

Abstract
Background
Raccoon dogs and red foxes are well-adapted hosts for Trichinella spp. The aims of this study were to estimate Trichinella infection prevalence and biomass and to investigate which Trichinella species circulated in these indicator hosts in Estonia.
Methods
From material collected for evaluating the effectiveness of oral vaccination program for rabies eradication in wildlife, samples from 113 raccoon dogs and 87 red foxes were included in this study. From each animal, 20 g of masseter muscle tissue was tested for the presence of Trichinella larvae using an artificial digestion method. The Trichinella larvae were identified to species level by multiplex polymerase chain reaction method.
Results
The majority of tested animals were infected with Trichinella spp. The parasite species identified were T. nativa and T. britovi. The apparent infection prevalence was 57.5% in raccoon dogs and 69.0% in red foxes, which were higher than previous estimates. In addition, the larval burden had also increased in both hosts. We estimated that in 2011–2012, the Trichinella spp. biomass was more than 15 times higher in raccoon dogs and almost two times higher in red foxes than in 1992–2000 (based on mean larval burden), and almost 20 times higher in raccoon dogs and almost five times higher in red foxes than in 2000–2002 (based on median larval burden).
Conclusions
Raccoon dogs and red foxes are relevant reservoirs for Trichinella spp. in Estonia. The biomass of Trichinella circulating in sylvatic cycles was substantial and had increased: there is substantial infection pressure in the sylvatic cycle.
Keywords: Trichinella infection, Trichinella nativa, Trichinella britovi, Prevalence, Sylvatic, Zoonosis
Background
Trichinella spp. are zoonotic parasitic nematodes transmitted by carnivorism. Sylvatic Trichinella infections are endemic in Estonia, a EU country located in north-eastern Europe that is bordered by Latvia in the south and Russia in the east [1–5]. For example, while anti-Trichinella antibodies were not detected in the domestic pigs investigated in our recent study, a substantial proportion of hunted wild boars were Trichinella seropositive [5]. Assessment of the sylvatic component and awareness about it are important because there is a risk of spill-over to domestic animals and humans [5].
The raccoon dog (Nyctereutes procyonoides) is a suitable indicator species and well-adapted reservoir host for all four Trichinella species circulating in Europe, and the red fox (Vulpes vulpes) particularly for T. spiralis and T. britovi [4, 6–10]. The invasive raccoon dog [11], and the native red fox are common and numerous sylvatic carnivores in Estonia [12]. A total of 12,577 raccoon dogs and 7144 red foxes were hunted in Estonia during the hunting season 2011–2012 [13].
The most recent epidemiological data on Trichinella infections in raccoon dogs and red foxes in Estonia were based on material collected in 2000–2002 [4]. The apparent prevalence of Trichinella spp. was 42.0% in raccoon dogs and 40.6% in red foxes, which did not differ significantly from estimates from 1992 to 2000 [14]. In both earlier studies, T. nativa and T. britovi were identified in the target hosts. Our study aimed to update the Trichinella infection prevalence estimates in raccoon dogs and red foxes in Estonia and to identify the Trichinella species causing the infections. We compared the findings with the two previous estimates and with those reported from other European countries and estimated how the biomass of Trichinella has changed in Estonia.
Methods
For the evaluation of the effectiveness of the oral vaccination program for rabies eradication in wildlife [15], head samples from 1214 raccoon dogs and 625 red foxes were collected from whole Estonian territory (average density 4.3 animals per 100 km2) from August 2011 to March 2012. The animals sampled were apparently healthy hunted animals, rabies indicator animals killed due to abnormal behavior near human settlements, and animals killed in traffic or found dead. We could investigate muscle samples from 200 of these heads (113 raccoon dogs and 87 red foxes), which was evaluated to be a sufficient sample size to estimate the infection prevalence with 80% confidence level.
To obtain a geographically representative sample for this study, the number of samples from each county was adjusted according to the surface area of the county, and a random sample was drawn from the samples available from there. Data on the estimated age (less than 1 year old = juvenile, at least 1 year old = adult) and gender of each animal had been collected on the submission forms. Age group was unknown for 31 animals and gender for 69 animals.
The samples were kept refrigerated until analysis, but few samples were or could have been frozen (n = 3 from raccoon dogs and n = 2 from foxes arrived frozen). From each animal, 20 g of masseter muscle tissue was analyzed for the presence of Trichinella spp. larvae using the European Union reference method, i.e. magnetic stirrer method, for artificial digestion [16]. The mean time between sampling and digestion was ten days (range: 1–92 days).
Larvae from each positive sample were evaluated morphologically and then counted, rinsed with water, collected, and stored in ethanol at room temperature until identification to species level. The species of Trichinella were identified using a previously described multiplex polymerase chain reaction method [17].
The sample size assessment and preliminary statistical calculations were done with OpenEpi software [18]. The confidence intervals (CI) of the prevalence estimates were calculated using Mid-P exact. Comparisons with the prevalence estimates, by host species and Trichinella species, from other European countries and previous Estonian studies were done using two by two tables. Two-tailed P-values (Mid-P exact) < 0.05 were considered statistically significant.
Logistic regression models were built with STATA 13.0 (Stata Corporation, College Station, Texas, USA) software for three outcomes: testing positive for Trichinella spp., testing positive for T. nativa, and testing positive for T. britovi. The variables we evaluated were ‘host species’ (raccoon dog or red fox), ‘age’ (juvenile or adult), ‘gender’ (female or male), ‘county’ (the 15 counties included as dummy variables i.e. allocated numbers that do not indicate any particular order), and ‘cause of death’ (whether the animal had been hunted, killed due to abnormal behavior, killed in traffic, or found dead). The variables with P-value ≤ 0.25 in univariable analysis were included in a multivariable model, followed by a stepwise backward elimination of those with P ≥ 0.05 that did not act as confounders.
Trichinella spp. biomass was quantitatively estimated for 1000 host animals and for the hunting bag, using estimate of weight of the host, estimate of proportion of muscle of the host weight (based on information available for small mammals of similar size), point estimate of Trichinella spp. prevalence, and mean or median larvae per gram of muscle tissue.
Results
The majority (62.5%, 125/200, 95% CI: 55.6–69.0) of the animals tested were infected with Trichinella spp. (Table 1). The apparent Trichinella spp. infection prevalence was 57.5% (65/113, 95% CI: 48.3–66.2) in raccoon dogs and 69.0% (60/87, 95% CI: 58.6–77.7) in red foxes. The prevalence was not significantly higher in red foxes than in raccoon dogs.
Table 1.
