

Abstract
Purpose
Previous studies have shown that melatonin (MEL) signaling is involved in the modulation of photoreceptor viability during aging. Recent work by our laboratory suggested that MEL may protect cones by modulating the Fas/FasL-caspase-3 pathway. In this study, we first investigated the presence of MEL receptors (MT1 and MT2) in 661W cells, then whether MEL can prevent H2O2-induced cell death, and last, through which pathway MEL confers protection.
Methods
The mRNA and proteins of the MEL receptors were detected with quantitative PCR (q-PCR) and immunocytochemistry, respectively. To test the protective effect of MEL, 661W cells were treated with H2O2 for 2 h in the presence or absence of MEL, a MEL agonist, and an antagonist. To study the pathways involved in H2O2–mediated cell death, a Fas/FasL antagonist was used before the exposure to H2O2. Finally, Fas/FasL and caspase-3 mRNA was analyzed with q–PCR and immunocytochemistry in cells treated with H2O2 and/or MEL. Cell viability was analyzed by using Trypan Blue.
Results
Both MEL receptors (MT1 and MT2) were detected at the mRNA and protein levels in 661W cells. MEL partially prevented H2O2-mediated cell death (20–25%). This effect was replicated with IIK7 (a melatonin receptor agonist) when used at a concentration of 1 µM. Preincubation with luzindole (a melatonin receptor antagonist) blocked MEL protection. Kp7–6, an antagonist of Fas/FasL, blocked cell death caused by H2O2 similarly to what was observed for MEL. Fas, FasL, and caspase-3 expression was increased in cells treated with H2O2, and this effect was prevented by MEL. Finally, MEL treatment partially prevented the activation of caspase-3 caused by H2O2.
Conclusions
The results demonstrate that MEL receptors are present and functional in 661W cells. MEL can prevent photoreceptor cell death induced by H2O2 via the inhibition of the proapoptotic pathway Fas/FasL-caspase-3.
Introduction
Retinal melatonin (MEL) is synthetized by the photoreceptors of many vertebrate species [1,2], and the levels of this neurohormone are high during the night and low (or undetectable) during the day. MEL exerts its actions mainly via the activation of two G protein-coupled receptors (GPCRs) known as MT1 and MT2 [3-7], which are both present in the retina [8-11]. In photoreceptor cells, MT1 and MT2 form heterodimers that modulate visual processing and sensitivity [8]. Additional studies have also shown that MEL may protect photoreceptors from oxidative stress [12] and can prevent photoreceptor degeneration [13,14]. A recent study reported that MEL signaling via MEL receptors can partially prevent photoreceptor death (rods and cones) during aging, possibly by decreasing the activation of the apoptotic pathway involving Fas signaling [15].
In recent years, the 661W cells (a cone-like photoreceptor cell line) originally developed by Al-Ubaidi [16] have gained significant recognition as a useful model for studying photoreceptor cell biology [17-21]. Using this cell line, several investigations have partially elucidated the molecular mechanisms underlying photoreceptor cell metabolism [20-22] and photoreceptor cell death following light exposure [23-27] or oxidative stress [28-30].
In this study, to gain a better understanding of the mechanisms by which melatonin can protect photoreceptors, we first investigated whether MEL could prevent cell death induced by oxidative stress in 661W cells, and then we investigated the molecular mechanisms by which MEL may prevent cell death. The data showed that 661W cells may represent a good model for studying melatonin signaling in photoreceptors and the mechanisms underlying the action of melatonin in the modulation of photoreceptor death.
Methods
Cell culture
The 661W photoreceptor cells were provided by Dr. Muayyad Al-Ubaidi (University of Oklahoma Health Sciences Center). The genetic profile of the cells was established by IDEXX Bioresearch (Columbia, MO). Nine short tandem repeat (STR) loci including a gender determining locus were analyzed. To ensure the identity of the cell line as a cone-like cell line, western blot for cone-arrestin, blue opsin and red/green opsin were performed using HEK293 cells as a negative control for the markers. Results obtained are shown in the Appendix 1. Cells were grown in Dulbecco’s Modified Eagle’s medium (DMEM; Gibco, Life Technologies, Carlsbad, CA) supplemented with fetal bovine serum (Gibco) and 1% penicillin/streptomycin, at 37 °C in a 95% O2 and 5% CO2 humidified atmosphere. In most assays, we used cultures seeded in a 75 mm flask at a concentration of 1 × 105 cells in a volume of 10 ml growth media and expanded to approximately 50% to 90%, depending on the experiment. In the immunocytochemistry experiments, the cells were seeded in sterile glass chamber slides and left to expand to 50%. All experimental procedures were performed in accordance with the NIH Guide on Care and Use of Laboratory Animals and were approved by the Morehouse School of Medicine Animal Care and Use Committee.
