

ABSTRACT
Archaeosomes are liposomal vesicles composed of ether glycerolipids unique to the domain of Archaea. Unlike conventional ester-linked liposomes, archaeosomes exhibit high stability and possess strong adjuvant and immunostimulatory properties making them an attractive vaccine delivery vehicle. Traditionally comprised of total polar lipids (TPL) or semi-synthetic phospho-glycerolipids of ether-linked isoprenoid phytanyl cores with varied glycol- and amino-head groups, archaeosomes can induce robust and long-lasting humoral and cell-mediated immune responses against antigenic cargo and provide protection in murine models of infectious disease and cancer. However, traditional TPL archaeosome formulations are relatively complex comprising several lipid species. Semi-synthetic archaeosomes tested previously contain a combination of several phospho-glycolipids (negative and neutral charged) to produce a stable, uniform-sized liposome formulation. Moreover, they involve many synthetic steps to arrive at the final desired glycolipid composition. Herein, we present a novel adjuvant formulation comprising a sulfated saccharide group covalently linked to the free sn-1 hydroxyl backbone of an archaeal core lipid (sulfated S-lactosylarchaeol, SLA). SLA individually or mixed with uncharged glyolipid (lactosylarchaeol, LA) constituted efficacious carrier vesicles for entrapped antigens (ovalbumin or melanoma associated tyrosinase-related protein 2 [TRP-2]) and induction of strong cell-mediated responses in mice and protection against subsequent B16 melanoma tumor challenge. Thus, semi-synthetic sulfated glycolipid archaeosomes represent a new class of adjuvants that will potentially ease manufacturing and scale-up, while retaining immunostimulatory activity.
KEYWORDS: Adjuvant, adjuvants, archaeosome, delivery, glycolipid, immune modulators, immune response, mice, vaccine, vaccinology
Introduction
Immunization has greatly reduced the incidence of mortality and morbidity due to infectious disease worldwide and is estimated to currently prevent between 2 to 3 million deaths annually.1 However, the battle against infectious disease is far from over and a need remains to either develop new prophylactic vaccines (for example against malaria, HIV, West Nile virus, Hepatitis C virus, etc.) or improve existing ones (e.g., influenza). In addition, in recent years there has also been a growing interest in therapeutic vaccines, such as those to treat chronic infections, cancer, etc. Since most antigens on their own are poorly immunogenic, the development of effective anti-tumor or anti-viral disease vaccines requires strong adjuvants to induce robust immune responses. Furthermore, it is conceived that many of these vaccines against intracellular pathogens and/or cancer will need to drive induction of strong cell-mediated immunity.
Ideally, adjuvants should be able to induce strong, rapid and long-lasting immune responses, permit dose sparing, and be simple, safe (including low reactogenicity) and cost-effective with the capability to be used with a broad range of antigen types. However, despite the availability of a broad range of adjuvants for preclinical testing, only a very few adjuvants have been approved for clinical use. Aluminum salts (hydroxide or phosphate) have been widely used in many vaccines for decades, and while they are able to enhance humoral responses do not elicit the strong cell-mediated immune responses required to treat chronic viral infections or cancer. More recently the squalene-based oil-in-water emulsion MF59 (Novartis) and Adjuvant System 04 (AS04) (GlaxoSmithKline), a combination of the toll-like receptor 4 agonist monophosphoryl lipid A (MPL) and aluminum hydroxide, have also been approved which will hopefully facilitate the approval of additional adjuvants for unmet medical needs.2, 3
Archaeosomes are liposomal vesicles constituted with archaeal lipids. Unlike conventional eukaryotic ester linked lipids, archaeal lipids possess fully saturated isoprenoid chains of constant length, with ether linkages to sn-2, 3 carbons of the glycerol backbone.4, 5 In comparison with conventional liposomes, archaeosomes exhibit high thermal and pH stability, low proton permeability, and possess strong adjuvant and immunostimulatory properties, particularly for CD8+ T cell responses.6 Archaeosomes have traditionally been composed of total polar lipids (TPL) extracted from archaea, such as those from the species Methanobrevibacter smithii, and have been shown to promote both MHC class I and MHC class II responses to entrapped antigen resulting in strong humoral and cell-mediated responses, and protection from pathogens such as Listeria monocytogenes as well as solid and metastatic tumors.6, 7, 8 However, TPL archaeosome formulations are relatively complex comprising several lipid species native to the archaeal strain. Sprott et al. have previously reported an improved archaeosome formulation using various semi-synthetic phospho-archaeol-based lipids which can induce strong, robust immune responses, including cell-mediated immunity.9 However, while these semi-synthetic archaeosomes are more easily and reproducibly produced than archaeal total polar lipid formulations, they typically require a combination of several phospho-glycolipids (negative and neutral charged) to produce a stable, uniform-sized liposome formulation, and incorporate ability to stimulate the immune system. Moreover, several phospho-glyco-archaeol lipids involve many complex synthetic steps to arrive at the final desired composition. This potentially may increase cost of manufacture but may also render the archaeosome more prone to enzymatic and chemical degradation via the phosphodiester linkages. Herein, we present a simplified archaeosome formulation, composed of a sulfated saccharide group covalently linked to the free sn-1 hydroxyl backbone of an archaeal core lipid (sulfated S-lactosylarchaeol, SLA) (Fig. 1). SLA individually or mixed with uncharged glycolipid (lactosylarchaeol, LA) constitutes robust delivery vesicles for induction of cell-mediated immunity to entrapped cargo (ovalbumin or melanoma associated tyrosinase-related protein [TRP]) as well as protection from B16 melanoma tumor challenge.