Trichinella species identified in raccoon dogs (Nyctereutes procyonoides) and red foxes (Vulpes vulpes) in 2011–2012 in Estonia
| Trichinella species | Raccoon dog (n = 113) | Red fox (n = 87) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Positive (n) | Prevalence (95% CI)a (%) | % of Trichinella-positive (95% CI) | Range of lpg | Mean (median) lpg | Positive (n) | Prevalence (95% CI)a (%) | % of Trichinella-positive (95% CI) | Range of lpg | Mean (median) lpg | |
| T. nativa only | 23 | 20.4 (13.7–29.0) | 35.4 (24.5–47.5) | 0.5–631.6 | 158.4 (135.0) | 19 | 21.8 (14.1–31.4) | 31.7 (20.9–44.2) | 0.1–636.8 | 82.4 (13.8) |
| T. britovi only | 15 | 13.3 (7.9–20.5) | 23.1(14.1–34.5) | 0.6–486.0 | 123.5 (58.8) | 23 | 26.4 (18.0–36.4) | 38.3 (26.7–51.1) | 0.1–409.5 | 44.7 (10.5) |
| T. nativa and T. britovi | 13 | 11.5 (6.6–18.4) | 20.0 (11.6–31.8) | 26.2–800.0 | 209.2 (98.0) | 8 | 9.2 (4.4–16.7) | 13.3 (6.4–23.8) | 2.3–28.6 | 9.1 (7.1) |
| T. nativa, totalb | 36 | 31.9 (23.8–40.9) | 55.4 (43.2–67.1) | 0.5–800.0 | 176.7 (130.0) | 27 | 31.0 (22.0–41.3) | 45.0 (32.8–57.7) | 0.1–636.8 | 60.7 (8.4) |
| T. britovi, totalb | 28 | 24.8 (17.5–33.4) | 43.1 (31.5–55.3) | 0.6–800.0 | 163.3 (83.2) | 31 | 35.6 (26.1–46.1) | 51.7 (39.1–64.1) | 0.1–409.5 | 8.2 (35.5) |
| Species-level result | 51 | 45.1 (36.1–54.4) | 78.5 (67.3–87.2) | 0.5–800.0 | 161.1 (101.4) | 50 | 57.5 (46.9–67.5) | 83.3 (72.3–91.2) | 0.1–636.8 | 53.3 (9.4) |
| No species-level result | 14 | 12.4 (7.2–19.5) | 21.5 (12.8–32.8) | 0.1–576.0 | 161.1 (43.6) | 10 | 11.5 (6.0–19.5) | 16.7 (8.8–27.7) | 0.1–142.9 | 21.4 (2.4) |
| Total | 65 | 57.5 (48.3–66.4) | 100 (95.5–100) | 0.1–800.0 | 161.1 (98.0) | 60 | 69.0 (58.7–78.0) | 100 (95.1–100) | 0.1–636.8 | 48.0 (8.2) |
Abbreviation: lpg, larvae per gram of muscle tissue
a95% confidence interval, Mid-P exact
bWith this particular Trichinella species, either as the only species or in mixed infection
The Trichinella species present were successfully identified from 80.8% of the animals that had larvae (Table 1). The success rate of Trichinella species identification was 82.7% (91/110, 95% CI: 74.8–89.0) from larvae from samples that were digested within the recommended 21 days after sampling [9], and 66.7% (10/15, 95% CI: 40.8–86.6) from samples stored longer. However, the difference was not significant.
The presence of two sylvatic species, T. nativa and T. britovi, was confirmed (Table 1). Trichinella nativa was detected as the only species present or in mixed infections in 31.9% of raccoon dogs and 31.0% of red foxes, and T. britovi was detected as the only species present or in mixed infections in 24.8% of raccoon dogs and 35.6% of red foxes. Of those animals that hosted Trichinella spp. larvae that were determined to the species level, T. nativa was detected as the only species present or in mixed infection in 70.6% of raccoon dogs and 54.0% of red foxes. Of those animals that hosted Trichinella spp. larvae that were determined to the species level, T. britovi was detected as the only species or in mixed infection in 54.9% of raccoon dogs and 62.0% of red foxes. The prevalence of mixed infections had increased in red foxes from the estimate of the previous Estonian study (Table 3) [4].
Table 3.
Prevalence of Trichinella spp. in red foxes (Vulpes vulpes) in European countries and comparison with the present study
| Country | Sampling period | Samples (n) | Prevalence of Trichinella spp. (95% CI) (%) | Prevalence of Tn (total) (95% CI) (%) | Prevalence of Tb (total) (95% CI) (%) | Prevalence of Ts (total) (95% CI) (%) | Prevalence of Tp (total) (95% CI) (%) | Prevalence of mixed infections (95% CI) (%) | Reference |
|---|---|---|---|---|---|---|---|---|---|
| Estonia | 2011–2012 | 87 | 69.0 (58.7–78.0) | 31.0 (22.0–41.3) | 35.6 (26.1–46.1) | 0.0 (0.0–3.4) | 0.0 (0.0–3.4) | 9.2 (4.4–16.7) | Present study |
| Estonia | 2000–2002 | 446 | 40.6*** (36.1–45.2) | 16.6** (13.4–20.3) | 13.5*** (10.5–16.7) | 0.0 (0.0–0.7) | 0.0 (0.0–0.7) | 2.9*(1.6–4.8) | [4] |
| Estonia | 1992–2000 | 21 | 42.9* (23.3–64.3) | 14.3 (3.8–34.1) | 28.6 (12.5–50.2) | 0.0 (0.0–13.3) | 0.0 (0.0–13.3) | 4.8 (0.2–21.3) | [3, 14, 57] |
| Austria | 1546 | 1.6*** (1.0–2.3) | 0.0*** (0.0–0.2) | 1.6*** (1.0–2.3) | 0.0 (0.0–0.2) | 0.0 (0.0–0.2) | 0.0*** (0.0–0.2) | [27] | |
| Belgium | 1996–2000 | 818 | 0.0*** (0.0–0.4) | 0.0*** (0.0–0.4) | 0.0*** (0.0–0.4) | 0.0 (0.0–0.4) | 0.0 (0.0–0.4) | 0.0*** (0.0–0.4) | [28] |
| Denmark | 1995–1996 | 3133 | 0.1*** (0.0–0.3) | [29] | |||||
| Denmark | 1997–1998 | 3008 | 0.0*** (0.0–0.1) | 0.0*** (0.0–0.1) | 0.0*** (0.0–0.1) | 0.0 (0.0–0.1) | 0.0 (0.0–0.1) | 0.0*** (0.0–0.1) | [29] |
| Finland | 1999–2005 | 1010 | 18.7*** (16.4–21.2) | [19] | |||||
| Finland | 1996–1998 | 158 | 36.7*** (29.5–44.4) | [8] | |||||
| France | 2006–2009 | 108 | 2.8*** (0.7–7.4) | 0.0*** (0.0–2.7) | 2.8*** (0.7–7.4) | 0.0 (0.0–2.7) | 0.0 (0.0–2.7) | 0.0*** (0.0–2.7) | [29] |
| France | 2006–2008 | 74 | 0.0*** (0.0–4.0) | 0.0*** (0.0–4.0) | 0.0*** (0.0–4.0) | 0.0 (0.0–4.0) | 0.0 (0.0–4.0) | 0.0*** (0.0–4.0) | [31] |
