

Abstract
The giant multifunctional protein “OBSCURIN” is encoded by OBSCN gene and is mostly expressed in cardiac and other skeletal muscles responsible for myofibrils organization. Loss of OBSCURIN affects the entire downstream pathway proteins vital for various cellular functions including cell integration and cell adhesion. The OBSCN gene mutations are more frequently observed in various muscular diseases, and cancers. Nevertheless, the direct role of OBSCN in tumorigenesis remains elusive. Interestingly, in clinical breast cancer samples a significant number of function changing mutations have been identified in OBSCN gene. In this study, we identified a significant role of OBSCN by conducting an integrative analysis of copy number alterations, functional mutations, gene methylation and expression data from various BRCA cancer projects data available on cBioPortal and TCGA firebrowse portal. Finally, we carried out genetic network analysis, which revealed that OBSCN gene plays a significant role in GPCR, RAS, p75 or Wnt signaling pathways. Similarly, OBSCN gene interacts with many cancer-associated genes involved in breast tumorigenesis. The OBSCN gene probably regulates breast cancer progression and metastasis and the prognostic molecular signatures such as copy number alterations and gene expression of OBSCN may serve as a tool to identify breast tumorigenesis and metastasis.
Keywords: breast cancer, OBSCN gene, OBSCURIN, cell adhesion, EMT
INTRODUCTION
Cancer in humans is a huge health burden in modern era worldwide. In recent years cancer genomics is assumed as a latest advancement in cancer research, which starts from disease identification and leads to personalized therapy. Owing to this scientific advancement, several key genetic elements have been identified and characterized in various cancer types. In human carcinogenesis events, cells lose their adhesion, integrity and others morphological characteristics and gain the invasive and migratory properties leading to cellular transitions which are called epithelial-to-mesenchymal transition (EMT), which is the most crucial step in initiating cancer metastasis [1, 2]. Large number of genes and their corresponding proteins are involved in EMT associated pathway including OBSCURIN. The giant OBSCURIN is encoded by unique gene called OBSCN located on chromosome 1 at loci q42 [3]. The giant OBSCURIN has two isoforms namely OBSCURIN-A & B [4–6]. The OBSCURIN-A comprises of immunoglobulin (Ig) and fibronectin type-III domains located in the amino terminal while in carboxyl terminal contains several signaling domains including Isoleucin & Glutamine calmodulin-binding (IQ), SRC homology-3 (SH3) domains, pleckstrin homology (PH) and a Rho-guanine nucleotide exchange factor (Rho-GEF) domains which are scattered in the non-modular sequences. The OBSCURIN-B or myosin light chain kinase (MLCK) isoform contains two serine/threonine kinase (STK) domains, which replace the non-modular carboxyl terminus of OBSCURIN-A. Additional to these domains, ERK kinase domain exists for phosphorylation along with two Ig domains [3, 4, 7, 8]. Every OBSCURIN domain regulates vast number of cellular and functional roles such as cell adhesion, migration and cell morphology, etc. Apart from these functional roles, OBSCURIN-A/B are also involved in cellular co-ordination that prevents cells from going in an EMT process [4, 9]. OBSCURIN MLCK family domains chiefly maintain the cellular organizations and contractility [10]. Moreover, a huge number of copy number alterations and mutations were observed in OBSCN gene in many cancer types, although, OBSCN is highly mutated in breast cancer. The main study is to uncover the functional mutations and the active role of OBSCN in breast tumorigenesis. A few studies on this gene also proved that, the reduced or altered OBSCN gene expression largely disturbs the cellular integration and activates cancer initiation; therefore, OBSCN may function as a tumor suppressor gene and prevent cellular transition [11, 12]. OBSCN gene is mostly expressed in many cancer types although the real functional association in cancer is still uncertain. Several studies on OBSCN gene mutations revealed potential roles of OBSCURIN in melanoma, glioblastoma, colorectal, lung, breast and pancreatic cancer [13–15]. Also, our recent study on breast cancer driver mutations genes also emphasized OBSCN as one among the 63 top candidate driver genes [16]. Here we will mainly focus on copy number variation, non-silent mutations, promotor methylation followed by epigenetic changes and their consequences in breast cancer formation.
Mutations and expression of OBSCN: genetic predisposition to tumorigenesis
Recent advances in high throughput sequencing support genomics in a magnificent way. In every cancer, vast numbers of genes are mutated, therefore the identification of real genetic elements responsible for carcinogenesis becomes quiet challenging. OBSCN gene is frequently and consistently mutated in various cancers with a strong correlation with breast, colorectal and other female related cancers. Recent studies revealed that TP53 and OBSCN genes are highly mutated among 189 candidate genes in breast and colorectal cancers [13, 17]. However, scientific background on OBSCN mutation and its impacts are limited in comparison to the other key genes involved in breast cancer [18, 19]. Mutations in OBSCN gene (> 15%) are observed in breast cancer patient samples published by TCGA. Furthermore, a recent study on OBSCN gene in breast cancer revealed that the loss of giant OBSCURIN protein increases the cell migration with more metastatic characteristics [20].
However, recent studies revealed that OBSCURIN expression loss causes functional abnormality, which increases the probability of cancer in human breast epithelial cells. Perry et al. (2012) found that knocking down OBSCN significantly affected breast epithelial cells in both growth and biological property, such as cellular adhesions, cell-cell communications, etc. As a result, expression level of other interlinked proteins such as E-cadherin, α and β-catenin involved in cell-cell junctions were largely affected. Meanwhile, an elevated level of p120-catenin prevents the cell adhesion and stimulates mesenchymal proteins, which stimulate cellular morphogenesis leading to metastasis [21–23]. The actin and microtubule cytoskeleton dysregulation is responsible for cellular transformation and migration found in many cancers and activation and deactivation of Rho-GTPase family proteins majorly regulates actin filaments through Ras homolog gene family, member A (RHOA) signaling. The loss of OBSCN caused RHOA signaling impairment leading to breast cancer initiation, progression and metastasis. [12, 24–28]. The consequent event of OBSCN loss may also affect tubulin microtentacles (McTN) formation. McTN is the key process in metastatic breast cancer and it increases metastatic probability and endothelial coupling followed by circulating tumor cells (CTC) [29–31].
Similarly, low levels of epithelial proteins accelerate phenotypic changes in cells, which further affect focal adhesion and increase of F-actin dynamics in cell-cell contacts [28]. Another interesting study proved that, an active oncogene, K-RAS in the OBSCN knockdown cells increases tumorigenesis probability [9]. The OBSCN down regulation also affected RHOA-mediated pathways leading to dramatic reduction of Rho activated kinase (ROCK) and its targeting proteins such as myosin light chain (MLC), lim kinase (LimK) and cofilin [4, 8, 22, 29, 30]. In this study, we reviewed and analyzed OBSCN gene and its association with breast cancer. The high throughput sequencing data for breast cancer mutations in clinical samples were retrieved from various breast cancer projects available in cBioPortal (www.cbioportal.org), Pan-Cancer (https://www.synapse.org), mRNAseq and Methylation data were retrieved from Firebrowse (http://firebrowse.org/) data portal, which provides multi-tier cancer analysis. The analyses of the overall mutation rates and the mutational patterns of OBSCN gene, as well as their impact on downstream pathways may help us to predict feasible drug targets for breast cancer.
