

Abstract
Just as evolutionary biologists endeavour to link phenotypes to fitness, sport scientists try to identify traits that determine athlete success. Both disciplines would benefit from collaboration, and to illustrate this, we used an analytical approach common to evolutionary biology to isolate the phenotypes that promote success in soccer, a complex activity of humans played in nearly every modern society. Using path analysis, we quantified the relationships among morphology, balance, skill, athleticism and performance of soccer players. We focused on performance in two complex motor activities: a simple game of soccer tennis (1 on 1), and a standard soccer match (11 on 11). In both contests, players with greater skill and balance were more likely to perform better. However, maximal athletic ability was not associated with success in a game. A social network analysis revealed that skill also predicted movement. The relationships between phenotypes and success during individual and team sports have potential implications for how selection acts on these phenotypes, in humans and other species, and thus should ultimately interest evolutionary biologists. Hence, we propose a field of evolutionary sports science that lies at the nexus of evolutionary biology and sports science. This would allow biologists to take advantage of the staggering quantity of data on performance in sporting events to answer evolutionary questions that are more difficult to answer for other species. In return, sports scientists could benefit from the theoretical framework developed to study natural selection in non-human species.
Keywords: performance, motor skill, sport, evolutionary sports science, soccer
1. Introduction
In any population, some individuals perform better than others. In humans, as with many other animals, individuals advertise their ability to perform under pressure through ritualized combat, usually called sport. Elite performance in a sport brings status, resources and mating opportunities. As in most sexual species, males engage in these ritualized displays of performance more often than females. In fact, studies of diverse cultures showed that men engage in competitive sports more often than women do [1,2], reinforcing the view that sporting events may help establish social hierarchies important for mate choice [3]. Consistent with this view, professional athletes are among the most admired and highest paid members of modern societies.
Sports scientists endeavour to identify the traits that determine athletic performance just as evolutionary biologists endeavour to link phenotypes to fitness. Yet sports scientists and evolutionary biologists operate in isolation of one another. At first glance, their goals may seem disparate. Sports scientists seek to identify the qualities that make an elite human competitor [4,5], whereas evolutionary biologists want to know how heritable phenotypes affect the reproductive success of wild animals [6–8]. However, both fields address the same question: what drives variation in performances among individuals? As such, much can be gained from a cross-disciplinary approach to the study of human performance and its consequences for success in organized sports.
Sports scientists and evolutionary biologists may pursue similar questions, but the way they approach these questions substantially differs between the disciplines [1]. Sports scientists have focused on proximate explanations for great athletic performance—or, how an athlete interacts with his or her environment (e.g. diet, training) to attain some capacity for success. By understanding how the environment shapes athletes, one can manipulate environmental conditions to optimize athletic performance. By contrast, evolutionary biologists seek ultimate explanations for the phenotypes observed within populations; thus, an evolutionary biologist would be more interested in why an ancestral environment selected for athletic ability. Still, the proximate mechanisms that link traits to performance serve as the raw material for natural selection. This view of evolutionary biology was formalized by Arnold [9] in his paradigm relating morphology, performance and fitness in non-human animals. This paradigm works equally well for studies of humans.
Sports scientists would benefit from the protocols that evolutionary biologists use to quantify natural selection. In sports science, talent identification programmes are used to predict which individuals are most likely to attain elite competitive status, based on differences in the characteristics of elite versus sub-elite athletes [5,10,11]. This approach has helped to identify and nurture talent in sports such as athletics, swimming and rowing; where power, speed and endurance determine success. However, there has been a limited uptake of quantitative protocols for talent identification across most professional team sports [5,12]. Protocols that compare a group of elite athletes with a group of non-elite athletes provide a coarse understanding of the attributes required to reach elite status, and yet the success of an individual depends on their ability relative to others. In team sports, predicting whether or not an individual will become ‘elite’ is less important than predicting how they will perform within elite competitions. In other words, identifying which people could become professional players is not the same as identifying which people could become the best professional players. The latter question requires analyses of variation among individuals, which is remarkably uncommon in sports science [13] yet routine in evolutionary biology, especially when quantifying natural selection [9,14,15].
Evolutionary biologists can also advance their discipline by working with sports scientists. Because sports generate a staggering quantity of data on the size, age, and performance of competitors, sport science offers evolutionary biologists unique opportunities to explore the drivers of success in complex activities. Studies of animal performance show that success in complex activities—such as fighting conspecifics or escaping predators—depend on differences in speed [8,16], size [17], strength [18–20] and physiology [21]. Still, these studies have almost exclusively focused on quantifying extreme capacities of animals, such as maximal strength, speed or endurance [22,23], disregarding traits such as agility and skill (but see [23–27]). Because agility and skill may be compromised at high speeds or forces [23,25,28], evolutionary biologists have missed an important aspect of animal performance that shapes natural selection. Although evolutionary biologists rarely use data from sports to address major theories, some researchers have explored a few areas, such as: (i) how body shape facilitates or constrains success [29–31], (ii) how costs and benefits influence deception [32], and (iii) how athletic performance deteriorates with age [33]. Additional studies of human athletes can help fill gaps in our knowledge about the correlational selection of traits.