Figure 1.
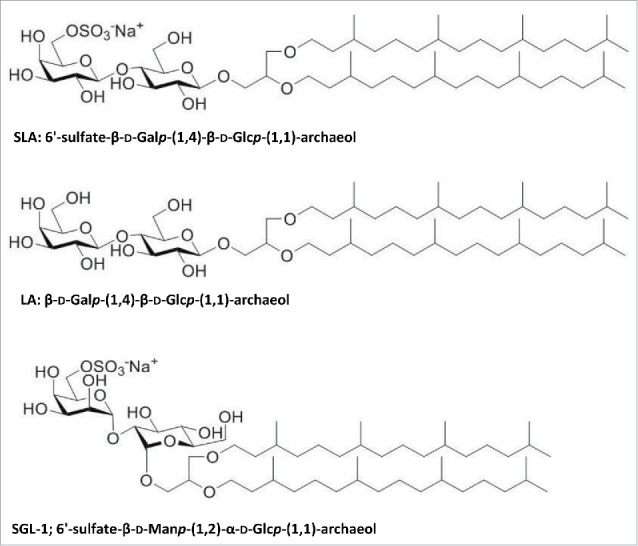
Structures of SLA, LA and SGL-1.
Results
Comparison of synthetic and naturally occurring sulphated glycolipids
Sulfated lactosyl archaeol (SLA) or lactosyl archaeol (LA) were synthesized by chemically adding the glycolipid head-groups to purified archaeol from halophilic archaea and exhibit the structures as outlined in Fig. 1. Archaeosomes prepared using the purified naturally sulfated glycolipid (SGL-1) from strain Haloferax volcanii were compared with those prepared using the semi-synthetic sulfated glycolipid (SLA) as adjuvant for ovalbumin (OVA) in mice. Following SC administration to mice, highest frequency of IFN-γ producing cells were observed in splenocytes from mice that received SLA as adjuvant, and responses were superior to SGL-1 archaeosomes which lacked adjuvant activity (Fig. 2; p ≤ 0.0001). Negative controls, including non-immunized mice (naive) and mice immunized with OVA alone induced only very weak baseline IFN-γ responses. Responses were shown to be antigen specific, since only low levels of IFN-γ were obtained in samples that were not stimulated with antigen (results not shown).
Figure 2.
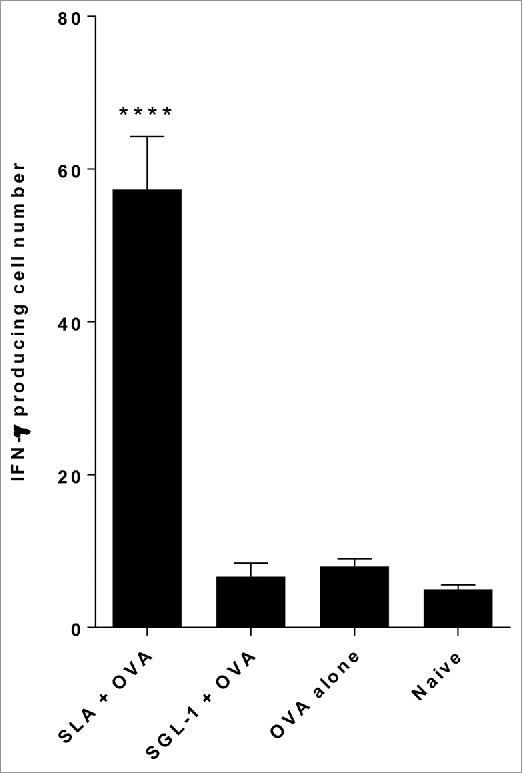
Comparison of CD8+ (T)cell responses induced by OVA entrapped in SLA and SGL-1 archaeosomes. C57BL/6 mice (n = 6/gp) were immunized with 20 µg OVA entrapped in SLA or SGL-1 archaeosomes on days 0 and 21. At 3 weeks post second immunization, representative mice (n = 2 per group) were killed, splenocytes isolated, stimulated with IL-2 (0.1 ng/mL) and OVA257–264 (10 μg/ml) and the frequency of IFN-gamma secreting cells in triplicate cultures enumerated by ELISPOT. Number of spots/106 spleen cells is indicated.
Comparison of SLA and SLA/LA as adjuvants
Preliminary studies demonstrated that archaeosome vesicles did not form using lactosyl-archaeol (LA) alone or other neutrally charged glycolipids (data not shown). In contrast, stable archaeosomes could be readily formed using SLA alone or in combination with LA. When administered to mice with OVA as antigen, SLA/LA archaeosomes induced as strong antigen specific humoral and CD8+ T cell responses as were induced by M. smithii total polar lipid (TPL) archaeosomes. This was demonstrated by equally high levels of OVA-specific IgG, equivalent number of IFN-γ producing cells upon stimulation with OVA-SIINFEKL CD8+ T cell epitope and equivalent functional in-vitro cytotoxic T lymphocyte (CTL) activity at 3 weeks post second immunization (Fig. 3). Archaeosomes comprised of just the single SLA lipid also had a strong adjuvant effect on IFN-γ production and CTL activity, although overall in repeated studies were noted to be slightly weaker than the combination of SLA/LA.