The OBSCN mutations in human cancers using high throughput analysis
Apart from breast cancer, OBSCN gene is widely mutated in many other cancer types as well (Supplementary Figure 1) (www.cbioportal.org). Among the various cancer types breast cancer cases are observed with significant numbers of OBSCN mutations (n = 819) with average mutation frequency of 18%. The overview of this work and study selection are given in the Figure 1. The overall mutations percentage of OBSCN gene in various breast cancer projects are illustrated in Figure 2 (www.cbioportal.org). The large numbers of functional mutations were observed and OBSCN copy number analyses were performed using GISTIC program and many copy gain, amplification and few deletion mutations were observed in major breast cancer projects data (Figure 3). From the overall DNA copy number analysis, we found that OBSCN gene had more number of copy gain and amplification mutations and these variations may potentially affect the giant OBSCURINs protein level and their subsequent expression.
Figure 1. Flow diagram summarizing selection and validation of the present meta-analysis on OBSCN gene.

Figure 2. Overall OBSCN gene mutations across various breast cancer projects data of cBioportal.

(A) number of OBSCN mutations/copy number variations found in each project; (B) Distribution of OBSCN gene mutations across the patient samples (%).
Figure 3. GISTIC copy number analyses of OBSCN gene in various breast cancer projects found with more number of gain and amplification mutations.
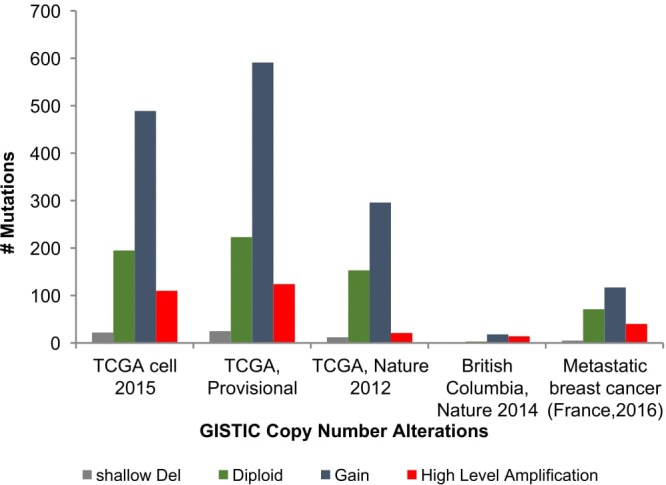
The significant copy number alterations are observed and their outcomes are predicted using GISTIC copy number analysis [32]. The overall mutation profiles of OBSCN gene in breast cancer shallow loss, diploid, gain and high level amplifications are classified based on distinctive copy number profiles and their clinical settings [33, 34]. This analysis also revealed a huge number of gain and high level amplification mutations in OBSCN gene found in invasive ductal carcinoma (IDC), followed by invasive lobular carcinoma (ILC) and so on. These results indicated that OBSCN gene might play an active role in invasion of cancer cells to distant organs and other metastatic processes. The subsequent mRNA expression analysis was performed using breast cancer patient data and based on the median Log2 value, the OBSCN gene is seen down regulated in majority of the mutated TCGA data. To confirm the OBSCN gene expression we also compared with other well-known breast cancer genes such as TP53, PIK3CA, ARID1A, BRCA1, etc. and found that these genes had closer expression levels and it indicated that OBSCN gene mutations may also have a positive correlation with breast cancer dispositions (Figure 4) (https://cran.r-project.org/); (www.cbioportal.org); (https://cancergenome.nih.gov/). In addition to that, OBSCN methylation profiles were also assessed using breast cancer methylation data by calculating beta value and found OBSCN gene apparently hyper-methylated. Then we compared methylation profiles of OBSCN gene over various cancer types and found the gene is hyper-methylated in breast cancer data, which indicates its methylation may also play an important role in disease etiology. We used promotor methylated and gene expression samples of OBSCN gene from TCGA wanderer (http://maplab.imppc.org/wanderer/). The OBSCN methylation and gene expression was compared with normal (n = 98) and tumor (n = 720) samples at cg09411356 and interestingly, the hyper methylation on OBSCN gene significantly reduced gene expression in tumor than the normal samples. The detailed methylation profile of single probe and its corresponding gene expression and comprehensive comparison of methylation pattern over major cancer types is depicted in Figure 5. (https://xenabrowser.net/#). The downregulation of OBSCN may affect cell-cell adhesion of breast epithelial cells and cells undergo EMT process [12]. Further we analyzed the expression of OBSCN gene in various immunohistochemical breast cancer molecular subtypes. Nearly 73% cases found with luminal A (46.99%) and luminal B (26.05%) type of disease followed by other subtypes including basal (10.7%), Her2(8.9%), normal (3.78%) and claudin-low (3.56%). These molecular subtypes classifications distinctly emphasized, OBSCN is vital gene may involve in the cell proliferation. Moreover, several previous studies on OBSCN gene mutations and its association with other cancers were identified by intensive literature search and the detailed information such as cancer types, mutation profiles, co-mutating genes, amino acid variations and supporting citations are tabulated (Table 1).
Figure 4. Comparative evaluation of OBSCN gene expression pattern with other known breast cancer genes shows OBSCN gene has similar expression pattern with some of the known breast cancer driver genes.

Figure 5.

(A) OBSCN methylation profiles of normal (n = 98) and breast tumor samples (n = 720) comparison; (B) Gene expression comparison between normal and tumor samples; (C) Comparison of methylation profiles showed that OBSCN gene largely hypermethylated in breast cancer than other cancer types.
Table 1. The OBSCN gene mutations and their associations with various cancer types.
| Disease Types | Most common co-mutation with OBSCN gene | Major mutations type | Specific Amino acid variations | References |
|---|---|---|---|---|
| Breast Cancer | TP53 | Missense | NA | [13, 21, 35] |
| Lung Cancer; Respiratory Disease | TP53, ARID1A, PTEN, KRAS, MYC, PIK3CA, BRAF, EGFR, NRAS | Missense | NA | [36, 37] |
| Gastrointestinal Stromal tumor and Leiomyosarcoma | C9orf65, TTN, SPEG | Missense | NA | [15, 38, 39] |
| Colorectal cancer | TP53 | Missense | NA | [13, 35] |
| Prostate cancer | CLDN7, STRA13, FLNA FAM83H, CLDN7, ARFGAP3, KDM2A, | Missense | NA | [40, 41] |
| Melanoma | EPHA3, TTN | Somatic Missense | E4574K | [14] |
| Cervical/ Endometrial and Ovarian cancer | Tp53, PIK3CA, FBXW7, NEB, DNAH11, ORAI2, RNF19B, SPTA1, UBA2, UTRN, BSN | Missense | NA | [42] |
| Liver & Pancreatic cancer | ATPAF1, TRPM4, MLL3, ARID1A, ARID1B, ARID2, SOS1, MROH1 | Missense | NA | [14, 19, 43, 44] |
| Glioblastoma | TTN | Germline Missense | R4558H | [13, 14] |
| Esophageal Cancer | C9orf65, TP53, ARID1A, MUC17 | Missense | NA | [45] |
| Sarcoma | C9orf65, PRUNE2 | Missense | NA | [15] |
| Nephroblastoma and other kidney cancers | PTHB1 | Missense | NA | [46, 47] |
| Cardiomyopathy | TTN, MYH7, DSP, VCL, LAMA4, MYOM1, TNNC1, TNNI3 | Truncating Mutation | NA | [47–51] |
Functional analysis of differentially expressed OBSCN gene in breast cancer
Mutations in the OBSCN gene largely affect various interlinked proteins involved in cell adhesion and integration process [52]. Functional mutations in OBSCN largely affects E-cadherin, α-catenin and β-catenin expressions which initiate epithelial cells dispersions and subsequently increase the level of F-actin followed by cell migration [20, 53–55]. Actin filaments are vital for cellular mechanistic amoeboid movements accelerated by filapodia or pseudopodia like structures formed in cytoplasmic regions [56, 57]. A recent study showed that OBSCN functional gene mutations are responsible for the loss of vital protein expressions [21]. The loss of OBSCN appears to be consequent event in primary to metastatic breast cancer majorly affecting cellular integration. The protein kinases (PKs) are from the larger protein family involved in various cellular, structural and functional mechanisms in humans. The Serine/Threonine Kinase (STK) is one of the important protein kinase involved in cellular cytoskeletal and integration roles such as cell proliferation, differentiation and apoptotic process, etc. [58, 59]. The STK actively binds with p120-catenin and regulates cadherin facilitated intercellular adhesions and E-cadherin-p120 bound complex also plays a crucial role in maintaining the equilibrium of cadherin levels [60, 61]. The OBSCN mutations affect E-cadherin level, lead to flux in adherens junctions and increase cell disintegration followed by cell movement [62]. The OBSCN gene mutations heavily affect STK protein activity and also act as a ligand for small ankyrin1 (sANK1). sANK1 are integral proteins to the fundamental spectrin-actin cytoskeleton involved in cellular movement activation and proliferation, which cause metastasis [63]. Moreover, OBSCN gene also acts as a kinome regulatory element that regulates few other kinase family proteins in either direct or indirect manner.
