

Key Points
Many CD4 T cells specific for FVIII escape thymic selection in healthy donors, revealing a low central tolerance to FVIII.
Some FVIII-specific CD4 T cells are differentiated into memory cells but do not expand.
Abstract
We investigated the frequency and subset origin of circulating factor VIII (FVIII)–specific CD4 T cells in healthy donors. Total CD4 T cells and purified CD4 T-cell subsets were stimulated with FVIII-loaded autologous dendritic cells and challenged for specificity in interferon-γ enzyme-linked immunospots. The number of specific T-cell lines allowed estimation of the frequency of T cells circulating in the blood of the donors. All the 16 healthy donors generated strong in vitro T-cell responses, leading to the generation of 154 FVIII-specific T-cell lines. The mean frequency of FVIII-specific CD4 T cells in healthy donors was similar to that of T cells specific for foreign antigens and greater than that of T cells specific for known immunogenic therapeutic proteins. Normal levels of endogenous FVIII in healthy donors therefore do not prevent a significant escape of FVIII-specific CD4 T cells from negative thymic selection. FVIII-specific T cells mainly originated from both the naïve and central memory cell subsets, but their frequencies remained low as compared with those of cells specific for foreign antigens in immunized donors. The observation of a spontaneous generation of FVIII-specific memory T cells without a global expansion suggests peculiar peripheral tolerance mechanisms to FVIII in healthy donors.
Visual Abstract
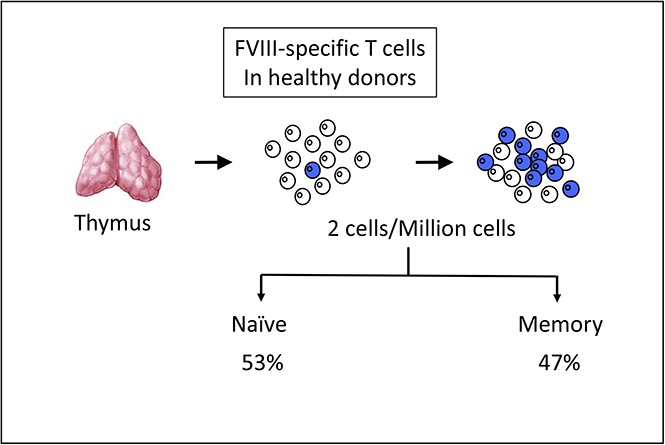
Introduction
A large array of self-antigens are the targets of natural antibodies found in healthy donors.1 In particular, up to 20% of healthy individuals possess immunoglobulin G reactive to procoagulant factor VIII (FVIII), which may potentially inhibit FVIII activity.2,3 Incidentally, loss of tolerance to endogenous FVIII may occur in individuals without previous abnormality in hemostasis, leading to acquired hemophilia, a severe autoimmune bleeding disorder.4 Taken together, these observations call into question the concept that tolerance to FVIII is ensured by the expression of the endogenous protein. This is in contrast to observations made in patients with congenital hemophilia A (HA), an X-linked hemorrhagic disease characterized by FVIII deficiency. The incidence of inhibitory alloantibodies to FVIII after replacement therapy using exogenous therapeutic FVIII is indeed higher in patients with no endogenous FVIII production than in patients with mild/moderate forms of the disease who produce a mutated FVIII.5 In agreement with their helper functions, FVIII-specific CD4 T cells participate in the loss of tolerance to FVIII in patients with HA.6,7 CD4 T cells are selected positively and negatively by self-peptides presented by HLA class 2 molecules carried by specialized thymic cells. After encountering antigen in the periphery, naïve T cells differentiate into effector cells that in turn become central memory cells and effector memory cells,8 the ultimate stage being effector memory reverting cells.9 FVIII-specific CD4 T cells have been mainly characterized in patients with moderate or severe HA,10,11 but pioneering studies have also documented the existence of FVIII-reactive CD4+ T cells in healthy individuals.12-14 However, no clear difference was found in terms of intensity of the anti-FVIII response or epitope specificity between T cells from healthy individuals and patients with HA who had developed anti-FVIII alloantibodies.12,14 To shed some light on the mechanisms underlying T-cell tolerance to FVIII under physiological conditions, we quantified and characterized naturally occurring FVIII-specific CD4+ T cells in healthy donors.
