoms in PD, dysbiosis among PD patients, and evidence that the microbiota impacts CNS function, we tested the hypothesis that gut bacteria regulate the hallmark motor deficits and pathophysiology of synucleinopathies. Herein, we report that the microbiota is necessary to promote αSyn pathology, neuroinflammation, and characteristic motor features in a validated mouse model. We identify specific microbial metabolites that are sufficient to promote disease symptoms. Remarkably, fecal microbes from PD patients impair motor function significantly more than microbiota from healthy controls when transplanted into mice. Together, these results suggest that gut microbes may play a critical and functional role in the pathogenesis of synucleinopathies such as PD.
Results
Gut Microbes Promote Motor and GI Dysfunction
The Thy1-αSyn (ASO; alpha-synuclein overexpressing) mouse displays progressive deficits in fine and gross motor function, as well as gut motility defects (Chesselet et al., 2012; Rockenstein et al., 2002). Recent evidence has linked unregulated αSyn expression in humans to a higher risk of PD (Soldner et al., 2016), providing an epidemiological foundation for the Thy1-αSyn mouse model. Defects in coordinated motor tasks become evident at 12 weeks of age (Fleming et al., 2004). Motor function was measured via four tests: beam traversal, pole descent, nasal adhesive removal, and hindlimb clasping reflexes, as previously validated in this model (Fleming et al., 2004). 12-13 week old ASO animals harboring a complex microbiota (SPF-ASO) require significantly more time to cross a challenging beam compared to wild-type littermates (SPF-WT), and also exhibit increased time to descend a pole, two measures of gross motor function (Figures 1A, B). Removal of an adhesive from the nasal bridge, a test of fine motor control, is impaired in SPF-ASO mice compared to SPF-WT mice (Figure 1C). Finally, the hindlimb clasping reflex, a measure of striatal dysfunction (Zhang et al., 2014), is defective in SPF-ASO mice (Figure 1D). To assess the contribution of gut bacteria, we re-derived ASO mice (GF-ASO) and wild-type mice (GF-WT) under germ-free conditions. Remarkably, 12-13 week old GF-ASO animals exhibit reduced deficits in beam traversal, pole descent, adhesive removal, and hindlimb clasping (Figures 1A-D). In fact, the execution of motor function tasks by GF-ASO mice resembles performance levels of WT animals in many cases. GF-ASO mice do not exhibit differences in weight compared to SPF-ASO animals (Figure S1A), while both SPF-ASO and GF-ASO animals display defects in the inverted grid assay, a measure of limb strength (Figure S1B)—thus, outcomes in motor tests are not due to weight or physical strength. At a later age (24-25 weeks old), SPF-ASO animals exhibit a progressive decline in motor function (Figures S1C-G), which is significantly delayed in GF-ASO animals (Figures S1C-G). We do not observe consistent differences in motor tasks between GF-WT and SPF-WT animals, providing evidence for gene-microbiome interactions.
Figure 1. Gut microbes promote motor and gastrointestinal dysfunction.

(A) Time to traverse beam apparatus
(B) Time to descend pole
(C) Time to remove adhesive from nasal bridge
(D) Hind-limb clasping reflex score
(E) Time course of fecal output in a novel environment over 15 minutes
(F) Total fecal pellets produced in 15 minutes
Animals were tested at 12-13 weeks of age. N=4-6, error bars represent the mean and standard error from 3 trials per animal. Data are representative of 2 experiments. *p ≤ 0.05; **p≤ 0.01; ***p≤ 0.001; ****p≤ 0.0001. SPF=specific pathogen free, GF=germ-free, WT=wild-type, ASO=Thy1-α-synuclein genotype. See also Figure S1.
As in PD, motor dysfunction in this mouse model co-occurs with decreased GI function and constipation (Verbaan et al., 2007; Wang et al., 2012). In SPF-ASO animals, we observe a marked decrease in the total output of fecal pellets, at both 12-13 weeks and 24-25 weeks of age, while fecal output is unaltered in GF-ASO animals (Figures 1E, F and Figures S1H, I). Further, fecal pellets produced by SPF-ASO mice contain reduced water content compared to GF-ASO mice (Figure S1J), together revealing reduced GI defects in GF animals. Indeed, compilation of all motor phenotypes into a principle component analysis (PCoA) displays a striking segregation by the SPF-ASO group, while GF-ASO animals cluster more similarly to WT mice (Figure S1K). Together, these data demonstrate that the presence of gut microbes promote the hallmark motor and intestinal dysfunction in a preclinical model of PD.
The Gut Microbiota is Required for αSyn Pathology
Motor deficits in PD coincide with the aggregation of αSyn. Utilizing an antibody that recognizes only conformation-specific αSyn aggregates and fibrils, we performed immunofluorescence microscopy to visualize αSyn inclusions in the brains of mice. Under SPF conditions, we observe notable aggregation of αSyn in the caudoputamen (CP) and substantia nigra (SN) of ASO animals (Figures 2A and B), brain regions of the nigrostriatal pathway affected in both mouse models and human PD (Brettschneider et al., 2015). Surprisingly, GF-ASO mice display appreciably less αSyn aggregates (Figures 2A and B). To quantify αSyn aggregation, we performed Western blots of brain extracts (Figure 2C). We reveal significantly less insoluble αSyn in brains of GF-ASO animals (Figures 2C-E). To further confirm these findings, we performed dot blot analysis for aggregated αSyn in the CP and inferior midbrain, where the SN is located, and observe similarly decreased αSyn aggregation in GF-ASO animals (Figures S2 A-C). Interestingly, we observe regional specificity of αSyn aggregation: in the frontal cortex (FC), GF-ASO animals harbor fewer αSyn aggregation than SPF animals, while in the cerebellum (CB), we observe nearly equal quantities of αSyn in SPF and GF mice (Figures S2D-H). To ensure that these findings do not reflect differences in transgene expression, we report similar levels of αSyn transcript and protein in the inferior midbrain and the CP between SPF and GF ASO animals (Figure 2F and G). These data suggest that the microbiota regulates pathways that promote αSyn aggregation and/or prevent the clearance of insoluble protein aggregates.
Figure 2. αSyn pathology is increased in mice harbouring a gut microbiota.
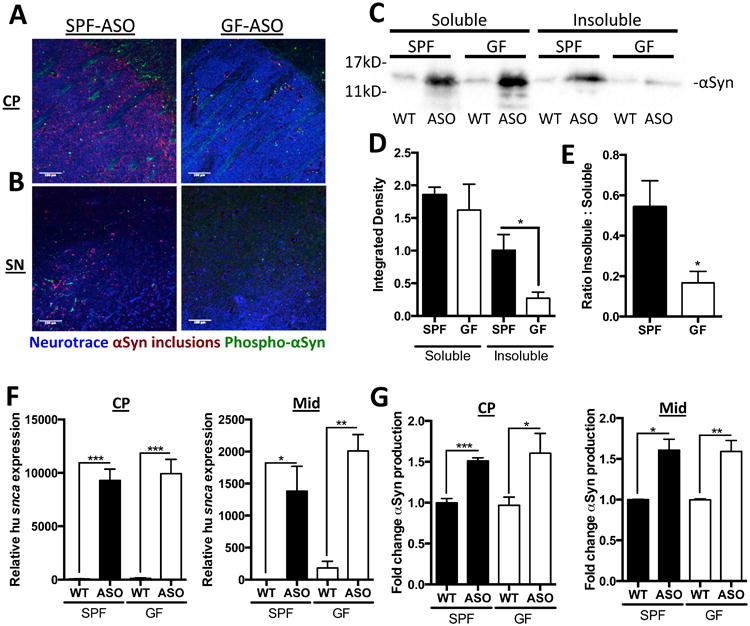
(A) Representative images of the caudoputamen (CP) from SPF-ASO or GF-ASO animals stained with aggregation-specific αSyn antibody (red), Phospho-Ser129-αSyn antibody (green), and Neurotrace/Nissl (blue)
(B) Representative images of the substantia nigra (SN) from SPF-ASO or GF-ASO animals, stained as above
(C) Representative western blot of triton soluble and insoluble brain homogenates, immunostained with anti-αSyn antibody
(D, E) Densitometry quantification of anti-αSyn western blots for (D) all αSyn and (E) ratio of insoluble to soluble αSyn staining
(F) qRT-PCR analysis of human αSyn in the CP or inferior midbrain (Mid)
(G) ELISA analysis of total αSyn present in homogenates from the the CP or inferior midbrain (Mid)
Tissues collected from mice at 12-13 weeks of age. N=3-4, error bars represent the mean and standard error. *p ≤ 0.05; **p≤ 0.01; ***p≤ 0.001. SPF=specific pathogen free, GF=germ-free, WT=wild-type, ASO=Thy1-α-synuclein genotype. See also Figure S2.
αSyn-Dependent Microglia Activation by the Microbiota
The microbiota modulates immune development in the CNS (Erny et al., 2015; Matcovitch-Natan et al., 2016), and αSyn aggregates activate immune cells, including brain-resident microglia (Kim et al., 2013; Sanchez-Guajardo et al., 2013). Microglia undergo significant morphological changes upon activation, transitioning from thin cell bodies with numerous branched extensions to round, amoeboid cells with fewer branches (Erny et al., 2015). In situ 3D reconstructions of individual microglia cells from confocal fluorescence microscopy reveals that wild-type GF animals harbor microglia that are distinct from SPF animals. Within the CP and SN, microglia in GF-WT mice display increased numbers and total lengths of microglia branches compared to SPF-WT animals (Figures 3A-C). These morphological features are indicative of an arrest in microglia maturation and/or a reduced activation state in GF animals, corroborating a recent report that gut bacteria affect immune cells in the brain (Erny et al., 2015).
Figure 3. αSyn-dependent microglia activation by the microbiota.
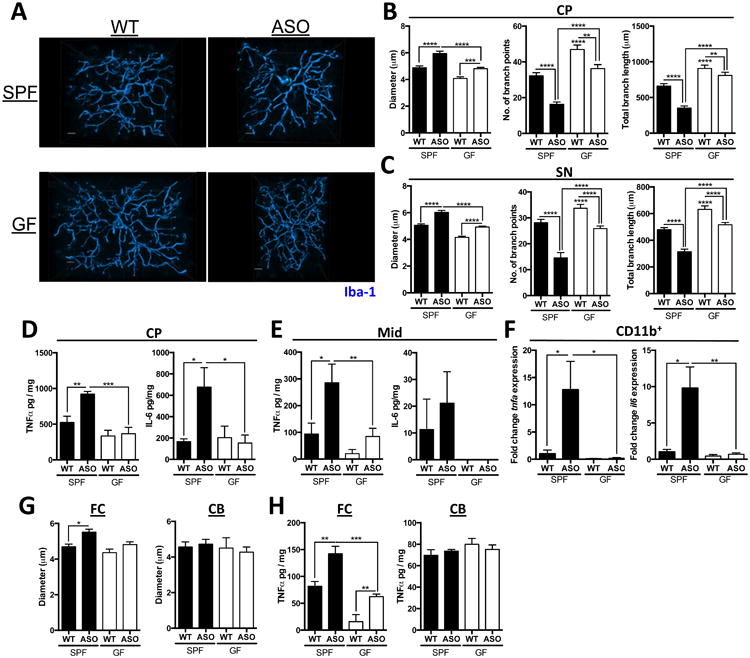
(A) Representative 3D reconstructions of Iba1-stained microglia residing in the caudoputamen (CP) of SPF-WT, SPF-ASO, GF-WT, and GF-ASO animals
(B) CP-resident microglia parameters diameter, number of branch points, and total branch length
(C) Substantia nigra (SN)-resident microglia parameters diameter, number of branch points, and total branch length
(D) ELISA analysis for TNF-α and IL-6 present in homogenates from the CP
(E) ELISA analysis for TNF-α and IL-6 present in homogenates from the inferior midbrain (Mid)
(F) qPCR analysis of CD11b+ cells derived from brain homogenate for tnfa and il6
(G) Diameter of microglia residing in the frontal cortex (FC) or cerebellum (CB)
(H) ELISA analysis for TNF-α present in homogenates from the FC or CB
Tissues collected from mice at 12-13 weeks of age. N=3-4, (with 20-60 cells per region per animal analyzed) error bars represent the mean and standard error. *p ≤ 0.05; **p≤ 0.01; ***p≤ 0.001; ****p≤ 0.0001. SPF=specific pathogen free, GF=germ-free, WT=wild-type, ASO=Thy1-α-synuclein genotype. See also Figure S2.