In this study, we explored the biological basis of success in soccer, a complex activity of humans that occurs in nearly every modern society. Our study was guided by the idea that the strengths of sports science and evolutionary biology can be integrated to study the biological fundamentals that shape variation in complex traits. In the sporting arena, as in the wild, excellence in physical activities relies on multiple dimensions of morphology and behaviour (as quantified by sports scientists). Furthermore, variation in traits and performance among individuals drives success (as quantified in evolutionary biologists). Soccer is an ideal sport for an integrative study such as ours, because soccer enables one to identify, isolate and quantify the traits that potentially underlie the relative success of many individuals in multiple populations [10,34]. Soccer is the world's most popular team sport, played by more than 200 million registered competitors and transcends national, cultural, religious and gender boundaries. Here, we quantified the relative importance of morphology, athleticism, motor skill and balance among elite soccer players to predict individual variation in simplified one-on-one soccer-tennis games, and standard 11-a-side competitive soccer matches. We do not only measure success in terms of their individual actions, but in the standard 11-a-side competitive matches we also quantified each player's importance within the network of the team using social network analyses.
2. Material and methods
We studied players from the University of Queensland Football Club that play in the semi-professional league of Brisbane Men's Football Premier and Reserves division. For each individual player (n = 32), we measured seven parameters of morphology, five parameters of maximum athletic performance, five parameters of motor skill function, and a single measure of static balance (table 1). We also quantified individual ability in two separate soccer-game scenarios: (i) simplified one-on-one soccer-tennis games (n = 27), and (ii) standard 11-a-side competitive soccer games (n = 24).
Table 1.
Measurements of morphology, balance, athleticism, motor skill and soccer performance, as quantified for the players used in our study.
| morphology | balance | maximum athletic performance | motor skill function | soccer performance |
|---|---|---|---|---|
| (i) height (m) (ii) full leg length (m) (iii) full arm length (m) (iv) upper-arm girth (m) (v) lower-arm girth (m) (vi) upper-leg girth (m) (vii) lower-leg girth (m) |
static balance: average of left and right foot | fastest sprint speed over 40 m (m s−1) fastest sprint speed through 44.8 m agility course (m s−1) fastest speed over 1500 m (m s−1) highest lower-body power: static jump distance (cm) greatest lower-body endurance: timed wall squat (s) |
juggling ability: keep-up ability of size 1 football using only feet (average) passing accuracy: passing towards target at 15 and 30 m volley accuracy: kicking served ball towards target at 5 and 10 m heading accuracy: heading served ball towards target at 5 and 10 m dribbling speed: fastest speed when kicking a ball through 44.8 m agility course (m s−1) |
soccer-tennis ability total accumulated score in tournament match performance (i) match success—across all tasks (ii) match activity—across all tasks (iii) match efficiency—across all tasks team performance social network analysis on ball movement |
(a). Morphology
For each individual, we measured upper and lower limb lengths, total height, torso-length, and the maximum circumference of leg and arm muscles using manual positioning of a tape measure. Mass was also measured using a standard commercially available balance (±0.01 kg). The first component of a principal component analysis (PCA) based on linear measurements (PCM) for all players from the soccer-tennis study explained 51% of the variation observed in the data (electronic supplementary material, table S1). As all vectors of PCM1 loaded in the same direction, this represented a measure of overall body size. The second component of the PCM (PCM2) explained 31% of the variation and represented a description of body shape (electronic supplementary material, table S1). Positive values of PCM2 were representative of larger arm and leg girths and shorter limbs and height.
(b). Maximum athletic ability
For each individual, we measured the following athletic ability traits: (i) maximum speed over 1500 m, (ii) total squat time, (iii) maximum jumping distance, (iv) fastest sprint speed over 40 m, (v) and fastest speed through an agility circuit. Performances were quantified using identical methods to those outlined in Wilson et al. [23].
The first component of a PCA based on measures of maximum athletic capacity from the soccer-tennis study explained 48% of the variation observed in the data (electronic supplementary material, table S3). All vectors of PCATH1 loaded in the same direction, and because larger positive values were indicative of better athleticism, PCATH1 represented a measure of overall maximum athletic performance. The second component of the PCATH explained 25% of the variation and described the negative association between acceleration/power with maximum sprinting speeds (electronic supplementary material, table S3). Positive values of PCATH2 were representative of faster speeds through the agility circuit and longer jump distances (rapid acceleration and high lower-body power) and lower top speeds over 40 m and 1500 m (low linear running speeds). The PCA based on the athletic performance of players from the 11-a-side competitive soccer game experiment are presented in electronic supplementary material, table S4.