Material and methods
Generation of FVIII- or KLH-specific T-cell lines
Peripheral blood mononuclear cells (PBMCs) were purified from the blood of anonymous healthy donors who gave informed consent (Etablissement Français du Sang, Rungis, France). Dendritic cells (DCs) were generated in 5-day cultures with 1000 U/mL of interleukin-4 and 1000 U/mL of granulocyte macrophage colony-stimulating factor (RD systems) of plastic-adherent cells isolated from 500 million to 1 billion PBMCs. CD4 T cells were isolated using magnetic microbeads (Miltenyi Biotech, Bergisch Gladbach, Germany) from 100 million nonadherent PBMCs. CD4 T-cell subsets were purified by flow cytometry (FACSARIA III; Becton Dickinson, San Jose, CA) using the following labeled antibodies: fluorescein isothiocyanate-labeled anti-CD3, phycoerythrin-labeled anti-CD45RA, peridinin chlorophyll protein complex cyanin 5.5–labeled anti-CD4, allophycocyanin-labeled anti-CD45RO, phycoerythrin cyanin 7–labeled anti-CD127, Brilliant Violet 421-labeled anti-CD197, and Brilliant Violet 711-labeled anti-CD62L (Becton Dickinson). CD4 T-cell lines were generated in vitro in Iscove modified Dulbecco medium supplemented with 10% human serum, 0.24 mM of glutamine, 0.55 mM of asparagine, 1.5 mM of arginine (all amino acids from Sigma-Aldrich), and 50 U/mL of penicillin/50 μg/mL of streptomycin (Invitrogen) by 3 once-per-week rounds of stimulation of CD4+ T cells (200 000 per well) with autologous DCs (20 000 per well) previously incubated overnight at 37°C with recombinant 0.2 µM of FVIII (Kogenate; Bayer, San Francisco, CA) or 0.25 µM of keyhole limpet hemocyanin (KLH; Sigma, St Louis, MO) as previously described.15
ELISPOT assay
The antigen specificity of the CD4 T-cell lines was analyzed by interferon-γ (IFN-γ) enzyme-linked immunospot (ELISPOT). T cells (30 000 cells per well) were incubated with autologous DCs alone (control) or with DCs previously loaded with FVIII (0.2 µM) or KLH (2 µM) in MultiScreen 96-well plates (Merck Millipore) previously coated with 2.5 µg/mL of anti-human IFN-γ monoclonal antibody (mAb; 1-D1K; Mabtech). After overnight incubation, spots were revealed using 0.25 µg/mL of biotinylated anti-human IFN-γ mAb (7-B6-1; Mabtech) in phosphate-buffered saline/bovine serum albumin 1%, extravidin-phosphatase (dilution 1:3000 in phosphate-buffered saline/Tween 20 0.05%/bovine serum albumin 1%; Sigma-Aldrich), and nitro blue tetrazolium/5-bromo-4-chloro-3-indolyl phosphate-toluidine salt (Sigma-Aldrich). CD4 T-cell lines were considered specific when a spot count was twofold higher in the presence of the protein than in its absence, with a minimal difference of 25 spots.