Extending these observations to a disease model, microglia from SPF-ASO mice display significant increases in cell body diameter, along with fewer processes of shorter length compared to GF-ASO mice (Figures 3A-C). Tissue homogenates from the CP and inferior midbrain of SPF-ASO mice contain a marked increase in the pro-inflammatory cytokines tumor necrosis factor-α (TNFα) and interleukin-6 (IL-6) compared to GF-ASO mice (Figures 3D and E). Both cytokines are elevated in the brains of PD patients (Mogi et al., 1994a; Mogi et al., 1994b). Gene expression analysis of RNA from enriched CD11b+ cells (primarily microglia) reveals increased tnfa and il6 expression in SPF-ASO animals, which is nearly absent in GF animals (Figure 3F). Neuroprotective bdnf and the cell cycle marker ddit4 levels are upregulated in GF animals (Figure S2I), as observed in previous studies (Erny et al., 2015; Matcovitch-Natan et al., 2016). Neuroinflammatory responses are region specific with increased in microglia diameter and TNFα production in the FC, but not the CB (Figures 3G and H). Overall, these findings support the hypothesis that gut microbes promote αSyn-dependent activation of microglia within specific brain regions involved in disease.
Postnatal Microbial Signals Modulate αSyn-Dependent Pathophysiology
The microbiota influence neurological outcomes during gestation, as well as via active gut-to-brain signaling in adulthood. In order to differentiate between these mechanisms, we treated SPF animals with an antibiotic cocktail to postnatally deplete the microbiota (Figure 4A). Conversely, we colonized groups of 5-6 week old GF mice with a complex microbiota from SPF-WT animals (Figure 4A). Remarkably, antibiotic-treated (Abx) animals display little αSyn-dependent motor dysfunction, closely resembling mice born under GF conditions (Figures 4B-E). Postnatal colonization of previously GF animals (Ex-GF) recapitulates the genotype effect observed in SPF mice, with mice that overexpress αSyn displaying significant motor dysfunction (Figures 4B-E). GI function, as measured by fecal output, is also significantly improved in Abx-treated animals, while Ex-GF mice exhibit an αSyn-dependent decrease in total fecal output (Figures 4F, G). Furthermore, in the transgenic ASO line, microglia from Ex-GF animals have increased cell body diameters comparable to those in SPF mice (Figures 4H and I). Abx-ASO animals, however, harbor microglia with diameters similar to GF animals (Figures 4H and I). While not excluding a role for the microbiota during prenatal neurodevelopment, modulation of microglia activation during adulthood contributes to αSyn-mediated motor dysfunction and neuroinflammation, suggesting active gut-brain signaling by the microbiota.
Figure 4. Postnatal microbial signals promote motor and gastrointestinal dysfunction.
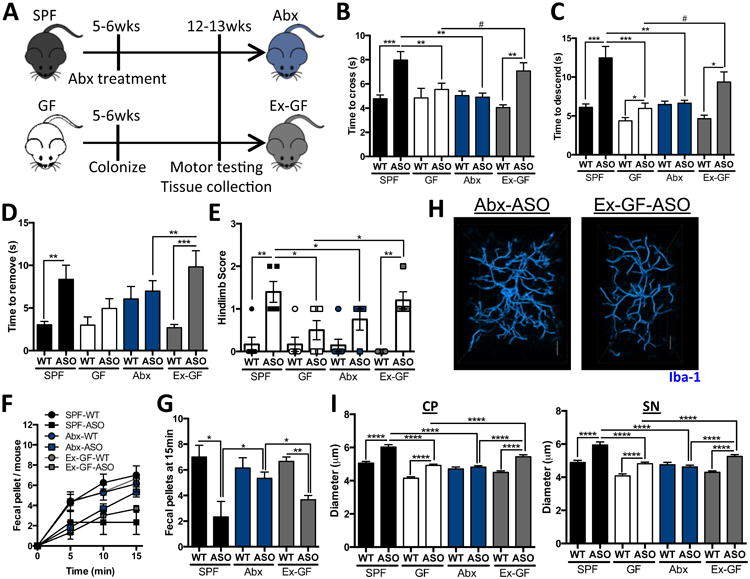
(A) Time course schema for animal treatment and testing
(B) Time to traverse beam apparatus
(C) Time to descend pole
(D) Time to remove nasal adhesive
(E) Hindlimb clasping reflex score
(F) Time course of fecal output in a novel environment over 15 minutes
(G) Total fecal pellets produced in 15 minutes
(H) Representative 3D reconstructions of Iba1-stained microglia residing in the caudoputamen (CP) of Abx-ASO or Ex-GF-ASO animals
(I) Diameter of microglia residing in the CP or substantia nigra (SN)
Animals were tested at 12-13 weeks of age. N=6-12, error bars represent the mean and standard error from 3 trials per animal, and compiled from 2 independent cohorts or 20-60 microglia per region analyzed. #0.05<p<0.1; *p ≤ 0.05; **p≤ 0.01; ***p≤ 0.001; ****p≤ 0.0001. SPF=specific pathogen free, GF=germ-free, Abx=antibiotic-treated, Ex-GF=recolonized germ-free animals, WT=wild-type, ASO=Thy1-α-synuclein genotype. See also Figure S3.
SCFAs are Sufficient to Promote αSyn-Mediated Neuroinflammation
Recently, it was revealed that gut bacteria modulate microglia activation during viral infection through production of microbial metabolites, namely short-chain fatty acids (SCFA) (Erny et al., 2015). Indeed, we observe lower fecal SCFA concentrations in GF and Abx-treated animals, compared to SPF mice (Figure S3A) (Smith et al., 2013). To address whether SCFAs impact neuroimmune responses in a mouse model of PD, we treated GF-ASO and GF-WT animals with a mixture of the SCFAs acetate, propionate, and butyrate (while the animals remained microbiologically sterile), and significantly restored fecal SCFA concentrations (Figure S3A). Within affected brain regions (i.e., CP and SN), microglia in SCFA-administered animals display morphology indicative of increased activation compared to untreated mice, and similar to cells from Ex-GF and SPF mice (Figures 5A and B, Figures S3B and C, see Figures 3 and 4). Microglia from GF-ASO mice fed SCFAs (SCFA-ASO) are significantly larger in diameter than those of GF-WT animals treated with SCFAs (SCFA-WT), with a concomitant decrease in the length and total number of branches. Abx-treated animals, however, display microglia morphology similar to GF animals (Figure 5B, Figures S3B and C, see Figures 3 and 4). Changes in microglia diameter are also observed in the FC, but not the CB, demonstrating region-specific responses (Figures S3D and E).
Figure 5. SCFAs promote αSyn-stimulated microglia activation and motor dysfunction.
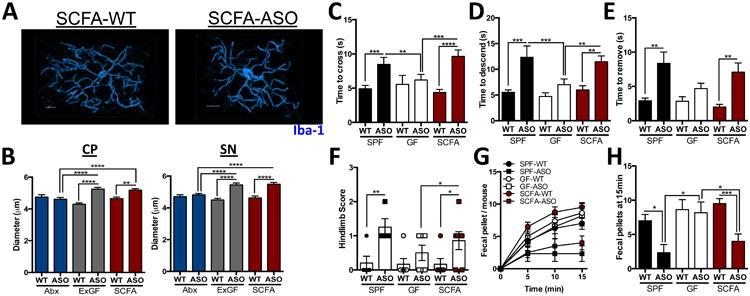
(A) Representative 3D reconstructions of Iba1-stained microglia residing in the caudoputamen (CP) of wild-type or ASO SCFA-treated animals
(B) Diameter of microglia residing in the CP or substantia nigra (SN)
(C) Time to traverse beam apparatus
(D) Time to descend pole
(E) Time to remove nasal adhesive
(F) Hindlimb clasping reflex score
(G) Time course of fecal output in a novel environment over 15 minutes
(H) Total fecal pellets produced in 15 minutes
Animals were tested at 12-13 weeks of age. N=6-12, error bars represent the mean and standard error from 3 trials per animal, and compiled from 2 independent cohorts or 20-60 microglia per region analyzed. Data are plotted with controls from Figure 4 for clarity. *p ≤ 0.05; **p≤ 0.01; ***p≤ 0.001; ****p≤ 0.0001. SPF=specific-pathogen free, GF=germ-free, SCFA=short-chain fatty acid-treated, WT=wild-type, ASO=Thy1-α-synuclein genotype. See also Figures S3-5.
Corresponding to microglia morphology, we reveal αSyn aggregates in mice administered SCFAs compared to untreated and Abx-treated mice, and similar to Ex-GF animals (Figures S3F-I). Strikingly, we observe that postnatal signaling by microbes induces increased αSyn aggregation in the CP, SN (Figures S3F and G), with no observable difference in the FC and CB (Figures S3H and I), confirmed by quantification and Western blot (Figures S3J-O). SCFAs either singly or in a mixture, over a range of concentrations, do not expedite the aggregation of human αSyn in vitro (Figures S4A-G); nor do they alter the overall structure of αSyn amyloid fibrils (Figures S4H and I). Though additional studies are needed, it appears SCFAs accelerate in vivo αSyn aggregation, albeit independently of direct molecular interactions.
SCFAs are Sufficient to Promote Motor Deficits
To explore a link between microbial metabolites and motor symptoms in the Thy1-αSyn model, GF animals were treated with the SCFA mixture beginning at 5-6 weeks of age, and motor function assessed at 12-13 weeks of age. SCFA-ASO mice display significantly impaired performance in several motor tasks compared to untreated GF-ASO animals (Figures 5C-F), including impairment in beam traversal, pole descent, and hindlimb reflex (compare GF-ASO to SCFA-ASO). All effects by SCFAs are genotype-specific to the Thy1-αSyn mice. GI deficits are also observed in the SCFA-treated transgenic animals (Figures 5G and H). Oral treatment of GF animals with heat-killed bacteria does not induce motor deficits (Figures S4J-M), suggesting bacteria need to be metabolically active. Additionally, oral treatment of SCFA-fed animals with the anti-inflammatory compound minocycline is sufficient to reduce TNFα production, reduce αSyn aggregation, and improve motor function, without altering transgene expression (Figures S5A-H). We propose that the microbiota actively produce metabolites, such as SCFAs, which are required for microglia activation and αSyn aggregation, contributing to motor dysfunction in a preclinical model of PD.
Dysbiosis of the PD Microbiome
Given recent evidence that PD patients display altered microbiomes (Hasegawa et al., 2015; Keshavarzian et al., 2015; Scheperjans et al., 2015), we sought to determine whether human gut microbes affect disease outcomes when transferred into GF mice. We collected fecal samples from 6 human subjects diagnosed with PD, as well as 6 matched healthy controls (Table S1). To limit confounding effects, only new onset, treatment naïve PD patients with healthy intestinal histology were chosen, among other relevant inclusion and exclusion criteria (see Methods and Resources and Table S1).