Gene interaction analysis revealed that OBSCN gene is positively correlated with protein binding and cell transition associated genes
The OBSCN gene mutations have significant relationship with genes responsible for cellular structural and functional roles. Also, the expression of OBSCN gene is positively correlated with few other genes such as TOP1MT, MYC, TARPBP1, ADCK3, TRIM17, and RND2. Initially we used metastatic breast cancer data (Metastatic Breast cancer-France 2016) for network analysis and revealed many participating genes such as RHOT2, RAC3, RHOD, ARHGEF7, PAK1, PDPK1, AKT3, RIPK2, ERBB2, PRKD1, etc. which are positively correlated with amplification mutations and have strong association with cell proliferation, differentiation, tumorigenesis and other metastasis features. Similarly, we used British Columbia Xenografts (British Columbia, Nature 2014) data linked with many interesting genes such as RHOB, ITSN1, RHOD, ERBB2, AKT3, BRAF, CDK1, MASTL along with eight more genes which were highly correlated with cell signaling and cell migration functions. The breast invasive carcinoma (TCGA, Provisional) data showed that the OBSCN had strong interaction with PAK1 gene through RHOU and RAC3 genes. The breast cancer oncogene, PAK1, acts as a hub gene connecting two sub networks. In first sub network, PDPK1 gene a central gene linked with other significant genes such as AKT3, ERBB2, SGK3, PRKCB, PRKCQ, etc. and the second sub network is connected with STK3, AURKA, CSNK1D, NEK2, etc. The PAK1 and PDPK1 genes are strongly associated with breast tumorigenesis and cell migration in metastatic event [64, 65]. Along with the afore-mentioned genes, OBSCN is interconnected with other genes such as PAK2, AKT1, TP53RK, NEK2, WNK1, CSNK1D, etc. which are frequently mutated in invasive breast carcinoma (Nature 2012) (Cell 2015). The detailed OBSCN mutation specific network neighbors were identified using functional protein association network using STRING (http://string-db.org/). Additional pathway specific networks were also used to identify and validate the genes involved in OBSCN associated pathway using PCViz (http://www.pathwaycommons.org/) and detailed networks are illustrated in Figure 6.
Figure 6.

(A) Mutation specific OBSCN functional association network and its interacting network partners identified using STRING database; (B) Gene specific interactions pathways were identified using pathway commons network visualizer and many interacting partners of OBSCN are cancer-associated genes.
OBSCN genetic alterations lead to cell survival and switches towards EMT
Substantial evidences propose that OBSCN functional mutations may have strong associations with Wnt signaling regulated by a series of activators along with β-catenin [66]. In the nucleus, β-catenin binds to transcription activation elements (TCF/LEF), leading to the activation of the target genes (specifically CCND1, c-MYC, and FN1) responsible for the regulation of cell proliferation and differentiation [67–69]. Mutational event of OBSCN gene majorly affects β-catenin downstream signaling leading to lower down or over expression of genes such as CCND1, c-MYC, and FN1, which stimulates several types of cancers including breast, ovarian, lung, pancreatic, colorectal, and uterine carcinoma [70–75]. The elevated level of fibronectin or loss of proteins such as E-CADHERIN, Β-CATENIN, etc. may cause cellular proliferation followed by EMT, which render patients overall survival. Henceforth, OBSCN gene is believed to be an indirect regulator of fibronectin expression in breast and colon cancer [72, 76]. The fibronectin malfunction may also affect vital proteins such as integrin and other transmembrane receptors responsible for cell-cell contact, extracellular matrix (ECM) communications, cell adhesion and accommodate several ligands such as collagen, laminin and vitronectin. Fibronectin is important protein which activates integrin and is involved in the regulation of cellular signal transduction, cell proliferation, gene expression and cytoskeletal reorganization [77]. Hence, the OBSCN gene mutation perhaps affects the regulation of β-catenin and other regulatory elements in Wnt signaling pathway largely affects mainly the downstream genes and proteins expressions involved [78]. Interestingly in metastatic cancer, OBSCN mutation observed with new list of interacting genes in Ras protein family members such as RHOD, RHOT2 and RAC3 are involved in major functions such as reorganization of actin cytoskeleton, GTPase activity, formation of lamellipodia, respectively [79–81, 99]. The RAC3 gene is important gene activates PAK1, which is vital to stimulates F-actin formation leads to cell migration. Similarly, PAK1 activates another important proliferation related protein, aurora kinase (AURKA) and it inhibits IKB-α, which inhibits apoptosis through NF-ĸB and CIAP1 genes [82, 83]. The detailed OBSCN gene involved cancer-associated pathway illustrated in Figure 7. The Panther database (http://pantherdb.org/) was used to further validate gene ontology based molecular functions analysis and further confirmed by evaluating fold enrichment scores [84]. The OBSCN gene and their networks neighbors were involved in binding and cell regulatory mechanisms including cell adhesion and binding with several receptors and signaling molecules (Figure 8).
Figure 7. The OBSCN gene involved either direct or indirect pathway of GPCR, Ras, p75 or Wnt signaling.

(In GPCR pathway, OBSCN-RhoGEF is simultaneously regulates mDIA, Rho and their downstream genes which are involved in cell differentiation and lamellipodia formation; the other chain of OBSCN, MLCK and its downstream pathway activates MLC leads to formation of stress fiber which cause cell differentiation and survival; On the other way RhoA activated from p75 (NTR) pathway also involved in Ras pathway regulates down regulates MLCK; in Wnt signaling OBSCN disequilibrium leads to β-catenin loss which leads to carcinogenesis.
Figure 8. Molecular functional classification analysis of OBSCN gene and its network neighbor genes showed that OBSCN gene majorly involved in binding activity with various molecules.