Drug stock preparation
Hydrogen peroxide 30% was diluted in DMEM to reach working concentrations. MEL (8 mg/ml; Sigma, St. Louis, MO) stock solutions were prepared in absolute ethanol and then diluted to the working concentration in DMEM. Luzindole (LUZ; 10 mM; Tocris, Ellisville, MO), IIK7 (30 mg/ml; Sigma), and Kp7–6 (90 mM; Fisher Scientific, Suwanee, GA) stock solutions were prepared in dimethyl sulfoxide (DMSO) and diluted to working solutions in DMEM.
Immunocytochemistry
The cells were directly washed in PBS (1X; KCl 2.67 mM, KH2PO4 1.47 mM, NaCl 139.9 mM, Na2HPO4, 8,1 mM, pH 7.4) and fixed in cold acetone for 2 min. Once the cells were fixed, they were washed in PBS twice for 10 min and incubated in BLOXALL (Vector Laboratories, Burlingame, CA) to block endogenous peroxidases and phosphatases for 10 min. Then, samples were treated with a biotin, streptavidin blocking kit (Vector Laboratories), washed twice with PBS for 10 min, blocked with PBS containing 0.02% Triton and 5% of bovine serum albumin (1 h at room temperature), and incubated with the primary antibodies MT1 and MT2 (1:200; Alomone Labs, Jerusalem, Israel) for 3 h at room temperature or cleaved caspase-3 (1:400; Cell Signaling, Danvers, MA) overnight at 4 °C. After incubation with the primary antibodies, the samples were washed three times in PBS and subsequently incubated with the secondary biotinylated goat anti-rabbit immunoglobulin G (IgG; 1:200; Vector Laboratories) for 1 h at room temperature. Then after two washes in PBS, the samples were incubated in streptavidin DyLight® 488 (1:100; Vector Laboratories) for 30 min at room temperature. After washing in PBS, the samples were incubated with propidium iodide (1:5,000) for 5 min. Finally, after washing twice in PBS, the chambers were detached from the slide and mounted with Vectashield mounting medium (Vector Laboratories). The slides were examined with a confocal microscope (Zeiss LSM700; Göttingen, Germany). Images were processed with Zeiss Cell software. Micrographs were generated in TIFF format and adjusted linearly for light and contrast before being assembled on plates using Adobe Photoshop CS6 (San Jose, CA). Nuclei size was analyzed by using the Cell Profiler software. To validate the MT1 and MT2 receptor antibodies, a secondary antibody control was used (avoiding the incubation with the primary antibody), as well as an absorption control (by preincubating the antibody with its specific blocking peptide).
Detection of MT1 and MT2 mRNA
RNA extraction (TRI® Reagent method, DNase treatment (Promega, Madison, WI), cDNA synthesis, and conventional PCR (One Step SYBR® PrimeScriptTM RT–PCR kit II, Takara, Mountain View, CA) reactions were performed following the manufacturers’ instructions. A quantity of 200 ng of RNA was used to perform the one-step PCR reactions that were developed in a final volume of 50 μl. The resulting PCR product (the expected sizes are 137 and 222 bp for MT1 and MT2, respectively) was run in an agarose 1.5% Tris- Acetate-EDTA buffer (TAE) gel and visualized by using an EZ-Vision kit (Amresco, VWR, Solon, OH) in an ultraviolet (UV) transilluminator.
Cell viability test
Cell viability was tested by using Trypan Blue. When the cells reached 90% confluence, they were treated with the vehicle, H2O2 (0.1, 1, or 10 mM), MEL (100 or 1,000 nM), IIK7 (10 or 1,000 nM), or a combination of the two for 2 h. After that time point, the cells were detached from the dish with a cell scraper and mixed with Trypan Blue in a 1:2 dilution (final concentration of Trypan Blue of 0.2%) to count viable and non-viable cells in an open field microscope using a Neubauer chamber (Horsham, PA). For the experiments where antagonists were used, LUZ (0.1, 1 and 10 µM) or Kp7–6 (1 mM) was added to the plates 15 min before the addition of MEL (100 nM) and/or H2O2 (1 mM).