| Germany | 2011–2012 | 3154 | 0.3*** (0.2–0.6) | [25] | |||||
| Germany | 2006–2007 | 100 | 1.0*** (0.1–4.8) | 0.0*** (0.0–3.0) | 0.0*** (0.0–3.0) | 0.0 (0.0–3.0) | 0.0 (0.0–3.0) | 0.0*** (0.0–3.0) | [20] |
| Hungary | 2008–2013 | 3304 | 2.1*** (1.6–2.6) | 0.0*** (0.0–0.1) | 1.8*** (1.4–2.3) | 0.2 (0.1–0.5) | 0.0 (0.0–0.1) | 0.0*** (0.0–0.1) | [32] |
| Hungary | 2007–2008 | 2116 | 1.7*** (1.2–2.3) | 0.0*** (0.0–0.1) | 1.4*** (1.0–2.0) | 0.2 (0.1–0.5) | 0.0 (0.0–0.2) | 0.0*** (0.0–0.1) | [33] |
| Italy | 2010–2014 | 153 | 8.5*** (4.8–13.8) | 0.0*** (0.0–1.9) | 8.5*** (4.8–13.8) | 0.0 (0.0–1.9) | 0.0 (0.0–1.9) | 0.0*** (0.0–1.9) | [34] |
| Italy | 2004–2014 | 480 | 5.0*** (3.3–7.2) | [35] | |||||
| Italy | 2001–2004 | 229 | 3.1*** (1.3–6.0) | 0.0*** (0.0–1.3) | 3.1*** (1.3–6.0) | 0.0 (0.0–1.3) | 0.0 (0.0–1.3) | 0.0*** (0.0–1.3) | [36] |
| Italy | 2001–2004 | 227 | 3.5*** (1.7–6.6) | 0.0*** (0.0–1.3) | 3.5*** (1.7–6.6) | 0.0 (0.0–1.3) | 0.0 (0.0–1.3) | 0.0*** (0.0–1.3) | [37] |
| Italy | 1997–2003 | 172 | 1.2*** (0.2–3.8) | [38] | |||||
| Ireland | 2002 | 454 | 0.9*** (0.3–2.1) | 0.0*** (0.0–0.7) | 0.0*** (0.0–0.7) | 0.9 (0.3–2.1) | 0.0 (0.0–0.7) | 0.0*** (0.0–0.7) | [39] |
| Latvia | 2010–2014 | 668 | 50.6** (46.8–54.4) | 1.5*** (0.8–2.7) | 40.9 (37.2–44.6) | 0.0 (0.0–0.4) | 0.0 (0.0–0.4) | 1.0*** (0.5–2.1) | [22] |
| Latvia | 2000–2002 | 1112 | 28.9*** (26.3–31.6) | 2.2*** (1.4–3.1) | 10.3*** (8.6–12.1) | 0.4 (0.2–1.0) | 0.0 (0.0–0.3) | 1.3*** (0.7–2.1) | [4] |
| Lithuania | 2001–2006 | 206 | 46.6*** (39.9–53.4) | [23] | |||||
| Lithuania | 2000–2002 | 567 | 40.0*** (36.1–44.1) | 0.9*** (0.3–1.9) | 23.3* (19.9–26.9) | 4.8* (3.2–6.8) | 0.2 (0.0–0.9) | 3.2* (2.0–4.9) | [4] |
| Netherlands | 2010–2013 | 369 | 0.3*** (0.0–1.3) | [40] | |||||
| Netherlands | 1996–1997 | 276 | 4.0*** (2.1–6.8) | 0.0*** (0.0–1.1) | 4.0*** (2.1–6.8) | 0.0 (0.0–1.1) | 0.0 (0.0–1.1) | 0.0*** (0.0–1.1) | [41] |
| Norway | 1994–1995, 2002–2005 | 393 | 4.8*** (3.0–7.3) | 4.6*** (2.8–7.0) | 0.3*** (0.0–1.2) | 0.0 (0.0–0.8) | 0.0 (0.0–0.8) | 0.0*** (0.0–0.8) | [42] |
| Poland | 2010–2015 | 1447 | 10.0*** (8.6–11.7) | 0.0*** (0.0–0.2) | 7.2*** (5.9–8.6) | 1.1 (0.7–1.8) | 0.1 (0.0–0.3) | 0.0*** (0.0–0.2) | [26] |
| Poland | 2011–2012 | 1634 | 2.7*** (2.0–3.6) | 0.1*** (0.0–0.3) | 2.0*** (1.4–2.7) | 0.6 (0.3–1.0) | 0.0 (0.0–0.2) | 0.1*** (0.0–0.3) | [25] |
| Portugal | 2008–2010 | 47 | 2.1*** (0.1–10.1) | 0.0*** (0.0–6.2) | 2.1*** (0.1–10.1) | 0.0 (0.0–6.2) | 0.0 (0.0–6.2) | 0.0*** (0.0–6.2) | [43] |
| Romania | 2012–2014 | 121 | 21.5*** (14.9–29.5) | 0.0*** (0.0–2.4) | 19.8* (13.5–27.7) | 0.8 (0.0–4.0) | 0.0 (0.0–2.4) | 0.0*** (0.0–2.4) | [44] |
| Romania | 2000–2005 | 71 | 7.0*** (2.6–14.9) | 0.0*** (0.0–4.1) | 5.6*** (1.8–13.0) | 1.4 (0.1–6.7) | 0.0 (0.0–4.1) | 0.0** (0.0–4.1) | [45] |
| Serbia | 2009–2010 | 57 | 12.3*** (5.5–22.8) | 0.0*** (0.0–5.1) | 3.5*** (0.6–11.1) | 8.8** (3.3–18.4) | 0.0 (0.0–5.1) | 3.5 (0.6–11.1) | [46] |
| Slovakia | 2000–2007 | 5270 | 11.5*** (10.7–12.4) | [47] | |||||
| Slovakia | 2007 | 601 | 20.3*** (17.2–23.7) | 0.0*** (0.0–0.5) | 20.3*** (17.2–23.7) | 0.3 (0.1–1.1) | 0.2 (0.0–0.8) | 0.5*** (0.1–1.4) | [47] |
| Slovakia | 2000–2006 | 4669 | 10.4*** (9.5–11.3) | 0.0*** (0.0–0.1) | 8.3*** (7.6–9.2) | 0.1 (0.0–0.2) | 0.0 (0.0–0.1) | 0.0*** (0.0–0.1) | [48] |
| Slovakia | 2000 | 545 | 6.1*** (4.3–8.3) | 0.0*** (0.0–0.5) | 6.1*** (4.3–8.3) | 0.0 (0.0–0.5) | 0.0 (0.0–0.5) | 0.0*** (0.0–0.5) | [47] |
| Spain | – | 400 | 15.5*** (12.2–19.3) | 15.3*** (12.0–19.0) | [49] | ||||
| Spain | 1997–1999 | 67 | 8.9*** (3.7–17.7) | [50] | |||||
| Spain | 1985–1997 | 227 | 2.6*** (1.1–5.4) | 0.0*** (0.0–1.3) | 1.8*** (0.6–4.2) | 0.9 (0.1–2.9) | 0.0 (0.0–1.3) | 0.0*** (0.0–1.3) | [51] |
| Spain | 1989–1993 | 84 | 1.2*** (0.1–5.7) | [52] | |||||
| Sweden | 1985–2003 | 1800 | 4.5*** (3.6–5.5) | [53] | |||||
| Switzerland | 2006–2007 | 1289 | 1.6*** (1.0–2.4) | 0.0*** (0.0–0.2) | 1.6*** (1.0–2.4) | 0.0 (0.0–0.2) | 0.0 (0.0–0.2) | 0.0*** (0.0–0.2) | [54] |
| United Kingdom (Great Britain) | 2003–2007 | 1144 | 0.0*** (0.0–0.3) | 0.0*** (0.0–0.3) | 0.0*** (0.0–0.3) | 0.0 (0.0–0.3) | 0.0 (0.0–0.3) | 0.0*** (0.0–0.3) | [55] |
| United Kingdom (Northern Ireland) | 2003–2004; 2006–2007 | 443 | 0.2*** (0.0–1.1) | 0.0*** (0.0–0.7) | 0.0*** (0.0–0.7) | 0.2 (0.0–1.1) | 0.0 (0.0–0.7) | 0.0*** (0.0–0.7) | [56] |
Abbreviations: Tn, Trichinella nativa; Tb, Trichinella britovi; Ts, Trichinella spiralis; Tp, Trichinella pseudospiralis
*P < 0.05, **P < 0.01, ***P < 0.001
The apparent Trichinella spp. infection prevalences estimated from the samples from 2011 to 2012 were higher than those from 2000 to 2002 in both raccoon dogs and red foxes (Tables 2 and 3) [4]. Moreover, the prevalence estimated from samples from 2011 to 2012 was higher than the one from 1992 to 2000 in red foxes; however, the estimate from 2011 to 2012 did not differ significantly from the estimate from 1992 to 2000 in raccoon dogs (Tables 2 and 3) [14].
Table 2.
