

Key Points
TA-316, a novel CMA, enhances human platelet ex vivo generation from iPS cells.
TA-316 exhibited biased megakaryopoiesis at levels comparable to rhTPO and superior to eltrombopag.
Abstract
Signaling by thrombopoietin (TPO) in complex with its receptor, c-MPL, is critical for hematopoietic stem cell (HSC) homeostasis and platelet generation. Here we show that TA-316, a novel chemically synthesized c-MPL agonist (CMA), is useful for ex vivo platelet generation from human-induced pluripotent stem (iPS) cell–derived immortalized megakaryocyte progenitor cell lines (imMKCLs). Moreover, the generation is clinically applicable, because self-renewal expansion and platelet release is tightly controllable. TA-316 but not eltrombopag, another CMA, promoted both the self-renewal and maturation of imMKCLs, leading to more than a twofold higher platelet production than that achieved with recombinant human TPO (rhTPO). Interestingly, TA-316 seemed to favor MK-biased differentiation from bone marrow CD34+ HSC/progenitors and imMKCLs through the upregulation of vascular endothelial growth factor A and fibroblast growth factor 2. This result suggests TA-316 could facilitate the development of an efficient and useful system to expand platelets from imMKCLs.
Visual Abstract
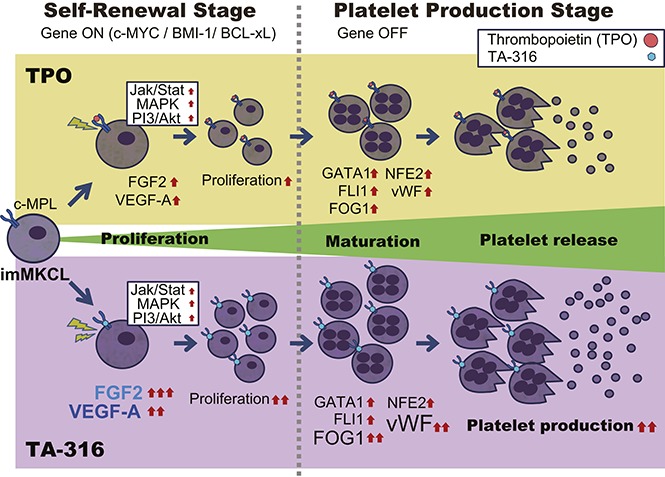
Introduction
Platelet transfusion plays a pivotal role in the management of patients with bleeding due to a severe decrease in platelet production or platelet dysfunction. However, platelets can be obtained only through blood donation, which represents an unstable supply of platelets for clinical use.1,2 Moreover, repeated transfusion leads to the induction of antibodies against allogeneic HLA and/or human platelet antigens on the donor-derived platelets, necessitating the use of identical HLA/human platelet antigens-matched platelets.3 To overcome these issues, there have been various attempts to expand human platelets ex vivo.4 It has been anticipated, for example, that the eventual use of human-induced pluripotent stem cells (hiPSCs) as a robust source of platelets would obviate or relieve the problem of allo-immunization.5 In that context, we established an in vitro iPS-Sac culture system for differentiation into megakaryocytes (MKs, platelet precursors),5,6 which also yields T lymphocytes.7 Unfortunately, the efficiency of the MK production is extremely low.5 We therefore established self-renewing immortalized MK progenitor cell lines (imMKCLs) from hiPSCs as a starting material of platelet supply, because they exhibit a capacity for the long-term expansion of MKs that produce a substantial yield of platelets.8
Thrombopoietin (TPO) is the primary regulator controlling human MK differentiation and platelet production, as well as the expansion of platelets from imMKCLs. However, TPO has failed as a standard therapy due to immunogenicity that worsens thrombocytopenia.8-10 Instead, several nonpeptidyl small molecules that activate c-MPL, the TPO receptor, have recently been developed.11,12 These include eltrombopag, an orally administered drug used to treat mostly immune-thrombocytopenia. The advantages of such chemically synthesized c-MPL agonists (CMAs) over peptide-based ligands include greater biological safety, lower immunogenicity, and lower manufacturing costs. Detailed examination of the activities of CMAs also promotes a better understanding of the signaling pathways that regulate megakaryopoiesis and thrombopoiesis. Considering that some CMAs activate signals that are preferentially specialized for platelet differentiation, we evaluated their effects on the expansion capacity of imMKCLs. This effort led us to identify TA-316 as a compound that increases platelet production from imMKCLs more efficiently and cost effectively than TPO or eltrombopag. Our results provide promising possibilities for the use of small-molecule compounds in the realization of regenerative medicine using hiPSCs.
Methods
Cell culture
Bone marrow (BM) CD34+ cells.
Human BM CD34+ cells were purchased from Lonza (Basel, Switzerland). Cells were suspended in StemSpan (Stemcell Technologies, Vancouver, BC, Canada) supplemented with 100 ng/mL recombinant human stem cell factor (rhSCF) (R&D Systems, Minneapolis, MN) and seeded onto 48-well plates (20 000 cells/500 μL per well). Thereafter, recombinant human TPO (rhTPO) (PeproTech, Rocky Hill, NJ), TA-316 (in-house synthesized), eltrombopag (ChemScene, Monmouth Junction, NJ, or synthesized in-house), or dimethyl sulfoxide (DMSO) (Wako Pure Chemical Industries [WAKO], Osaka, Japan) was added to the cell culture for 10 days in a humidified incubator at 37°C with 5% CO2. We confirmed no significant differences in the purities and proliferation of cells treated with purchased (ChemScene) and in-house synthesized eltrombopag (data not shown). In-house synthesized eltrombopag was used for all experiments unless otherwise noted.