Gene expression analysis
Gene expression (Fas, FasL, and caspase-3) was measured with qPCR in cells that were treated with vehicle, MEL (100 or 1,000 nM), and/or H2O2 (1 mM) for 2 h. The 18S gene was used as a reference gene. Specific primers (Invitrogen, Camarillo, CA) and Gene Data Bank reference numbers are shown in Table 1. RNA extraction (TRI® Reagent method, DNase treatment (Promega), cDNA synthesis, and real-time PCR reactions (iTaq™ SYBR® Green Supermix in a CFX96TM Real-Time System; Bio-Rad Laboratories, Hercules, CA) were performed following the manufacturers’ instructions with minor modifications. Total RNA (1 μg) was retrotranscribed and PCR reactions were developed in a final volume of 10 μl (1 μl of cDNA per sample). PCR conditions were 10 min at 95 °C, and 40 cycles consisting of 5 s at 95 °C and 30 s at 60 °C. Calibration curves were made with serial dilutions of cDNA, exhibiting efficiencies around 100%. The specificity of the amplifications was ensured with melting curves. The relative mRNA expression was determined with the ΔΔCt method [31].
Table 1. Accession numbers of the genes and primers sequences employed in quantitative RT–PCR studies.
| Target gene | Accession number | Strand | Primer sequences 5′ → 3′ | Product (bp) |
|---|---|---|---|---|
| Caspase-3 | NM_001284409.1; NM_009810.3 | Forward | GGGCGTGTTTCTGTTTTGTT | 138 |
| Reverse | TTGAGGTAGCTGCACTGTGG | |||
| FasL | NM_010177.4; NM_001205243.1 | Forward | GGTCAGTTTTTCCCTGTCCA | 113 |
| Reverse | CAGTGGGAGTGGTTGTGATG | |||
| Fas | NM_007987.2; NM_001146708.1 | Forward | AGGCGGGTTCGTGAAACTGA | 81 |
| Reverse | GGTTGGCATGGTTGACAGCA | |||
| MT1 | NM_008639.2 | Forward | GAGCTGCTCAATGCCACTCA | 137 |
| Reverse | ACAGGATGACCAGCAGGTTG | |||
| MT2 | NM_145712.2 | Forward | GCGCACCTTGTGCATCTATC | 222 |
| Reverse | GATGACAAGCAGGTTCCCCA | |||
| 18S | K01365.1 | Forward | CTCTGTTCCGCCTAGTCCTG | 123 |
| Reverse | GGCCGTGCGTACTTAGACAT |
The two accession numbers in Caspase-3, FasL and Fas genes correspond with different splicing variants.
Statistical analysis
A one-way ANOVA followed by the post hoc Student–Newman–Keuls (SNK) test was performed for data from the viability, nuclei size, and gene expression experiments. A p value of less than 0.05 was considered statistically significant on all tests. Results are shown as the mean ± standard error of the mean (SEM).
Results
MT1 and MT2 receptors are present in 661W cells
To determine whether 661W cells express melatonin receptors, we first amplified the MT1 and MT2 transcripts from RNA obtained from the 661W cells (Figure 1), and then we performed immunochemistry with the previously validated MT1 and MT2 antibodies. From the data obtained with RT–PCR, we consistently detected MT1 and MT2 immunoreactivity in the 661W cells (Figure 1).
Figure 1.

MEL receptor type 1 (MT1) and melatonin receptor type 2 (MT2) in 661W cells. The top left panel shows MT1 and MT2 RNA expression. Ladder=100 bp. MT1 and MT2 immunoreactivity (green) was detected in 661W cells. Secondary antibody control without a primary antibody (the bottom left panel) and absorption control using a blocking peptide (the bottom center and right panels) were performed. Cell nuclei are shown in red. Scale bar=100 µm.
MEL increases cell viability following H2O2 treatment
Cell viability in 661W cells was significantly reduced in a concentration-dependent manner after 2 h treatment with H2O2 at the concentrations of 1 and 10 mM (Figure 2A). Cotreatment with MEL (100 or 1,000 nM) partially prevented (around 20%) the cell death caused by H2O2 (Figure 2B). When the 661W cells were preincubated with LUZ at 0.1, 1, and 10 μΜ, the protection observed with MEL disappeared gradually (Figure 2C). Cotreatment with IIK7 at a concentration of 10 nM (Figure 2D) did not reduce the rate of cell death, whereas cotreatment of IIK7 at a concentration of 1,000 nM reduced cell death in a comparable amount to that observed with MEL (Figure 2D).
Figure 2.