Mutational tolerance profiles and their consequences in proteome level
Recent studies on OBSCN mutations uncover key amino acids mutations, which are crucial factor for many human diseases including cardiomyopathy and cancers. [49, 51, 85]. In this study, we also attempted to know the deep impact of OBSCN mutations including the proteome level. The OBSCN gene mutational protein variants retrieved from Ensembl genome browser (http://asia.ensembl.org/) and their effects were predicted using Variant Effect Predictor (VEP) tool (http://asia.ensembl.org/Tools/VEP) and wANNOVAR (http://wannovar.wglab.org/) [86, 87]. The SIFT (http://sift.jcvi.org/) and PolyPhen-2 (http://genetics.bwh.harvard.edu/pph2/) tools were used to achieve the most reliable protein variants prediction, and we also considered few other variants impact prediction algorithms such as Log ratio test (LRT), Mutation Taster (http://www.mutationtaster.org/), Mutation Assessor (http://mutationassessor.org/r3/), FATHMM (http://fathmm.biocompute.org.uk/) and Variant effect scoring tool 3 (VEST3)(http://karchinlab.org/apps/appVest.html), etc. Most of the abovementioned tools predict the variants and their impacts based on sequence changes and very few tools also consider structural changes [88–96]. The variants predicted by the maximum number of tools were used to filter and used for further analysis (Figure 9) (http://compbio.berkeley.edu/proj/varant/manual.html). From the overall analysis, it was seen that OBSCN gene had a low tolerance against few functional mutations (total of 251 protein variants), half of the variants (~48%) exhibited deleterious and probably damaging effects (Supplementary Table 1), and rest of the variants came in deleterious & possibly damaging and deleterious with benign categories [95, 97, 98]. The overlapping mutational impact variants were taken and further analyzed the somatic mutations using COSMIC database. The detailed OBSCN mutational impact on protein levels are emphasized using lollipop plot of maftools (Bioconductor R-package) and amino acids variations and their impacts are indicated in various colors representations along with mutations count (Figure 10) (https://github.com/PoisonAlien/mafTools). In addition, the key mutational variants, types, amino acid variations and impacts in domain levels are listed in Table 2.
Figure 9.

(A) SIFT and POLYPHEN-2 programs were used to predict the most common and significant OBSCN mutational impact variants; (B) To validate the high-impact variants and their mutations impact are predicted using 10 potential impact prediction tools.
Figure 10. Lollipop plot function of maftools represents functional amino acids mutations and their impacts on various domains of OBSCN gene are indicated in different colors representation.

Table 2. The significant OBSCN mutation variants and their impacts filtered from cBioPortal breast cancer patient tumor samples.
| Genomic Coordinate | Reference and Variants | Mutation Type | Copy Number | Amino acid Variant | Major Domain Affected |
|---|---|---|---|---|---|
| 228402602 | C>G | Missense | AMP | T544S | Fibronectin-III |
| 228412169 | NA | Missense | Gain | V888G | Ig-Domain |
| 228432150 | T>G | Missense | Gain | V1212G | Ig-Domain |
| 228432150 | NA | Missense | Gain | V1120G | Ig-Domain |
| 228432244 | G>T | Missense | Gain | Q1243H | Ig-Domain |
| 228434340 | C>T | Missense | Diploid | T1382M | Ig-Domain |
| 228461665 | G>A | Missense | Gain | V1778M | Fibronectin-III |
| 228464221 | G>- | Frameshift DEL | Del | V2282fs | Ig-Domain |
| 228464225 | ->G | Frameshift INS | Gain | Q2283fs | Ig-Domain |
| 228479785 | NA | Missense | Gain | G3509V | Ig-Domain |
| 228479785 | G>T | Missense | Gain | G3938V | Ig-Domain |
| 228479785 | G>T | Missense | Gain | G3693V | Ig-Domain |
| 228480325 | T>C | Missense | Gain | W3998R | Ig-Domain |
| 228480325 | T>C | Missense | Gain | W3753R | Ig-Domain |
| 228480325 | NA | Missense | Gain | W3569R | Ig-Domain |
| 228481236 | C>T | Missense | Gain | R3868C | Ig-Domain |
| 228491624 | AC>- | Frameshift DEL | Del | L4419fs | Ig-Domain |
| 228492905 | G>C | Missense | Gain | S4538T | Ig-Domain |
| 228494273 | G>A | Missense | Gain | E4911K | Ig-Domain |
| 228494273 | NA | Missense | Gain | E3954K | Ig-Domain |
| 228494979 | NA | Missense | AMP | K4071N | Ig-Domain |
| 228497221 | G>A | Missense | Gain | V5282M | Ig-Domain |
| 228497221 | NA | Missense | Gain | V4325M | Ig-Domain |
| 228497221 | G>A | Missense | Gain | V5037M | Ig-Domain |
| 228503574 | G>A | Missense | Gain | E5059Q | Ig-Domain |
| 228506585 | T>C | Missense | Gain | V5668A | SH3 |
| 228506585 | NA | Missense | Gain | V4711A | Ig-Domain |
| 228506585 | T>C | Missense | Gain | V5423A | Ig-Domain |
| 228506947 | G>A | Missense | Gain | A5544T | NA |
| 228509882 | G>A | Missense | Gain | E5826K | NA |
| 228521392 | NA | Missense | Gain | S5322C | Ig-Domain |
| 228521466 | C>T | Missense | Gain | R6304W | NA |
| 228521466 | NA | Missense | Gain | R5347W | Ig-Domain |
| 228521466 | C>T | Missense | Gain | R6059W | Ig-Domain |
| 228537694 | ->T | Frameshift INS | Gain | Y6796fs | NA |
| 228538601 | C>T | Nonsense | Gain | Q6838* | Ig-Domain |
| 228547542 | G>A | Missense | Gain | G6317R | NA |
| 228556454 | G>T | Missense | Gain | R7312L | STK-cat Domain |
| 228559643 | C>A | Missense | Gain | P7767Q | NA |
| 228559663 | C>T | Missense | Gain | P7774S | Protein Kinase 2 |
*Ig- Immunoglobulin. AMP-Amplification; Del-Deletion
OBSCN mediated targeted anti-cancer therapy: a present and future perspectives
Several interesting genes are interlinked with tumorigenesis including OBSCN gene, which is considered the key gene among them. The OBSCN gene may act as unique target for anti-cancer therapy in breast and other cancers, since it is having multiple interactions with intra and inter-cellular levels of numerous interconnecting proteins. In breast cancer targeted anti-cancer therapy, several key genes are targeted such as CTNND1, CDC42 and DVL. Over expression of P120-CATENIN tremendously inhibited several signaling molecules such as RHOA, RAC1 and CDC42 which are involved in the regulation of numerous cellular functions [100]. Similarly, downregulation of CDC42 initiated cancer. Interestingly OBSCN gene is an active target regulating CDC42 downstream pathway accounting for the tumorigenesis [101]. DVL (Dishevelled homology1) protein phosphorylation are vital phenomenon accelerated by CK1ε and RIPK4 in Wnt signaling pathway [102]. Hence, reduced β-catenin level and LRP6 association facilitates further downstream signaling of β-catenin, which activates transcription factors/ lymphoid enhancer-binding factors (TCF/LEF) involved in numerous cellular regulations and cell migrations. An alternative and most interesting target is DVL expression responsible for calmodulin-dependent protein kinase (CamK-II) activation followed by expression of F-actin. The F-actin is the key protein involved in cell division, contraction and locomotion and altered F-actin dynamics is most vital feature for epithelial to mesenchymal transition (EMT) of metastatic breast cancer. Hence, anti-cancer drug targets may actively repress DVL and it may potentially block F-actin dynamics, which inhibits EMT process in cancer cells, and the overall phenomena, is indirectly regulated by OBSCN gene.
Concluding remarks
We have analyzed OBSCN gene mutation, expression and methylation data and the data revealed that OBSCN gene is one of most frequently mutated gene in various cancer types, especially in breast cancer. The OBSCN gene mutation may play an essential role during cancer initiation and progression, largely distressed by various kinds of mutations in gene level due to loss of heterozygosity, various oncogenic factors, intra and intercellular environmental stress. Mutations in OBSCN gene largely affects multiple properties of cells including cell adhesion and increase the integration and render cellular transitions and many more. Evaluation of OBSCN mutational status may help early prognosis of metastatic potential of breast cancer. Mutational, copy number and epigenetic analysis of OBSCN gene status may serve as a tool for the prediction of targeted anti-cancer drugs, which will be helpful for targeted breast cancer therapy [11, 20]. Further functional studies on OBSCN gene in various model systems with gene overexpression and/or targeted disruption should also greatly facilitate these processes.