C3H10T1/2 cells.
Mouse C3H10T1/2 cells (RIKEN BioResource Center, Ibaraki, Japan) were maintained in Basal Medium Eagle (Thermo Fisher Scientific, Waltham, MA) containing 10% fetal bovine serum (FBS) (Thermo Fisher Scientific). Cells were cultured in a humidified incubator at 37°C with 5% CO2.
hiPSC-derived induced hematopoietic progenitor cells (iHPCs).
hiPSCs (clone; Tkda3-4) were maintained in Dulbecco’s modified Eagle medium /Ham’s F12 medium (WAKO) containing 20% StemSure Serum Replacement (WAKO), 1% minimal non-essential amino acids solution (Thermo Fisher Scientific), 1% StemSure (WAKO), 50 mM monothioglycerol solution (×100) (WAKO), 292 μg/mL l-glutamine (Thermo Fisher Scientific), 100 U/mL penicillin (Thermo Fisher Scientific), and 100 μg/mL streptomycin (Thermo Fisher Scientific). Colonies were removed from the dish using dissociation solution for human embryonic stem/iPS cells (ReproCELL, Kanagawa, Japan) and seeded onto 0.1% gelatin-coated plates or dishes, along with mitomycin C (WAKO) treated (10 μg/mL, 1.5 to 2 hours) C3H10T1/2 cells (200 000 cells per well in 6-well plates or 800 000 cells per 100-mm dish). The plated cells were cultured in endothelial basal medium (EBM) (Iscove modified Dulbecco medium [Sigma-Aldrich, St Louis, MO]), supplemented with 15% FBS (Thermo Fisher Scientific), 292 μg/mL l-glutamine (Thermo Fisher Scientific), 100 U/mL penicillin (Thermo Fisher Scientific), 100 μg/mL streptomycin (Thermo Fisher Scientific), 0.05 mg/mL ascorbic acid (Sigma-Aldrich), 0.45 mM 1-thioglycerol (Sigma-Aldrich), and 1% insulin-transferrin-selenium supplement (Thermo Fisher Scientific), in the presence of 20 ng/mL recombinant human vascular endothelial growth factor (VEGF)-A165 (WAKO), 5 ng/mL recombinant human basic fibroblast growth factor (FGF) (ReproCELL), and 10 μM rock inhibitor (Y-27632, WAKO). The medium was changed every 2 to 4 days. hiPSCs were cultured for 14 days in a humidified incubator (37°C, 5% CO2) and formed sac-like structures (iPS-Sacs) containing iHPCs inside. Crude iHPCs were collected from iPS-Sacs on day 14 and further sorted using FACSAria III (BD Biosciences, San Jose, CA) into a CD34+/CD43+/CD41−/glycophorin-A (GPA−) fraction. Sorted iHPCs and unsorted iHPCs were seeded onto mitomycin C-treated C3H10T1/2 cells and cultured for 3 to 10 days in EBM medium supplemented with 50 ng/mL rhSCF (R&D Systems) and one of 100 ng/mL rhTPO, 100 ng/mL TA-316, 3000 ng/mL eltrombopag, or 0.05% DMSO.
imMKCLs.
imMKCL clone 7 was established from hiPSCs as previously reported.8 imMKCLs bear 3 doxycycline (DOX)-inducible transgenes, c-MYC, BMI1, and BCL-XL. As such, the addition of DOX turns on the expression of these transgenes (genes on), leading to proliferation of imMKCLs, whereas the depletion of DOX (genes off) induces their maturation. imMKCL clone 7 (which we named imMKCL Sev2) was cultured in EBM medium supplemented with rhSCF (R&D Systems) and 1 μg/mL DOX (TAKARA, Shiga, Japan) in the presence of rhTPO (PeproTech), eltrombopag (ChemScene), TA-316, or DMSO (vehicle). Cells were seeded onto 0.1% gelatin-coated 6-well plates (100 000 cells/mL, 2 mL per well) along with mitomycin C-treated C3H10T1/2 cells, and cultured in a humidified incubator at 37°C and 5% CO2, and passaged every 3 to 4 days.
UT-7/TPO cells and UT-7/erythropoietin (EPO) cells.
UT-7/TPO and UT-7/EPO cells were obtained from N. Komatsu (Juntendo University, Tokyo, Japan),13-15 and cultured in Iscove modified Dulbecco medium containing 10% FBS in the presence of 10 ng/mL rhTPO and 0.5 U/mL recombinant human EPO (rhEPO) (Eposin; Chugai Pharmaceutical, Tokyo, Japan), respectively. Cells were cultured in a humidified incubator at 37°C and 5% CO2.
Ba/F3 cells.
Murine pro–B-cell line Ba/F3 was obtained from RIKEN Cell Bank (Tsukuba, Japan). Stable transfectants such as Ba/F3-HuMpl and Ba/F3-MuMpl were established to transfect human or murine c-mpl.16 Ba/F3 cells and their transfectants were maintained in RPMI 1640 medium supplemented with 10% FBS and 10 ng/mL recombinant mouse interleukin-3 (rmIL-3) or 30 to 50 ng/mL rhTPO.