Results and discussion
To detect KLH- and FVIII-specific CD4 T lymphocytes, purified CD4 T lymphocytes collected from healthy individuals were distributed into multiple wells and stimulated weekly by autologous DCs previously loaded with FVIII or KLH. After 3 rounds of stimulation, each independent T-cell line (CD4 T cells present in a single well) was evaluated for its specificity by IFN-γ ELISPOT under culture conditions favoring the differentiation of T cells into TH1 cells. KLH was introduced as a control antigen to assess the ability of the donors to mount a CD4 T-cell response. As shown Figure 1A, 9 T-cell lines specific for FVIII were obtained from donor #329, corresponding to almost half of the seeded wells at the initiation of the culture. As shown for donor #330, FVIII-specific T-cell lines did not react with KLH, and KLH-specific T-cell lines were not activated by FVIII (Figure 1B). A total of 154 FVIII-specific T-cell lines were isolated from the 16 donors (supplemental Table 1), all of them responding to FVIII. As demonstrated previously,15 the distribution of the antigen-specific CD4 T cells at the initiation of our assay follows a Poisson distribution. This allowed us to calculate a frequency of FVIII-specific CD4 T cells between 0.3 and 4.2 per million of total CD4 T cells, with a mean of 1.7 cells per million. The frequency of FVIII-specific CD4 T cells was ∼10-fold greater than that calculated in the case of CD4+ T cells specific for human recombinant IFN-β (S.M., Aurélien Azam, Marie de Bourayne, E.M., and B.M., manuscript in preparation), investigated using the same set of donors and methods previously used to calculate frequency in the cases of CD4+ T cells specific for therapeutic antibodies,15 human erythropoietin, antitrypsin, antithrombin, and insulin.16 Strikingly, the frequency of FVIII-specific CD4 T cells was in the range of that calculated in the case of foreign antigens such as mouse mAbs,15 ovalbumin,15 and HIV peptide vaccine.17 It was however below the frequency found for strong immunogens, such as KLH,15,18 and it was too low to be detected without any in vitro culture phase. The data suggest that the basal level of expression of endogenous FVIII (ie, 0.1 µg/mL or 0.3 nM in normal plasma19) does not preclude a large escape of FVIII-specific CD4 T cells from negative thymic selection. Whether such a large pool of circulating FVIII-specific CD4 T cells provides help to natural FVIII-specific B cells and accounts for the presence of natural anti-FVIII antibodies in a substantial percentage of healthy individuals3,20 and whether the uncontrolled recruitment of this pool leads to the development of acquired hemophilia remain to be established. Furthermore, it is tempting to speculate that, because of the impaired production of FVIII that characterizes HA, the pool of FVIII-specific CD4 T cells is enlarged in patients with HA, although formal evaluation of the size of the preexisting T-cell repertoire in previously untreated patients is ethically challenging. The hypothesis is substantiated by recent ancillary results from the SIPPET study that document the existence of FVIII-binding antibodies in the plasma of 8% of previously untreated patients.21
Figure 1.

Generation of FVIII-specific CD4 T cells from healthy individuals. CD4 T-cell lines were generated as described in Material and Methods. (A) FVIII-specific T-cell lines generated from donor #329. (B) Cross-reactivity analysis between KLH- and FVIII-specific T-cell lines. (C) Frequencies of CD4 T cells specific for FVIII and KLH estimated for 16 healthy blood donors. The frequency of CD4 T-cell precursors was calculated using Poisson distribution according to the following formula: Frequency = −Ln([number of negative wells/total number of wells tested])/(number of CD4 T cells per well). Statistical significance was assessed using the Wilcoxon match-pairs signed rank test. ****P < .0001. Ctrl, control.