Figure 3.
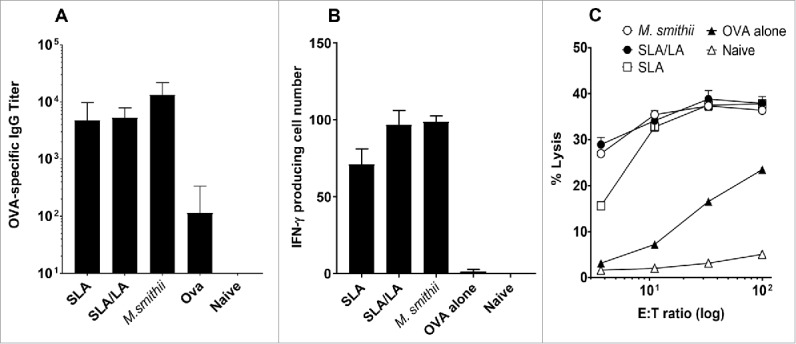
Immune responses induced by OVA entrapped in different archaeosomes. C57/BL6 mice (n = 6/gp) were immunized with 20 µg OVA alone or entrapped in SLA, SLA:LA or M. smithii archaeosomes on days 0 and 21. At 3 weeks post second immunization, serum was collected and OVA-specific IgG titers measured by ELISA (panel A). Representative mice (n = 2 per group) were killed and splenocytes isolated. Pooled splenocytes were stimulated with IL-2 (0.1 ng/mL) and OVA257–264 (10 μg/ml) and the frequency of IFN-gamma secreting cells in triplicate cultures enumerated by ELISPOT. Number of spots/106 spleen cells is indicated (panel B). Splenocytes were stimulated with OVA275–264 for 5 d before assessing CTL activity against 51Cr-labeled targets. CTL data represent percentage of specific lysis of triplicate cultures ± SD at various E:T ratios (panel C).
To determine whether an optimal SLA/LA formulation existed, OVA archaeosomes were prepared using different ratios of SLA:LA and tested in mice. High levels of OVA-specific IgG were induced by all archaeosomes formulations (Fig. 4A). Antigen-specific IFN-γ production by CD8+ T cells was evaluated in an ELISPOT assay (Fig. 4B). At 3 wks post second immunization, an equivalent number of IFN-γ secreting cells were induced by OVA encapsulated in either SLA/LA (50:50) or M. smithii TPL archaeosomes (p = 0.26) which were both significantly higher than all other groups (p < 0.05). The combination of SLA/LA induced equivalent CD8+ T cells responses to SLA alone, provided archaeosomes contained at least 30% SLA.
Figure 4.

Immune responses induced by OVA entrapped in different SLA/LA archaeosomes. C57/BL6 mice (n = 6/gp) were immunized with 20 µg OVA entrapped in SLA, SLA:LA (at various ratios) or M. smithii archaeosomes on days 0 and 21. At 3 weeks post second immunization, serum was collected and OVA-specific IgG titers measured by ELISA (panel A). Representative mice (n = 2 per group) were killed and splenocytes isolated. Pooled splenocytes were stimulated with IL-2 (0.1 ng/mL) and OVA257–264 (10 μg/ml) and the frequency of IFN-gamma secreting cells in triplicate cultures enumerated by ELISPOT. Number of spots/106 spleen cells is indicated (panel B). Splenocytes were also stimulated with OVA275–264 for 5 d before assessing CTL activity against 51Cr-labeled targets. CTL data represent percentage of specific lysis of triplicate cultures ± SD at various E:T ratios (panel C). Zeta potential (mV) was measured as an indicator of archaeosomes stability and plotted relative to %SLA in different archaeosomes (panel D). The zeta potential (mV) of each archaeosome was also plotted against its respective CD8+ T cell response, as measured by the frequency of IFN-gamma secreting cells enumerated by ELISPOT (panel E).
Cytotoxic T lymphocyte activity was measured following re-stimulation of splenocytes with OVA275–264 for 5 d. As expected, strongest CTL activity was measured in splenocytes from mice immunized with OVA encapsulated in M. smithii TPL archaeosomes. Nevertheless, an equivalently strong CTL activity was induced using OVA entrapped in SLA:LA (50:50) archaeosomes. Weaker responses were obtained with other SLA:LA ratios or with SLA alone, although responses were still much greater than obtained with naïve mice (Fig. 4C).
When zeta potential was measured as an indicator of archaeosomes stability, higher amounts of SLA in the archaeosome formulation resulted in an increase in zeta potential suggesting better dispersion stability as the strong electrostatic repulsion interactions will likely prevent aggregation (Table 1, Fig. 4D). Interestingly, when CD8+ responses were evaluated, best responses were obtained with a zeta potential of ∼−40 mV (Fig. 4E), highlighting the importance of archaeosome stability on adjuvanticity.
Table 1.