Cell growth assay
UT-7/TPO, UT-7/EPO, Ba/F3, Ba/F3-HuMpl, and Ba/F3-MuMpl cells were seeded onto 96-well plates (5000 cells/100 μL per well) and cultured for 4 days as described earlier in medium supplemented with rhTPO, TA-316, eltrombopag, rhEPO, or rmIL-3 in a humidified incubator at 37°C and 5% CO2. Cell viability was assessed using WST-8 assays (Cell Counting Kit-8; Dojindo Laboratories, Kumamoto, Japan). A one-tenth volume of WST-8 solution (10 μL) was added to the cell culture and incubated for 3 to 4 hours in a humidified incubator at 37°C and 5% CO2. The absorbance at 450 nm was measured using an absorbance spectrometer (SpectraMax 190; Molecular Devices, Sunnyvale, CA), and the number of viable cells was measured after subtracting the absorbance of the medium alone.
Real-time quantitative reverse transcription polymerase chain reaction (RT-PCR)
RNA was extracted using RNeasy Mini Kits (Qiagen, Valencia, CA) according to the manufacturer’s instructions, after which complementary DNAs were synthesized using PrimeScript RT Master Mix (Perfect Real Time; TAKARA). RT-PCR was carried out for 40 to 50 cycles of 1 minute at 60°C and 15 seconds at 95°C in an ABI PRISM 7700 Sequence Detector (Applied Biosystems, Foster City, CA). All Taqman primers and probes were obtained from Applied Biosystems: glyceraldehyde-3-phosphate dehydrogenase (GAPDH; Hs02758991_g1); GATA binding protein 1 (GATA1; Hs01085823_m1); zinc finger protein, FOG family member 1 (ZFPM1; Hs00542350_m1); nuclear factor, erythroid 2 (NFE2; Hs00232351_m1); Fli-1 proto-oncogene ETS transcription factor (FLI1; Hs00956711_m1); von Willebrand factor (VWF; Hs04397751_m1); VEGF A (VEGFA; Hs00173626_m1); and basic FGF2 (Hs00266645_m1). GAPDH served as an internal control.
Megakaryopoiesis/thrombopoiesis of imMKCLs
imMKCLs were cultured in EBM medium supplemented with rhSCF and 1 μg/mL DOX in the presence of rhTPO, eltrombopag, TA-316, or DMSO (vehicle). Cells were seeded onto 0.1% gelatin-coated 6-well plates (100 000 cells/mL, 2 mL per well) along with mitomycin C-treated C3H10T1/2 cells, and cultured in a humidified incubator at 37°C and 5% CO2 and passaged every 3 to 4 days. Cell proliferation was assessed based on 0.4% Trypan blue staining (Thermo Fisher Scientific), and the cumulative number of cells was calculated until day 11. To induce platelet production, c-MYC, BMI1, and BCL-XL genes were turned off by culturing the imMKCLs in DOX-free medium. After 4 days of culture, platelets and MKs were gently collected with their medium by pipetting, and acid citrate dextrose solution (85 mM trisodium citrate [WAKO], 104 mM D-glucose [WAKO], and 65 mM citric acid [WAKO]) was added to each sample (10% [volume-to-volume ratio] final concentration). MKs were centrifuged for 8 minutes at 440 g, after which the pellets were suspended in phosphate-buffered saline (PBS) containing 2% FBS.
Flow cytometry
Samples were analyzed on a FACSAria III using FACSDiva software (BD Biosciences). To analyze megakaryocytic differentiation and maturation, cultured BM CD34+ cells, iHPCs, and imMKCLs were collected and stained with antibodies as follows, and added to samples at the manufacturer’s recommended concentrations: phycoerythrin (PE)-conjugated anti-human CD34 (BD Biosciences), allophycocyanin (APC)-conjugated anti-human CD41 (BioLegend, San Diego, CA), Pacific blue-conjugated anti-human CD41 (Affymetrix, San Diego, CA), PE-conjugated anti-human CD42b (Affymetrix), APC-conjugated anti-human CD43 (Affymetrix), and APC-conjugated CD235a (GPA; BD Biosciences). To evaluate the number of multinucleate MKs, Hoechst 33342 (Thermo Fisher Scientific) was added to part of each sample (20 μg/mL final concentration), and the samples were incubated for 45 minutes in a water bath at 37°C and then washed with cold 2% FBS/PBS. All samples were then incubated for 30 minutes on ice in the dark. After washing with 2% FBS/PBS, cells were centrifuged for 5 minutes at 1500 rpm and resuspended with 2% FBS/PBS. Dead cells were excluded based on propidium iodide (Biomol, Hamburg, Germany) staining at a final concentration of 1 μg/mL.
Platelet production from imMKCLs
imMKCL-derived platelets were collected as described earlier. Total platelet numbers were determined using BD TruCount tubes (BD Biosciences), in combination with APC-conjugated anti-human CD41 (BioLegend) and PE-conjugated anti-human CD42b (Affymetrix) antibodies. Cells were stained for 15 minutes at room temperature and analyzed using fluorescence-activated cell sorting (FACS).