Fecal microbiota from PD patients or controls were transplanted into individual groups of GF recipient animals via oral gavage. Fecal pellets were collected from “humanized” mice, bacterial DNA extracted, and 16S rRNA sequencing performed. Sequences were annotated into Operational Taxonomic Units (OTUs), using closed reference picking against the Greengenes database and metagenome function was predicted by PICRUSt. Recipient animal groups were most similar to their respective human donor's profile in unweighted UniFrac (Lozupone and Knight, 2005), based on PCoA (Figures 6A, B). Strikingly, the disease status of the donor had a strong effect on the microbial communities within recipient mice. Humanized mouse groups from PD donors are significantly more similar to each other than to communities transplanted from healthy donors, with this trend persisting when stratified by genetic background (Figures 6C, D). Furthermore, there are significant differences between the healthy and PD donors in the ASO background compared to WT recipients, suggesting genotype effects on microbial community configuration (Figures 6C, D).
Figure 6. Microbiome dysbiosis of PD patient samples after transplant into germ-free mice.
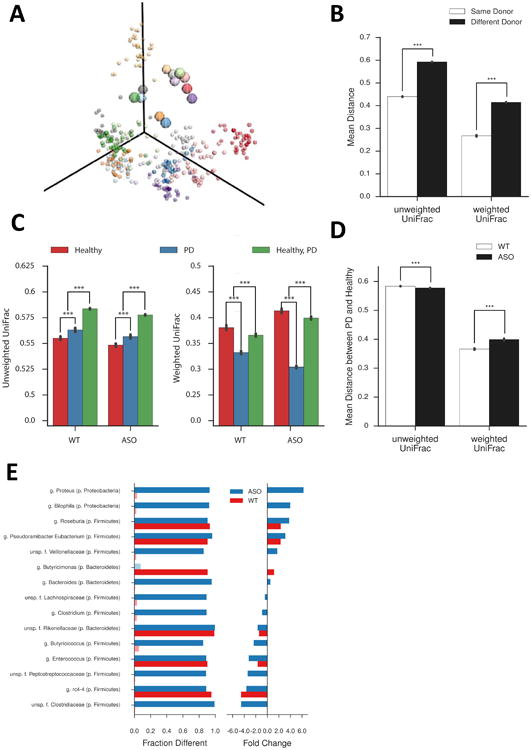
(A) Unweighted UniFrac Principle Coordinate Analysis of microbial communities of human donors (large circles) and recipient mice (small circles). Each donor and recipient sample are matched by color.
(B) Unweighted and weighted UniFrac analysis of microbial communities in recipient animals based on donor identity
(C) Unweighted and weighted UniFrac analysis of microbial communities in recipient animals based on mouse genotype
(D) Comparison of unweighted and weighted UniFrac analysis of microbial communities in recipient animals
(E) Taxa-level analysis of individual genera altered between PD and healthy donors as a function of recipient mouse genotype. Left column indicates percentage with significant differences observed; right column indicates fold change between PD and healthy donors. Light colors indicate non-statistically significant differences
N=3-6, over 3 time points post-colonization. ***p≤ 0.001, 999 permutations. HC=germ-free mice colonized with fecal microbes from healthy controls, PD=germ-free mice colonized with fecal microbes from Parkinson's disease patients, WT=wild-type, ASO=Thy1-α-synuclein genotype. See also Figure S6.
We identified a number of genera that are altered in animals colonized with microbiota derived from PD donors, compared to healthy controls (Figure 6E), as well as altered KEGG pathways between these groups as indicated by Bray-Curtis distances (Figures S6 A-C). OTUs increased in abundance in mice with PD microbiomes include Proteus sp., Bilophila sp., and Roseburia sp., with a concomitant loss of members of families Lachnospiraceae, Rikenellaceae, and Peptostreptococcaceae, as well as Butyricicoccus sp. (Figure 6E). Interestingly, some taxa are altered only in ASO animals (e.g. Proteus sp., Bilophila sp., and Lachnospiraceae), while others display significant changes independent of mouse genotype (e.g. Rosburia sp., Rikenellaceae, and Enterococcus sp.) (Figure 6E). Intriguingly, the abundance of three SCFA-producing KEGG families (K00929, butyrate kinase; K01034 and K01035, acetate CoA/acetoacetate CoA transferase alpha and beta) are increased in mice that received fecal microbes derived from PD donors (Figure S6D). Further, we observe that animals receiving PD donor-derived microbiota display a significantly altered SCFA profile, with a lower concentration of acetate, and higher relative abundances of propionate and butyrate, compared to animals colonized with microbes from healthy controls (Figure S6E). Together, these data indicate that differences in fecal microbial communities between PD patients and controls can be maintained following transfer into mice. Further, αSyn overexpression engenders distinct alterations to the gut microbiome profile following transplantation.
PD-Derived Gut Microbiota Promotes Motor Dysfunction
To assess microbiota function, groups of humanized animals from each of the donor pairs were tested for motor function. Consistent among four of the six pairs (Pairs #1, 3, 4, and 5), microbiota derived from individuals with PD promote increased αSyn-mediated motor dysfunction (Figures 7A-F). Beam traversal, pole descent and nasal adhesive removal are significantly impaired in ASO animals colonized with PD microbiota compared to genotype-matched recipient mice harboring gut bacteria from healthy controls. Hindlimb reflex scores, on the other hand, are generally not different between individual donors. Interestingly, microbiota from one pair of samples did not induce significant genotype effects in the beam traversal and pole descent tasks (Pair #2, Figure 7B), reflecting potential heterogeneity in the population that needs to be addressed through well-powered cohort studies. We observed no notable effects in motor function by WT recipient animals colonized with microbiota from either donor group (Figures 7A-F). This finding in a preclinical mouse model supports the notion the PD microbiota contributes to disease symptoms in genetically susceptible hosts. Notably, recipient animals display little alteration to weight and GI function as measured by fecal output (Figures S7A-F). Compilation of performance data from all groups reveals that microbiota from PD patients induce increased motor impairment in ASO animals compared to microbes from healthy controls in 3 of 4 tests used in this study (Figure 7G). In fact, depicting all motor function by PCoA displays striking global differences between animals colonized with microbiota from PD donors, compared to those colonized with gut bacteria derived from healthy individuals (Figure S7G). The observation that gut bacteria from PD patients compared to healthy controls enhance motor deficits in a mouse model provides evidence for a functional contribution by the microbiota to synuclienopathies.
Figure 7. Microbiota from PD patients induce increased αSyn-mediated motor deficits.

(A-F) Time to cross a beam, time to descend the pole, time to remove nasal adhesive, and hindlimb clasping reflex scores of mice humanized with microbiota from either PD patients or matched healthy controls
(G) Compilation of all independent cohorts in each motor task: beam traversal, pole descent, adhesive removal, and hindlimb clasping reflex score, grouped by health status of fecal donor
Animals were tested at 12-13 weeks of age. N=3-6, error bars represent the mean and standard error from 3 trials per animal. #0.05<p<0.1; *p ≤ 0.05; **p≤ 0.01; ***p≤ 0.001; ****p≤ 0.0001. HC=germ-free mice colonized with fecal microbes from healthy controls, PD=germ-free mice colonized with fecal microbes from Parkinson's disease patients, WT=wild-type, ASO=Thy1-α-synuclein genotype. See also Figure S7 and Table S1.
Discussion
Parkinson's disease represents a growing health concern for an ever-aging population. While genetic risks have been identified, environmental factors and gene-environment interactions likely account for most PD cases (Nalls et al., 2014; Ritz et al., 2016). We provide evidence that the gut microbiota are required for postnatal events that promote hallmark motor deficits in an animal model. Under GF conditions, or when bacteria are depleted with antibiotics, transgenic animals overexpressing human αSyn display reduced microglia activation, αSyn inclusions, and motor deficits compared to animals with a complex microbiota. Treatment with microbially-produced SCFAs restores all major features of disease in GF mice, identifying potential molecular mediators involved in gut-brain signaling. Exacerbated motor symptoms in humanized mice transplanted with a PD microbiota compared to healthy controls suggest that αSyn overexpression (genetics) and dysbiosis (environment) combine to influence disease outcomes in mice. Extrapolation of these preclinical findings to humans may embolden the concept that gene-microbiome interactions represent a previously unrecognized etiology for PD.
Mechanisms by which gut bacteria promote αSyn-mediated pathophysiology are likely complex; herein, we have identified one potential pathway requiring microbiota-dependent effects on microglia. Recent studies have demonstrated an active role for the gut microbiota in promoting full maturation and inflammatory capabilities of microglia through the production of SCFAs (Erny et al., 2015). Despite a requirement for the SCFA receptor FFAR2 for microglia maturation, these cells are not known to express FFAR2, but do express other SCFA responsive genes such as the histone deactylases that modulate gene expression (Erny et al., 2015). SCFAs may cross the BBB and impact the physiology of cells in the CNS (Mitchell et al., 2011), or they may have peripheral effects, which indirectly activate and mature microglia by currently unknown mechanisms (Erny et al., 2015). Further, insoluble aggregates and oligomeric forms of αSyn activate microglia (Kim et al., 2013; Sanchez-Guajardo et al., 2013). Increases in the activation state of microglia and the production of pro-inflammatory cytokines alter neuronal function and increase cell death in models of PD and other neurodegenerative diseases (Kannarkat et al., 2013; Sanchez-Guajardo et al., 2013). Intriguingly, an inflammatory environment is known to enhance αSyn aggregation, which may further activate microglia upon contact and promote a feed-forward cascade that leads to additional αSyn aggregation and propagation, and progression of disease (Gao et al., 2011). If true, possible future treatment options may include targeting immune activation by the microbiota, a notion consistent with research into anti-inflammatory therapeutic modalities for PD (Valera and Masliah, 2016).
While the microbiota promote microglia maturation, there are likely other disease-modifying processes that remain undiscovered. These include effects by the microbiota on autophagy (Lin et al., 2014), a cellular recycling process that is genetically linked to PD risk, and when impaired may lead to reduced clearance of αSyn aggregates (Beilina and Cookson, 2015; Nalls et al., 2014). Additionally, intestinal bacteria have been shown to modulate proteasome function (Cleynen et al., 2014), which may also aid in the clearance of αSyn inclusions. The protective effects of autophagy and the proteasome are not specific to synuclienopathies, and the ability of the microbiota to modulate these critical cellular functions suggests that other amyloid disorders, such as Alzheimer's and Huntington's diseases, may be impacted by gut bacteria. In fact, recent studies have implicated the gut microbiota in promoting amyloid beta pathology in a model of Alzheimer's disease (Minter et al., 2016). Though we have explored postnatal effects of the microbiota in a model of neurodegenerative disease, our findings do not address the likely important role of microbial signals during prenatal neurodevelopment. Whether gut microbes alter the development of the dopaminergic system, perhaps by modulating neurogenesis or neural differentiation in utero or early life, remains unexplored. Furthermore, gut microbes can produce dopamine and its precursors from dietary substrates, with almost half of the body's dopamine generated in the GI tract (Eisenhofer et al., 1997; Wall et al., 2014). Deciphering microbiota effects on microglia activation, cellular protein clearance pathways, neurotransmitter production and/or other mechanisms may offer an integrated approach to understand the pathogenesis of a complex and enigmatic disorder such as PD.
We reveal that gut bacteria from PD patients promote enhanced motor impairment compared to microbiota from healthy controls when transplanted into genetically susceptible ASO mice. This surprising finding suggests that distinct microbes associated with PD, rather than general microbial stimulation, manifest disease symptoms. Several bacterial taxa are altered in mice receiving fecal transplants from PD patients compared to healthy controls. Additionally, a number of bacterial genera are changed specifically in ASO animals, but not WT mice, receiving microbes from the same donor. These include depletions in members of family Lachnospiraceae and Ruminococceae in recipient mice, a notable finding as these same genera are significantly reduced in fecal samples directly from PD patients (Keshavarzian et al., 2015). Conversely, the gut microbiomes in human subjects with PD contain an increased abundance of Proteobacteria (Hasegawa et al., 2015; Keshavarzian et al., 2015; Scheperjans et al., 2015; Unger et al., 2016), remarkably similar to our results in mice. Whether these specific microbes play a role in disease processes remains unknown. Intriguingly, a recent study demonstrated alterations in fecal SCFA ratios between patients and healthy controls, including an elevated relative concentration of butyrate, possibly implicating a role for SCFAs in PD (Unger et al., 2016). Accordingly, we observe altered SCFA abundances in animals colonized with PD donor-derived microbiota, and the discovery that SCFAs are sufficient to generate αSyn-reactive microglia in the brain is consistent with expansive literature showing altered microbial communities impact immune responses in the gut and periphery (Hooper et al., 2012).