SUPPLEMENTARY MATERIALS FIGURES AND TABLES
ACKNOWLEDGMENTS AND FUNDING
We thank members of the Professor Deng laboratory for critical reading and discussion. This work is supported by the Chair Professor Grant (CPG2017-00026-FHS), MYRG2016-00132-FHS and MYRG2016-00139 of University of Macau and FDCT grants (065/2015/A2 and 094/2015/A3) to Chu-Xia Deng.
Footnotes
CONFLICTS OF INTEREST
Authors declare no conflicts of interest.
REFERENCES
- 1.Kalluri R, Weinberg RA. The basics of epithelial-mesenchymal transition. J Clin Invest. 2009;119:1420–1428. doi: 10.1172/JCI39104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Thiery JP, Acloque H, Huang RY, Nieto MA. Epithelial-mesenchymal transitions in development and disease. Cell. 2009;139:871–890. doi: 10.1016/j.cell.2009.11.007. [DOI] [PubMed] [Google Scholar]
- 3.Russell MW, Raeker MO, Korytkowski KA, Sonneman KJ. Identification, tissue expression and chromosomal localization of human Obscurin-MLCK, a member of the titin and Dbl families of myosin light chain kinases. Gene. 2002;282:237–246. doi: 10.1016/s0378-1119(01)00795-8. [DOI] [PubMed] [Google Scholar]
- 4.Young P, Ehler E, Gautel M. Obscurin, a giant sarcomeric Rho guanine nucleotide exchange factor protein involved in sarcomere assembly. J Cell Biol. 2001;154:123–136. doi: 10.1083/jcb.200102110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Borisov AB, Raeker MO, Kontrogianni-Konstantopoulos A, Yang K, Kurnit DM, Bloch RJ, Russell MW. Rapid response of cardiac obscurin gene cluster to aortic stenosis: differential activation of Rho-GEF and MLCK and involvement in hypertrophic growth. Biochem Biophys Res Commun. 2003;310:910–918. doi: 10.1016/j.bbrc.2003.09.035. [DOI] [PubMed] [Google Scholar]
- 6.Kontrogianni-Konstantopoulos A, Catino DH, Strong JC, Bloch RJ. De novo myofibrillogenesis in C2C12 cells: evidence for the independent assembly of M bands and Z disks. Am J Physiol Cell Physiol. 2006;290:C626–637. doi: 10.1152/ajpcell.00442.2005. [DOI] [PubMed] [Google Scholar]
- 7.Kontrogianni-Konstantopoulos A, Ackermann MA, Bowman AL, Yap SV, Bloch RJ. Muscle giants: molecular scaffolds in sarcomerogenesis. Physiol Rev. 2009;89:1217–1267. doi: 10.1152/physrev.00017.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Fukuzawa A, Idowu S, Gautel M. Complete human gene structure of obscurin: implications for isoform generation by differential splicing. J Muscle Res Cell Motil. 2005;26:427–434. doi: 10.1007/s10974-005-9025-6. [DOI] [PubMed] [Google Scholar]
- 9.Petersen TE, Thogersen HC, Skorstengaard K, Vibe-Pedersen K, Sahl P, Sottrup-Jensen L, Magnusson S. Partial primary structure of bovine plasma fibronectin: three types of internal homology. Proc Natl Acad Sci USA. 1983;80:137–141. doi: 10.1073/pnas.80.1.137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kamm KE, Stull JT. Signaling to myosin regulatory light chain in sarcomeres. J Biol Chem. 2011;286:9941–9947. doi: 10.1074/jbc.R110.198697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Perry NA, Vitolo MI, Martin SS, Kontrogianni-Konstantopoulos A. Loss of the obscurin-RhoGEF downregulates RhoA signaling and increases microtentacle formation and attachment of breast epithelial cells. Oncotarget. 2014;5:8558–8568. doi: 10.18632/oncotarget.2338. https://doi.org/10.18632/oncotarget.2338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Shriver M, Stroka KM, Vitolo MI, Martin S, Huso DL, Konstantopoulos K, Kontrogianni-Konstantopoulos A. Loss of giant obscurins from breast epithelium promotes epithelial-to-mesenchymal transition, tumorigenicity and metastasis. Oncogene. 2015;34:4248–4259. doi: 10.1038/onc.2014.358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Sjöblom T, Jones S, Wood LD, Parsons DW, Lin J, Barber TD, Mandelker D, Leary RJ, Ptak J, Silliman N, Szabo S, Buckhaults P, Farrell C, et al. The consensus coding sequences of human breast and colorectal cancers. Science. 2006;314:268–274. doi: 10.1126/science.1133427. [DOI] [PubMed] [Google Scholar]
- 14.Balakrishnan A, Bleeker FE, Lamba S, Rodolfo M, Daniotti M, Scarpa A, van Tilborg AA, Leenstra S, Zanon C, Bardelli A. Novel somatic and germline mutations in cancer candidate genes in glioblastoma, melanoma, and pancreatic carcinoma. Cancer Res. 2007;67:3545–3550. doi: 10.1158/0008-5472.CAN-07-0065. [DOI] [PubMed] [Google Scholar]
- 15.Price ND, Trent J, El-Naggar AK, Cogdell D, Taylor E, Hunt KK, Pollock RE, Hood L, Shmulevich I, Zhang W. Highly accurate two-gene classifier for differentiating gastrointestinal stromal tumors and leiomyosarcomas. Proc Natl Acad Sci USA. 2007;104:3414–3419. doi: 10.1073/pnas.0611373104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Rajendran BK, Deng CX. Characterization of potential driver mutations involved in human breast cancer by computational approaches. Oncotarget. 2017;8:50252–50272. doi: 10.18632/oncotarget.17225. https://doi.org/10.18632/oncotarget.17225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Liu Y, Hu Z, DeLisi C. Mutated Pathways as a Guide to Adjuvant Therapy Treatments for Breast Cancer. Mol Cancer Ther. 2016;15:184–189. doi: 10.1158/1535-7163.MCT-15-0601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Waalkes S, Atschekzei F, Kramer MW, Hennenlotter J, Vetter G, Becker JU, Stenzl A, Merseburger AS, Schrader AJ, Kuczyk MA, Serth J. Fibronectin 1 mRNA expression correlates with advanced disease in renal cancer. BMC Cancer. 2010;10:503. doi: 10.1186/1471-2407-10-503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bii VM, Trobridge GD. Identifying Cancer Driver Genes Using Replication-Incompetent Retroviral Vectors. Cancers (Basel) 2016:8. doi: 10.3390/cancers8110099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Shriver M, Marimuthu S, Paul C, Geist J, Seale T, Konstantopoulos K, Kontrogianni-Konstantopoulos A. Giant obscurins regulate the PI3K cascade in breast epithelial cells via direct binding to the PI3K/p85 regulatory subunit. Oncotarget. 2016;7:45414–45428. doi: 10.18632/oncotarget.9985. https://doi.org/10.18632/oncotarget.9985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Perry NA, Shriver M, Mameza MG, Grabias B, Balzer E, Kontrogianni-Konstantopoulos A. Loss of giant obscurins promotes breast epithelial cell survival through apoptotic resistance. FASEB J. 2012;26:2764–2775. doi: 10.1096/fj.12-205419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Anastasiadis PZ, Reynolds AB. The p120 catenin family: complex roles in adhesion, signaling and cancer. J Cell Sci. 2000;113:1319–1334. doi: 10.1242/jcs.113.8.1319. [DOI] [PubMed] [Google Scholar]
- 23.Yap AS. The morphogenetic role of cadherin cell adhesion molecules in human cancer: a thematic review. Cancer Invest. 1998;16:252–261. doi: 10.3109/07357909809039774. [DOI] [PubMed] [Google Scholar]