First procaspase activating compound (PAC-1) binding assay
The procedures used were described previously.8
Intracellular phospho-specific flow cytometry
The phosphorylation of signaling proteins in UT-7/TPO cells and imMKCLs was evaluated using the BD Phosflow assay (BD Biosciences). Before the tests, imMKCLs were cultured for 24 hours without rhTPO. UT-7/TPO cells and imMKCLs were then collected, washed twice with PBS, and resuspended in TPO-free culture medium (1 000 000 cells/500 µL per sample). Each sample was pre-incubated in a humidified incubator at 37°C and 5% CO2 for 30 minutes and stimulated with rhTPO, TA-316, eltrombopag, or DMSO for 15 to 240 minutes. During the last 30 minutes, APC-conjugated anti-human CD41a (BioLegend) antibody was added to the cells. For the 15-minute stimulation samples, antibodies were added at the same time as the TPO receptor agonists. Stimulated cells were fixed immediately using the same volume of pre-warmed BD Cytofix (BD Biosciences) (37°C, 10 minutes) and centrifuged at 600 g for 6 minutes. The pellets were resuspended in 1 mL of cooled Perm Buffer III (BD Biosciences) and incubated on ice for 30 minutes. After permeabilization, the cells were washed 3 times with 3 mL of 2% FBS/PBS (centrifuged at 600 g for 6 minutes) and stained with antibodies (PE-conjugated anti-STAT5 [pY694], Alexa 488-conjugated anti-STAT3 [pY705], PE-conjugated anti-STAT1 [pY701], PE-conjugated anti-AKT [pS473], Alexa 488-conjugated anti-extracellular signal-regulated kinase 1/2 [pT202/pY204], and/or corresponding isotype controls; BD Biosciences) for 60 minutes at room temperature in the dark. The cells were then washed with 3 mL of 2% FBS/PBS, centrifuged at 600 g for 6 minutes, resuspended in 200 μL of 2% FBS/PBS, and analyzed by FACS. Signals were normalized to no-agonist (Vehicle, DMSO alone) samples.
Statistical analysis
All results are presented as the mean ± standard deviation (SD). Statistical significance was analyzed using the Dunnett test. The level of significance was set at 0.05.
Results
Identification of TA-316 as a novel chemically synthesized CMA
We first assessed the abilities of CMAs synthesized in-house to promote the proliferation of UT-7/TPO cells, a human leukemia cell line that expresses c-MPL, and selected the most potent candidate, TA-316 (Figure 1) (data not shown). These effects by TA-316 were dependent on c-MPL, as evidenced by TA-316 enhancing the growth of UT-7/TPO (Figure 2A) and Ba/F3 cells with c-MPL (Ba/F3-HuMpl) (Figure 2C-D), but not UT-7/EPO (Figure 2B), Ba/F3 without human c-MPL (Ba/F3), or Ba/F3 with murine c-MPL (Ba/F3-MuMpl) cells (Figure 2C). The median effective concentrations (ie, EC50) of TA-316, rhTPO, and eltrombopag were 0.3 ± 0.03 ng/mL, 1.6 ± 0.17 ng/mL, and 55.3 ± 5.04 ng/mL, respectively, in assays using UT-7/TPO cells (Figure 2A), and 0.65 ± 0.23 ng/mL, 11.7 ± 0.74 ng/mL, and 20.7 ± 2.0 ng/mL, respectively, in assays using Ba/F3-HuMpl cells (Figure 2D). We confirmed TA-316 stimulated UT-7/TPO and Ba/F3-HuMpl cell proliferation more strongly than either rhTPO or eltrombopag (Figure 2A,D). Intracellular phosphoprotein analysis revealed that TA-316 induced sustained STAT5 phosphorylation (Figure 2E), which is likely to have elicited the more potent effect by TA-316.
Figure 1.

Structures of TA-316 and eltrombopag (olamine).
Figure 2.

TPO receptor-dependent cell proliferation and STAT5 phosphorylation. (A) UT-7/TPO and (B) UT-7/EPO cells were cultured for 4 days with various doses of rhTPO, TA-316, eltrombopag, or rhEPO, and the number of cells were counted using the WST-8 assay. (C) Ba/F3 cells, Ba/F3 cells with human c-MPL (Ba/F3-HuMpl), and Ba/F3 cells with murine c-MPL (Ba/F3-MuMpl) were similarly cultured and counted after 4 days with rmIL-3 (1 ng/mL), rhTPO (100 ng/mL), TA-316 (1 ng/mL), or eltrombopag (ChemScene; 100 ng/mL). (D) Ba/F3-HuMpl cells were similarly cultured and counted after 4 days with various doses of rhTPO, TA-316, or eltrombopag (ChemScene). (E) STAT5 phosphorylation in UT-7/TPO cells stimulated for 15 to 360 minutes with rhTPO (100 ng/mL), TA-316 (100 ng/mL), eltrombopag (ChemScene; 1000 ng/mL), or DMSO (vehicle) were evaluated using BD Phosflow (STAT5, pY694). △MFI values of pSTAT5 are shown as relative values to that of vehicle treatment. Data represent the mean and SD; *P < .05, †P < .05. MFI, mean fluorescence intensity; min, minutes; O.D., optical density; pSTAT5, phosphorylated STAT5.