What causes dysbiosis in PD? Physiological functions in affected individuals, such as altered intestinal absorption, reduced gastric motility, or dietary habits, represent factors that may change the microbiome. Epidemiological evidence has linked specific pesticide exposure to the incidence of PD (Ritz et al., 2016), with some pesticides known to impact microbiome configuration (Gao et al., 2016). Given the structure of αSyn and its ability to associate with membranes (Jo et al., 2000), it is tempting to speculate that extracellular αSyn may act as an antimicrobial, similar to recent observations with amyloid beta (Kumar et al., 2016), and shape the PD microbiome. Whether microbial community alterations are caused by extrinsic or intrinsic factors, the PD microbiota may be missing or reduced in protective microbes, harbor increased pathogenic resident microbes, or both. In turn, dysbiosis will result in differential production of microbial molecules in the gut. Metabolites produced by a deranged microbiota may enter the circulation (or even the brain) and impact neurological function. Identification of bacterial taxa or microbial metabolites that are altered in PD may serve as disease biomarkers or even drug targets, and interventions that correct dysbiosis may provide safe and effective treatments to slow or halt the progression of often debilitating motor symptoms.
Our findings establish that the microbiota are required for the hallmark motor and GI dysfunction in a mouse model of PD, via postnatal gut-brain signaling by microbial molecules that impact neuroinflammation and αSyn aggregation. Coupled with emerging research that has linked gut bacteria to disorders such as anxiety, depression and autism, we propose the provocative hypothesis that certain neurologic conditions which have classically been studied as disorders of the brain may also have etiologies in the gut.
Contact For Reagent and Resource Sharing
Sarkis K. Mazmanian, California Institute of Technology, sarkis@caltech.edu
Experimental Model and Subject Details
Mice
Female BDF1 background, Thy1-αSyn animals heterozygous for the Thy1-α-synuclein transgene on the X-chromosome were bred with wild-type male BDF1 mice to generate the male ASO and WT littermates used in the study (Chesselet et al., 2012; Rockenstein et al., 2002). Male BDF1 were bred by crossing female C57BL/6 with DBA/2 males (Charles River, Hollister, CA). Breeding pairs were replenished every 6 months with transgenic females and newly generated BDF1 males. Germ-free (GF) Thy1-αSyn breeding pairs were generated via caesarian section and males newly generated every 6 months. Following surgical removal of the uterus and delivery of pups, microbiologically-sterile animals were fostered by GF Swiss-Webster dams. SPF, antibiotic-treated, and ex-GF animals were housed in autoclaved, ventilated, microisolator caging. GF and SCFA-treated animals were housed in open-top caging within flexible film isolators and maintained microbiologically sterile. Microbial sterility was confirmed on a biweekly basis through 16s rRNA PCR from fecal-derived DNA and plating of fecal pellets on Brucella blood agar media under anaerobic conditions and tryptic soy blood agar under aerobic conditions. All animals, irrespective of colonization status, received autoclaved food (LabDiet Laboratory Autoclavable Diet 5010, St Louis, MO) and water ad libitum, were maintained on the same 12-hour light-dark cycle, and housed in the same facility. Antibiotic-treated animals were provided ampicillin (1g/L; Sigma Aldrich, St. Louis, MO), vancomycin (0.5g/L; Sagent Pharmaceuticals, Schaumburg, IL), neomycin (0.5g/L; Fisher Scientific), gentamycin (100mg/L; Sigma Aldrich), and erythromycin (10mg/L; Sigma Aldrich) in drinking water beginning at 5-6 weeks of age through 12-13 weeks of age. Ex-GF animals were generated by colonizing 5-6 week old GF animals with cecal contents from 3 wild-type BDF1 males resuspended in sodium biocarbonate buffer prior to oral gavage. SCFA treated animals were provided with drinking containing sodium acetate (67.5mM; Sigma Aldrich), sodium propionate (25mM; Sigma Aldrich), and sodium butyrate (40mM; Sigma Aldrich) beginning at 5-6 weeks of age until 12-13 weeks of age (Smith et al., 2013). Minocycline (Arcos Organics) treatment was provided in drinking water ad libitum at 2g/L, concurrently with SCFAs from 5-6 weeks of age until 12-13 weeks (Kohman et al., 2013). GF animals treated with heat-killed bacteria were provided ∼5×108 cfu/mL of lysogeny broth (LB)-grown Escherichia coli MC4100 (a kind gift from Matthew Chapman, U. of Michigan), washed twice in phosphate buffered saline and boiled for 45min, in drinking water ad libitum. All animal husbandry and experiments were approved by the California Institute of Technology's Institutional Animal Care and Use Committee (IACUC).
Human Donor and Criteria
Human donors were selected from patients seen at the Movement Disorder Clinic at Rush University. PD was diagnosed according to the UK Brain Bank Criteria. Exclusion criteria for PD subjects: atypical or secondary Parkinsonism; the use of probiotics or antibiotics within three months prior to sample collection; use of NSAIDs; primary gastrointestinal pathology; history of chronic GI illness (including IBD and celiac disease); unstable medical, neurological, or psychiatric illness; low platelet count (<80k); uncorrectable prolonged PT 9>15sec); or history of bleeding that precludes biopsies. All patients had normal mucosa in their rectum and sigmoid by sigmoidoscopy and by H&E histology. Healthy controls were matched as closely as possible to PD patients. Inclusion criteria for healthy subjects: normal physical exam and blood work; no digestive complaints, symptoms, or history of disease; no neurodegenerative disease; no probiotic, antibiotic, NSAIDs, or prescription medication use at least three months prior to sample collection. All human studies were approved under both Rush University and California Institute of Technology Institutional Review Board (IRB).
Method Details
Motor Function and Gastrointestinal Testing
Excluding humanized animals, all motor function assessment was performed in identical, gnotobiotic animal facility. Humanized animals were tested within a laminar-flow biosafety cabinet in the same facility. Motor function for all animals was tested between hours 7 and 9 of the light-phase. All tests were performed similarly to previous studies (Fleming et al., 2004). Beam traversal was performed first, before allowing animals to rest for ∼1hr and testing on pole descent. The following day, adhesive removal and hindlimb scoring was performed. Fecal output was performed within 3 days and immediately prior to tissue collection. Beam traversal- A 1 meter plexiglass beam (Stark's Plastics, Forest Park, OH) was constructed of four segments of 0.25m in length. Each segment was of thinner widths 3.5cm, 2.5cm, 1.5cm, and 0.5cm, with 1cm overhangs placed 1cm below the surface of the beam. The widest segment acted as a loading platform for the animals and the narrowest end placed into home cage. Animals had two days of training to traverse the length of the beam before testing. On the first day of training, animals received 1 trial with the home cage positioned close to the loading platform and guided the animals forward along the narrowing beam. Animals received two more trials with limited or no assistance to encourage forward movement and stability on the beam. On the second day of training, animals had three trials to traverse the beam and generally did not require assistance in forward movement. On the third day, animals were timed over three trials to traverse from the loading platform and to the home cage. Timing began when the animals placed their forelimbs onto the 2.5cm segment and ended when one forelimb reached the home cage. Pole descent- A 0.5m long pole, 1cm in diameter, wrapped with non-adhesive shelf liner to facilitate the animals grip, was placed into the home cage. Animals received two days of training to descend from the top of the pole and into the home cage. On day one of training, animals received 3 trials. The first trial, animals were placed head-down 1/3 the distance above the floor, the second trial from 2/3 the distance, and the third trial animals were placed at the top. The second day of training, animals were given 3 trials to descend, head-down, from the top of the pole. On the test day, animals were placed head-down on the top of the pole and timed to descend back into the home cage. Timing began when the experimenter released the animal and ended when one hind-limb reached the home cage base. Adhesive removal- ¼” round adhesive labels (Avery, Glendale, CA) were placed on the nasal bridge between the nostrils and forehead. Animals were placed into their home cage (with cage mates removed) and timed to completely remove the sticker. Animals were recorded over three trials. Hindlimb clasping reflex scoring- Animals were gently lifted upwards by the mid-section of the tail and observed over ∼5-10 seconds (Zhang et al., 2014). Animals were assigned a score of 0, 1, 2, 3 based on the extent to which the hindlimbs clasped inward. 0, indicating no clasping, was given to animals that freely moved both their limbs and extended them outwards. A score of 1 was assigned to animals which clasped one hindlimb inward for the duration of the restraint or if both legs exhibited partial inward clasping. A score of 2 was given if both legs clasped inward for the majority of the observation, but still exhibited some flexibility. A score of 3 was assigned if animals displayed complete paralysis of hindlimbs that immediately clasped inward and exhibited no signs of flexibility. Inverted grid- Animals were placed in the center of a 30cm by 30cm screen with 1cm wide mesh. The screen was inverted head-over-tail and placed on supports ∼40cm above an open cage with deep bedding. Animals were timed until they released their grip or remained for 60s. Fecal Output- Animals were removed from their home cages and placed into a 12cm × 25cm translucent cylinder. Fecal pellets were counted every 5 minutes, cumulative over 15 minutes. Principal component analysis of all motor function was performed using MATLAB software (MathWorks) using behavioral data collected from subjects that performed at least 3 tasks. Data was centered and standardized (σ = 1) prior to running the pca function. Only PC1 and PC2, which accounted for 70.5% of the variance, were plotted using the corresponding factor loadings for each individual subject.
Immunostaining and Microglia Reconstructions
Animals were sedated with pentobarbital and well-perfused with phosphate-buffered saline, brains were dissected and hemispheres fixed in 4%(w/v) paraformaldehyde. 50 μm saggital sections were generated via vibratome. Free-floating sections were stained with anti-aggregated/fibril αSyn MJFR1 (1:1000; rabbit; AbCam, Cambridge UK), anti-phosphoSer129 αSyn (1:1000; mouse; Biolegend, San Diego, CA), and Neurotrace (Life Technologies, Carlsbad, CA), or with anti-Iba1 (1:1000; rabbit; Wako, Richmond, VA) and subsequently stained with anti-mouse IgG-AF488 and anti-rabbit IgG-AF546 (1:1000; Life Technologies). Sections were mounted with ProFade Diamond (Life Technologies), and imaged with a 10× objective on a Zeiss LSM800 confocal microscope. 2-3 fields per region per animal were imaged and compiled in ImageJ software for analysis. For microglia reconstructions, z-stacks were imaged at 1μm steps and subsequently analyzed using Imaris software, as previously described (Erny et al., 2015). Semi-automated reconstruction of microglia cell bodies and processes were performed, whereby the experimenter designates individual cell bodies and the software quantifies diameter, dendrite length, and branch points from each given cell body. 20-60 cells per region per animal were analyzed.
CD11b enrichment and qPCR analysis
Perfused whole brains were homogenized in PBS via passage through a 100μm mesh filter, myelin debris were removed using magnetic separation with Myelin Removal Beads (Miltenyi Biotec, San Diego CA), according to manufacturer's instructions. CD11b enrichment was performed similarly, with magnetic enrichment by Microglia Microbeads (Miltenyi Biotec), according to manufacturer's instructions. Generally, greater than 90% of cells enriched stained positive for CD11b by immunofluorescence microscopy. For RNA analysis, dissected tissue (frontal cortex, caudoputamen, inferior midbrain, and cerebellum) or CD11b-enriched cell pellets, were lysed in Trizol for DirectZol RNA extraction (Zymo Research, Irvine, CA). cDNA was generated via iScript cDNAsynthesis kit (BioRad, Hercules, CA). qRT-PCR was performed with SybrGreen master mix (Applied Biosystems, Foster City, CA) on an AB 7900ht instrument using primers derived from PrimerBank for the indicated target genes and quantified as ΔΔCT, relative to gapdh (Primers listed in associated Resource Table).