Prevalence of Trichinella spp. in raccoon dogs (Nyctereutes procyonoides) in European countries and comparison with the present study
| Country | Sampling period | Samples (n) | Prevalence of Trichinella spp. (95% CI) (%) | Prevalence of Tn (total) (95% CI) (%) | Prevalence of Tb (total) (95% CI) (%) | Prevalence of Ts (total) (95% CI) (%) | Prevalence of Tp (total) (95% CI) (%) | Prevalence of mixed infections (95% CI) (%) | Reference |
|---|---|---|---|---|---|---|---|---|---|
| Estonia | 2011–2012 | 113 | 57.5 (48.3–66.4) | 31.9 (23.8–40.9) | 24.8 (17.5–33.4) | 0.0 (0.0–2.6) | 0.0 (0.0–2.6) | 11.5 (7.9–20.5) | Present study |
| Estonia | 2000–2002 | 157 | 42.0* (34.5–49.9) | 15.9** (10.8–22.3) | 15.9 (10.8–22.3) | 0.0 (0.0–5.1) | 0.0 (0.0–5.1) | 5.1 (2.4–9.4) | [4] |
| Estonia | 1992–2000 | 33 | 45.5 (29.2–62.5) | 15.2 (5.8–30.4) | 27.3 (14.2–44.2) | 0.0 (0.0–8.7) | 0.0 (0.2–8.7) | 3.0 (0.2–14.1) | [3, 14, 57] |
| Finland | 1999–2005 | 662 | 28.1*** (24.8–31.6) | [19] | |||||
| Finland | 1996–1998 | 199 | 37.7*** (31.2–44.6) | [8] | |||||
| Germany | 2006–2007 | 146 | 4.8*** (2.1–9.3) | 0.0*** (0.0–2.0) | 0.0*** (0.0–2.0) | 4.1* (1.7–8.3) | 2.7 (0.9–6.5) | 1.4*** (0.2–4.5) | [20] |
| Germany | 2000–2014 | 1527 | 1.9*** (1.3–2.7) | 0.0*** (0.0–0.2) | 0.1*** (0.0–0.3) | 1.7 (1.1–2.4) | 0.1 (0.0–0.3) | 0.0 (0.0–0.2)*** | [21] |
| Latvia | 2010–2014 | 394 | 37.3*** (32.6–42.2) | 2.8*** (1.5–4.8) | 35.0* (30.4–39.9) | 0.5 (0.1–1.7) | 0.0 (0.0–0.8) | 3.0*** (1.7–5.1) | [22] |
| Latvia | 2000–2002 | 17 | 35.3 (15.7–59.5) | [4] | |||||
| Lithuania | 2001–2006 | 75 | 29.3*** (19.9–40.4) | [23] | |||||
| Lithuania | 2000–2002 | 83 | 32.5*** (23.1–43.1) | 2.4*** (0.4–7.7) | 25.3 (16.8–35.5) | 4.8* (1.6–11.2) | 0.0 (0.0–3.5) | 4.8 (1.6–11.2) | [4] |
| Poland | 2012 | 39 | 5.1*** (0.9–15.9) | 0.0*** (0.0–7.4) | 0.0*** (0.0–7.4) | 5.1 (0.9–15.9) | 0.0 (0.0–7.4) | 0.0* (0.0–7.4) | [24] |
Abbreviations: Tn, Trichinella nativa; Tb, Trichinella britovi; Ts, Trichinella spiralis; Tp, Trichinella pseudospiralis
*P < 0.05, **P < 0.01, ***P < 0.001
The number of Trichinella spp. larvae recovered per gram muscle tissue (lpg) was higher in raccoon dogs (median: 98.0, mean: 161.1, range: 0.1–800.0 lpg) than in red foxes (median: 8.2, mean: 48.0, range: 0.1–636.8 lpg), and varied by Trichinella species (Table 1). The highest larval burden, 800 lpg, was detected in a raccoon dog with mixed infection.
The median larval burden had increased in both raccoon dogs and red foxes from those reported in the previous study: from 7.2 lpg to 98.0 lpg in raccoon dogs and from 3.0 lpg to 8.2 lpg in red foxes [4]. Furthermore, the proportion of animals with low larval burden (< 1 lpg) had decreased from 18.1% to 7.7% in raccoon dogs, and from 23.7% to 11.7% in red foxes, further indicating that the circulating parasite biomass of Trichinella larvae had increased [4]. The Trichinella spp. biomass was estimated to have increased 18.6-fold in raccoon dogs and 4.6-fold in red foxes (based on median larval burden) (Table 4).
Table 4.
Calculation of the change in Trichinella spp. biomass in raccoon dogs and red foxes in Estonia
| Raccoon dog | Red fox | Reference | |
|---|---|---|---|
| Hunting bag 1995, n animals | 1723 | 3326 | [67] |
| Hunting bag 2001, n animals | 4259 | 6628 | [67] |
| Hunting bag 2011, n animals | 12,577 | 7144 | [13] |
| With Trichinella larvae (%) | |||
| 1992–2000 | 45.5 | 42.9 | [14] |
| 2000–2002 | 42.0 | 40.6 | [4] |
| 2011–2012 | 57.5 | 69.0 | Present study |
| Mean body weight of host, g | 4830 | 4890 | [68, 69] |
| Muscle tissue of body weight, % | 60 | 60 | [70] |
| Median (mean) Trichinella lpg | |||
| 1992–1996 | nd (13.4) | nd (43.1) | [2] |
| 2000–2002 | 7.2 (nd) | 3.0 (nd) | [4] |
| 2011–2012 | 98.0 (161.1) | 8.2 (48.0) | Current study |
| Trichinella biomass 1992–2000, median (mean) n larvae | |||
| in 1000 animals | nd (17,669,106) | nd (54,249,367) | |
| in the hunting baga | nd (30,443,870) | nd (180,433,393) | |
| Trichinella biomass 2000–2002, median (mean) n larvae | |||
| in 1000 animals | 8,763,552 (nd) | 3,573,612 (nd) | |
| in the hunting bagb | 37,323,968 (nd) | 23,685,900 (nd) | |
| Trichinella biomass 2011–2012, median (mean) n larvae | |||
| in 1000 animals | 163,302,300 (268,448,985) | 16,600,572 (97,174,080) | |
| in the hunting bagc | 2,053,853,027 (3,376,282,884) | 118,594,486 (694,211,628) | |
| Increase in Trichinella biomass from 1992 to 2000 to 2011–2012, calculated from median (mean) n larvae | |||
| in 1000 animals | nd (15.2-fold) | nd (1.8-fold) | |
| in the hunting bag | nd (110.9-fold) | nd (3.8-fold) | |
| Increase in Trichinella biomass from 2000 to 2002 to 2011–2012, calculated from median (mean) n larvae | |||
| in 1000 animals | 18.6-fold (nd) | 4.6-fold (nd) | |
| in the hunting bag | 55.0-fold (nd) | 5.0-fold (nd) | |
Abbreviations: nd, no data; lpg, larvae per gram of muscle tissue
a n larvae = n animals 1995 × % with larvae 1992–2000 × (mean body weight of host, g × muscle tissue of body weight, %) × median (mean) lpg (1992–1996)
b n larvae = n animals 2001 × % with larvae 2000–2002 × (mean body weight of host, g × muscle tissue of body weight, %) × median (mean) lpg (2000–2002)
c n larvae = n animals 2011 × % with larvae 2011–2012 × (mean body weight of host, g × muscle tissue of body weight, %) × median (mean) lpg (2011–2012)
Trichinella nativa was not detected in samples from the large islands Saaremaa and Hiiumaa, nor the most southeastern county Võrumaa, while T. britovi was found in samples collected from all counties (Fig. 1).
Fig. 1.