- 24.Vega FM, Ridley AJ. Rho GTPases in cancer cell biology. FEBS Lett. 2008;582:2093–2101. doi: 10.1016/j.febslet.2008.04.039. [DOI] [PubMed] [Google Scholar]
- 25.Rossman KL, Der CJ Sondek J. GEF means go: turning on RHO GTPases with guanine nucleotide-exchange factors. Nat Rev Mol Cell Biol. 2005;6:167–180. doi: 10.1038/nrm1587. [DOI] [PubMed] [Google Scholar]
- 26.Hall A. The cytoskeleton and cancer. Cancer Metastasis Rev. 2009;28:5–14. doi: 10.1007/s10555-008-9166-3. [DOI] [PubMed] [Google Scholar]
- 27.Sahai E. Mechanisms of cancer cell invasion. Curr Opin Genet Dev. 2005;15:87–96. doi: 10.1016/j.gde.2004.12.002. [DOI] [PubMed] [Google Scholar]
- 28.Yamaguchi H, Condeelis J. Regulation of the actin cytoskeleton in cancer cell migration and invasion. Biochim Biophys Acta. 2007:642–652. doi: 10.1016/j.bbamcr.2006.07.001. 1773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Whipple RA, Balzer EM, Cho EH, Matrone MA, Yoon JR, Martin SS. Vimentin filaments support extension of tubulin-based microtentacles in detached breast tumor cells. Cancer Res. 2008;68:5678–5688. doi: 10.1158/0008-5472.CAN-07-6589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Balzer EM, Whipple RA, Thompson K, Boggs AE, Slovic J, Cho EH, Matrone MA, Yoneda T, Mueller SC, Martin SS. c-Src differentially regulates the functions of microtentacles and invadopodia. Oncogene. 2010;29:6402–6408. doi: 10.1038/onc.2010.360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Matrone MA, Whipple RA, Thompson K, Cho EH, Vitolo MI, Balzer EM, Yoon JR, Ioffe OB, Tuttle KC, Tan M, Martin SS. Metastatic breast tumors express increased tau, which promotes microtentacle formation and the reattachment of detached breast tumor cells. Oncogene. 2010;29:3217–3227. doi: 10.1038/onc.2010.68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Mermel CH, Schumacher SE, Hill B, Meyerson ML, Beroukhim R, Getz G. GISTIC2.0 facilitates sensitive and confident localization of the targets of focal somatic copy-number alteration in human cancers. Genome Biol. 2011;12:R41. doi: 10.1186/gb-2011-12-4-r41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Shen R, Olshen AB, Ladanyi M. Integrative clustering of multiple genomic data types using a joint latent variable model with application to breast and lung cancer subtype analysis. Bioinformatics. 2009;25:2906–2912. doi: 10.1093/bioinformatics/btp543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Ali HR, Rueda OM, Chin SF, Curtis C, Dunning MJ, Aparicio SA, Caldas C. Genome-driven integrated classification of breast cancer validated in over 7,500 samples. Genome Biol. 2014;15:431. doi: 10.1186/s13059-014-0431-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Caldas C, Edwards PA. A quantum leap in our knowledge of breast cancer mutations. Breast Cancer Res. 2006;8:304. doi: 10.1186/bcr1624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Babur O, Gonen M, Aksoy BA, Schultz N, Ciriello G, Sander C, Demir E. Systematic identification of cancer driving signaling pathways based on mutual exclusivity of genomic alterations. Genome Biol. 2015;16:45. doi: 10.1186/s13059-015-0612-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kim JH, Park B, Pasaje CF, Kim Y, Bae JS, Park JS, Uh ST, Kim YH, Kim MK, Choi IS, Cho SH, Choi BW, Koh I, et al. Contribution of the OBSCN nonsynonymous variants to aspirin exacerbated respiratory disease susceptibility in Korean population. DNA Cell Biol. 2012;31:1001–1009. doi: 10.1089/dna.2011.1436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Greenman C, Stephens P, Smith R, Dalgliesh GL, Hunter C, Bignell G, Davies H, Teague J, Butler A, Stevens C, Edkins S, O'Meara S, Vastrik I, et al. Patterns of somatic mutation in human cancer genomes. Nature. 2007;446:153–158. doi: 10.1038/nature05610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Zang ZJ, Ong CK, Cutcutache I, Yu W, Zhang SL, Huang D, Ler LD, Dykema K, Gan A, Tao J, Lim S, Liu Y, Futreal PA, et al. Genetic and structural variation in the gastric cancer kinome revealed through targeted deep sequencing. Cancer Res. 2011;71:29–39. doi: 10.1158/0008-5472.CAN-10-1749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Nalla AK, Williams TF, Collins CP, Rae DT, Trobridge GD. Lentiviral vector-mediated insertional mutagenesis screen identifies genes that influence androgen independent prostate cancer progression and predict clinical outcome. Mol Carcinog. 2016;55:1761–1771. doi: 10.1002/mc.22425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Schinke EN, Bii V, Nalla A, Rae DT, Tedrick L, Meadows GG, Trobridge GD. A novel approach to identify driver genes involved in androgen-independent prostate cancer. Mol Cancer. 2014;13:120. doi: 10.1186/1476-4598-13-120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kuhn E, Wu RC, Guan B, Wu G, Zhang J, Wang Y, Song L, Yuan X, Wei L, Roden RB, Kuo KT, Nakayama K, Clarke B, et al. Identification of molecular pathway aberrations in uterine serous carcinoma by genome-wide analyses. J Natl Cancer Inst. 2012;104:1503–1513. doi: 10.1093/jnci/djs345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Fujimoto A, Totoki Y, Abe T, Boroevich KA, Hosoda F, Nguyen HH, Aoki M, Hosono N, Kubo M, Miya F, Arai Y, Takahashi H, Shirakihara T, et al. Whole-genome sequencing of liver cancers identifies etiological influences on mutation patterns and recurrent mutations in chromatin regulators. Nat Genet. 2012;44:760–764. doi: 10.1038/ng.2291. [DOI] [PubMed] [Google Scholar]
- 44.Ranzani M, Annunziato S, Calabria A, Brasca S, Benedicenti F, Gallina P, Naldini L, Montini E. Lentiviral vector-based insertional mutagenesis identifies genes involved in the resistance to targeted anticancer therapies. Mol Ther. 2014;22:2056–2068. doi: 10.1038/mt.2014.174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Shimizu T, Marusawa H, Matsumoto Y, Inuzuka T, Ikeda A, Fujii Y, Minamiguchi S, Miyamoto S, Kou T, Sakai Y, Crabtree JE, Chiba T. Accumulation of somatic mutations in TP53 in gastric epithelium with Helicobacter pylori infection. Gastroenterology. 2014;147:407–417 e403. doi: 10.1053/j.gastro.2014.04.036. [DOI] [PubMed] [Google Scholar]
- 46.Vernon EG, Malik K, Reynolds P, Powlesland R, Dallosso AR, Jackson S, Henthorn K, Green ED, Brown KW. The parathyroid hormone-responsive B1 gene is interrupted by a t(1;7)(q42;p15) breakpoint associated with Wilms’ tumour. Oncogene. 2003;22:1371–1380. doi: 10.1038/sj.onc.1206332. [DOI] [PubMed] [Google Scholar]
- 47.Perry NA, Ackermann MA, Shriver M, Hu LY, Kontrogianni-Konstantopoulos A. Obscurins: unassuming giants enter the spotlight. IUBMB Life. 2013;65:479–486. doi: 10.1002/iub.1157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Wu Y, Bell SP, Trombitas K, Witt CC, Labeit S, LeWinter MM, Granzier H. Changes in titin isoform expression in pacing-induced cardiac failure give rise to increased passive muscle stiffness. Circulation. 2002;106:1384–1389. doi: 10.1161/01.cir.0000029804.61510.02. [DOI] [PubMed] [Google Scholar]