Archaeosome vaccine characteristics and stability.
| Zeta potential (mV) |
Average Diameter (nm) |
||||
|---|---|---|---|---|---|
| Archaeosome | Loading (μg antigen/mg archaeosome) | At first injection | 12 weeks | At first injection | 12 weeks |
| SLA | 35.0 | −48.1 | −52.7 | 172 | 176 |
| SLA/LA (90/10) | 41.8 | −47.3 | −49.4 | 197 | 190 |
| SLA/LA (70/30) | 12.0 | −43.6 | −45.0 | 202 | 186 |
| SLA/LA (50/50) | 25.3 | −37.0 | −39.7 | 165 | 169 |
| SLA/LA (30/70) | 44.1 | −26.9 | −29.6 | 167 | 172 |
| SLA/LA (10/90) | 25.9 | −18.8 | −14.7 | 196 | 1035 |
| M. smithii TPL | 40.1 | −40.7 | −38.2 | 168 | 169 |
To determine whether the strong CD8+ T cell responses induced with OVA entrapped in SLA/LA archaeosomes could be induced with another antigen, we also entrapped the cancer self-antigen peptide TRP-2 (SVYDFFVWL)) in archaeosomes prepared using different ratios of SLA:LA and tested these in mice. As seen previously with OVA as antigen, delivery of TRP-2 peptide in SLA/LA archaeosomes induced as strong CD8+ T cell responses as were induced by TRP-2 peptide entrapped in M. smithii TPL archaeosomes for both frequency of IFN-γ secreting cells, and functional in vitro cytotoxic response against tumor targets (Fig. 5A, B). In addition, following challenge of immunized mice with a B16 cancer cell line an equivalent reduction in tumor volume compared with naïve unvaccinated mice was obtained with TRP-2 in SLA/LA and M. smithii archaeosomes on day 14 after tumor challenge (Fig. 5C).
Figure 5.
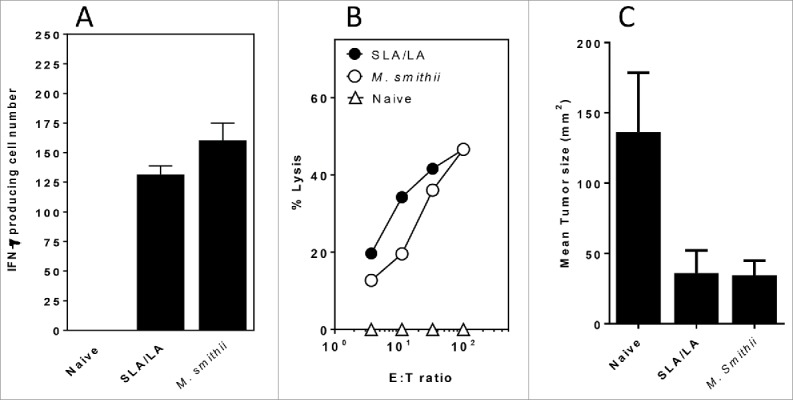
CD8+ (T)cell responses and protection against B16 melanoma challenge induced by TRP-2 entrapped in different archaeosomes. C57/BL6 mice (n = 6/gp) were immunized 15 µg tyrosinase-related protein 2 entrapped in SLA:LA or M. smithii archaeosomes on days 0, 25 and 60. At ∼11 weeks post first immunization, representative mice (n = 2 per group) were killed and splenocytes isolated. Pooled splenocytes were stimulated with IL-2 (0.1 ng/mL) and TRP-2180–188 (10 μg/ml) and the frequency of IFN-gamma secreting cells in triplicate cultures enumerated by ELISPOT. Number of spots/106 spleen cells is indicated (panel A). Splenocytes were also stimulated with TRP2180–189 for 5 d before assessing CTL activity against 51Cr-labeled targets. CTL data represent percentage of specific lysis of triplicate cultures ± SD at various E:T ratios (panel B). Remaining mice (n = 4/gp) were injected with B16 melanoma tumor cells and tumor size monitored over time. The mean tumor size at day 14 following tumor cell injection is shown (panel C).
Discussion
Archaeosomes, primarily composed of total polar lipids of various genera have been used as adjuvants in pre-clinical studies for many years and have been shown to promote strong humoral and cell-mediated responses to entrapped antigen10, 11 to protect immunized mice from pathogens such as Listeria monocytogenes,7, 12 Trypanosoma cruzi,13 and Mycobacterium tuberculosis,14 as well as to protect against solid and metastatic tumors in murine models.8, 15 While typically administered by systemic routes such as subcutaneous or intramuscular injection, the formulation of archaeol TPLs with cations results in the formation of aggregated structures termed archaeal mucosal adjuvant delivery system (AMVAD) which is suitable for mucosal delivery via intranasal routes.16, 17, 18 The effectiveness of the AMVAD system was demonstrated by the induction of strong antigen-specific immune responses and protection against bacterial infection using a murine model of intranasal Francisella tularensis LVS (LVS) challenge to mice immunized intranasally with cell free extract of LVS adjuvanted with the AMVAD system.16 Furthermore, the addition of a detergent, sodium cholate, to TPL archaeosomes has been shown to generate ultradeformable archaeosomes (UDA) for topical delivery of vaccines to the skin.19, 20 Nevertheless, despite these successes, the TPL archaeosome formulations typically used are relatively complex and comprise multiple lipid species. In attempts to overcome this, we have previously tested various semi-synthetic phospho-glyco-archaeol archaeosomes which offer some advantages over traditional TPL formulations while retaining adjuvanticity. However, an optimal formulation of such semi-synthetic archeaosomes still requires the presence of several glycolipids, some of which have a relatively complex synthesis process.9, 21 In this study, we have formulated archaeosomes comprised of sulfated glycol-archaeol lipids and demonstrated that a single or 2 lipid formulation can retain adjuvant properties similar to TPL archaeosomes.