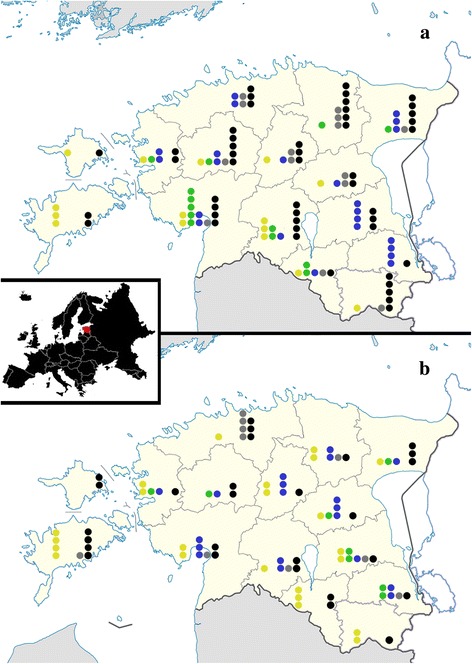
Trichinella spp. in raccoon dogs (Nyctereutes procyonoides) (a) and red foxes (Vulpes vulpes) (b) in 2011–2012 in Estonia, by counties. Key: yellow dot, T. britovi; green dot, T. britovi + T. nativa; blue dot, T. nativa; grey dot, Trichinella spp. (no species-level result); black dot, no larvae detected
None of the variables were significant factors for testing positive for Trichinella spp. in either of the hosts nor in both hosts together. The final model for testing positive for T. nativa had two variables, ‘age’ and ‘county’, and the area under the receiver operating characteristic (ROC) curve was 0.72. The odds of testing positive for T. nativa were 3.6 times (P = 0.009, 95% CI: 1.4–9.3) higher in adults than in juveniles, and higher in counties Põlvamaa and Pärnumaa when compared with Harjumaa where the capital is located (P = 0.009, OR = 15.2, 95% CI: 2.0–117.3, and P = 0.029, OR = 7.6, 95% CI: 1.2–47.1, respectively). The final model for testing T. britovi positive included only the variable ‘county’, and the area under the ROC curve was 0.68. The odds of an animal testing T. britovi positive were higher in the counties Valgamaa, Saaremaa, Läänemaa, and Pärnumaa (P = 0.019, OR = 16.8, 95% CI: 1.6–176.2; P = 0.023, OR = 14.0, 95% CI: 1.4–137.3; P = 0.040, OR =11.7, 95% CI: 1.1–122.4; and P = 0.043, OR = 9.7, 95% CI: 1.1–87.4, respectively) than in the reference county Harjumaa.
Discussion
We summarized the results of European studies on Trichinella spp. infection prevalence in raccoon dogs (Table 2) and red foxes (Table 3). Lower prevalences than our estimate from Estonia have been observed in both hosts in Finland, Latvia, Lithuania, Poland and Germany (Tables 2 and 3) [19–26]. Moreover, in red foxes, the Trichinella spp. infection prevalence was higher in Estonia than what has been reported in Austria, Belgium, Denmark, France, Great Britain, Hungary, Italy, Ireland, Netherlands, Norway, Northern Ireland, Portugal, Romania, Serbia, Slovakia, Spain and Switzerland (Table 3) [26–56]. However, as different sampling schemes, sample sizes, sample material, and detection methods were used, these studies are not all directly comparable with our study.
In Europe, according to the International Trichinella Reference Centre [57], the northern species T. nativa has been found in raccoon dogs in Estonia, Finland, Latvia, Russia, and Sweden; and in red foxes in Estonia, Finland, Germany, Latvia, Norway, Poland, Sweden and Ukraine. The published studies on T. nativa in raccoon dogs and red foxes report lower prevalences (single and mixed infections included) in Latvia, Lithuania and Norway than our estimate from Estonia [4, 22, 42]. In Poland and Germany, T. nativa has been found in red foxes (Table 3) [20, 21, 24–26]. When comparing the result of our study with that from the previous Estonian study, the T. nativa infection prevalence had increased in both raccoon dogs and red foxes (Tables 2 and 3) [4].
In Europe, T. britovi has been found in raccoon dogs in Estonia, Finland, Germany, Latvia and Lithuania (Table 2) [4, 57]. It is the most common Trichinella species in red foxes in Europe [10]. The prevalence of T. britovi we observed in raccoon dogs in single and mixed infections was similar to that reported from Lithuania, lower than that from Latvia, and higher than those from western Poland and Germany (Table 2) [4, 21, 22, 24]. The prevalence of T. britovi we observed in red foxes, including both single and mixed infections, was higher than those reported from Austria, France, Hungary, Norway, Poland, Portugal, Romania, Serbia, Slovakia and Switzerland (Table 3) [21–24, 32, 33, 45–48, 50, 54–56]. A similar to our prevalence estimate for T. britovi was detected in red foxes in Latvia [22]. When comparing the result of our study with that from the previous Estonian study, the T. britovi infection prevalence had increased in red foxes (Table 3) [4]. Moreover, mixed infections were more common in our study than what was observed in raccoon dogs and red foxes in the neighboring country Latvia and in red foxes in Lithuania (Tables 2 and 3) [4, 22].
In this study, the odds of being Trichinella-infected were not significantly different in raccoon dogs and red foxes, whereas the mean larval burden was 3.2 times higher in raccoon dogs than in red foxes. In Latvia, red foxes had higher odds to test positive (P = 0.010, OR = 1.41, 95% CI: 1.08–1.83) than raccoon dogs, but raccoon dogs had 2.9 times higher mean larval burden than red foxes [22]. In Finland, both indicators were higher in raccoon dogs than in red foxes (P < 0.001, OR = 1.70, 95% CI: 1.35–2.14; 3.8 times higher mean larval burden) [19]. A higher larval burden in raccoon dogs than in red foxes has also been described in other studies [4, 8].
Despite the fact that we used 20 g of tissue for the digestion, our study likely underestimated the actual infection prevalence and larval burdens, because the available material was not optimal for finding Trichinella larvae [9, 58, 59]. In experimentally infected raccoon dogs, the T. nativa larval density in masseter muscle was about half of that in foreleg muscles [58]. The storage conditions and transport time could also affect the results [29, 40, 60].
The high Trichinella infection prevalence in raccoon dogs and red foxes, as well as the overall circulation of the parasites in the sylvatic cycle, may be supported by human behavior. For example, the local hunters use carcasses of hunted raccoon dogs as baits [61], which might help the transmission. According to winter tracking index and hunters’ estimations, after the rabies vaccination program started in 2005 [62], the red fox population size first increased, with a peak in 2009–2010, and then decreased [12]. The raccoon dog population size has increased since the second half of last century [62] and has relatively stabilized after 2011–2012 [12]. These changes are also reflected in the increased hunting bag sizes [13] and may have relevance beyond simply higher numbers. There was an association between the abundance index of raccoon dogs and the proportion of Trichinella-infected raccoon dogs and red foxes in Finland [19].
Estonia is located in the transition zone of maritime and continental climate [63]. The coldest months with mean air temperature below zero are December to February [64]. According to data covering these three months from six weather stations located in Harjumaa, Lääne-Virumaa, Pärnumaa, Saaremaa, Tartumaa and Võrumaa, the mean number of days with snow cover was 12.7% (from 4% in Pärnumaa to 26% in Saaremaa) higher in 2002–2011 than in 1992–2001 (data received on request from Estonian Environment Agency). The snow cover could reduce the destructive effect of freezing-thawing cycles on carcasses of infected animals and thus facilitate survival of Trichinella larvae [65, 66].
Raccoon dogs and red foxes act as reservoir hosts for Trichinella spp. in the sylvatic cycle, where the infection can spread to game animals, such as wild boars, that are hunted for human consumption. The Trichinella seroprevalence in wild boars is high in Estonia [5], and the odds of testing Trichinella-seropositive were higher if the wild boar was hunted in certain counties, including Pärnumaa and Saaremaa, when compared with Harjumaa. In this study, a similar comparison was made, with Harjumaa as the reference county. Raccoon dogs and red foxes had higher odds to test T. nativa positive in Pärnumaa, whereas the odds to test T. britovi positive were higher in Pärnumaa and Saaremaa. Moreover, the highest larval burden was detected in a young raccoon dog from Pärnumaa. This raccoon dog had a mixed infection. These two counties could thus be interesting for further studies.
We estimated that in 2011–2012, the Trichinella spp. biomass was more than 15 times higher in raccoon dogs and almost two times higher in red foxes than in 1992–2000 (based on mean larval burden), and almost 20 times higher in raccoon dogs and almost five times higher in red foxes than in 2000–2002 (based on median larval burden) (Table 4). Using the increased hunting bag in the calculation as an indication of increased population size or as an indication of biomass removed from the circulation by hunting, the role of these hosts as reservoirs was clearly illustrated (Table 4). The widespread distribution of Trichinella infections in Estonian wildlife underlines that there is a high infection pressure within the eastern European sylvatic cycles. Moreover, the results of this study indicate that there is an increase in the infection pressure. Trichinella spp. thrive in Estonia, and there is a continuous risk of spill-over to domestic animals and humans.