- 49.Arimura T, Matsumoto Y, Okazaki O, Hayashi T, Takahashi M, Inagaki N, Hinohara K, Ashizawa N, Yano K, Kimura A. Structural analysis of obscurin gene in hypertrophic cardiomyopathy. Biochem Biophys Res Commun. 2007;362:281–287. doi: 10.1016/j.bbrc.2007.07.183. [DOI] [PubMed] [Google Scholar]
- 50.Borisov AB, Sutter SB, Kontrogianni-Konstantopoulos A, Bloch RJ, Westfall MV, Russell MW. Essential role of obscurin in cardiac myofibrillogenesis and hypertrophic response: evidence from small interfering RNA-mediated gene silencing. Histochem Cell Biol. 2006;125:227–238. doi: 10.1007/s00418-005-0069-x. [DOI] [PubMed] [Google Scholar]
- 51.Marston S, Montgiraud C, Munster AB, Copeland O, Choi O, Dos Remedios C, Messer AE, Ehler E, Knoll R. OBSCN Mutations Associated with Dilated Cardiomyopathy and Haploinsufficiency. PLoS One. 2015;10:e0138568. doi: 10.1371/journal.pone.0138568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Schlessinger J, Ullrich A. Growth factor signaling by receptor tyrosine kinases. Neuron. 1992;9:383–391. doi: 10.1016/0896-6273(92)90177-f. [DOI] [PubMed] [Google Scholar]
- 53.Rimm DL, Koslov ER, Kebriaei P, Cianci CD, Morrow JS. Alpha 1(E)-catenin is an actin-binding and -bundling protein mediating the attachment of F-actin to the membrane adhesion complex. Proc Natl Acad Sci USA. 1995;92:8813–8817. doi: 10.1073/pnas.92.19.8813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Shankar J, Nabi IR. Actin cytoskeleton regulation of epithelial mesenchymal transition in metastatic cancer cells. PLoS One. 2015;10:e0119954. doi: 10.1371/journal.pone.0119954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Mege RM, Ishiyama N. Integration of Cadherin Adhesion and Cytoskeleton at Adherens Junctions. Cold Spring Harb Perspect Biol. 2017. [DOI] [PMC free article] [PubMed]
- 56.Arjonen A, Kaukonen R, Ivaska J. Filopodia and adhesion in cancer cell motility. Cell Adh Migr. 2011;5:421–430. doi: 10.4161/cam.5.5.17723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Jacquemet G, Hamidi H, Ivaska J. Filopodia in cell adhesion, 3D migration and cancer cell invasion. Curr Opin Cell Biol. 2015;36:23–31. doi: 10.1016/j.ceb.2015.06.007. [DOI] [PubMed] [Google Scholar]
- 58.Capra M, Nuciforo PG, Confalonieri S, Quarto M, Bianchi M, Nebuloni M, Boldorini R, Pallotti F, Viale G, Gishizky ML, Draetta GF, Di Fiore PP. Frequent alterations in the expression of serine/threonine kinases in human cancers. Cancer Res. 2006;66:8147–8154. doi: 10.1158/0008-5472.CAN-05-3489. [DOI] [PubMed] [Google Scholar]
- 59.Lahiry P, Torkamani A, Schork NJ, Hegele RA. Kinase mutations in human disease: interpreting genotype-phenotype relationships. Nat Rev Genet. 2010;11:60–74. doi: 10.1038/nrg2707. [DOI] [PubMed] [Google Scholar]
- 60.Ireton RC, Davis MA, van Hengel J, Mariner DJ, Barnes K, Thoreson MA, Anastasiadis PZ, Matrisian L, Bundy LM, Sealy L, Gilbert B, Roy VF, Reynolds AB. A novel role for p120 catenin in E-cadherin function. J Cell Biol. 2002;159:465–476. doi: 10.1083/jcb.200205115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Xia X, Mariner DJ, Reynolds AB. Adhesion-associated and PKC-modulated changes in serine/threonine phosphorylation of p120-catenin. Biochemistry. 2003;42:9195–9204. doi: 10.1021/bi034597h. [DOI] [PubMed] [Google Scholar]
- 62.Heuberger J, Birchmeier W. Interplay of cadherin-mediated cell adhesion and canonical Wnt signaling. Cold Spring Harb Perspect Biol. 2010;2:a002915. doi: 10.1101/cshperspect.a002915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Kontrogianni-Konstantopoulos A, Jones EM, Van Rossum DB, Bloch RJ. Obscurin is a ligand for small ankyrin 1 in skeletal muscle. Mol Biol Cell. 2003;14:1138–1148. doi: 10.1091/mbc.E02-07-0411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Shrestha Y, Schafer EJ, Boehm JS, Thomas SR, He F, Du J, Wang S, Barretina J, Weir BA, Zhao JJ, Polyak K, Golub TR, Beroukhim R, Hahn WC. PAK1 is a breast cancer oncogene that coordinately activates MAPK and MET signaling. Oncogene. 2012;31:3397–3408. doi: 10.1038/onc.2011.515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Du J, Yang M, Chen S, Li D, Chang Z, Dong Z. PDK1 promotes tumor growth and metastasis in a spontaneous breast cancer model. Oncogene. 2016;35:3314–3323. doi: 10.1038/onc.2015.393. [DOI] [PubMed] [Google Scholar]
- 66.Gottardi CJ, Gumbiner BM. Distinct molecular forms of beta-catenin are targeted to adhesive or transcriptional complexes. J Cell Biol. 2004;167:339–349. doi: 10.1083/jcb.200402153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Gao ZH, Lu C, Wang MX, Han Y, Guo LJ. Differential beta-catenin expression levels are associated with morphological features and prognosis of colorectal cancer. Oncol Lett. 2014;8:2069–2076. doi: 10.3892/ol.2014.2433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Maretzky T, Reiss K, Ludwig A, Buchholz J, Scholz F, Proksch E, de Strooper B, Hartmann D, Saftig P. ADAM10 mediates E-cadherin shedding and regulates epithelial cell-cell adhesion, migration, and beta-catenin translocation. Proc Natl Acad Sci. USA. 2005;102:9182–9187. doi: 10.1073/pnas.0500918102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Haspel J, Grumet M. The L1CAM extracellular region: a multi-domain protein with modular and cooperative binding modes. Front Biosci. 2003;8:s1210–1225. doi: 10.2741/1108. [DOI] [PubMed] [Google Scholar]
- 70.Mylona E, Tzelepis K, Theohari I, Giannopoulou I, Papadimitriou C, Nakopoulou L. Cyclin D1 in invasive breast carcinoma: favourable prognostic significance in unselected patients and within subgroups with an aggressive phenotype. Histopathology. 2013;62:472–480. doi: 10.1111/his.12013. [DOI] [PubMed] [Google Scholar]
- 71.Xu P, Zhao M, Liu Z, Liu Y, Chen Y, Luo R, Fang W. Elevated nuclear CCND1 expression confers an unfavorable prognosis for early stage lung adenocarcinoma patients. Int J Clin Exp Pathol. 2015;8:15887–15894. [PMC free article] [PubMed] [Google Scholar]
- 72.Williams CM, Engler AJ, Slone RD, Galante LL, Schwarzbauer JE. Fibronectin expression modulates mammary epithelial cell proliferation during acinar differentiation. Cancer Res. 2008;68:3185–3192. doi: 10.1158/0008-5472.CAN-07-2673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Xu J, Chen Y, Huo D, Khramtsov A, Khramtsova G, Zhang C, Goss KH, Olopade OI. beta-catenin regulates c-Myc and CDKN1A expression in breast cancer cells. Mol Carcinog. 2016;55:431–439. doi: 10.1002/mc.22292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Ross JS, Ali SM, Wang K, Palmer G, Yelensky R, Lipson D, Miller VA, Zajchowski D, Shawver LK, Stephens PJ. Comprehensive genomic profiling of epithelial ovarian cancer by next generation sequencing-based diagnostic assay reveals new routes to targeted therapies. Gynecol Oncol. 2013;130:554–559. doi: 10.1016/j.ygyno.2013.06.019. [DOI] [PubMed] [Google Scholar]