Cytokine and αSyn ELISAs and Western blots
Tissue homogenates were prepared in RIPA buffer containing protease inhibitor cocktail (ThermoFisher, Pittsburgh, PA) and diluted into PBS. TNF-α and IL-6 ELISAs (eBioscience, San Diego, CA) and αSyn ELISAs (ThermoFisher) were performed according to manufacturer's instructions. For dot blot quantification of αSyn fibrils, 1μg of tissue homogenate from the specified region was spotted in 1μL volume aloquats onto 0.45 μm nitrocellulose membranes. For Triton X- soluble vs insoluble fraction western blots, brain hemispheres were homogenized in RIPA buffer containing 1% Triton X-100, centrifuged at 15k × g for 60 min at 4°C to precipitate insoluble proteins from the Triton soluble supernatant. The insoluble fraction was solubilized in 10% sodium dodecyl sulfate, as previously described (Klucken et al., 2006). 5μg of each fraction was separated by 4-20% SDS-PAGE (ThermoFisher), blotted onto PVDF membranes. All membranes were blocked with 5% dry skim milk in Tris-buffered saline with 0.1% Tween-20. Anti-aggregated αSyn antibody (1:2000; rabbit; Abcam) or anti-αSyn (1:1000; mouse; BD) was diluted in skim milk and incubated overnight at 4°C. Membranes were probed with anti-rabbit or anti-mouse IgG HRP (1:1000; Cell Signaling Technology). All blots were detected with the Clarity chemiluminescence substrate (BioRad) on a BioRad GelDoc XR. Densitometry was performed using ImageJ software.
αSyn aggregation assays
For in vitro aggregation kinetics, 70μM αSyn was purified as described previously (Chorell et al., 2015), and incubated in phosphate-buffered saline solution (0.01 M phosphate buffer, 0.0027 M potassium chloride, 0.137M sodium chloride, pH 7.4) in the presence of 12μM of Thioflavin T (ThT; Sigma Aldrich) and increasing concentrations of SCFA. A nonbinding 96-well plate with half area (Corning #3881) was used for each experiment and a 2 mm diameter glass bead was added to each well to accelerate the aggregation. The ThT fluorescence signal was recorded using a microplate reader (Fluostar OPTIMA Microplate reader, BMG Labtech) with the excitation filter of 440 ± 10nm and an emission filter of 490 ± 10nm under intermittent shaking conditions at 37°C. The kinetic curves were normalized to the fluorescence maxima and the time to reach half-maximum intensity quantified. For atomic force microscopy (AFM) imaging, samples were diluted with ultrapure water to ∼3μM total protein concentration, and 50μls were pipetted onto freshly cleaved mica and left to dry. The samples were imaged with a Modular scanning probe microscope NTEGRA Prima (NT-MDT) in intermittent contact mode in air using a gold-coated single crystal silicon cantilever (spring constant of ∼5.1 N/m) with a resonance frequency of ∼150 kHz. AFM images were processed with Gwyddion open source software.
SCFA extraction and analysis
Fecal samples were collected from animals at 12 weeks of age. Each fecal pellet was mixed with 1mL sterile 18 Ω de-ionized water. The pellet-water mixtures were homogenized by mixing at 3200 rpm for five minutes and centrifuged for 15 minutes at 13,000 rpm at 4 °C. Supernatants were filtered using Acrodisc LC 13 mm sterile syringe filters with 0.2 μm PVDF membranes (Pall Life Sciences). The filtrates were used for high performance liquid chromatography (HPLC) analysis. Short chain fatty acids (SCFAs) were analyzed using HPLC (LC-20AT, Shimadzu) equipped with a carbohydrate column (Aminex HPX-87H column, Biorad) and photodiode array detector (PDA, Shimadzu). The eluent was 5 mM H2SO4, fed at a flowrate of 0.6 mL/min, and the column temperature was 50 °C. The run time was 60 minutes. Standard curves were generated by diluting 10 mM volatile fatty acid standard solution (acetic acid, butyric acid, formic acid, valeric acid, isovaleric acid, caproic acid, isocaproic acid, and heptanoic acid) to 50 nM to 5000 nM. Concentrations of SCFAs were normalized to soluble chemical oxygen demand. sCOD values of the fecal supernatants were measured with high range (20-1500 mg/L) Hach COD digestion tubes (Hach Company, Loveland) as recommended by the manufacturer. The wavelength used to measure COD with Hach spectrophotometer was 620 nm.
Microbiome Profiling
Fecal pellets were collected at day 7, 14, 21, and 49 post fecal transplant, from animals housed in groups of 1-3 by genotype and donor. Samples were sequenced according to the Earth Microbiome Project protocols (Gilbert et al., 2014). Briefly, DNA was extracted using a MoBio Power soil kit (Carlsbad, CA), and the V4 region of the 16S rRNA gene was amplified using barcoded primers (Walters et al., 2016). Sequencing was performed using an Illumina MiSeq. Operational Taxonomic Units (OTUs) were picked closed reference using SortMeRNA 2.0 (Kopylova et al., 2012) against the August 2013 release of Greengenes (McDonald et al., 2012) in QIIME 1.9 (Caporaso et al., 2010). The table was rarefied to 7500 sequences per sample for alpha and beta diversity calculations. Differential abundance was performed on a table filtered to exclude samples with less than 7500 sequences. Weighted and unweighted UniFrac (Lozupone and Knight, 2005) distances were calculated in QIIME 1.9. Principle Coordinate Analysis (PCoA) projections were visualized using Emperor 0.9.4 (Vazquez-Baeza et al., 2013). Function was inferred using PICRUSt 1.0 (Langille et al., 2013); predicted functional repertoires were compared using Bray Curtis distance. Significance tests were performed using permanova in scikit-bio 0.4.2 and permutative t tests in QIIME 1.9, both with 999 permutations per test. Differential abundance calculations were performed using genus-level taxa and KEGG-based relative abundance of all counts offset by one. Tests were performed using ANCOM (Mandal et al., 2015) in scikit-bio 0.4.2 with a one-way ANOVA test with a Bonferroni-corrected alpha of 0.1 as the rejection threshold. Mice colonized with samples from healthy donors or donors with PD were compared in the BDF1 or Thy1-αSyn genetic backgrounds. Significantly different taxa were compared between the groups, and classified as significant in both, significant in the Thy1-αSyn background only, or significant in the BDF1 background. Plots were generated using Seaborn 0.7.0.
Quantification and Statistical Analysis
Microbiome population statistics are described in detail above. Excluding these, data sets were analyzed within GraphPad Prism 6 software. Pair-wise comparisons were generated with two-tailed t tests. Comparisons of groups were generated with one-way ANOVA. P –values, n values, definition of center and dispersion measurements are indicated in the associated figure legends for each figure.
Data and Software Availability
16s sequencing data and metadata are available online through the QIITA website (https://qiita.ucsd.edu/), with the study accession #10483 and the EMBL ENA database (http://www.ebi.ac.uk/ena) with the study accession # ERP019564.
Supplementary Material
Related to Figure 1.
(A) Body weight of 12-13 week old animals
(B) Time to fall from inverted grid by 12-13 week old animals
(C) Time to traverse beam by 24-25 week old animals
(D) Time to descend pole by 24-25 week old animals
(E) Time to removal adhesive from nasal bridge by 24-25 week old animals
(F) Hindlimb clasping reflex scores by 24-25 week old animals
(G) Body weight of 24-25 week old animals
(H) Time course of fecal output in a novel environment over 15 minutes by 24-25 week old animals
(I) Total fecal pellets produced in 15 minutes by 24-25 week old animals
(J) Percent water content in fecal pellets from 12-13 week old animals
(K) Principle Component Analysis compiling all motor function scores from SPF-WT, SPF-ASO, GF-WT, and GF-ASO cohorts
N=4-6, error bars represent the mean and standard error. From 3 trials per animal for motor tests. Data are representative of 2 experiments. #0.1>p>0.05; * p≤ 0.05; **p≤ 0.01; ***p≤ 0.001; ****p≤ 0.0001. SPF= specific-pathogen free, GF=germ-free, WT=wild-type, ASO=Thy1-α- synuclein genotype.
(A) Aggregate-specific αSyn dot blots derived from caudoputamen (CP) and inferior midbrain (Mid) homogenates from SPF-ASO and GF-ASO animals
(B, C) Densitometry quantification of dot blots of the (B) CP or (C) inferior midbrain
(D) Representative images of the frontal cortex (FC) from SPF-ASO or GF-ASO animals stained with aggregation-specific αSyn antibody (red), Phospho-Ser129-αSyn antibody (green), and Neurotrace/Nissl (blue).
(E) Representative images of the cerebellum (CB) from SPF-ASO or GF-ASO animals, stained as above.
(F) Dot blot images of homogenates from the FC or CB from SPF-ASO and GF-ASO animals, immunostained with aggregation-specific αSyn antibody
(G, H) Densitometry quantification of dot blots from the (G) FC and (H) CB.
(I) qPCR analysis of CD11b+ cells derived from brain homogenate for bdnf and ddit4
Animals were tested at 12-13 weeks of age. N= 3-4, error bars represent the mean and standard error. *p ≤ 0.05. SPF=specific pathogen free, GF=germ-free, WT=wild-type, ASO=Thy1-α- synuclein genotype.
(A) Fecal concentrations of acetate, propionate, and butyrate, normalized by soluble chemical oxygen demand (sCOD).
(B) Caudoputamen (CP)-resident microglia parameters: number of branch points and total branch length
(C) Substantia nigra (SN)-resident microglia parameters: number of branch points and total branch length
(D, E) Diameter of microglia resident in the (D) frontal cortex (FC) and (E) cerebellum (CB)
(F-I) Representative images of the (F) CP, (G) SN, (H) FC, and (I) CB from Abx-ASO, ex-GF-ASO, or SCFA-ASO animals stained with aggregation-specific αSyn antibody (red), Phospho-Ser129-αSyn antibody (green), and Neurotrace/Nissl (blue)
(J) Dot blot images of CP, inferior midbrain (Mid), FC, and CB homogenate from Abx-ASO, ex-GF-ASO, and SCFA-ASO animals immunostained with aggregation-specific αSyn antibody
(K-N) Densitometry quantification of dot blots from the (K) CP, (L) inferior midbrain, (M) FC, and (N) CB
(O) Western blot for αSyn from Triton soluble and insoluble fractions of CP homogenates derived from Abx- and SCFA-ASO animals.
Animals were tested at 12-13 weeks of age. N=3-6, with 20-60 microglia per region analyzed. Error bars represent the mean and standard error. *p ≤ 0.05; **p≤ 0.01; ***p≤ 0.001; ****p≤ 0.0001. SPF=specific pathogen free, GF=germ-free, Abx=antibiotic-treated animals, Ex-GF=recolonized germ-free animals, SCFA=short-chain fatty acid-treated animals, WT=wild-type, ASO=Thy1-α-synuclein genotype.
Related to Figure 5.
(A-C) αSyn aggregation kinetics, as measured by ThT fluorescence in the presence of the indicated concentrations of (A) sodium acetate, (B) sodium propionate, or (C) sodium butyrate
(D, E) αSyn aggregation kinetics, as measured by ThT fluorescence in the presence of independent mixtures of SCFAs. (D) SCFA Mix 1- 29.6μM acetate, 11μM propionate, and 18.5μM butyrate; SCFA Mix 2- 88.8μM acetate, 33μM propionate, and 55.5μM butyrate. (E) SCFA Mix 3- 0.4mM acetate, 0.15mM propionate, and 0.24mM butyrate; SCFA Mix 4-0.8mM acetate, 0.3mM propionate, and 0.47mM butyrate; SCFA Mix 5- 2.0mM acetate, 0.74mM propionate, and 1.18mM butyrate
(F, G) Time to half-max fluorescence intensity for (F) individual SCFA treatments or (G) SCFA mixtures. N=3, bars represent the mean and standard error.