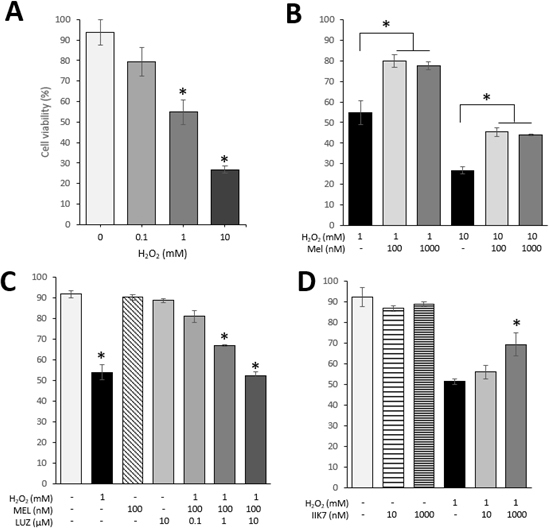
H2O2-induced cell death is partially rescued by MEL and IIK7. A: Cell viability after 2 h of treatment with H2O2. Asterisks indicate a significant effect H2O2-inducing cell death (one-way ANOVA and the Student–Newman–Keuls (SNK) test; p<0.05). B: Cell viability after 2 h of treatment with melatonin (MEL) and/or H2O2. Asterisks indicate a significant effect of MEL preventing H2O2-induced death (one-way ANOVA and the Student–Newman–Keuls (SNK) test; p<0.05). C: Cell viability after pretreatment with luzindole (LUZ), a melatonin antagonist, after 2 h of treatment with MEL and/or H2O2. Asterisks indicate significant differences compared to the control group (treated only with vehicle; one-way ANOVA and the SNK test; p<0.05). D: Cell viability after 2 h of treatment with IIK7 and/or H2O2. Asterisk indicates a significant effect of IIK7, which protects the cells from death (one-way ANOVA and the SNK test; p<0.05). In all the cases, results are shown as mean ± standard error of the mean (SEM; n=6).
MEL prevents the activation of the Fas/FasL-caspase-3 pathway following H2O2 treatment
The activation of the Fas/FasL-caspase-3 pathway was investigated to determine the mechanisms by which MEL protects 661W cells from death induced by H2O2. Blockage of Fas with a Fas/FasL antagonist (Kp7–6) partially prevented 661W cell death after H2O2 treatment (Figure 3) in a similar manner to what was observed with MEL.
Figure 3.
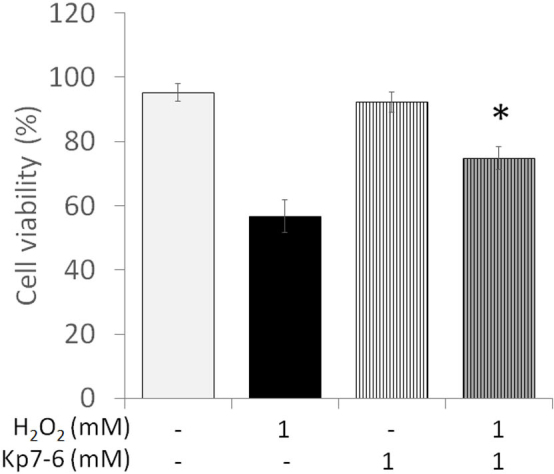
Cell viability is affected by H2O2 via activation of the Fas/FasL pathway. Viability of cells pretreated with Kp7–6, a Fas/FasL antagonist, or vehicle and treated for 2 h with or without H2O2. Results shown as mean ± standard error of the mean (SEM; n=5). Asterisk indicates significant effect of Kp7–6 preventing cell death caused by H2O2 (one-way ANOVA and the Student–Newman–Keuls (SNK) test; p<0.001).
The 2 h treatment with H2O2 induced the gene expression of Fas, FasL, and caspase-3 (Figure 4A). MEL cotreatment (100 or 1,000 nM) partially returned the expression of the three genes studied to basal levels (Figure 4A). The effect of melatonin was blocked in a concentration-dependent manner when the cells were previously treated with LUZ (0.1–10 µM; Figure 4B). Treatment with IIK7 (1,000 nM) was as effective as treatment with MEL in reducing the mRNA levels for the Fas, FasL, and caspase-3 transcripts following H2O2 treatment, whereas treatment with the 10 nM concentration had no effect on the Fas, FasL, and caspase-3 mRNA levels (Figure 4C). Treatment for 2 h with H2O2 (1 mM) increased the signal of cleaved caspase-3, and such an increase was partially abolished by MEL (100 nM; Figure 5). In addition, shrinkage of the nuclei in the cells treated with H2O2 in the absence or presence of MEL was detected (Figure 5), with a size around 70% smaller than the cells treated with vehicle or MEL (data not shown).
Figure 4.

Fas/FasL-caspase-3 pathway is responsible for H2O2-induced cell death, and MEL blocks the activation of this pathway. A: Gene expression levels of Fas, FasL, and caspase-3 after 2 h of treatment with H2O2 and/or melatonin (MEL). Asterisk indicates significant effect of MEL preventing Fas/FasL-caspase-3 pathway activation (one-way ANOVA and the Student–Newman–Keuls (SNK) test; p<0.05). B: Gene expression levels of Fas, FasL, and caspase-3 of cells pretreated with luzindole (LUZ) after 2 h of treatment with MEL and/or H2O2. Asterisks indicate a significant effect of a LUZ-blocking MEL-protective effect (one-way ANOVA and SNK test; p<0.05). C: Gene expression levels of Fas, FasL, and caspase-3 after 2 h of treatment with IIK7 and/or H2O2. Asterisk indicates a significant effect of IIK7 preventing Fas/FasL-caspase-3 pathway activation (one-way ANOVA and the SNK test; p<0.05). In all the cases, results shown as mean ± standard error of the mean (SEM; n=6).