To assess the phenotypes of the anti-FVIII CD4 T cells in healthy individuals, we separated the CD4 T cells from 5 different donors into 4 main subsets, namely naïve T cells, central memory cells, effector memory cells and effector cells CD45RO+, and effector memory cells and effector cells CD45RA+ (supplemental Figure 1; supplemental Table 2) and generated FVIII-specific T-cell lines. As shown in the case of donors #393 and #423, all T-cell subsets gave rise to FVIII-specific T-cell lines, albeit with different efficiencies (Figure 2). A large number of T-cell lines originated from naïve cells and to a lesser extent from central memory cells (Figure 2A). The frequency of FVIII-specific T cells was calculated in each subset with respect to the whole population (supplemental Table 2). Naïve and central memory cells (Figure 2B) represented 53% and 29% of the FVIII-specific T cells, respectively, whereas the 2 other subsets were less represented (12% for effector memory cells and effector cells CD45RO+ and 6% for effector memory cells and effector cells CD45RA+). Although striking, the existence of self-reactive memory cells in normal individuals has already been observed by others22 and by us for another self-antigen.23 Importantly, FVIII-specific memory CD4 T cells were found at frequencies equivalent to that of naïve CD4 T cells, suggesting that they did not expand, as memory cells specific for foreign antigens generally do.18,24 Indeed, memory responses to foreign antigens lead to frequencies of memory T cells that are ≥100-fold greater than the frequencies of naïve T cells specific for the same antigens.18,24 It is however probable that FVIII-specific memory CD4 T cells expand in patients with HA who have developed inhibitors. The apparent lack of expansion of FVIII-specific memory CD4 T cells seen in our study with healthy donors may have resulted from a suboptimal priming of the T cells by the low concentration of endogenous FVIII in the absence of inflammatory cytokines or from active regulation by regulatory T cells.25,26 Importantly, in our analysis of the CD4 T-cell subsets, antigen-specific regulatory T cells were removed before initiation of the in vitro culture and accounted for <0.1% of the growing CD4 T cells (data not shown).
Figure 2.

Characterization of FVIII-specific CD4 T-cell subsets. CD4 T cells were separated by flow cytometry into 4 subsets: naïve T cells (CD45RA+ CD45RO− CD127+ CD62L+ CD197+), central memory cells (TCMs; CD45RA− CD45RO+ CD127+ CD62L+ CD197+), effector memory cells and effector cells CD45RO+ (TEM&TeffRO; CD45RA− CD45RO+ CD127+ CD62L−), and effector memory cells and effector cells CD45RA+ (TEMRA&TeffRA; CD45RA+ CD45RO− CD127+ CD62L−; supplemental Figure 1). CD4 T-cell lines were generated from the cells of these subsets by 3 once-per-week rounds of in vitro stimulation with autologous DCs loaded with 0.2 µM of FVIII or 0.25 µM of KLH (for naïve cells only). Specificity of CD4 T-cell lines raised against FVIII or KLH was analyzed by IFN-γ ELISPOT. (A) FVIII-specific T-cell lines obtained from the different CD4 T-cell subsets isolated from 2 representative donors: #393 and #423. (B) Frequencies of FVIII-specific CD4 T cells isolated from the different CD4 T-cell subsets of all donors. Statistical significance was assessed using the Wilcoxon match-pairs signed rank test. *P < .05. Ctrl, control.
The high frequency of preexisting FVIII-specific CD4 T cells reported in the present study suggests a deficiency in the central tolerance to FVIII. It is reminiscent of the immune ignorance27 previously proposed in the case of endogenous erythropoietin.16 We also observed that a significant portion of the preexisting FVIII-specific CD4 T cells had a memory phenotype without overt sign of expansion. Maintenance of tolerance to FVIII in healthy donors might rely on such an apparent lack of expansion.
Supplementary Material
The full-text version of this article contains a data supplement.
Acknowledgments
The authors thank Jeanette Lo (Bayer, San Francisco, CA) for the kind gift of Kogenate.
The research leading to these results was supported by the Innovative Medicines Initiative Joint Undertaking ABIRISK (Anti-Biopharmaceutical Immunization Risk) project under grant agreement #115303, the resources of which comprise financial contribution from the European Union’s Seventh Framework Program (FP7/2007-2013) and in-kind contributions from European Federation of Pharmaceutical Industries and Associations companies. This work was also supported by the Labex in Research on Medication and Therapeutic Innovation (LERMIT) and the Commissariat à l’Énergie Atomique (B.M.).