TA-316 promotes biased megakaryopoiesis and upregulated MK lineage markers
We then examined the effect of TA-316 on BM CD34+ cell differentiation and observed that TA-316 increased the number of CD41+ MK lineage cells after 10 days of culture (Figure 3A) with continued enhancement of STAT5 activation in UT-7/TPO cells (data not shown and Figure 2E). Interestingly, TA-316 also upregulated the gene expression of 5 specific markers of the MK lineage (GATA1, ZFPM1, FLI1, NFE2, and VWF) in cultured BM CD34+ cells to a greater degree than rhTPO, whereas eltrombopag did not show this effect (Figure 3B).
Figure 3.

TA-316 induces megakaryopoietic differentiation from BM CD34+cells. BM CD34+ cells were cultured for 10 days with rhTPO (50 ng/mL), TA-316 (800 ng/mL), eltrombopag (3000 ng/mL), or DMSO (0.05%). The CD41+ cell population was analyzed using FACS (A), and the expressions of GATA1, ZFPM1, FLI1, NFE2, and VWF mRNA were evaluated using RT-PCR (B). GAPDH served as an internal control. Data represent the mean and SD; *P < .05, **P < .01. mRNA, messenger RNA.
TA-316 expands MKs from hiPSC-derived iHPCs ex vivo more efficiently than eltrombopag
We next prepared iHPCs using our iPS-Sac system5 and tested whether TA-316 affects MK production. We found that at 100 ng/mL, TA-316 increased MK numbers more efficiently than rhTPO and eltrombopag (supplemental Figure 1A). TA-316 also tends to promote multinuclearity and increase ploidy (supplemental Figure 1B). When the CD43+CD34+CD41−GPA− population was sorted from crude iHPCs using flow cytometry and cultured for another 5 days in the presence of rhTPO, TA-316, or eltrombopag, TA-316 and rhTPO increased the number of CD41+ MKs best (Figure 4). Collectively, the results summarized to this point indicate that TA-316 is a specific CMA with a greater ability to stimulate biased and efficient human megakaryopoiesis ex vivo that is comparable with rhTPO and superior to eltrombopag.
Figure 4.

TA-316 induces megakaryopoietic differentiation from iPS-Sac–derived hematopoietic progenitor cells. (A) Scheme of megakaryopoiesis and thrombopoiesis in iPS-Sac–derived iHPCs. hiPSCs cultured for 14 days with VEGF-formed sac-like structures (iPS-Sacs). CD43+/CD34+/CD41−/GPA− iHPCs were sorted from crude iHPCs in iPSC-Sacs using FACS and then cultured with rhTPO (100 ng/mL), TA-316 (100 ng/mL), or eltrombopag (3000 ng/mL). (B) The number of CD41+ cells was evaluated using FACS on days 3 and 5. Data represent the mean and SD; **P < .01, ***P < .001. n.s., not significant.
TA-316 expands MKs and platelets from imMKCLs ex vivo more efficiently than TPO or eltrombopag
We next cultured imMKCLs for 11 days in the presence of rhTPO, TA-316, or eltrombopag to reach the self-renewal stage, and then for an additional 4 days to reach the platelet production stage (Figure 5A). On day 11, the total number of cells obtained in culture with 200 ng/mL TA-316 was 1.5-fold to greater than twofold higher than in cultures with 50 ng/mL rhTPO and even 3000 ng/mL eltrombopag, respectively (Figure 5B). When we turned off c-MYC, BMI1, and BCL-XL expression to induce platelet production, TA-316 increased the number of CD41+CD42b+ platelets by more than twice that of rhTPO (P < .001), whereas eltrombopag had little effect on platelet production (Figure 5C). The relative number of CD41+CD42b+ platelets produced per single imMKCL cell treated with TA-316 was 1.3-fold higher than that with rhTPO. We attributed this higher platelet yield by TA-316 treatment to the combination of enhanced imMKCL expansion and platelet yield per cell. In addition, imMKCLs stimulated with TA-316 or rhTPO showed high levels of DNA content (Figure 6A; supplemental Figure 2). The level of binding to PAC-1 on platelets by thrombin stimulation, which is a key parameter of αIIbβ3 integrin-dependent platelet functionality, was the same in platelets generated using TA-316 or rhTPO (Figure 6B). An electron microscope image of platelets obtained under the TA-316 condition showed normal platelet structures (supplemental Figure 3). These results indicate that TA-316 enhanced the production of imMKCL-derived platelets and that the platelets showed a competency comparable to those produced with rhTPO.
Figure 5.

TA-316 promotes megakaryopoiesis and thrombopoiesis in imMKCLs. (A) System to generate platelets from imMKCLs cultured with rhTPO, TA-316, eltrombopag, or DMSO (vehicle). Cells were seeded onto mitomycin C-treated C3H10T1/2 feeder cells and passaged every 3 to 4 days. To turn off the c-MYC, BMI1, and BCL-XL genes in order to induce platelet production, beginning on day 11, the cells were cultured in DOX-free medium. After 4 days of culture, matured MKs and platelets were collected. (B) The number of imMKCLs was counted on day 11 based on Trypan blue assays. (C) On day 15, imMKCL-derived platelets were collected, and CD41+/CD42b+ platelets were counted using FACS. Data represent the mean and SD; **P < .01, ***P < .001.
Figure 6.