Naturally occurring sulfated glycolipids are found in some archaea such as Halobacterium halobium, Halococcus morrhuae and Haloferax volcanii and are contained in archaeosomes made from the total polar lipids (TPL) of these archaea.11, 22 However, while archaeosomes containing naturally occurring sulfated glycolipids possess some adjuvant activity,11 it is no better and in some cases is weaker than observed with archaeosomes, such as M. smithii or Thermoplasma acidophilum, that do not contain sulfated glycolipids,23 suggesting that the presence of sulfated glycolipids is not a key component of their adjuvant properties. Therefore it was somewhat surprising to find that whereas the naturally occurring purified glycolipid SGL-1 had no adjuvant activity, the synthetic glycolipid SLA could induce a high frequency of antigen-specific IFN-γ secreting CD8+ T cells to the entrapped cargo. Furthermore, whether used alone or in combination with an uncharged glycolipid, SLA was capable of inducing CD8+ CTL activity and protection from tumor challenge in a B16 melanoma model equivalent to that obtained with M. smithii TPL archaeosomes. While there are some structural similarities between SGL-1 and SLA such as the archaeal core lipid, the number of sugar moieties and their attachment via the sn-1 hydroxyl group of the glycerol on the archaeal core lipid as well as and 6′-position of attachment the sulfate moiety, there are some key differences including configuration (α vs β), sugar types (mannose/glucose vs galactose/glucose) and the linkage between sugars (1′-2′ vs 1′-4′). The relative importance of these differences has not yet been fully evaluated and it is likely a combination of structural features that contribute to the stability and adjuvanticity associated with sulfated glycolipids.
Phospholipids are primarily used in conventional eukaryotic liposomes for delivering vaccine and/or drug cargo. Such phospholipids tend often to be inert, and liposomal formulations require addition of other immunomodulatory components such as TLR agonists, to render them adjuvants. A key difference of archaeal phosphoglycolipids is their ability to provide immunostimulation, and we have shown that is related to their glyco- and amino-head groups.9 In contrast, eukaryotic sulfoglycolipids (SGLs) that are produced by tubercle bacilli are suggested to play a role in their virulence and pathogenesis.24 A diacylated sulfoglycolipid purified from Mycobacterium tuberculosis was found to be immunogenic, presented by CD1b molecules to T cells leading to release of interferon-γ .25 Furthermore, the fatty acyl structure as well as sulfation was considered important for the binding to CD1b grove. Future studies will be needed to determine the mechanism of interaction of SLA with immune cells and influence of archaeal backbone and stereochemistry of SLA on adjuvant activity.
Although most of archaeosomes still utilize TPL archaeosomes, there have been several different approaches proposed to modify TPL archaeosomes or their use. These include priming of animals with TLR agonists before immunization with archaeosomes vaccines,26 preparation of stealth archaeosomes through PEGylation of archaeal lipids to increase blood circulation longevity for drug delivery applications,27 and combination with hydrogels to enhance dermal delivery.28 The archaeosome formulation presented herein, composed of the sulfated glycolipid SLA alone or mixed with the uncharged glycolipid LA offers a simplified archaeosome adjuvant formulation.
Materials and methods
Archaeosome production
Halobacterium salinarum was grown and archaeol was purified as described previously.9 Structural identity and purity of archaeol was confirmed by both NMR spectroscopy and negative-ion fast atom bombardment mass spectrometry. Thereafter, lactosylarchaeol (LA; β-D-Galp-(1,4)-β-D-Glcp-(1,1)-archaeol) and sulfated lactosylarchaeol (SLA; 6′-sulfate-β-D-Galp-(1,4)-β-D-Glcp-(1,1)-archaeol) were synthesized as reported previously.21, 29 To enable comparison of a synthetic sulfated glycolipid with a sulfated glycolipid found in nature, a naturally occurring sulfated glycolipid-1 (SGL-1) with structure 6′-HSO3-D-Manp-α-1,2-D-Glcp-α-1,1-archaeol was purified from Haloferax volcanii.22
Archaeosomes were formed by hydrating 20–30 mg dried lipid at 40°C in 2 ml PBS buffer (10 mM sodium phosphate, 160 mM NaCl, pH 7.1) with the protein antigen OVA type VI (Sigma-Aldrich, MO, US) dissolved at 10 mg/ml. Vesicle size was reduced to about 100 - 150 nm diameter by brief sonication in a sonic bath (Fisher Scientific), and the portion of OVA antigen not entrapped was removed by centrifugation from 7 ml PBS followed by 2 washes (200,000 x g max for 30 mins). Vesicle pellets were re-suspended in PBS and filter sterilized through 0.45 μm Millipore filters (EMD Millipore, ON, Canada). Sterile conditions and pyrogen-free water were used throughout. Archaeosomes composed of various molar ratios of SLA:LA were prepared (100/0; 90/10: 70/30; 50/50; 30/70; 10/90) (see Table 1). LA alone, did not hydrate well and hence homogenous vesicles could not be constituted, so a pure LA archaeosomes could not be evaluated. Control archaeosomes were also prepared from the total polar lipids of Methanobrevibacter smithii as described previously.30
Quantification of antigen loading was conducted by separating protein(s) from lipids using SDS polyacrylamide gel electrophoresis as described previously.23 Loading of synthetic archaeosomes with antigens was also determined using SDS Lowry with standard curves prepared for the respective antigen. Loading was based on μg protein/mg salt corrected dry weight of lipid. Average diameters based on Intensity and Zeta potentials were measured using a Malvern Nano Zetasizer with a He/Ne laser (Spectra Research Corp., ON, Canada).