Authorship
Contribution: S.M. designed research, performed research, analyzed data, and wrote the paper; C.M. and E.M. performed research and analyzed data; and S.L.-D. and B.M. designed research, analyzed data, and wrote the paper.
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Correspondence: Bernard Maillère, CEA-Saclay, Institute Frederic Joliot, SIMOPRO, 91191 Gif sur Yvette, France; e-mail: bernard.maillere@cea.fr.
References
- 1.Marchalonis JJ, Kaveri S, Lacroix-Desmazes S, Kazatchkine MD. Natural recognition repertoire and the evolutionary emergence of the combinatorial immune system. FASEB J. 2002;16(8):842-848. [DOI] [PubMed] [Google Scholar]
- 2.Lacroix-Desmazes S, Misra N, Bayry J, Mohanty D, Kaveri SV, Kazatchkine MD. Autoantibodies to factor VIII. Autoimmun Rev. 2002;1(1-2):105-110. [DOI] [PubMed] [Google Scholar]
- 3.Whelan SFJ, Hofbauer CJ, Horling FM, et al. . Distinct characteristics of antibody responses against factor VIII in healthy individuals and in different cohorts of hemophilia A patients. Blood. 2013;121(6):1039-1048. [DOI] [PubMed] [Google Scholar]
- 4.Franchini M, Mannucci PM. Acquired haemophilia A: a 2013 update. Thromb Haemost. 2013;110(6):1114-1120. [DOI] [PubMed] [Google Scholar]
- 5.Pandey GS, Yanover C, Miller-Jenkins LM, et al. ; PATH (Personalized Alternative Therapies for Hemophilia) Study Investigators. Endogenous factor VIII synthesis from the intron 22-inverted F8 locus may modulate the immunogenicity of replacement therapy for hemophilia A. Nat Med. 2013;19(10):1318-1324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Jacquemin MG, Desqueper BG, Benhida A, et al. . Mechanism and kinetics of factor VIII inactivation: study with an IgG4 monoclonal antibody derived from a hemophilia A patient with inhibitor. Blood. 1998;92(2):496-506. [PubMed] [Google Scholar]
- 7.Bray GL, Kroner BL, Arkin S, et al. . Loss of high-responder inhibitors in patients with severe hemophilia A and human immunodeficiency virus type 1 infection: a report from the Multi-Center Hemophilia Cohort Study. Am J Hematol. 1993;42(4):375-379. [DOI] [PubMed] [Google Scholar]
- 8.Sallusto F, Geginat J, Lanzavecchia A. Central memory and effector memory T cell subsets: function, generation, and maintenance. Annu Rev Immunol. 2004;22:745-763. [DOI] [PubMed] [Google Scholar]
- 9.Di Mitri D, Azevedo RI, Henson SM, et al. . Reversible senescence in human CD4+CD45RA+CD27- memory T cells. J Immunol. 2011;187(5):2093-2100. [DOI] [PubMed] [Google Scholar]
- 10.Jacquemin M, Vantomme V, Buhot C, et al. . CD4+ T-cell clones specific for wild-type factor VIII: a molecular mechanism responsible for a higher incidence of inhibitor formation in mild/moderate hemophilia A. Blood. 2003;101(4):1351-1358. [DOI] [PubMed] [Google Scholar]
- 11.Ettinger RA, James EA, Kwok WW, Thompson AR, Pratt KP. HLA-DR-restricted T-cell responses to factor VIII epitopes in a mild haemophilia A family with missense substitution A2201P. Haemophilia. 2010;16(102):44-55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Reding MT, Wu H, Krampf M, et al. . Sensitization of CD4+ T cells to coagulation factor VIII: response in congenital and acquired hemophilia patients and in healthy subjects. Thromb Haemost. 2000;84(4):643-652. [PubMed] [Google Scholar]
- 13.Hu G-L, Okita DK, Diethelm-Okita BM, Conti-Fine BM. Recognition of coagulation factor VIII by CD4+ T cells of healthy humans. J Thromb Haemost. 2003;1(10):2159-2166. [DOI] [PubMed] [Google Scholar]