(H, I) Representative atomic force microscopy of the final product from the above αSyn aggregation assays in the (H) absence of SCFA or (I) presence of SCFA Mix 1
(J) Time to cross beam
(K) Time to descend pole
(L) Time to remove nasal adhesive
(M) Hindlimb reflex scores
Animals were tested at 12-13 weeks of age. N=6-12, error bars represent the mean and standard error from 3 trials per animal. Data are compiled from 2 independent cohorts and plotted with controls from Figure 4 for clarity. *p≤ 0.05; **p≤ 0.01; ***p≤ 0.001; ****p≤ 0.0001. SPF=specific pathogen free; GF=germ-free; HK=heat-killed bacteria-treated; WT=wild-type, ASO=Thy1-α-synuclein genotype.
Related to Figure 5.
(A) ELISA analysis for TNFα present in the caudoputamen (CP) and inferior midbrain (Mid)
(B) Dot blot images of CP, inferior midbrain (Mid) homogenates for aggregate-specific αSyn
(C) Densitometry quantification of dot blots from the CP and inferior midbrain (Mid)
(D) qPCR analysis for human snca expression in total brain homogenate
(E) Time to traverse beam apparatus
(F) Time to descend pole
(G) Time to remove nasal adhesive
(H) Hindlimb clasping reflex score
Animals were tested at 12-13 weeks of age. N=6-12, error bars represent the mean and standard error from 3 trials per animal. Data are compiled from 2 independent cohorts. #0.05<p<0.1; *p ≤ 0.05; **p≤ 0.01; ***p≤ 0.001; ****p≤ 0.0001. SPF=specific pathogen free; GF=germ-free; WT=wild-type, ASO=Thy1-α-synuclein genotype.
Related to Figure 6.
(A) Bray-Curtis distance comparisons between humanized groups
(B, C) Bray-Curtis mean distance comparisons between (B) identical vs different donors, or (C) wild-type (WT) vs Thy1- α-synuclein (ASO) genotypes
(D) PICRUSt analysis for specific KEGG families involved in SCFA production, light circles =healthy control derived microbes, dark circles = PD derived microbes
(E) Fecal concentrations and relative abundances of acetate, propionate, and butyrate from humanized animals, normalized to soluble chemical oxygen demans. Compiled from 6 independent donor pairs, HC= healthy controls; PD= Parkinson's disease.
N=3-6, over 3 time points post-colonization for KEGG analysis, N=21-24 for SCFA abundances. *p ≤ 0.05; **p≤ 0.01; ***p≤ 0.001
Related to Figure 7.
(A-F) Body weight and fecal output following 15min in novel environment for each PD and HC humanized pair. (A) Pair #1, (B) Pair #2, (C) Pair #3, (D) Pair #4, (E) Pair #5, (F) Pair #6. (G) Principle Component Analysis of compiled motor function between humanized animals Animals were tested at 12-13 weeks of age. N=3-6, error bars represent the mean and standard error from 3 trials per animal. #0.05<p<0.1; *p ≤ 0.05; **p≤ 0.01. HC=germ-free mice colonized with fecal microbes from healthy controls, PD=germ-free mice colonized with fecal microbes from Parkinson's disease patients, WT=wild-type, ASO=Thy1-α-synuclein genotype.
Highlights.
Gut microbes promote α-synuclein-mediated motor deficits and brain pathology
Depletion of gut bacteria reduces microglia activation
SCFAs modulate microglia and enhance PD pathophysiology
Human gut microbiota from PD patients induce enhanced motor dysfunction in mice
Acknowledgments
We thank E. Hsiao, M. Sampson, and the Mazmanian laboratory for helpful critiques. We are grateful to K. Ly, A. Maskell, M. Quintos for animal husbandry, Y. Garcia-Flores G. Ackermann, G. Humphrey, and H. Derderian for technical support. Imaging and analysis was performed in the Caltech Biological Imaging Facility, with the support of the Caltech Beckman Institute and the Arnold and Mabel Beckman Foundation. T.R.S. is a Larry L. Hillblom Foundation postdoctoral fellow. This project was supported by funds from the Knut and Alice Wallenberg Foundation and Swedish Research Council to P.W.S.; a gift from Mrs. and Mr. Larry Field to A. K.; the Heritage Medical Research Institute to V.G. and S.K.M.; and a National Institutes of Health grant NS085910 to S.K.M.
Footnotes
Author Contributions: Conceptualization, T.R.S., C.E.S., M.F.C., and S.K.M; Formal analysis, J.W.D., S.J., and C.C.; Investigation, T.R.S., T.T., G.G.S., Z.E.I., and S.R.; Resources, A.K. and K.M.S; Writing-Original Draft, T.R.S. and S.K.M; Writing-Review and Editing, all authors; Supervision, V.G., R.K.B., P.W.S., R.K., and S.K.M; Funding Acquisition, T.R.S., V.G., M.F.C., A.K., P.W.S., R.K., and S.K.M
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Beilina A, Cookson MR. Genes associated with Parkinson's disease: regulation of autophagy and beyond. J Neurochem. 2015 doi: 10.1111/jnc.13266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bercik P, Denou E, Collins J, Jackson W, Lu J, Jury J, Deng Y, Blennerhassett P, Macri J, McCoy KD, et al. The intestinal microbiota affect central levels of brain-derived neurotropic factor and behavior in mice. Gastroenterology. 2011;141:599–609. 609 e591–593. doi: 10.1053/j.gastro.2011.04.052. [DOI] [PubMed] [Google Scholar]
- Braak H, Rub U, Gai WP, Del Tredici K. Idiopathic Parkinson's disease: possible routes by which vulnerable neuronal types may be subject to neuroinvasion by an unknown pathogen. Journal of neural transmission. 2003;110:517–536. doi: 10.1007/s00702-002-0808-2. [DOI] [PubMed] [Google Scholar]
- Braniste V, Al-Asmakh M, Kowal C, Anuar F, Abbaspour A, Toth M, Korecka A, Bakocevic N, Ng LG, Kundu P, et al. The gut microbiota influences blood-brain barrier permeability in mice. Sci Transl Med. 2014;6:263ra158. doi: 10.1126/scitranslmed.3009759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brettschneider J, Del Tredici K, Lee VM, Trojanowski JQ. Spreading of pathology in neurodegenerative diseases: a focus on human studies. Nat Rev Neurosci. 2015;16:109–120. doi: 10.1038/nrn3887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burke RE, Dauer WT, Vonsattel JP. A critical evaluation of the Braak staging scheme for Parkinson's disease. Annals of neurology. 2008;64:485–491. doi: 10.1002/ana.21541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, Fierer N, Pena AG, Goodrich JK, Gordon JI, et al. QIIME allows analysis of high-throughput community sequencing data. Nat Methods. 2010;7:335–336. doi: 10.1038/nmeth.f.303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chesselet MF, Richter F, Zhu C, Magen I, Watson MB, Subramaniam SR. A progressive mouse model of Parkinson's disease: the Thy1-aSyn (“Line 61”) mice. Neurotherapeutics. 2012;9:297–314. doi: 10.1007/s13311-012-0104-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chorell E, Andersson E, Evans ML, Jain N, Gotheson A, Aden J, Chapman MR, Almqvist F, Wittung-Stafshede P. Bacterial Chaperones CsgE and CsgC Differentially Modulate Human alpha-Synuclein Amyloid Formation via Transient Contacts. PLoS One. 2015;10:e0140194. doi: 10.1371/journal.pone.0140194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clarke G, Grenham S, Scully P, Fitzgerald P, Moloney RD, Shanahan F, Dinan TG, Cryan JF. The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Mol Psychiatry. 2013;18:666–673. doi: 10.1038/mp.2012.77. [DOI] [PubMed] [Google Scholar]
- Cleynen I, Vazeille E, Artieda M, Verspaget HW, Szczypiorska M, Bringer MA, Lakatos PL, Seibold F, Parnell K, Weersma RK, et al. Genetic and microbial factors modulating the ubiquitin proteasome system in inflammatory bowel disease. Gut. 2014;63:1265–1274. doi: 10.1136/gutjnl-2012-303205. [DOI] [PubMed] [Google Scholar]
- Del Tredici K, Braak H. A not entirely benign procedure: progression of Parkinson's disease. Acta neuropathologica. 2008;115:379–384. doi: 10.1007/s00401-008-0355-5. [DOI] [PubMed] [Google Scholar]
- Devos D, Lebouvier T, Lardeux B, Biraud M, Rouaud T, Pouclet H, Coron E, Bruley des Varannes S, Naveilhan P, Nguyen JM, et al. Colonic inflammation in Parkinson's disease. Neurobiol Dis. 2013;50:42–48. doi: 10.1016/j.nbd.2012.09.007. [DOI] [PubMed] [Google Scholar]
- Diaz Heijtz R, Wang S, Anuar F, Qian Y, Bjorkholm B, Samuelsson A, Hibberd ML, Forssberg H, Pettersson S. Normal gut microbiota modulates brain development and behavior. Proc Natl Acad Sci U S A. 2011;108:3047–3052. doi: 10.1073/pnas.1010529108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dinan TG, Cryan JF. The impact of gut microbiota on brain and behaviour: implications for psychiatry. Curr Opin Clin Nutr Metab Care. 2015;18:552–558. doi: 10.1097/MCO.0000000000000221. [DOI] [PubMed] [Google Scholar]
- Eisenhofer G, Aneman A, Friberg P, Hooper D, Fandriks L, Lonroth H, Hunyady B, Mezey E. Substantial production of dopamine in the human gastrointestinal tract. J Clin Endocrinol Metab. 1997;82:3864–3871. doi: 10.1210/jcem.82.11.4339. [DOI] [PubMed] [Google Scholar]
- Erny D, Hrabe de Angelis AL, Jaitin D, Wieghofer P, Staszewski O, David E, Keren-Shaul H, Mahlakoiv T, Jakobshagen K, Buch T, et al. Host microbiota constantly control maturation and function of microglia in the CNS. Nat Neurosci. 2015;18:965–977. doi: 10.1038/nn.4030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fleming SM, Salcedo J, Fernagut PO, Rockenstein E, Masliah E, Levine MS, Chesselet MF. Early and progressive sensorimotor anomalies in mice overexpressing wild-type human alpha-synuclein. J Neurosci. 2004;24:9434–9440. doi: 10.1523/JNEUROSCI.3080-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gao B, Bian X, Mahbub R, Lu K. Gender-Specific Effects of Organophosphate Diazinon on the Gut Microbiome and Its Metabolic Functions. Environ Health Perspect. 2016 doi: 10.1289/EHP202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gao HM, Zhang F, Zhou H, Kam W, Wilson B, Hong JS. Neuroinflammation and alpha-synuclein dysfunction potentiate each other, driving chronic progression of neurodegeneration in a mouse model of Parkinson's disease. Environ Health Perspect. 2011;119:807–814. doi: 10.1289/ehp.1003013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilbert JA, Jansson JK, Knight R. The Earth Microbiome project: successes and aspirations. BMC Biol. 2014;12:69. doi: 10.1186/s12915-014-0069-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hasegawa S, Goto S, Tsuji H, Okuno T, Asahara T, Nomoto K, Shibata A, Fujisawa Y, Minato T, Okamoto A, et al. Intestinal Dysbiosis and Lowered Serum Lipopolysaccharide-Binding Protein in Parkinson's Disease. PLoS One. 2015;10:e0142164. doi: 10.1371/journal.pone.0142164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoban AE, Stilling RM, Ryan FJ, Shanahan F, Dinan TG, Claesson MJ, Clarke G, Cryan JF. Regulation of prefrontal cortex myelination by the microbiota. Transl Psychiatry. 2016;6:e774. doi: 10.1038/tp.2016.42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holmqvist S, Chutna O, Bousset L, Aldrin-Kirk P, Li W, Bjorklund T, Wang ZY, Roybon L, Melki R, Li JY. Direct evidence of Parkinson pathology spread from the gastrointestinal tract to the brain in rats. Acta neuropathologica. 2014;128:805–820. doi: 10.1007/s00401-014-1343-6. [DOI] [PubMed] [Google Scholar]