Conclusions
In Estonia, the proportion of both raccoon dogs and red foxes that hosted Trichinella were higher than ten years earlier. In addition, the larval burdens had also increased in these hosts, and an increased biomass of Trichinella larvae was circulating in sylvatic cycles. Trichinella nativa and T. britovi were found in both host species. There is a substantial and increasing Trichinella infection pressure to the food chains and humans.
Acknowledgements
We thank the hunters and Estonian Hunters Society as well as veterinarians, Veterinary and Food Laboratory, Veterinary and Food Board, Estonian Environment Agency, and European Union Reference Laboratory for their contributions to this study.
Funding
The work was partly supported by project funding M14143VLVP from the Strategic Development Fund of the Estonian University of Life Sciences and by project funding 8P160014VLVP from Base Financing of Estonian University of Life Sciences.
Availability of data and materials
All data used and analyzed during the current study are included in this article.
Abbreviations
- CI
confidence interval
- nd
no data
- OR
odds ratio
- ROC curve
receiver operating characteristic (ROC) curve
- Tb
Trichinella britovi
- Tn
Trichinella nativa
- Tp
Trichinella pseudospiralis
- Ts
Trichinella spiralis
Authors’ contributions
AK presented the idea and designed the study. EN designed and organized the sampling, and KP was responsible for the sampling at necropsy. LH performed the artificial digestion analyses, and AV performed the molecular analyses. AK, PJ and BL analyzed the data and drafted the manuscript. All authors contributed to the writing, read and approved the final manuscript.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Age Kärssin, Email: age.karssin@vetlab.ee.
Liidia Häkkinen, Email: liidia.hakkinen@vetlab.ee.
Enel Niin, Email: enel.niin@vet.agri.ee.
Katrin Peik, Email: katrin.peik@vetlab.ee.
Annika Vilem, Email: annika.vilem@vetlab.ee.
Pikka Jokelainen, Email: pikka.jokelainen@helsinki.fi.
Brian Lassen, Email: brian.lassen@gmail.com.
References
- 1.Kapel CMO. Trichinella in arctic, subarctic and temperate regions: Greenland, the Scandinavian countries and the Baltic states. Southeast Asian J Trop Med Public Health. 1997;28(Suppl.):S14–S19. [PubMed]
- 2.Pozio E, Miller I, Järvis T, Kapel CMO, La Rosa G. Distribution of sylvatic species of Trichinella in Estonia according to climate zones. J Parasitol. 1998;84:193–195. doi: 10.2307/3284561. [DOI] [PubMed] [Google Scholar]
- 3.Järvis T, Miller I, Pozio E. Epidemiological studies on animal and human trichinellosis in Estonia. Parasite. 2001;8:S86–S87. doi: 10.1051/parasite/200108s2086. [DOI] [PubMed] [Google Scholar]
- 4.Malakauskas A, Paulauskas V, Järvis T, Keidans P, Eddi C, Kapel CMO. Molecular epidemiology of Trichinella spp. in three Baltic countries: Lithuania, Latvia, and Estonia. Parasitol Res. 2007;100(4):687–693. doi: 10.1007/s00436-006-0320-y. [DOI] [PubMed] [Google Scholar]
- 5.Kärssin A, Velström K, Gómez-Morales MA, Saar T, Jokelainen P, Lassen B. Cross-sectional study of anti-Trichinella antibody prevalence in domestic pigs and hunted wild boars in Estonia. Vector-Borne Zoonotic Dis. 2016;16(9):604–610. doi: 10.1089/vbz.2016.1943. [DOI] [PubMed] [Google Scholar]
- 6.Pozio E. Trichinellosis in the European Union: epidemiology, ecology and economic impact. Parasitol Today. 1998;14(1):35–38. doi: 10.1016/S0169-4758(97)01165-4. [DOI] [PubMed] [Google Scholar]
- 7.Kapel CMO. Host diversity and biological characteristics of the Trichinella genotypes and their effect on transmission. Vet Parasitol. 2000;93(3–4):263–278. doi: 10.1016/S0304-4017(00)00345-9. [DOI] [PubMed] [Google Scholar]
- 8.Oivanen L, Kapel CMO, Pozio E, La Rosa G, Mikkonen T, Sukura A. Associations between Trichinella species and host species in Finland. J Parasitol. 2002;88(1):84. doi: 10.1645/0022-3395(2002)088[0084:ABTSAH]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- 9.Pozio E, Rossi P. Guidelines for the identification and development of sampling methods and design of suitable protocols for monitoring of Trichinella infection in indicator species. Ann Ist Super di Sanita. 2008;44(2):200–204. [PubMed] [Google Scholar]
- 10.Pozio E, Rinaldi L, Marucci G, Musella V, Galati F, Cringoli G, et al. Hosts and habitats of Trichinella spiralis and Trichinella britovi in Europe. Int J Parasitol. 2009;39(1):71–79. doi: 10.1016/j.ijpara.2008.06.006. [DOI] [PubMed] [Google Scholar]
- 11.Lavrov NP. Results of raccoon dog introductions in different parts of the soviet union. Trudy kafedry biologii MGZPI. 1971;29:101–160. [Google Scholar]
- 12.Veeroja R, Männil P. Status of game populations in Estonia and proposal for hunting in 2016. Estonian Environment Agency. http://www.keskkonnaagentuur.ee/sites/default/files/seirearuanne_2016.pdf. Accessed 19.10.2016.
- 13.Estonian Environment Agency. Jahiulukite küttimine Eestis 2011/2012 aasta jahihooajal. http://www.keskkonnaagentuur.ee/failid/Jahiulukite_kyttimine_2011_12.pdf. Accessed 19.10.2016.
- 14.Järvis T, Miller I. Trichinella infection in animals and humans in Estonia. Agraarteadus. J Agric Sci. 2002;5:266–270. [Google Scholar]
- 15.Cliquet F, Robardet E, Must K, Laine M, Peik K, Picard-Meyer E, et al. eliminating rabies in Estonia. PLoS Negl Trop Dis. 2012;6(2):e1535. doi: 10.1371/journal.pntd.0001535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.European Comission Commission regulation (EC) no 2075/2005 of 5 December 2005 laying down specific rules on official controls for Trichinella in meat. OJEC. 2005;L338:60–82. [Google Scholar]
- 17.European Union Reference Laboratory for Parasites. Istituto Superiore di Sanita. Identification of Trichinella muscle stage larvae at the species level by multiplex PCR. http://www.iss.it/binary/crlp/cont/MI_02_WEB_SITE.pdf. Accessed 07 Dec 2017.
- 18.Dean A, Sullivan K, Soe M. OpenEpi: Open Source Epidemiological Statistics for Public Health, Version 3.01. http://www.openepi.com. Accessed 15.12.2016.