Figure 5.

Caspase-3 cleavage induced by H2O2 is blocked by MEL. Caspase-3 immunoreactivity (green) in cells treated with H2O2 and/or melatonin (MEL) for 2 h. Cell nuclei are shown in red. Scale bar=20 µm.
Discussion
Previous investigations have reported that administration of exogenous melatonin can delay photoreceptor loss in a mouse model of retinitis pigmentosa [13,32] and may also slow down the progression of photoreceptor loss in patients affected by age-related macular degeneration [33]. However, it is unclear whether the protective action of melatonin on the photoreceptor is due to the direct action of melatonin as an antioxidant [12] or whether this protective action is mediated by melatonin’s GPCRs [7].
A recent study conducted by our laboratory reported that MEL signaling may play an important role in the protection of photoreceptor viability during aging. Mice lacking melatonin receptors (MT1 or MT2) show a significant reduction in the number of photoreceptors during aging [14,15]. Interestingly, we observed that the number of cones in mice lacking melatonin receptors is reduced by about 30% at 18 months of age with respect to age-matched controls [15]. The protective action of MEL on the photoreceptor seems to involve the modulation of the Fas/FasL pathway [15].
In this study, we investigated the molecular mechanism by which melatonin signaling may protect cones using 661W cells, a well-established cell line used by many laboratories to study photoreceptor biology [17-21]. The present study data showed that 661W cells express MT1 and MT2 receptors and activation of these receptors by MEL partially protects these cells from oxidative stress (Figure 2). The protective effect of MEL on cell death was concentration dependent and was abolished in the presence of LUZ (MT1 and MT2 antagonist). The fact that IIK7 replicates the effects obtained with MEL further indicated that the protective action of MEL on these cells is mediated by its GPCRs. We have previously shown that IIK7 is specific for MT2 at a concentration of 50 nM, whereas at higher concentration (5 µM) this agonist can also activate MT1 [8]. The present results indicate that IIK7 does not produce a significant effect at a concentration in which only MT2 receptors are activated, whereas at a concentration where both receptors are activated, IIK7 produces a similar effect to MEL (Figure 2 and Figure 3). This result suggests that in 661W cells, MT1 and MT2 may also heterodimerize as seen in mouse photoreceptors [8].
Treatment with H2O2 is a widely used protocol for studying oxidative stress–induced cell death in a variety of cell lines [34-37]. For 661W cells, previous studies used H2O2 as a model to mimic the oxidative stress damage common in photoreceptor cells [22,28-30,38]. The concentration of H2O2 used in these experiments (1 mM) has been previously reported to increase cell death in 661W cells at different incubation durations [22,28,30]. In addition, in many of these studies, H2O2 treatment results in activation of apoptosis [22,28-30,38].
The activation of the Fas/FasL pathway has been previously linked with photoreceptor death [26,39-41]. Along with the progression of different eye diseases, such as retinal detachment or age-related macular degeneration, it has been reported that several proapoptotic factors, including the Fas/FasL pathways (which are mainly in the photoreceptors and the RPE) are activated [39,41]. This activation has been described as a consequence of an increase in the levels of oxidative stress in the retina [26,41]. The presence of Fas on the cell surface of 661W cells has been demonstrated, along with the involvement of Fas in light-induced cell death [26]. Interestingly, previous studies have shown that in response to FasL, 661W cells exhibited specific changes in the morphology of the cells considered markers of apoptosis, including shrinkage and loss of cellular adhesion and cytoplasmic condensation. We also observed this phenotype in the present study experiments after 2 h of H2O2 treatment, as can be partially detected in Figure 5, where the size and compactness of the cells are modified in the H2O2-treated group, compared with the control cells and the cells treated with MEL only or with MEL and H2O2. Furthermore, the same study showed that the activation of the Fas/FasL pathway in 661W cells leads to the subsequent activation of caspase-3 [26], which is a result that also agrees with our observations. Our results indicate that MEL (via its associated receptors) can partially protect photoreceptor cells against oxidative stress damage by inhibiting the activation of Fas/FasL and thus prevents caspase-3-mediated cell death.