- Hooper LV, Littman DR, Macpherson AJ. Interactions between the microbiota and the immune system. Science. 2012;336:1268–1273. doi: 10.1126/science.1223490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jenner P. Molecular mechanisms of L-DOPA-induced dyskinesia. Nat Rev Neurosci. 2008;9:665–677. doi: 10.1038/nrn2471. [DOI] [PubMed] [Google Scholar]
- Jo E, McLaurin J, Yip CM, St George-Hyslop P, Fraser PE. alpha-Synuclein membrane interactions and lipid specificity. J Biol Chem. 2000;275:34328–34334. doi: 10.1074/jbc.M004345200. [DOI] [PubMed] [Google Scholar]
- Kannarkat GT, Boss JM, Tansey MG. The role of innate and adaptive immunity in Parkinson's disease. J Parkinsons Dis. 2013;3:493–514. doi: 10.3233/JPD-130250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keshavarzian A, Green SJ, Engen PA, Voigt RM, Naqib A, Forsyth CB, Mutlu E, Shannon KM. Colonic bacterial composition in Parkinson's disease. Mov Disord. 2015;30:1351–1360. doi: 10.1002/mds.26307. [DOI] [PubMed] [Google Scholar]
- Kim C, Ho DH, Suk JE, You S, Michael S, Kang J, Joong Lee S, Masliah E, Hwang D, Lee HJ, et al. Neuron-released oligomeric alpha-synuclein is an endogenous agonist of TLR2 for paracrine activation of microglia. Nat Commun. 2013;4:1562. doi: 10.1038/ncomms2534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klucken J, Ingelsson M, Shin Y, Irizarry MC, Hedley-Whyte ET, Frosch M, Growdon J, McLean P, Hyman BT. Clinical and biochemical correlates of insoluble alpha-synuclein in dementia with Lewy bodies. Acta neuropathologica. 2006;111:101–108. doi: 10.1007/s00401-005-0027-7. [DOI] [PubMed] [Google Scholar]
- Kohman RA, Bhattacharya TK, Kilby C, Bucko P, Rhodes JS. Effects of minocycline on spatial learning, hippocampal neurogenesis and microglia in aged and adult mice. Behav Brain Res. 2013;242:17–24. doi: 10.1016/j.bbr.2012.12.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kopylova E, Noe L, Touzet H. SortMeRNA: fast and accurate filtering of ribosomal RNAs in metatranscriptomic data. Bioinformatics. 2012;28:3211–3217. doi: 10.1093/bioinformatics/bts611. [DOI] [PubMed] [Google Scholar]
- Kumar DK, Choi SH, Washicosky KJ, Eimer WA, Tucker S, Ghofrani J, Lefkowitz A, McColl G, Goldstein LE, Tanzi RE, et al. Amyloid-beta peptide protects against microbial infection in mouse and worm models of Alzheimer's disease. Sci Transl Med. 2016;8:340ra372. doi: 10.1126/scitranslmed.aaf1059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Langille MG, Zaneveld J, Caporaso JG, McDonald D, Knights D, Reyes JA, Clemente JC, Burkepile DE, Vega Thurber RL, Knight R, et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat Biotechnol. 2013;31:814–821. doi: 10.1038/nbt.2676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ley RE, Peterson DA, Gordon JI. Ecological and evolutionary forces shaping microbial diversity in the human intestine. Cell. 2006;124:837–848. doi: 10.1016/j.cell.2006.02.017. [DOI] [PubMed] [Google Scholar]
- Lin R, Jiang Y, Zhao XY, Guan Y, Qian W, Fu XC, Ren HY, Hou XH. Four types of Bifidobacteria trigger autophagy response in intestinal epithelial cells. J Dig Dis. 2014;15:597–605. doi: 10.1111/1751-2980.12179. [DOI] [PubMed] [Google Scholar]
- Lozupone C, Knight R. UniFrac: a new phylogenetic method for comparing microbial communities. Appl Environ Microbiol. 2005;71:8228–8235. doi: 10.1128/AEM.71.12.8228-8235.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luk KC, Kehm V, Carroll J, Zhang B, O'Brien P, Trojanowski JQ, Lee VM. Pathological alpha-synuclein transmission initiates Parkinson-like neurodegeneration in nontransgenic mice. Science. 2012;338:949–953. doi: 10.1126/science.1227157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mandal S, Van Treuren W, White RA, Eggesbo M, Knight R, Peddada SD. Analysis of composition of microbiomes: a novel method for studying microbial composition. Microb Ecol Health Dis. 2015;26:27663. doi: 10.3402/mehd.v26.27663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matcovitch-Natan O, Winter DR, Giladi A, Vargas Aguilar S, Spinrad A, Sarrazin S, Ben-Yehuda H, David E, Zelada Gonzalez F, Perrin P, et al. Microglia development follows a stepwise program to regulate brain homeostasis. Science. 2016;353:aad8670. doi: 10.1126/science.aad8670. [DOI] [PubMed] [Google Scholar]
- Mayer EA, Padua D, Tillisch K. Altered brain-gut axis in autism: comorbidity or causative mechanisms? Bioessays. 2014;36:933–939. doi: 10.1002/bies.201400075. [DOI] [PubMed] [Google Scholar]
- McDonald D, Price MN, Goodrich J, Nawrocki EP, DeSantis TZ, Probst A, Andersen GL, Knight R, Hugenholtz P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012;6:610–618. doi: 10.1038/ismej.2011.139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Minter MR, Zhang C, Leone V, Ringus DL, Zhang X, Oyler-Castrillo P, Musch MW, Liao F, Ward JF, Holtzman DM, et al. Antibiotic-induced perturbations in gut microbial diversity influences neuro-inflammation and amyloidosis in a murine model of Alzheimer's disease. Sci Rep. 2016;6:30028. doi: 10.1038/srep30028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitchell RW, On NH, Del Bigio MR, Miller DW, Hatch GM. Fatty acid transport protein expression in human brain and potential role in fatty acid transport across human brain microvessel endothelial cells. J Neurochem. 2011;117:735–746. doi: 10.1111/j.1471-4159.2011.07245.x. [DOI] [PubMed] [Google Scholar]
- Mogi M, Harada M, Kondo T, Riederer P, Inagaki H, Minami M, Nagatsu T. Interleukin-1 beta, interleukin-6, epidermal growth factor and transforming growth factor-alpha are elevated in the brain from parkinsonian patients. Neurosci Lett. 1994a;180:147–150. doi: 10.1016/0304-3940(94)90508-8. [DOI] [PubMed] [Google Scholar]
- Mogi M, Harada M, Riederer P, Narabayashi H, Fujita K, Nagatsu T. Tumor necrosis factor-alpha (TNF-alpha) increases both in the brain and in the cerebrospinal fluid from parkinsonian patients. Neurosci Lett. 1994b;165:208–210. doi: 10.1016/0304-3940(94)90746-3. [DOI] [PubMed] [Google Scholar]
- Mohle L, Mattei D, Heimesaat MM, Bereswill S, Fischer A, Alutis M, French T, Hambardzumyan D, Matzinger P, Dunay IR, et al. Ly6Chi Monocytes Provide a Link between Antibiotic-Induced Change in Gut Microbiota and Adult Hippocampal Neurogenesis. Cell Reports. 2016 doi: 10.1016/j.celrep.2016.04.074. [DOI] [PubMed] [Google Scholar]
- Nalls MA, Pankratz N, Lill CM, Do CB, Hernandez DG, Saad M, DeStefano AL, Kara E, Bras J, Sharma M, et al. Large-scale meta-analysis of genome-wide association data identifies six new risk loci for Parkinson's disease. Nat Genet. 2014;46:989–993. doi: 10.1038/ng.3043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neufeld KM, Kang N, Bienenstock J, Foster JA. Reduced anxiety-like behavior and central neurochemical change in germ-free mice. Neurogastroenterol Motil. 2011;23:255–264. e119. doi: 10.1111/j.1365-2982.2010.01620.x. [DOI] [PubMed] [Google Scholar]
- Prusiner SB, Woerman AL, Mordes DA, Watts JC, Rampersaud R, Berry DB, Patel S, Oehler A, Lowe JK, Kravitz SN, et al. Evidence for alpha-synuclein prions causing multiple system atrophy in humans with parkinsonism. Proc Natl Acad Sci U S A. 2015;112:E5308–5317. doi: 10.1073/pnas.1514475112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ritz BR, Paul KC, Bronstein JM. Of Pesticides and Men: a California Story of Genes and Environment in Parkinson's Disease. Curr Environ Health Rep. 2016;3:40–52. doi: 10.1007/s40572-016-0083-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rockenstein E, Mallory M, Hashimoto M, Song D, Shults CW, Lang I, Masliah E. Differential neuropathological alterations in transgenic mice expressing alpha-synuclein from the platelet-derived growth factor and Thy-1 promoters. J Neurosci Res. 2002;68:568–578. doi: 10.1002/jnr.10231. [DOI] [PubMed] [Google Scholar]
- Rooks MG, Garrett WS. Gut microbiota, metabolites and host immunity. Nat Rev Immunol. 2016;16:341–352. doi: 10.1038/nri.2016.42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sacchettini JC, Kelly JW. Therapeutic strategies for human amyloid diseases. Nat Rev Drug Discov. 2002;1:267–275. doi: 10.1038/nrd769. [DOI] [PubMed] [Google Scholar]
- Sanchez-Guajardo V, Barnum CJ, Tansey MG, Romero-Ramos M. Neuroimmunological processes in Parkinson's disease and their relation to alpha-synuclein: microglia as the referee between neuronal processes and peripheral immunity. ASN Neuro. 2013;5:113–139. doi: 10.1042/AN20120066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scheperjans F, Aho V, Pereira PA, Koskinen K, Paulin L, Pekkonen E, Haapaniemi E, Kaakkola S, Eerola-Rautio J, Pohja M, et al. Gut microbiota are related to Parkinson's disease and clinical phenotype. Mov Disord. 2015;30:350–358. doi: 10.1002/mds.26069. [DOI] [PubMed] [Google Scholar]
- Schroeder BO, Backhed F. Signals from the gut microbiota to distant organs in physiology and disease. Nat Med. 2016;22:1079–1089. doi: 10.1038/nm.4185. [DOI] [PubMed] [Google Scholar]
- Selkrig J, Wong P, Zhang X, Pettersson S. Metabolic tinkering by the gut microbiome: Implications for brain development and function. Gut Microbes. 2014;5:369–380. doi: 10.4161/gmic.28681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shannon KM, Keshavarzian A, Dodiya HB, Jakate S, Kordower JH. Is alpha-synuclein in the colon a biomarker for premotor Parkinson's disease? Evidence from 3 cases. Mov Disord. 2012;27:716–719. doi: 10.1002/mds.25020. [DOI] [PubMed] [Google Scholar]