- 19.Airas N, Saari S, Mikkonen T, Virtala A-M, Pellikka J, Oksanen A, et al. Sylvatic Trichinella spp. infection in Finland. J Parasitol. 2010;96(1):67–76. doi: 10.1645/GE-2202.1. [DOI] [PubMed] [Google Scholar]
- 20.Pannwitz G, Mayer-Scholl A, Balicka-Ramisz A, Nöckler K. Increased prevalence of Trichinella spp., northeastern Germany, 2008. Emerg Infect Dis. 2010;16(6):936–942. doi: 10.3201/eid1606.091629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Mayer-Scholl A, Reckinger S, Schulze C, Nöckler K. Study on the occurrence of Trichinella spp. in raccoon dogs in Brandenburg, Germany. Vet Parasitol. 2016;231:102–105. doi: 10.1016/j.vetpar.2016.04.027. [DOI] [PubMed] [Google Scholar]
- 22.Deksne G, Seglina Z, Jahundoviča I, Esite Z, Bakasejevs E, Bagrade G, et al. High prevalence of Trichinella spp. in sylvatic carnivore mammals of Latvia. Vet Parasitol. 2016;231:118–123. doi: 10.1016/j.vetpar.2016.04.012. [DOI] [PubMed] [Google Scholar]
- 23.Bružinskaitė-Schmidhalter R, Šarkūnas M, Malakauskas A, Mathis A, Torgerson PR, Deplazes P. Helminths of red foxes (Vulpes vulpes) and raccoon dogs (Nyctereutes procyonoides) in Lithuania. Parasitology. 2012;139(1):120–127. doi: 10.1017/S0031182011001715. [DOI] [PubMed] [Google Scholar]
- 24.Osten-Sacken N, Solarczyk P. Trichinella spiralis in road-killed raccoon dogs (Nyctereutes procyonoides) in western Poland. Ann Parasitol. 2016;62(1):77–79. doi: 10.17420/ap6201.36. [DOI] [PubMed] [Google Scholar]
- 25.Chmurzyńska E, Rózycki M, Bilska-Zajac E, Nöckler K, Mayer-Scholl A, Pozio E, et al. Trichinella nativa in red foxes (Vulpes vulpes) of Germany and Poland: possible different origins. Vet Parasitol. 2013;198(1–2):254–257. doi: 10.1016/j.vetpar.2013.07.034. [DOI] [PubMed] [Google Scholar]
- 26.Cybulska A, Kornacka A, Bień J, Goździk K, Kalisińska E, Lanocha-Arendarczyk N, et al. The occurrence of Trichinella spp. in red foxes (Vulpes vulpes) in different regions of Poland: current data. Vector-Borne Zoonotic Dis. 2016;16(11):717–721. doi: 10.1089/vbz.2016.1996. [DOI] [PubMed] [Google Scholar]
- 27.Krois E, Nöckler K, Duscher G, Joachim A, Kapel CMO, Prosl H. Trichinella britovi in Austrian red foxes (Vulpes vulpes). Wien Tierarztl Monatsschr 2005;92:308–314. (In German).
- 28.Vercammen F, Vervaeke M, Dorny P, Brandt J, Brochier B, Geerts S, Verhagen R. Survey for Trichinella spp. in red foxes (Vulpes vulpes) in Belgium. Vet Parasitol. 2002;103(1–2):83–88. doi: 10.1016/S0304-4017(01)00579-9. [DOI] [PubMed] [Google Scholar]
- 29.Enemark HL, Bjørn H, Henriksen SA, Nielsen B. Screening for infection of Trichinella in red fox (Vulpes vulpes) in Denmark. Vet Parasitol. 2000;88(3–4):229–237. doi: 10.1016/S0304-4017(99)00219-8. [DOI] [PubMed] [Google Scholar]
- 30.Aoun O, Lacour S, Levieuge A, Marié J-L, Vallée I, Davoust B. Screening for Trichinella britovi infection in red fox (Vulpes vulpes) and wild boar (Sus scrofa) in southeastern France. J Wild Dis. 2012;48(1):223–225. doi: 10.7589/0090-3558-48.1.223. [DOI] [PubMed] [Google Scholar]
- 31.Richomme C, Lacour SA, Ducrot C, Gilot-Fromont E, Casabianca F, Maestrini O, et al. Epidemiological survey of trichinellosis in wild boar (Sus scrofa) and fox (Vulpes vulpes) in a French insular region, Corsica. Vet Parasitol. 2010;172(1–2):150–154. doi: 10.1016/j.vetpar.2010.04.026. [DOI] [PubMed] [Google Scholar]
- 32.Tolnai Z, Széll Z, Marucci G, Pozio E, Sréter T. Environmental determinants of the spatial distribution of Trichinella britovi and Trichinella spiralis in Hungary. Vet Parasitol. 2014;204(3–4):426–429. doi: 10.1016/j.vetpar.2014.04.024. [DOI] [PubMed] [Google Scholar]
- 33.Széll Z, Marucci G, Bajmóczy E, Cséplo A, Pozio E, Sréter T. Spatial distribution of Trichinella britovi, T. pseudospiralis and T. spiralis in red foxes (Vulpes vulpes) in Hungary. Vet Parasitol. 2008;156(3–4):210–215. doi: 10.1016/j.vetpar.2008.06.014. [DOI] [PubMed] [Google Scholar]
- 34.Bandino E, Goddi L, Mulas M, Murgia MC, Soddu M, Marucci G, et al. Trichinella britovi from domestic to wild animals of Sardinia, Italy. Vet Parasitol. 2015;212(3–4):262–266. doi: 10.1016/j.vetpar.2015.07.020. [DOI] [PubMed] [Google Scholar]
- 35.Badagliacca P, Di Sabatino D, Salucci S, Romeo G, Cipriani M, Sulli N, et al. The role of the wolf in endemic sylvatic Trichinella britovi infection in the Abruzzi region of central Italy. Vet Parasitol. 2016;231:124–7. [DOI] [PubMed]
- 36.Balestrieri A, Remonti L, Ferrari N, Ferrari A, Lo Valvo T, Robetto S, Orusa R. Sarcoptic mange in wild carnivores and its co-occurrence with parasitic helminths in the western Italian Alps. Eur J Wildl Res. 2006;52(3):196–201.
- 37.Remonti L, Balestrieri A, Domenis L, Banchi C, Lo Valvo T, Robetto S, Orusa R. Red fox (Vulpes vulpes) cannibalistic behaviour and the prevalence of Trichinella britovi in NW Italian alps. Parasitol Res. 2005;97(6):431–435. doi: 10.1007/s00436-005-1481-9. [DOI] [PubMed] [Google Scholar]
- 38.Di Cerbo AR, Manfredi MT, Bregoli M, Milone NF, Cova M. Wild carnivores as source of zoonotic helminths in north-eastern Italy. Helminthologia. 2008;45(1):13–19. doi: 10.2478/s11687-008-0002-7. [DOI] [Google Scholar]
- 39.Rafter P, Marucci G, Brangan P, Pozio E. Rediscovery of Trichinella spiralis in red foxes (Vulpes vulpes) in Ireland after 30 years of oblivion. J Inf Secur. 2005;50(1):61–65. doi: 10.1016/j.jinf.2004.02.004. [DOI] [PubMed] [Google Scholar]
- 40.Franssen F, Deksne G, Esíte Z, Havelaar A, Swart A, Van Der Giessen J. Trend analysis of Trichinella in a red fox population from a low endemic area using a validated artificial digestion and sequential sieving technique. Vet Res. 2014;45(1):120. doi: 10.1186/s13567-014-0120-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Van Der Giessen JWB, Rombout Y, Vand Der Veen A, Pozio E. Diagnosis and epidemiology of Trichinella infections in wildlife in the Netherlands. Parasite. 2001;8:S103–S105. doi: 10.1051/parasite/200108s2103. [DOI] [PubMed] [Google Scholar]
- 42.Davidson RK, Gjerde B, Vikøren T, Lillehaug A, Handeland K. Prevalence of Trichinella larvae and extra-intestinal nematodes in Norwegian red foxes (Vulpes vulpes) Vet Parasitol. 2006;136(3–4):307–316. doi: 10.1016/j.vetpar.2005.11.015. [DOI] [PubMed] [Google Scholar]
- 43.Lopes AP, Vila-Viçosa MJ, Coutinho T, Cardoso L, Gottstein B, Müller N, Cortes HCE. Trichinella britovi in a red fox (Vulpes vulpes) from Portugal. Vet Parasitol. 2015;210(3–4):260–263. doi: 10.1016/j.vetpar.2015.03.025. [DOI] [PubMed] [Google Scholar]