In conclusion, the present study shows that activation of melatonin signaling via its associated receptors can protect photoreceptor cells against oxidative stress via modulation of the Fas/FasL pathway. These data expand our previous study by demonstrating that melatonin acting via its associated receptors may represent a useful tool for protecting photoreceptor (cones) cells against oxidative stress and thus increase the viability of these cells during aging. Finally, the present study also indicates that 661W cells may represent a very important new tool for studying melatonin signaling in photoreceptors and possibly the mechanisms by which MT1 and MT2 form heterodimers.
Acknowledgments
This work was supported by grants from the National Institutes of Health Grants EY022216, R01EY026291 to G.T. and by 5U54NS083932, S21MD000101, G12-RR03034, U54RR026137 to Morehouse School of Medicine.
Appendix 1. Expression of specific cone markers in 661W cells compared with HEK293 cells (negative control).
To access the data, click or select the words “Appendix 1” Protein extracts were incubated overnight at 4°C with cone arrestin (1:5000, Millipore, 42 KDa), blue opsin (39 KDa), red/green opsin (40.5 KDa) and α E-catenin (100 KDa; 1:5000, Cell signaling) primary antibodies. Then, they were incubated with antirabbit HRP (1:10000, Cell signaling) and developed.
References
- 1.Klein DC, Coon SL, Roseboom PH, Weller JL, Bernard M, Gastel JA, Zatz M, Iuvone PM, Rodriguez IR, Bégay V, Falcón J, Cahill GM, Cassone VM, Baler R. The melatonin rhythm-generating enzyme: molecular regulation of serotonin N-acetyltransferase in the pineal gland. Recent Prog Horm Res. 1997;52:307–57. [PubMed] [Google Scholar]
- 2.Iuvone PM, Tosini G, Pozdeyev N, Haque R, Klein DC, Chaurasia SS. Circadian clocks, clock networks, arylalkylamine N-acetyltransferase, and melatonin in the retina. Prog Retin Eye Res. 2005;24:433–56. doi: 10.1016/j.preteyeres.2005.01.003. [DOI] [PubMed] [Google Scholar]
- 3.Dubocovich ML, Delagrange P, Krause DN, Sugden D, Cardinali DP, Olcese J. International Union of Basic and Clinical Pharmacology. LXXV. Nomenclature, classification, and pharmacology of G protein-coupled melatonin receptors. Pharmacol Rev. 2010;62:343–80. doi: 10.1124/pr.110.002832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Reppert SM. Melatonin receptors: molecular biology of a new family of G protein-coupled receptors. J Biol Rhythms. 1997;12:528–31. doi: 10.1177/074873049701200606. [DOI] [PubMed] [Google Scholar]
- 5.Tosini G, Owino S, Guillame J-L, Jockers R. Melatonin receptors: latest insights from mouse models. BioEssays News Rev Mol Cell Dev Biol. 2014;36:778–87. doi: 10.1002/bies.201400017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Cecon E, Oishi A, Jockers R. Melatonin receptors: molecular pharmacology and signaling in the context of system bias. Br J Pharmacol. 2017;(July) doi: 10.1111/bph.13950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Jockers R, Delagrange P, Dubocovich ML, Markus RP, Renault N, Tosini G, Cecon E, Zlotos DP. Update on melatonin receptors: IUPHAR Review 20. Br J Pharmacol. 2016;173:2702–25. doi: 10.1111/bph.13536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Baba K, Benleulmi-Chaachoua A, Journé AS, Kamal M, Guillaume JL, Dussaud S, Gbahou F, Yettou K, Liu C, Contreras-Alcantara S, Jockers R, Tosini G. Heteromeric MT1/MT2 melatonin receptors modulate photoreceptor function. Sci Signal. 2013;6:ra89. doi: 10.1126/scisignal.2004302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Scher J, Wankiewicz E, Brown GM, Fujieda H. MT(1) melatonin receptor in the human retina: expression and localization. Invest Ophthalmol Vis Sci. 2002;43:889–97. [PubMed] [Google Scholar]
- 10.Savaskan E, Jockers R, Ayoub M, Angeloni D, Fraschini F, Flammer J, Eckert A, Müller-Spahn F, Meyer P. The MT2 melatonin receptor subtype is present in human retina and decreases in Alzheimer’s disease. Curr Alzheimer Res. 2007;4:47–51. doi: 10.2174/156720507779939823. [DOI] [PubMed] [Google Scholar]