TA-316 induces maturation of imMKCLs and increases functional platelets. imMKCLs cultured with rhTPO (50 ng/mL), TA-316 (200 ng/mL), eltrombopag (1000 ng/mL), or DMSO (0.05%) were collected on day 11 (genes on) and day 15 (after 4 days without DOX, genes off). (A) Multinucleated (≥8 n) MKs and (B) PAC-1 binding on platelets were analyzed by FACS. Platelets were stimulated for 15 minutes with 1 U/mL thrombin from human plasma. Human washed platelets served as a positive control. (C) mRNA expressions of GATA1, ZFPM1 (FOG1), NFE2, FLI1, and VWF (genes on or off) were evaluated using Taqman PCR assays. GAPDH served as an internal control. Data represent the mean and SD; *P < .05, **P < .01. PLT, platelets.
We also observed that TA-316 persistently activated STAT5 in imMKCLs (supplemental Figure 4A) as it did in UT-7/TPO cells (Figure 2E). Further analysis revealed that TA-316 induced the phosphorylation of components in 3 major TPO signaling pathways: JAK/STAT, MAPK, and phosphatidylinositol 3-kinase/AKT, within imMKCLs (supplemental Figures 4B-E and 5). These results indicate that TA-316 acts on imMKCL as a TPO receptor agonist, and enhances imMKCL expansion and platelet production.
TA-316 preferentially enhances VEGFA and FGF2 expression
Consistent with the data of BM CD34+ cells shown in Figure 3B, GATA1, ZFPM1, FLI1, NFE2, and VWF were upregulated in imMKCLs at the platelet production stage to a greater degree with TA-316 than with rhTPO (Figure 6C), whereas eltrombopag showed little capacity to enhance the expression of these thrombopoiesis-specific genes in imMKCLs. To determine the molecular mechanisms underlying the beneficial actions of TA-316 on platelet production from imMKCLs, we performed DNA microarray-mediated transcriptional profiling of imMKCLs cultured with TA-316 and found greater upregulation of growth factor activity-related genes, including VEGFA and FGF2, than that seen with rhTPO (supplemental Table 1). Real-time quantitative PCR confirmed the superior effect of TA-316 on the induction of these genes in imMKCLs (Figure 7A). Moreover, earlier observations that VEGFA and FGF2 are involved in the MK maturation associated with thrombopoiesis17,18 suggest the greater efficacy of TA-316 may reflect VEGFA and FGF2 upregulation. This hypothesis was confirmed by pharmacologic effects using an anti-VEGF receptor antagonist, axitinib (5 nM) or an anti-FGF receptor antagonist, PD173074 (100 nM). These drugs reversed the cell proliferation of imMKCLs in the presence of TA-316, whereas epithelial growth factor receptor antagonist, gefitinib, did not (Figure 7B). Neither axitinib nor PD173074 showed an effect on platelet production or the expression of differentiation markers (CD41 and CD42b) in imMKCLs (data not shown).
Figure 7.

An inhibitor of FGF or VEGF receptors eliminates the superior effect of TA-316 on imMKCL proliferation. imMKCLs cultured with rhTPO (50 ng/mL), TA-316 (200 ng/mL), eltrombopag (1000 ng/mL), or DMSO (0.05%). (A) Cells were collected on day 11 (genes on). mRNA expressions of VEGFA and FGF2 (genes on) were evaluated using Taqman PCR assays. GAPDH served as an internal control. (B) FGFR antagonist PD173074 (100 nM; WAKO), VEGFR antagonist axitinib (5 nM; Toronto Research Chemicals, Toronto, Canada), or epithelial growth factor receptor antagonist gefitinib (1 μM, Santa Cruz Biotechnology, Dallas, TX) was added to the imMKCLs, which were cultured with TA-316 (200 ng/mL) for 4 days (genes on). Cell proliferation was assessed based on 0.4% Trypan blue staining (Thermo Fisher Scientific). Data represent the mean and SD; *P < .05, **P < .01.
Discussion
A stable supply for platelet transfusion by hiPSC technology needs an efficient and robust cell culture system that generates large quantities of platelets at low cost. To address this issue, we have developed stable imMKCLs from hiPSCs, which enable the long-term expansion of MKs and efficient production of platelets.8 In the present study, we focused particularly on TPO/c-MPL signaling, which plays a primary role in megakaryopoiesis and thrombopoiesis, and identified TA-316 as a novel small-molecule CMA that promotes the proliferation and differentiation of human c-MPL–expressing cells. Indeed, in proliferation assays using UT-7/TPO cells and Ba/F3-HuMpl cells, TA-316 showed an EC50 of 0.3 ng/mL and 0.65 ng/mL, values superior to either rhTPO (1.6 ng/mL and 11.7 ng/mL) or another CMA, eltrombopag (55.3 ng/mL and 20.7 ng/mL) (Figure 2A,D). Of the 3 CMAs examined, TA-316 was most potent for the proliferation of UT-7/TPO cells, Ba/F3-HuMpl cells, and imMKCLs (Figure 5B). In differentiation settings using BM CD34+ cells and human iPSCs, TA-316 had a comparable effect to rhTPO and a more potent one than eltrombopag (Figures 3A and 4B). Interestingly, TA-316 seemed to be also most potent for the terminal maturation of MKs from human hiPSCs (supplemental Figure 1A) and platelet yield from imMKCLs (Figure 5C).