- 44.Imre K, Pozio E, Tonanzi D, Sala C, Ilie MS, Imre M, Morar A. The red fox (Vulpes vulpes) plays a minor role in the epidemiology of the domestic cycle of Trichinella in Romania. Vet Parasitol. 2015;212(3–4):448–450. doi: 10.1016/j.vetpar.2015.06.032. [DOI] [PubMed] [Google Scholar]
- 45.Blaga R, Gherman C, Cozma V, Zocevic A, Pozio E, Boireau P. Trichinella species circulating among wild and domestic animals in Romania. Vet Parasitol. 2009;159(3–4):218–221. doi: 10.1016/j.vetpar.2008.10.034. [DOI] [PubMed] [Google Scholar]
- 46.Zivojinovic M, Sofronic-Milosavljevic L, Cvetkovic J, Pozio E, Interisano M, Plavsic B, et al. Trichinella infections in different host species of an endemic district of Serbia. Vet Parasitol. 2013;194(2–4):136–138. doi: 10.1016/j.vetpar.2013.01.039. [DOI] [PubMed] [Google Scholar]
- 47.Hurníková Z, Dubinský P. Long-term survey on Trichinella prevalence in wildlife of Slovakia. Vet Parasitol. 2009;159(3–4):276–280. doi: 10.1016/j.vetpar.2008.10.056. [DOI] [PubMed] [Google Scholar]
- 48.Miterpáková M, Hurníková Z, Antolová D, Dubinský P. Endoparasites of red fox (Vulpes vulpes) in the Slovak Republic with the emphasis on zoonotic species Echinococcus multilocularis and Trichinella spp. Helminthologia. 2009;46:73–79. doi: 10.2478/s11687-009-0015-x. [DOI] [Google Scholar]
- 49.Lledó L, Giménez-Pardo C, Saz JV, Serrano JL. Wild red foxes (Vulpes vulpes) as sentinels of parasitic diseases in the province of Soria, northern Spain. Vector-Borne Zoonotic Dis. 2015;12:743–749. doi: 10.1089/vbz.2014.1766. [DOI] [PubMed] [Google Scholar]
- 50.Criado-Fornelio A, Gutierrez-Garcia L, Rodriguez-Caabeiro F, Reus-Garcia E, Roldan-Soriano MA, Diaz-Sanchez MAA. Parasitological survey of wild red foxes (Vulpes vulpes) from the province of Guadalajara, Spain. Vet Parasitol. 2000;92(4):245–251. doi: 10.1016/S0304-4017(00)00329-0. [DOI] [PubMed] [Google Scholar]
- 51.Pérez-Martín JE, Serrano F, Reina D, Mora J, Navarette I. Sylvatic trichinellosis in southwestern Spain. J Wildl Dis. 2000;36(3):531–534. doi: 10.7589/0090-3558-36.3.531. [DOI] [PubMed] [Google Scholar]
- 52.Gortázar C, Villafuerte R, Lucientes J, Fernández-de-Luco D. Habitat related differences in helminth parasites of red foxes in the Ebro valley. Vet Parasitol. 1998;80(1):75–81. doi: 10.1016/S0304-4017(98)00192-7. [DOI] [PubMed] [Google Scholar]
- 53.Pozio E, Christensson D, Stéen M, Marucci G, La Rosa G, Bröjer C, et al. Trichinella pseudospiralis foci in Sweden. Vet Parasitol. 2004;125(3–4):335–342. doi: 10.1016/j.vetpar.2004.07.020. [DOI] [PubMed] [Google Scholar]
- 54.Frey CF, Schuppers ME, Müller N, Ryser-Degiorgis MP, Gottstein B. Assessment of the prevalence of Trichinella spp. in red foxes and Eurasian lynxes from Switzerland. Vet Parasitol. 2009;159(3–4):295–299. doi: 10.1016/j.vetpar.2008.10.060. [DOI] [PubMed] [Google Scholar]
- 55.Zimmer IA, Hunter SJ, Morgan CP, Hunt KR, Smith GC, Howell M, Taylor MA. Detection and surveillance for animal trichinellosis in GB. Vet Parasitol. 2008;151(2–4):233–241. doi: 10.1016/j.vetpar.2007.11.001. [DOI] [PubMed] [Google Scholar]
- 56.Zimmer IA, Fee SA, Spratt-Davison S, Hunter SJ, Boughtflower VD, Morgan CP, et al. Report of Trichinella spiralis in a red fox (Vulpes vulpes) in Northern Ireland. Vet Parasitol. 2009;159(3–4):300–303. doi: 10.1016/j.vetpar.2008.10.066. [DOI] [PubMed] [Google Scholar]
- 57.International Trichinella Reference Center. Database of Trichinella strains. https://www.iss.it/site/Trichinella. Accessed 19.10.2016.
- 58.Mikkonen T, Oivanen L, Näreaho A, Helin H, Sukura A. Predilection muscles and physical condition of raccoon dogs (Nyctereutes procyonoides) experimentally infected with Trichinella spiralis and Trichinella nativa. Acta Vet Scand. 2001;42(4):441–452. doi: 10.1186/1751-0147-42-441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kapel CMO, Webster P, Gamble HR. Muscle distribution of sylvatic and domestic Trichinella larvae in production animals and wildlife. Vet Parasitol. 2005;132(1–2):101–105. doi: 10.1016/j.vetpar.2005.05.036. [DOI] [PubMed] [Google Scholar]
- 60.Davidson RK, Handeland K, Kapel CMO. High tolerance to repeated cycles of freezing and thawing in different Trichinella nativa isolates. Parasitol Res. 2008;103(5):1005–1010. doi: 10.1007/s00436-008-1079-0. [DOI] [PubMed] [Google Scholar]
- 61.Süld K, Valdmann H, Laurimaa L, Soe E, Davison J, Saarma U. An invasive vector of zoonotic disease sustained by anthropogenic resources: the raccoon dog in northern Europe. PLoS One. 2014;9(5) [DOI] [PMC free article] [PubMed]
- 62.Niin E, Laine M, Guiot AL, Demerson JM, Cliquet F. Rabies in Estonia: situation before and after the first campaigns of oral vaccination of wildlife with SAG2 vaccine bait. Vaccine. 2008;26(29–30):3556–3565. doi: 10.1016/j.vaccine.2008.04.056. [DOI] [PubMed] [Google Scholar]
- 63.Jaagus J. Climatic changes in Estonia during the second half of the 20th century in relationship with changes in large-scale atmospheric circulation. Theor Appl Climatol. 2006;83:77–88. doi: 10.1007/s00704-005-0161-0. [DOI] [Google Scholar]
- 64.Estonian Weather Service. Climate normals. Average air temperature (°C) 1981–2010. http://www.ilmateenistus.ee/kliima/kliimanormid/ohutemperatuur/?lang=en. Accessed 30.11.2016.
- 65.Kirjušina M, Deksne G, Marucci G, Bakasejevs E, Jahundoviča I, Daukšte A, et al. A 38-year study on Trichinella spp. in wild boar (Sus scrofa) of Latvia shows a stable incidence with an increased parasite biomass in the last decade. Parasit Vectors. 2015;8:137. doi: 10.1186/s13071-015-0753-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Pozio E. Adaptation of Trichinella spp. for survival in cold climates. FAWPAR. 2016;4:4–12. [Google Scholar]
- 67.Statistikaamet, 2017. www.stat.ee. Accessed 16.09.2017.
- 68.Laurimaa L, Süld K, Davison J, Moks E, Valdmann H, Saarma U. Alien species and their zoonotic parasites in native and introduced ranges: the raccoon dog example. Vet Parasitol. 2016;219:24–33. doi: 10.1016/j.vetpar.2016.01.020. [DOI] [PubMed] [Google Scholar]
- 69.Laurimaa L, Moks E, Soe E, Valdmann H, Saarma U. Echinococcus multilocularis and other zoonotic parasites in red foxes in Estonia. Parasitology. 2016;143:1450–1458. doi: 10.1017/S0031182016001013. [DOI] [PubMed] [Google Scholar]
- 70.Pitts GC, Bullard TR. Some interspecific aspects of body composition in mammals. In: Body composition in animal and man. Proceedings of symposium held May 4, 5 and 6, 1967 at the University of Missouri, Columbia. National Academy of Sciences Washington DC; 1968. p. 45–70.
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All data used and analyzed during the current study are included in this article.