- Sharon G, Sampson TR, Geschwind DH, Mazmanian SK. The Central Nervous System and the Gut Microbiome. Cell. 2016;167:915–932. doi: 10.1016/j.cell.2016.10.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith PM, Howitt MR, Panikov N, Michaud M, Gallini CA, Bohlooly YM, Glickman JN, Garrett WS. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science. 2013;341:569–573. doi: 10.1126/science.1241165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soldner F, Stelzer Y, Shivalila CS, Abraham BJ, Latourelle JC, Barrasa MI, Goldmann J, Myers RH, Young RA, Jaenisch R. Parkinson-associated risk variant in distal enhancer of alpha-synuclein modulates target gene expression. Nature. 2016;533:95–99. doi: 10.1038/nature17939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sudo N, Chida Y, Aiba Y, Sonoda J, Oyama N, Yu XN, Kubo C, Koga Y. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. The Journal of physiology. 2004;558:263–275. doi: 10.1113/jphysiol.2004.063388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Svensson E, Horvath-Puho E, Thomsen RW, Djurhuus JC, Pedersen L, Borghammer P, Sorensen HT. Vagotomy and subsequent risk of Parkinson's disease. Annals of neurology. 2015;78:522–529. doi: 10.1002/ana.24448. [DOI] [PubMed] [Google Scholar]
- Unger MM, Spiegel J, Dillmann KU, Grundmann D, Philippeit H, Burmann J, Fassbender K, Schwiertz A, Schafer KH. Short chain fatty acids and gut microbiota differ between patients with Parkinson's disease and age-matched controls. Parkinsonism Relat Disord. 2016 doi: 10.1016/j.parkreldis.2016.08.019. [DOI] [PubMed] [Google Scholar]
- Valera E, Masliah E. Combination therapies: The next logical Step for the treatment of synucleinopathies? Mov Disord. 2016;31:225–234. doi: 10.1002/mds.26428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vazquez-Baeza Y, Pirrung M, Gonzalez A, Knight R. EMPeror: a tool for visualizing high-throughput microbial community data. Gigascience. 2013;2:16. doi: 10.1186/2047-217X-2-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verbaan D, Marinus J, Visser M, van Rooden SM, Stiggelbout AM, van Hilten JJ. Patient-reported autonomic symptoms in Parkinson disease. Neurology. 2007;69:333–341. doi: 10.1212/01.wnl.0000266593.50534.e8. [DOI] [PubMed] [Google Scholar]
- Wall R, Cryan JF, Ross RP, Fitzgerald GF, Dinan TG, Stanton C. Bacterial neuroactive compounds produced by psychobiotics. Adv Exp Med Biol. 2014;817:221–239. doi: 10.1007/978-1-4939-0897-4_10. [DOI] [PubMed] [Google Scholar]
- Walters W, Hyde ER, Berg-Lyons D, Ackermann G, Humphrey G, Parada A, Gilbert JA, Jansson JK, Caporaso JG, Fuhrman JA, et al. Improved Bacterial 16S rRNA Gene (V4 and V4-5) and Fungal Internal Transcribed Spacer Marker Gene Primers for Microbial Community Surveys. mSystems. 2016;1 doi: 10.1128/mSystems.00009-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang L, Magen I, Yuan PQ, Subramaniam SR, Richter F, Chesselet MF, Tache Y. Mice overexpressing wild-type human alpha-synuclein display alterations in colonic myenteric ganglia and defecation. Neurogastroenterol Motil. 2012;24:e425–436. doi: 10.1111/j.1365-2982.2012.01974.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yano JM, Yu K, Donaldson GP, Shastri GG, Ann P, Ma L, Nagler CR, Ismagilov RF, Mazmanian SK, Hsiao EY. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell. 2015;161:264–276. doi: 10.1016/j.cell.2015.02.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang J, Saur T, Duke AN, Grant SG, Platt DM, Rowlett JK, Isacson O, Yao WD. Motor impairments, striatal degeneration, and altered dopamine-glutamate interplay in mice lacking PSD-95. J Neurogenet. 2014;28:98–111. doi: 10.3109/01677063.2014.892486. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Related to Figure 1.
(A) Body weight of 12-13 week old animals
(B) Time to fall from inverted grid by 12-13 week old animals
(C) Time to traverse beam by 24-25 week old animals
(D) Time to descend pole by 24-25 week old animals
(E) Time to removal adhesive from nasal bridge by 24-25 week old animals
(F) Hindlimb clasping reflex scores by 24-25 week old animals
(G) Body weight of 24-25 week old animals
(H) Time course of fecal output in a novel environment over 15 minutes by 24-25 week old animals
(I) Total fecal pellets produced in 15 minutes by 24-25 week old animals
(J) Percent water content in fecal pellets from 12-13 week old animals
(K) Principle Component Analysis compiling all motor function scores from SPF-WT, SPF-ASO, GF-WT, and GF-ASO cohorts
N=4-6, error bars represent the mean and standard error. From 3 trials per animal for motor tests. Data are representative of 2 experiments. #0.1>p>0.05; * p≤ 0.05; **p≤ 0.01; ***p≤ 0.001; ****p≤ 0.0001. SPF= specific-pathogen free, GF=germ-free, WT=wild-type, ASO=Thy1-α- synuclein genotype.
(A) Aggregate-specific αSyn dot blots derived from caudoputamen (CP) and inferior midbrain (Mid) homogenates from SPF-ASO and GF-ASO animals
(B, C) Densitometry quantification of dot blots of the (B) CP or (C) inferior midbrain
(D) Representative images of the frontal cortex (FC) from SPF-ASO or GF-ASO animals stained with aggregation-specific αSyn antibody (red), Phospho-Ser129-αSyn antibody (green), and Neurotrace/Nissl (blue).
(E) Representative images of the cerebellum (CB) from SPF-ASO or GF-ASO animals, stained as above.
(F) Dot blot images of homogenates from the FC or CB from SPF-ASO and GF-ASO animals, immunostained with aggregation-specific αSyn antibody
(G, H) Densitometry quantification of dot blots from the (G) FC and (H) CB.
(I) qPCR analysis of CD11b+ cells derived from brain homogenate for bdnf and ddit4
Animals were tested at 12-13 weeks of age. N= 3-4, error bars represent the mean and standard error. *p ≤ 0.05. SPF=specific pathogen free, GF=germ-free, WT=wild-type, ASO=Thy1-α- synuclein genotype.
(A) Fecal concentrations of acetate, propionate, and butyrate, normalized by soluble chemical oxygen demand (sCOD).
(B) Caudoputamen (CP)-resident microglia parameters: number of branch points and total branch length
(C) Substantia nigra (SN)-resident microglia parameters: number of branch points and total branch length
(D, E) Diameter of microglia resident in the (D) frontal cortex (FC) and (E) cerebellum (CB)
(F-I) Representative images of the (F) CP, (G) SN, (H) FC, and (I) CB from Abx-ASO, ex-GF-ASO, or SCFA-ASO animals stained with aggregation-specific αSyn antibody (red), Phospho-Ser129-αSyn antibody (green), and Neurotrace/Nissl (blue)
(J) Dot blot images of CP, inferior midbrain (Mid), FC, and CB homogenate from Abx-ASO, ex-GF-ASO, and SCFA-ASO animals immunostained with aggregation-specific αSyn antibody
(K-N) Densitometry quantification of dot blots from the (K) CP, (L) inferior midbrain, (M) FC, and (N) CB
(O) Western blot for αSyn from Triton soluble and insoluble fractions of CP homogenates derived from Abx- and SCFA-ASO animals.
Animals were tested at 12-13 weeks of age. N=3-6, with 20-60 microglia per region analyzed. Error bars represent the mean and standard error. *p ≤ 0.05; **p≤ 0.01; ***p≤ 0.001; ****p≤ 0.0001. SPF=specific pathogen free, GF=germ-free, Abx=antibiotic-treated animals, Ex-GF=recolonized germ-free animals, SCFA=short-chain fatty acid-treated animals, WT=wild-type, ASO=Thy1-α-synuclein genotype.
Related to Figure 5.
(A-C) αSyn aggregation kinetics, as measured by ThT fluorescence in the presence of the indicated concentrations of (A) sodium acetate, (B) sodium propionate, or (C) sodium butyrate
(D, E) αSyn aggregation kinetics, as measured by ThT fluorescence in the presence of independent mixtures of SCFAs. (D) SCFA Mix 1- 29.6μM acetate, 11μM propionate, and 18.5μM butyrate; SCFA Mix 2- 88.8μM acetate, 33μM propionate, and 55.5μM butyrate. (E) SCFA Mix 3- 0.4mM acetate, 0.15mM propionate, and 0.24mM butyrate; SCFA Mix 4-0.8mM acetate, 0.3mM propionate, and 0.47mM butyrate; SCFA Mix 5- 2.0mM acetate, 0.74mM propionate, and 1.18mM butyrate
(F, G) Time to half-max fluorescence intensity for (F) individual SCFA treatments or (G) SCFA mixtures. N=3, bars represent the mean and standard error.
(H, I) Representative atomic force microscopy of the final product from the above αSyn aggregation assays in the (H) absence of SCFA or (I) presence of SCFA Mix 1
(J) Time to cross beam
(K) Time to descend pole
(L) Time to remove nasal adhesive
(M) Hindlimb reflex scores
Animals were tested at 12-13 weeks of age. N=6-12, error bars represent the mean and standard error from 3 trials per animal. Data are compiled from 2 independent cohorts and plotted with controls from Figure 4 for clarity. *p≤ 0.05; **p≤ 0.01; ***p≤ 0.001; ****p≤ 0.0001. SPF=specific pathogen free; GF=germ-free; HK=heat-killed bacteria-treated; WT=wild-type, ASO=Thy1-α-synuclein genotype.
Related to Figure 5.
(A) ELISA analysis for TNFα present in the caudoputamen (CP) and inferior midbrain (Mid)
(B) Dot blot images of CP, inferior midbrain (Mid) homogenates for aggregate-specific αSyn
(C) Densitometry quantification of dot blots from the CP and inferior midbrain (Mid)
(D) qPCR analysis for human snca expression in total brain homogenate
(E) Time to traverse beam apparatus
(F) Time to descend pole
(G) Time to remove nasal adhesive
(H) Hindlimb clasping reflex score
Animals were tested at 12-13 weeks of age. N=6-12, error bars represent the mean and standard error from 3 trials per animal. Data are compiled from 2 independent cohorts. #0.05<p<0.1; *p ≤ 0.05; **p≤ 0.01; ***p≤ 0.001; ****p≤ 0.0001. SPF=specific pathogen free; GF=germ-free; WT=wild-type, ASO=Thy1-α-synuclein genotype.
Related to Figure 6.
(A) Bray-Curtis distance comparisons between humanized groups
(B, C) Bray-Curtis mean distance comparisons between (B) identical vs different donors, or (C) wild-type (WT) vs Thy1- α-synuclein (ASO) genotypes
(D) PICRUSt analysis for specific KEGG families involved in SCFA production, light circles =healthy control derived microbes, dark circles = PD derived microbes
(E) Fecal concentrations and relative abundances of acetate, propionate, and butyrate from humanized animals, normalized to soluble chemical oxygen demans. Compiled from 6 independent donor pairs, HC= healthy controls; PD= Parkinson's disease.
N=3-6, over 3 time points post-colonization for KEGG analysis, N=21-24 for SCFA abundances. *p ≤ 0.05; **p≤ 0.01; ***p≤ 0.001
Related to Figure 7.
(A-F) Body weight and fecal output following 15min in novel environment for each PD and HC humanized pair. (A) Pair #1, (B) Pair #2, (C) Pair #3, (D) Pair #4, (E) Pair #5, (F) Pair #6. (G) Principle Component Analysis of compiled motor function between humanized animals Animals were tested at 12-13 weeks of age. N=3-6, error bars represent the mean and standard error from 3 trials per animal. #0.05<p<0.1; *p ≤ 0.05; **p≤ 0.01. HC=germ-free mice colonized with fecal microbes from healthy controls, PD=germ-free mice colonized with fecal microbes from Parkinson's disease patients, WT=wild-type, ASO=Thy1-α-synuclein genotype.