The peptide HLA.A2/H-2Kb TRP-2180–188 (SVYDFFVWL), the CTL epitope from tyrosinase related protein-2, was synthesized. TRP-2180–188 was entrapped using the same methodology for antigen entrapment as described above for ovalbumin. Peptide amount was assayed by RP-HPLC using a Zorbax C-18 reverse-phase column (150× 4.6 mm) with a guard cartridge installed in a DX-300 Dionex dual piston HPLC system (Sunnyvale, CA). The peptide was eluted at a flow rate of 1 ml/min using a gradient aqueous mobile phase from 2% acetonitrile in 0.1% TFA to 70% acetonitrile in 0.085% TFA over 60 min, and revealed by UV absorbance at a 216 nm wavelength. Integration was done by a Dionex 4290 integrator. Quantification was performed using a calibration curve based on known amounts of the peptide.
Animals
Female C56BL/6 mice (6 – 8 weeks) were obtained from the Jackson Laboratory (Bar Harbor, Maine, US). Mice were maintained at the National Research Council Canada (NRC) in accordance with the guidelines of the Canadian Council on Animal Care. All procedures performed on animals in this study were in accordance with regulations and guidelines reviewed and approved by the NRC Human Health Therapeutics Ottawa Animal Care Committee.
Immunization of mice
Female C56BL/6 mice (6 – 8 weeks; n = 6/gp) were injected subcutaneously (SC) on days 0 and 21 with 20 µg ovalbumin (OVA; type VI, Sigma-Aldrich) entrapped in archaeosomes of various compositions. In separate experiments, mice (n = 6/gp) were injected SC on days 0, 25 and 60 with 15 µg TRP-2180–188 entrapped in archaeosomes of various compositions as outlined in Table 1. Antigen with no adjuvant and non-immunized mice were included as negative controls. All animal experiments were repeated on at least one independent occasion to ensure reproducibility of results.
Measurement of immune responses
Antigen-specific IgG levels in the serum of individual mice at various time-points were measured by ELISA as described previously.10 CD8+ T cell responses were quantified by sacrificing representative mice (n = 2 per group) at different time-points and pooling their splenic cells. Representative animals were selected based on Ab titers to ensure animals selected were not hyper- or hypo-responders. These were assayed in triplicate for antigen-specific responses by ELISPOT and CTL methods.31 In brief, splenocytes were stimulated with 0.01 μg/mL of the appropriate antigen (OVA275–264 or TRP2180–189) for 5 d in vitro, and the ensuing effectors were used in a standard 51Cr release CTL assay against non-specific (EL-4) and antigen-specific target cells. CTL data represents percentage of specific lysis of triplicate cultures ± SD at various E:T ratios. Splenocytes were also stimulated with IL-2 (0.1 ng/mL) and 10 µg/mL OVA275–264 or TRP2180–189 and the frequency of IFN-gamma secreting cells enumerated by ELISPOT. To ensure that responses were antigen-specific, control wells contained medium alone. Number of spots/106 spleen cells is indicated.
B-16 melanoma tumor cells were grown in the laboratory as per previously published methods.31 Mice were injected with 106 B16 tumor cells (in PBS plus 0.5% normal mouse serum) in the shaved lower dorsal region, 11.5 weeks post first vaccination. From day 5 onwards, palpable solid tumors were measured using digital calipers. Tumor size, expressed in mm2, was obtained by multiplication of diametrically perpendicular measurements. Mice were killed when the tumor sizes reached a maximum of 300 mm2.
Statistical analysis
Data were analyzed using GraphPad Prism (GraphPad Software, San Diego, CA). Statistical significance of the difference between 2 groups was calculated by Student's 2-tailed t-test and between 3 or more groups by 1-factor analysis of variance (ANOVA) followed by post-hoc analysis using either Dunnett's (comparison with control group) or Tukey's (comparison between groups) multiple comparison tests. Differences were considered to be not significant with p > 0.05.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Acknowledgments
The authors would like to acknowledge John Shelvey and Peter Fleming for producing the archaeal biomass, Dennis Sprott and Dennis Whitfield for designing and providing the archaeal lipids as well as their initial input in the concept of using sulfated glycolipids as adjuvants, as well Chantal Dicaire, Komal Gurnani and Ahmed Zafer for excellent laboratory technical assistance. We would also like to thank Janelle Sauvageau for providing lipid structure figures and helpful discussion.