- 11.Fujieda H, Hamadanizadeh SA, Wankiewicz E, Pang SF, Brown GM. Expression of mt1 melatonin receptor in rat retina: evidence for multiple cell targets for melatonin. Neuroscience. 1999;93:793–9. doi: 10.1016/s0306-4522(99)00111-6. [DOI] [PubMed] [Google Scholar]
- 12.Marchiafava PL, Longoni B. Melatonin as an antioxidant in retinal photoreceptors. J Pineal Res. 1999;26:184–9. doi: 10.1111/j.1600-079x.1999.tb00582.x. [DOI] [PubMed] [Google Scholar]
- 13.Liang FQ, Aleman TS. ZaixinYang null, Cideciyan AV, Jacobson SG, Bennett J. Melatonin delays photoreceptor degeneration in the rds/rds mouse. Neuroreport. 2001;12:1011–4. doi: 10.1097/00001756-200104170-00029. [DOI] [PubMed] [Google Scholar]
- 14.Baba K, Pozdeyev N, Mazzoni F, Contreras-Alcantara S, Liu C, Kasamatsu M, Martinez-Merlos T, Strettoi E, Iuvone PM, Tosini G. Melatonin modulates visual function and cell viability in the mouse retina via the MT1 melatonin receptor. Proc Natl Acad Sci USA. 2009;106:15043–8. doi: 10.1073/pnas.0904400106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gianesini C, Hiragaki S, Laurent V, Hicks D, Tosini G. Cone Viability Is Affected by Disruption of Melatonin Receptors Signaling. Invest Ophthalmol Vis Sci. 2016;57:94–104. doi: 10.1167/iovs.15-18235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.al-Ubaidi MR, Font RL, Quiambao AB, Keener MJ, Liou GI, Overbeek PA, Baehr W. Bilateral retinal and brain tumors in transgenic mice expressing simian virus 40 large T antigen under control of the human interphotoreceptor retinoid-binding protein promoter. J Cell Biol. 1992;119:1681–7. doi: 10.1083/jcb.119.6.1681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Al-Ubaidi MR, Matsumoto H, Kurono S, Singh A. Proteomics profiling of the cone photoreceptor cell line, 661W. Adv Exp Med Biol. 2008;613:301–11. doi: 10.1007/978-0-387-74904-4_35. [DOI] [PubMed] [Google Scholar]
- 18.Duricka DL, Brown RL, Varnum MD. Defective trafficking of cone photoreceptor CNG channels induces the unfolded protein response and ER-stress-associated cell death. Biochem J. 2012;441:685–96. doi: 10.1042/BJ20111004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Tan E, Ding X-Q, Saadi A, Agarwal N, Naash MI, Al-Ubaidi MR. Expression of Cone-Photoreceptor–Specific Antigens in a Cell Line Derived from Retinal Tumors in Transgenic Mice. Invest Ophthalmol Vis Sci. 2004;45:764–8. doi: 10.1167/iovs.03-1114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Farrell SMJ, Groeger G, Bhatt L, Finnegan S, O’Brien CJ, Cotter TG. bFGF-mediated redox activation of the PI3K/Akt pathway in retinal photoreceptor cells. Eur J Neurosci. 2011;33:632–41. doi: 10.1111/j.1460-9568.2010.07559.x. [DOI] [PubMed] [Google Scholar]
- 21.Ozaki T, Nakazawa M, Yamashita T, Tomita H, Ebina Y, Ishiguro S. Decrease of ATP by mitochondrial m-calpain inhibitory peptide in the rat retinas. Cell Struct Funct. 2013;38:207–23. doi: 10.1247/csf.13008. [DOI] [PubMed] [Google Scholar]
- 22.Layton CJ. Diabetic levels of glucose increase cellular reducing equivalents but reduce survival in three models of 661W photoreceptor-like cell injury. BMC Ophthalmol. 2015;15:174. doi: 10.1186/s12886-015-0164-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Natoli R, Rutar M, Lu YZ, Chu-Tan JA, Chen Y, Saxena K, Madigan M, Valter K, Provis JM. The Role of Pyruvate in Protecting 661W Photoreceptor-Like Cells Against Light-Induced Cell Death. Curr Eye Res. 2016;41:1473–81. doi: 10.3109/02713683.2016.1139725. [DOI] [PubMed] [Google Scholar]
- 24.Krishnamoorthy RR, Crawford MJ, Chaturvedi MM, Jain SK, Aggarwal BB, Al-Ubaidi MR, Agarwal N. Photo-oxidative stress down-modulates the activity of nuclear factor-kappaB via involvement of caspase-1, leading to apoptosis of photoreceptor cells. J Biol Chem. 1999;274:3734–43. doi: 10.1074/jbc.274.6.3734. [DOI] [PubMed] [Google Scholar]