- 14.Reding MT, Okita DK, Diethelm-Okita BM, Anderson TA, Conti-Fine BM. Human CD4+ T-cell epitope repertoire on the C2 domain of coagulation factor VIII. J Thromb Haemost. 2003;1(8):1777-1784. [DOI] [PubMed] [Google Scholar]
- 15.Delluc S, Ravot G, Maillere B. Quantitative analysis of the CD4 T-cell repertoire specific to therapeutic antibodies in healthy donors. FASEB J. 2011;25(6):2040-2048. [DOI] [PubMed] [Google Scholar]
- 16.Delluc S, Ravot G, Maillere B. Quantification of the preexisting CD4 T-cell repertoire specific for human erythropoietin reveals its immunogenicity potential. Blood. 2010;116(22):4542-4545. [DOI] [PubMed] [Google Scholar]
- 17.Castelli FA, Szely N, Olivain A, et al. . Hierarchy of CD4 T cell epitopes of the ANRS Lipo5 synthetic vaccine relies on the frequencies of pre-existing peptide-specific T cells in healthy donors. J Immunol. 2013;190(11):5757-5763. [DOI] [PubMed] [Google Scholar]
- 18.Geiger R, Duhen T, Lanzavecchia A, Sallusto F. Human naive and memory CD4+ T cell repertoires specific for naturally processed antigens analyzed using libraries of amplified T cells. J Exp Med. 2009;206(7):1525-1534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Butenas S, Parhami-Seren B, Undas A, Fass DN, Mann KG. The “normal” factor VIII concentration in plasma. Thromb Res. 2010;126(2):119-123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Algiman M, Dietrich G, Nydegger UE, Boieldieu D, Sultan Y, Kazatchkine MD. Natural antibodies to factor VIII (anti-hemophilic factor) in healthy individuals. Proc Natl Acad Sci USA. 1992;89(9):3795-3799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Cannavò A, Valsecchi C, Garagiola I, et al. ; SIPPET study group. Nonneutralizing antibodies against factor VIII and risk of inhibitor development in severe hemophilia A. Blood. 2017;129(10):1245-1250. [DOI] [PubMed] [Google Scholar]
- 22.Richards DM, Ruggiero E, Hofer A-C, et al. . The contained self-reactive peripheral T cell repertoire: size, diversity, and cellular composition. J Immunol. 2015;195(5):2067-2079. [DOI] [PubMed] [Google Scholar]
- 23.Chevaleyre C, Benhamouda N, Favry E, et al. . The tumor antigen cyclin B1 hosts multiple CD4 T cell epitopes differently recognized by pre-existing naive and memory cells in both healthy and cancer conors. J Immunol. 2015;195(4):1891-1901. [DOI] [PubMed] [Google Scholar]
- 24.Kwok WW, Tan V, Gillette L, et al. . Frequency of epitope-specific naive CD4(+) T cells correlates with immunodominance in the human memory repertoire. J Immunol. 2012;188(6):2537-2544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.James EA, Kwok WW, Ettinger RA, Thompson AR, Pratt KP. T-cell responses over time in a mild hemophilia A inhibitor subject: epitope identification and transient immunogenicity of the corresponding self-peptide. J Thromb Haemost. 2007;5(12):2399-2407. [DOI] [PubMed] [Google Scholar]
- 26.Kallas A, Kuuse S, Maimets T, Pooga M. Naturally occurring CD4+ CD25+ cells in modulating immune response to administered coagulation factor VIII in factor VIII-deficient mice. Haemophilia. 2011;17(1):143-151. [DOI] [PubMed] [Google Scholar]
- 27.Ohashi PS, Oehen S, Buerki K, et al. . Ablation of “tolerance” and induction of diabetes by virus infection in viral antigen transgenic mice. Cell. 1991;65(2):305-317. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.