- 75.Chen Y, McGee J, Chen X, Doman TN, Gong X, Zhang Y, Hamm N, Ma X, Higgs RE, Bhagwat SV, Buchanan S, Peng SB, Staschke KA. Identification of druggable cancer driver genes amplified across TCGA datasets. PLoS One. 2014;9:e98293. doi: 10.1371/journal.pone.0098293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Gavert N, Conacci-Sorrell M, Gast D, Schneider A, Altevogt P, Brabletz T, Ben-Ze’ev A. L1, a novel target of beta-catenin signaling, transforms cells and is expressed at the invasive front of colon cancers. J Cell Biol. 2005;168:633–642. doi: 10.1083/jcb.200408051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Yao ES, Zhang H, Chen YY, Lee B, Chew K, Moore D, Park C. Increased beta1 integrin is associated with decreased survival in invasive breast cancer. Cancer Res. 2007;67:659–664. doi: 10.1158/0008-5472.CAN-06-2768. [DOI] [PubMed] [Google Scholar]
- 78.Gradl D, Kuhl M, Wedlich D. The Wnt/Wg signal transducer beta-catenin controls fibronectin expression. Mol Cell Biol. 1999;19:5576–5587. doi: 10.1128/mcb.19.8.5576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.De Langhe S, Haataja L, Senadheera D, Groffen J, Heisterkamp N. Interaction of the small GTPase Rac3 with NRBP, a protein with a kinase-homology domain. Int J Mol Med. 2002;9:451–459. [PubMed] [Google Scholar]
- 80.Shimizu F, Watanabe TK, Okuno S, Omori Y, Fujiwara T, Takahashi E, Nakamura Y. Isolation of a novel human cDNA (rhoHP1) homologous to rho genes. Biochim Biophys Acta. 1997:13–16. doi: 10.1016/s0167-4781(97)00008-0. 1351. [DOI] [PubMed] [Google Scholar]
- 81.Fransson S, Ruusala A, Aspenstrom P. The atypical Rho GTPases Miro-1 and Miro-2 have essential roles in mitochondrial trafficking. Biochem Biophys Res Commun. 2006;344:500–510. doi: 10.1016/j.bbrc.2006.03.163. [DOI] [PubMed] [Google Scholar]
- 82.Hannak E, Kirkham M, Hyman AA, Oegema K. Aurora-A kinase is required for centrosome maturation in Caenorhabditis elegans. J Cell Biol. 2001;155:1109–1116. doi: 10.1083/jcb.200108051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Zhao ZS, Lim JP, Ng YW, Lim L, Manser E. The GIT-associated kinase PAK targets to the centrosome and regulates Aurora-A. Mol Cell. 2005;20:237–249. doi: 10.1016/j.molcel.2005.08.035. [DOI] [PubMed] [Google Scholar]
- 84.Mi H, Muruganujan A, Casagrande JT, Thomas PD. Large-scale gene function analysis with the PANTHER classification system. Nat Protoc. 2013;8:1551–1566. doi: 10.1038/nprot.2013.092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Ackermann MA, Shriver M, Perry NA, Hu LY, Kontrogianni-Konstantopoulos A. Obscurins: Goliaths and Davids take over non-muscle tissues. PLoS One. 2014;9:e88162. doi: 10.1371/journal.pone.0088162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.McLaren W, Gil L, Hunt SE, Riat HS, Ritchie GR, Thormann A, Flicek P, Cunningham F. The Ensembl Variant Effect Predictor. Genome Biol. 2016;17:122. doi: 10.1186/s13059-016-0974-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Yang H, Wang K. Genomic variant annotation and prioritization with ANNOVAR and wANNOVAR. Nat Protoc. 2015;10:1556–1566. doi: 10.1038/nprot.2015.105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Chun S, Fay JC. Identification of deleterious mutations within three human genomes. Genome Res. 2009;19:1553–1561. doi: 10.1101/gr.092619.109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Schwarz JM, Rodelsperger C, Schuelke M, Seelow D. MutationTaster evaluates disease-causing potential of sequence alterations. Nat Methods. 2010;7:575–576. doi: 10.1038/nmeth0810-575. [DOI] [PubMed] [Google Scholar]
- 90.Reva B, Antipin Y, Sander C. Predicting the functional impact of protein mutations: application to cancer genomics. Nucleic Acids Res. 2011;39:e118. doi: 10.1093/nar/gkr407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Shihab HA, Gough J, Mort M, Cooper DN, Day IN, Gaunt TR. Ranking non-synonymous single nucleotide polymorphisms based on disease concepts. Hum Genomics. 2014;8:11. doi: 10.1186/1479-7364-8-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Carter H, Douville C, Stenson PD, Cooper DN, Karchin R. Identifying Mendelian disease genes with the variant effect scoring tool. BMC Genomics. 2013;14:S3. doi: 10.1186/1471-2164-14-S3-S3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Kircher M, Witten DM, Jain P, O’Roak BJ, Cooper GM, Shendure J. A general framework for estimating the relative pathogenicity of human genetic variants. Nat Genet. 2014;46:310–315. doi: 10.1038/ng.2892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Davydov EV, Goode DL, Sirota M, Cooper GM, Sidow A, Batzoglou S. Identifying a high fraction of the human genome to be under selective constraint using GERP++ PLoS Comput Biol. 2010;6:e1001025. doi: 10.1371/journal.pcbi.1001025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Kumar P, Henikoff S, Ng PC. Predicting the effects of coding non-synonymous variants on protein function using the SIFT algorithm. Nat Protoc. 2009;4:1073–1081. doi: 10.1038/nprot.2009.86. [DOI] [PubMed] [Google Scholar]
- 96.Adzhubei IA, Schmidt S, Peshkin L, Ramensky VE, Gerasimova A, Bork P, Kondrashov AS, Sunyaev SR. A method and server for predicting damaging missense mutations. Nat Methods. 2010;7:248–249. doi: 10.1038/nmeth0410-248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Adzhubei I, Jordan DM, Sunyaev SR. Predicting functional effect of human missense mutations using PolyPhen-2. Curr Protoc Hum Genet. 2013. Chapter 7:Unit7 20. [DOI] [PMC free article] [PubMed]
- 98.Chang X, Wang K. wANNOVAR: annotating genetic variants for personal genomes via the web. J Med Genet. 2012;49:433–436. doi: 10.1136/jmedgenet-2012-100918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Dovas A, Couchman JR. RhoGDI: multiple functions in the regulation of Rho family GTPase activities. Biochem J. 2005;390:1–9. doi: 10.1042/BJ20050104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Qadir MI, Parveen A, Ali M. Cdc42: Role in Cancer Management. Chem Biol Drug Des. 2015;86:432–439. doi: 10.1111/cbdd.12556. [DOI] [PubMed] [Google Scholar]
- 101.Gao C, Xiao G, Hu J. Regulation of Wnt/beta-catenin signaling by posttranslational modifications. Cell Biosci. 2014;4:13. doi: 10.1186/2045-3701-4-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Akhshi TK, Wernike D, Piekny A. Microtubules and actin crosstalk in cell migration and division. Cytoskeleton (Hoboken) 2014;71:1–23. doi: 10.1002/cm.21150. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.