- 25.Chen W-J, Wu C, Xu Z, Kuse Y, Hara H, Duh EJ. Nrf2 protects photoreceptor cells from photo-oxidative stress induced by blue light. Exp Eye Res. 2017;154:151–8. doi: 10.1016/j.exer.2016.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Chang Q, Peter ME, Grassi MA. Fas ligand-Fas signaling participates in light-induced apoptotic death in photoreceptor cells. Invest Ophthalmol Vis Sci. 2012;53:3703–16. doi: 10.1167/iovs.11-8928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kuse Y, Tsuruma K, Kanno Y, Shimazawa M, Hara H. CCR3 Is Associated with the Death of a Photoreceptor Cell-line Induced by Light Exposure. Front Pharmacol. 2017;8:207. doi: 10.3389/fphar.2017.00207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Fabiani C, Zulueta A, Bonezzi F, Casas J, Ghidoni R, Signorelli P, Caretti A. 2-Acetyl-5-tetrahydroxybutyl imidazole (THI) protects 661W cells against oxidative stress. Naunyn Schmiedebergs Arch Pharmacol. 2017;390:741–51. doi: 10.1007/s00210-017-1374-3. [DOI] [PubMed] [Google Scholar]
- 29.Mackey AM, Sanvicens N, Groeger G, Doonan F, Wallace D, Cotter TG. Redox survival signalling in retina-derived 661W cells. Cell Death Differ. 2008;15:1291–303. doi: 10.1038/cdd.2008.43. [DOI] [PubMed] [Google Scholar]
- 30.Chen H, Tran J-TA, Anderson RE, Mandal MNA. Caffeic acid phenethyl ester protects 661W cells from H2O2-mediated cell death and enhances electroretinography response in dim-reared albino rats. Mol Vis. 2012;18:1325–38. [PMC free article] [PubMed] [Google Scholar]
- 31.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001;25:402–8. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 32.Xu X-J, Wang S-M, Jin Y, Hu Y-T, Feng K, Ma Z-Z. Melatonin delays photoreceptor degeneration in a mouse model of autosomal recessive retinitis pigmentosa. J Pineal Res. 2017;(June) doi: 10.1111/jpi.12428. [DOI] [PubMed] [Google Scholar]
- 33.Yi C, Pan X, Yan H, Guo M, Pierpaoli W. Effects of melatonin in age-related macular degeneration. Ann N Y Acad Sci. 2005;1057:384–92. doi: 10.1196/annals.1356.029. [DOI] [PubMed] [Google Scholar]
- 34.Xiang J, Wan C, Guo R, Guo D. Is Hydrogen Peroxide a Suitable Apoptosis Inducer for All Cell Types? BioMed Res Int. doi: 10.1155/2016/7343965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kim M-H, Chung J, Yang J, Chung S-M, Kwag N-H, Yoo J-S. Hydrogen peroxide-induced cell death in a human retinal pigment epithelial cell line, ARPE-19. Korean J Ophthalmol KJO. 2003;17:19–28. doi: 10.3341/kjo.2003.17.1.19. [DOI] [PubMed] [Google Scholar]
- 36.Park WH. The effect of MAPK inhibitors and ROS modulators on cell growth and death of H2O2-treated HeLa cells. Mol Med Rep. 2013;8:557–64. doi: 10.3892/mmr.2013.1551. [DOI] [PubMed] [Google Scholar]
- 37.Hirsch I, Prell E, Weiwad M. Assessment of cell death studies by monitoring hydrogen peroxide in cell culture. Anal Biochem. 2014;456:22–4. doi: 10.1016/j.ab.2014.04.009. [DOI] [PubMed] [Google Scholar]
- 38.Nishida T, Kuse Y, Mochizuki K, Shimazawa M, Yamamoto T, Hara H. Protective effects of fluoroquinolones on UV-induced damage of cultured ocular cell lines. Eur J Pharmacol. 2017;806:59–66. doi: 10.1016/j.ejphar.2017.04.004. [DOI] [PubMed] [Google Scholar]
- 39.Zacks DN, Zheng Q-D, Han Y, Bakhru R, Miller JW. FAS-mediated apoptosis and its relation to intrinsic pathway activation in an experimental model of retinal detachment. Invest Ophthalmol Vis Sci. 2004;45:4563–9. doi: 10.1167/iovs.04-0598. [DOI] [PubMed] [Google Scholar]
- 40.Dunaief JL, Dentchev T, Ying GS, Milam AH. The role of apoptosis in age-related macular degeneration. Arch Ophthalmol. 2002;120:1435–42. doi: 10.1001/archopht.120.11.1435. [DOI] [PubMed] [Google Scholar]
- 41.Xiao J, Yao J, Jia L, Lin C, Zacks DN. Protective Effect of Met12, a Small Peptide Inhibitor of Fas, on the Retinal Pigment Epithelium and Photoreceptor After Sodium Iodate Injury. Invest Ophthalmol Vis Sci. 2017;58:1801–10. doi: 10.1167/iovs.16-21392. [DOI] [PMC free article] [PubMed] [Google Scholar]