References
- [1].World Health Organization http://www.who.int/mediacentre/factsheets/fs378/en/.2016 Immunization Coverage: Fact Sheet Number 378. [Google Scholar]
- [2].O'Hagan DT, Ott GS, Nest GV, Rappuoli R, Giudice GD. The history of MF59® adjuvant: a phoenix that arose from the ashes. Expert Rev Vaccines 2013; 12(1):13-30; PMID:23256736; http://dx.doi.org/ 10.1586/erv.12.140 [DOI] [PubMed] [Google Scholar]
- [3].Garcon N, Morel S, Didierlaurent A, Descamps D, Wettendorff M, Van MM. Development of an AS04-adjuvanted HPV vaccine with the adjuvant system approach. BioDrugs 2011; 25(4):217-26; PMID:21815697; http://dx.doi.org/ 10.2165/11591760-000000000-00000 [DOI] [PubMed] [Google Scholar]
- [4].Kates M. Archaebacterial lipids: structure, biosynthesis and function. Biochem Soc Symp 1992; 58:51-72; PMID:1445410 [PubMed] [Google Scholar]
- [5].Sprott GD. Structures of archaebacterial membrane lipids. J Bioenerg Biomembr 1992; 24(6):555-66; PMID:1459987; http://dx.doi.org/ 10.1007/BF00762348 [DOI] [PubMed] [Google Scholar]
- [6].Haq K, Jia Y, Krishnan L. Archaeal lipid vaccine adjuvants for induction of cell-mediated immunity. Expert Rev Vaccines 2016; 15(12):1557-66; PMID:27276183; http://dx.doi.org/ 10.1080/14760584.2016.1195265 [DOI] [PubMed] [Google Scholar]
- [7].Conlan JW, Krishnan L, Willick GE, Patel GB, Sprott GD. Immunization of mice with lipopeptide antigens encapsulated in novel liposomes prepared from the polar lipids of various Archaeobacteria elicits rapid and prolonged specific protective immunity against infection with the facultative intracellular pathogen, Listeria monocytogenes. Vaccine 2001; 19(25–26):3509-17; PMID:11348718; http://dx.doi.org/ 10.1016/S0264-410X(01)00041-X [DOI] [PubMed] [Google Scholar]
- [8].Krishnan L, Deschatelets L, Stark FC, Gurnani K, Sprott GD. Archaeosome adjuvant overcomes tolerance to tumor-associated melanoma antigens inducing protective CD8 T cell responses. Clin Dev Immunol 2010; 2010:578432; PMID:21318177; http://dx.doi.org/ 10.1155/2010/578432 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Sprott GD, Yeung A, Dicaire CJ, Yu SH, Whitfield DM. Synthetic archaeosome vaccines containing triglycosylarchaeols can provide additive and long-lasting immune responses that are enhanced by archaetidylserine. Archaea 2012; 2012:513231; PMID:23055819; http://dx.doi.org/ 10.1155/2012/513231 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].Krishnan L, Dicaire CJ, Patel GB, Sprott G. Archaeosome vaccine adjuvants induce strong humoral, cell-mediated, and memory responses: comparison to conventional liposomes and alum. Infect Immun 2000; 68(1):54-63; PMID:10603368; http://dx.doi.org/ 10.1128/IAI.68.1.54-63.2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Sprott GD, Sad S, Fleming LP, Dicaire CJ, Patel GB, Krishnan L. Archaeosomes varying in lipid composition differ in receptor-mediated endocytosis and differentially adjuvant immune responses to entrapped antigen. Archaea 2003; 1(3):151-64; PMID:15803661; http://dx.doi.org/ 10.1155/2003/569283 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Ansari MA, Zubair S, Tufail S, Ahmad E, Khan MR, Quadri Z, Owais M. Ether lipid vesicle-based antigens impart protection against experimental listeriosis. Int J Nanomedicine 2012; 7:2433-47; PMID:22745536; http://dx.doi.org/ 10.2147/IJN.S25875 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].Higa LH, Corral RS, Morilla MJ, Romero EL, Petray PB. Archaeosomes display immunoadjuvant potential for a vaccine against Chagas disease. Hum Vaccin Immunother 2013; 9(2):409-12; PMID:23291939; http://dx.doi.org/ 10.4161/hv.22780 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Ansari MA, Zubair S, Mahmood A, Gupta P, Khan AA, Gupta UD, Arora A, Owais M. RD antigen based nanovaccine imparts long term protection by inducing memory response against experimental murine tuberculosis. PLoS One 2011; 6(8):e22889; PMID:21853054; http://dx.doi.org/ 10.1371/journal.pone.0022889 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Stark FC, McCluskie MJ, Krishnan L. Homologous Prime-Boost Vaccination with OVA Entrapped in Self-Adjuvanting Archaeosomes Induces High Numbers of OVA-Specific CD8(+) T Cells that Protect Against Subcutaneous B16-OVA Melanoma. Vaccines (Basel) 2016; 4(4):pii: E44; PMID:27869670 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Patel GB, Zhou H, Ponce A, Harris G, Chen W. Intranasal immunization with an archaeal lipid mucosal vaccine adjuvant and delivery formulation protects against a respiratory pathogen challenge. PLoS One 2010; 5(12):e15574; PMID:21206916; http://dx.doi.org/ 10.1371/journal.pone.0015574 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Patel GB, Chen W. Archaeal lipid mucosal vaccine adjuvant and delivery system. Expert Rev Vaccines 2010; 9(4):431-40; PMID:20370552; http://dx.doi.org/ 10.1586/erv.10.34 [DOI] [PubMed] [Google Scholar]