In addition to the fact that both TA-316 and eltrombopag showed no effect on mouse c-mpl (Figure 2C) and showed later activation patterns (Figure 2E; supplemental Figures 4A and 5), TA-316 and eltrombopag share chemical structures (Figure 1). Another chemical, NIP-004, from which TA-316 was derived, also has no effect on mouse c-mpl.16 Therefore, despite the different potencies, these compounds may function similarly at the molecular level, but in a way is distinct from TPO.
We also demonstrated that TA-316 prevails over TPO and eltrombopag in the ex vivo production of platelets from hiPSCs, as evidenced by the final platelet yield for which TA-316 induced a greater than twofold expansion compared with rhTPO or eltrombopag (Figure 5C). These findings suggest that the combination of TA-316 with imMKCLs is applicable to a culture system for the clinical supply of hiPSC-derived platelets.8
To further investigate the effect of TA-316 on the platelet differentiation of c-MPL–expressing cells, we analyzed the gene expression of specific markers of MK lineage in culture with rhTPO, eltrombopag, or TA-316. TA-316 tended to enhance the gene expression of GATA1, ZFPM1 (also known as FOG1), FLI1, NFE2, and VWF in BM CD34+ cells (Figure 3B) and imMKCLs (Figure 6C) at levels higher than rhTPO or eltrombopag. We also noted a unique effect of TA-316 on the expression of growth factor activity-related genes in imMKCLs. The expressions of VEGFA and FGF2 genes were preferentially increased in imMKCLs cultured with TA-316 (Figure 7A). Consistent with data that show TA-316 has superior activity for the promotion of imMKCL proliferation, VEGFA reportedly plays an important role in hematopoietic stem cell (HSC) survival.19 Considering that TA-316 stimulates a direct pathway to MKs from HSCs or iHPCs20,21 as well as the induction of VEGFA, we propose that TA-316 may promote the self-renewal activity of imMKCLs as well as HSCs/iHPCs. As shown in Figure 2E and supplemental Figure 4A, TA-316 exhibited its maximum signal intensity at times (120 to 240 minutes) later than rhTPO (15 minutes) in terms of STAT5 activation. The differences in TPO-receptor binding and dissociation kinetics between rhTPO and TA-316 may provide a key to understanding why TA-316 more effectively mediates platelet production (Figure 5C).
Chemically synthesized compounds other than CMAs have been reported to show positive effects in the production of MKs and platelets from HSCs/iHPCs ex vivo. MK1, a platelet-derived growth factor receptor agonist, promotes megakaryopoiesis from CD34+ cells.22 In addition, StemRegenin 1, an antagonist of the aryl hydrocarbon receptor, increases the number of proplatelet-producing MKs from CD34+ cells by repressing CYP1B1 expression.23 We confirmed that StemRegenin 1 increases the number of platelets from imMKCLs when cultured with TA-316 (data not shown). Thus, TA-316 could be applied in combination with other small molecule modulators that function independent of the TPO/c-MPL signal.
In conclusion, our findings demonstrate that TA-316 enhances the production of human platelets from imMKCLs and that TA-316 has the potential to become an indispensable tool for the expansion of platelets from hiPSCs, in combination with an imMKCL system. Moreover, TA-316 could help elucidate the roles of factors involved in biased megakaryopoiesis and thrombopoiesis from HSCs/iHPCs, such as VWF, VEGFA, and FGF2.
Supplementary Material
The full-text version of this article contains a data supplement.
Acknowledgments
The authors thank N. Komatsu for providing the UT-7/TPO and UT-7/EPO cells, H. Hoshino for technical assistance, and P. Karagiannis for reading the manuscript.
This work was supported in part by the Highway Program for Realization of Regenerative Medicine, Research Center Network for Realization of Regenerative Medicine from the Ministry of Education, Science and Technology/Japan Agency for Medical Research and Development (AMED) (K.E.), a Regenerative Medicine grant from the Ministry of Health, Labor and Welfare/AMED (K.E.), and by the Core Center for iPS Cell Research, Research Center Network for Realization of Regenerative Medicine from AMED (S.N., N.S., and K.E.). This work was also supported in part by Nissan Chemical Industries, Ltd. (Tokyo, Japan).
Authorship
Contribution: A.A. designed and performed the experimens, and wrote the paper; T.K., N.A., S.N., and A.S. performed the experiments; N.S. interpreted the data and wrote the paper; T. Nakamura and H.N. were involved in the initial idea of the project; and T. Nishino and K.E. designed the experiments, interpreted the data, and wrote the paper.
Conflict-of-interest disclosure: T. Nishino, T. Nakamura, H.N., and K.E. have applied for a patent related to this work; H.N. is a scientific advisory member of ReproCELL and Megakaryon Corporation; and H.N. and K.E. are founders of Megakaryon Corporation. The interests of K.E. were reviewed and are managed by Kyoto University in accordance with its conflict-of-interest policies. The remaining authors declare no competing financial interests.
Correspondence: Koji Eto, Department of Clinical Application, Center for iPS Cell Research and Application, Kyoto University, 53 Kawaharacho, Shogoin, Sakyoku, Kyoto 606-8507, Japan; e-mail: kojieto@cira.kyoto-u.ac.jp.
References
- 1.Basu D, Kulkarni R. Overview of blood components and their preparation. Indian J Anaesth. 2014;58(5):529-537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Cid J, Harm SK, Yazer MH. Platelet transfusion - the art and science of compromise. Transfus Med Hemother. 2013;40(3):160-171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Schiffer CA. Diagnosis and management of refractoriness to platelet transfusion. Blood Rev. 2001;15(4):175-180. [DOI] [PubMed] [Google Scholar]
- 4.Walasek MA, van Os R, de Haan G. Hematopoietic stem cell expansion: challenges and opportunities. Ann N Y Acad Sci. 2012;1266:138-150. [DOI] [PubMed] [Google Scholar]
- 5.Takayama N, Nishimura S, Nakamura S, et al. Transient activation of c-MYC expression is critical for efficient platelet generation from human induced pluripotent stem cells. J Exp Med. 2010;207(13):2817-2830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Takayama N, Nishikii H, Usui J, et al. Generation of functional platelets from human embryonic stem cells in vitro via ES-sacs, VEGF-promoted structures that concentrate hematopoietic progenitors. Blood. 2008;111(11):5298-5306. [DOI] [PubMed] [Google Scholar]
- 7.Nishimura T, Kaneko S, Kawana-Tachikawa A, et al. Generation of rejuvenated antigen-specific T cells by reprogramming to pluripotency and redifferentiation. Cell Stem Cell. 2013;12(1):114-126. [DOI] [PubMed] [Google Scholar]
- 8.Nakamura S, Takayama N, Hirata S, et al. Expandable megakaryocyte cell lines enable clinically applicable generation of platelets from human induced pluripotent stem cells. Cell Stem Cell. 2014;14(4):535-548. [DOI] [PubMed] [Google Scholar]
- 9.Kaushansky K, Lok S, Holly RD, et al. Promotion of megakaryocyte progenitor expansion and differentiation by the c-Mpl ligand thrombopoietin. Nature. 1994;369(6481):568-571. [DOI] [PubMed] [Google Scholar]
- 10.Bartley TD, Bogenberger J, Hunt P, et al. Identification and cloning of a megakaryocyte growth and development factor that is a ligand for the cytokine receptor Mpl. Cell. 1994;77(7):1117-1124. [DOI] [PubMed] [Google Scholar]
- 11.Ikeda Y, Miyakawa Y. Development of thrombopoietin receptor agonists for clinical use. J Thromb Haemost. 2009;7(suppl 1):239-244. [DOI] [PubMed] [Google Scholar]
- 12.Erickson-Miller CL, Delorme E, Tian SS, et al. Preclinical activity of eltrombopag (SB-497115), an oral, nonpeptide thrombopoietin receptor agonist. Stem Cells. 2009;27(2):424-430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Komatsu N, Kunitama M, Yamada M, et al. Establishment and characterization of the thrombopoietin-dependent megakaryocytic cell line, UT-7/TPO. Blood. 1996;87(11):4552-4560. [PubMed] [Google Scholar]
- 14.Komatsu N, Nakauchi H, Miwa A, et al. Establishment and characterization of a human leukemic cell line with megakaryocytic features: dependency on granulocyte-macrophage colony-stimulating factor, interleukin 3, or erythropoietin for growth and survival. Cancer Res. 1991;51(1):341-348. [PubMed] [Google Scholar]
- 15.Komatsu N, Yamamoto M, Fujita H, et al. Establishment and characterization of an erythropoietin-dependent subline, UT-7/Epo, derived from human leukemia cell line, UT-7. Blood. 1993;82(2):456-464. [PubMed] [Google Scholar]
- 16.Nakamura T, Miyakawa Y, Miyamura A, et al. A novel nonpeptidyl human c-Mpl activator stimulates human megakaryopoiesis and thrombopoiesis. Blood. 2006;107(11):4300-4307. [DOI] [PubMed] [Google Scholar]
- 17.Casella I, Feccia T, Chelucci C, et al. Autocrine-paracrine VEGF loops potentiate the maturation of megakaryocytic precursors through Flt1 receptor. Blood. 2003;101(4):1316-1323. [DOI] [PubMed] [Google Scholar]
- 18.Bruno E, Cooper RJ, Wilson EL, Gabrilove JL, Hoffman R. Basic fibroblast growth factor promotes the proliferation of human megakaryocyte progenitor cells. Blood. 1993;82(2):430-435. [PubMed] [Google Scholar]
- 19.Gerber HP, Malik AK, Solar GP, et al. VEGF regulates haematopoietic stem cell survival by an internal autocrine loop mechanism. Nature. 2002;417(6892):954-958. [DOI] [PubMed] [Google Scholar]
- 20.Notta F, Zandi S, Takayama N, et al. Distinct routes of lineage development reshape the human blood hierarchy across ontogeny. Science. 2016;351(6269):aab2116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Yamamoto R, Morita Y, Ooehara J, et al. Clonal analysis unveils self-renewing lineage-restricted progenitors generated directly from hematopoietic stem cells. Cell. 2013;154(5):1112-1126. [DOI] [PubMed] [Google Scholar]
- 22.Boitano AE, de Lichtervelde L, Snead JL, Cooke MP, Schultz PG. An image-based screen identifies a small molecule regulator of megakaryopoiesis. Proc Natl Acad Sci USA. 2012;109(35):14019-14023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Strassel C, Brouard N, Mallo L, et al. Aryl hydrocarbon receptor-dependent enrichment of a megakaryocytic precursor with a high potential to produce proplatelets. Blood. 2016;127(18):2231-2240. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.