- [18].Patel GB, Zhou H, Ponce A, Chen W. Mucosal and systemic immune responses by intranasal immunization using archaeal lipid-adjuvanted vaccines. Vaccine 2007; 25(51):8622-36; PMID:17959279; http://dx.doi.org/ 10.1016/j.vaccine.2007.09.042 [DOI] [PubMed] [Google Scholar]
- [19].Higa LH, Arnal L, Vermeulen M, Perez AP, Schilrreff P, Mundina-Weilenmann C, Yantorno O, Vela ME, Morilla MJ, Romero EL. Ultradeformable Archaeosomes for Needle Free Nanovaccination with Leishmania braziliensis Antigens. PLoS One 2016; 11(3):e0150185; PMID:26934726; http://dx.doi.org/ 10.1371/journal.pone.0150185 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [20].Higa LH, Schilrreff P, Perez AP, Iriarte MA, Roncaglia DI, Morilla MJ, Romero EL. Ultradeformable archaeosomes as new topical adjuvants. Nanomedicine 2012; 8(8):1319-28; PMID:22366598; http://dx.doi.org/ 10.1016/j.nano.2012.02.008 [DOI] [PubMed] [Google Scholar]
- [21].Whitfield DM, Yu SH, Dicaire CJ, Sprott GD. Development of new glycosylation methodologies for the synthesis of archaeal-derived glycolipid adjuvants. Carbohydr Res 2010; 345(2):214-29; PMID:20004891; http://dx.doi.org/ 10.1016/j.carres.2009.10.011 [DOI] [PubMed] [Google Scholar]
- [22].Sprott GD, Larocque S, Cadotte N, Dicaire CJ, McGee M, Brisson JR. Novel polar lipids of halophilic eubacterium Planococcus H8 and archaeon Haloferax volcanii. Biochim Biophys Acta 2003; 1633(3):179-88; PMID:14499737; http://dx.doi.org/ 10.1016/j.bbalip.2003.08.001 [DOI] [PubMed] [Google Scholar]
- [23].Sprott GD, Patel GB, Krishnan L. Archaeobacterial ether lipid liposomes as vaccine adjuvants. Methods Enzymol 2003; 373:155-72; PMID:14714403 [DOI] [PubMed] [Google Scholar]
- [24].Layre E, Paepe DC, Larrouy-Maumus G, Vaubourgeix J, Mundayoor S, Lindner B, Puzo G, Gilleron M. Deciphering sulfoglycolipids of Mycobacterium tuberculosis. J Lipid Res 2011; 52(6):1098-110; PMID:21482713; http://dx.doi.org/ 10.1194/jlr.M013482 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Gilleron M, Stenger S, Mazorra Z, Wittke F, Mariotti S, Bohmer G, Prandi J, Mori L, Puzo G, De LG. Diacylated sulfoglycolipids are novel mycobacterial antigens stimulating CD1-restricted T cells during infection with Mycobacterium tuberculosis. J Exp Med 2004; 199(5):649-59; PMID:14981115; http://dx.doi.org/ 10.1084/jem.20031097 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Mohammad O, Kaur J, Singh G, Faisal SM, Azhar A, Rauf MA, Gupta UD, Gupta P, Pal R, Zubair S. TLR agonist augments prophylactic potential of acid inducible antigen Rv3203 against mycobacterium tuberculosis H37Rv in experimental animals. PLoS One 2016; 11(3):e0152240; PMID:27023750; http://dx.doi.org/ 10.1371/journal.pone.0152240 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Barbeau J, Cammas-Marion S, Auvray P, Benvegnu T. Preparation and characterization of stealth archaeosomes based on a synthetic PEGylated archaeal tetraether lipid. J Drug Deliv 2011; 2011:396068; PMID:21603209; http://dx.doi.org/ 10.1155/2011/396068 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Gonzalez-Paredes A, Clares-Naveros B, Ruiz-Martinez MA, Durban-Fornieles JJ, Ramos-Cormenzana A, Monteoliva-Sanchez M. Delivery systems for natural antioxidant compounds: Archaeosomes and archaeosomal hydrogels characterization and release study. Int J Pharm 2011; 421(2):321-31; PMID:21986112; http://dx.doi.org/ 10.1016/j.ijpharm.2011.09.042 [DOI] [PubMed] [Google Scholar]
- [29].Whitfield DM, Sprott GD, Krishnan L. Sulfated-Glycolipids As Adjuvants for Vaccines. January 14, 2016 [Google Scholar]
- [30].Krishnan L, Gurnani K, Dicaire CJ, van Faassen H, Zafer A, Kirschning CJ, Sad S, Sprott GD. Rapid clonal expansion and prolonged maintenance of memory CD8+ T cells of the effector (CD44highCD62Llow) and central (CD44highCD62Lhigh) phenotype by an archaeosome adjuvant independent of TLR2. J Immunol 2007; 178(4):2396-406; PMID:17277146; http://dx.doi.org/ 10.4049/jimmunol.178.4.2396 [DOI] [PubMed] [Google Scholar]
- [31].Krishnan L, Sad S, Patel GB, Sprott GD. Archaeosomes induce enhanced cytotoxic T lymphocyte responses to entrapped soluble protein in the absence of interleukin 12 and protect against tumor challenge. Cancer Res 2003; 63(10):2526-34; PMID:12750276 [PubMed] [Google Scholar]