(c). Motor skill function
For each player, we measured performance in five soccer-specific motor-skill tasks: (i) maximum dribbling speed, (ii) average juggling (i.e. keep-up) ability of a size 1 football, (iii) static-ball passing accuracy, (iv) volley-kick accuracy, and (v) heading accuracy. Motor skill traits were quantified using identical methods to those outlined in Wilson et al. [23] and represent repeatable measures of individual skill. The first component of the PCA based on measures of skill performance (PCSK1) from the soccer-tennis study explained 48% of the variation observed in the data (electronic supplementary material, table S5). All vectors of PCSK1 loaded in the same direction, and because larger positive values were indicative of better skill in each trait, PCSK1 represented a measure of overall skill performance. The second component of the PCA for measures of motor skill performance (PCSK2) explained 18% of the variation and described the negative association between juggling ability and passing ability (volley and passing accuracy) (electronic supplementary material, table S5). The PCA based on the athletic performance of players from the 11-a-side competitive soccer game experiment are presented in the electronic supplementary material, table S6.
(d). Balance
An individual's relative balance was taken from the average of three trials (for each leg) where we recorded the total time taken to lose balance when standing on high-density foam (40 × 40 × 7.5 cm high-density physiotherapy foam) with their eyes closed and non-standing leg flexed at the knee at a right angle.
(e). Soccer-tennis performance
We quantified an individual's soccer-tennis performance using a tournament design involving 30 players. Soccer-tennis is a commonly employed training activity for professional soccer players and provides a statistically independent measure of individual success that probably relies upon a similar suite of underlying morphological and performance traits as 11-a-side soccer matches. Games of soccer-tennis were played on a 16 m long by 8 m wide grass court with a 1 m high net in the middle. Players alternated a kick-serve (after bouncing the ball when dropped from their hands) from behind the baseline and each player was allowed a maximum of two touches of the soccer ball and one bounce on their side of the court before they had to return the ball into the opponent's side of the court. Games consisted of a total of 10 points and players accumulated points throughout a tournament based on their score in each game. Thus, if an individual played three games and their results were an 8-2 win, 10-0 win and 5-5 draw, then their total score was 23 points (with a maximum of 30 points).
For our experiment, one-on-one soccer-tennis games were conducted in a tournament design [35] where each individual played 15 games, thus allowing a detailed hierarchy of success to be determined for each player. Individuals were randomly placed into five groups of six individuals, with all individuals playing each other once. After this first round, all players were then randomly distributed to another five groups of six individuals, with all individuals playing each other once again. A third round was also conducted using five groups of six individuals. Analyses were conducted using only the 29 players whose morphology, balance, motor skill and athletic ability were also quantified.
(f). Match performance
We explored the underlying basis of success in 11-a-side soccer games to replicate the more realistic, complex and interactive competitive games that are played by most soccer players and encompass many team sports. To quantify individual match ability, we recorded footage from 10 in-season matches (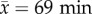 observed per match) from two Division 1 leagues (premier league, n = 5; reserve league, n = 5). Four players participated in both leagues. Matches were recorded from an elevated position using a Sony HD camcorder that enabled individuals to be identified using post-match analyses. Each ball involvement by an individual was categorized into the following five skilled match tasks: challenging (player and opposition compete for the ball using a header or tackle), dribbling (player has three or more consecutive touches of the ball), intercepting (player gains ball from opponent), passing (player delivers the ball to another player on his team) and shooting (player strikes ball at opposition's goal). For each action we recorded whether this was successful or unsuccessful. An action was classified as successful when the player's team retained possession following the execution of a skilled task, with the exception of shooting where only an on-target shot was considered as successful. We only chose actions and defined success in a way that ensured that individual success always had a net positive for the team. This does not mean that an individual made the best choice (i.e. a player could have completed an alternative action that provided a greater help to their team), but regardless, any successful action was still at least a positive action for the team.
observed per match) from two Division 1 leagues (premier league, n = 5; reserve league, n = 5). Four players participated in both leagues. Matches were recorded from an elevated position using a Sony HD camcorder that enabled individuals to be identified using post-match analyses. Each ball involvement by an individual was categorized into the following five skilled match tasks: challenging (player and opposition compete for the ball using a header or tackle), dribbling (player has three or more consecutive touches of the ball), intercepting (player gains ball from opponent), passing (player delivers the ball to another player on his team) and shooting (player strikes ball at opposition's goal). For each action we recorded whether this was successful or unsuccessful. An action was classified as successful when the player's team retained possession following the execution of a skilled task, with the exception of shooting where only an on-target shot was considered as successful. We only chose actions and defined success in a way that ensured that individual success always had a net positive for the team. This does not mean that an individual made the best choice (i.e. a player could have completed an alternative action that provided a greater help to their team), but regardless, any successful action was still at least a positive action for the team.
For each of the five match activities, three measures of performance were quantified: (i) successes (number of attempted skilled actions that were successful), (ii) attempts (number of attempted skilled actions involving the ball), and (iii) efficiency (success/attempts). These three different match traits were selected to represent the different types of players involved in 11-a-side games. We also calculated measures of match success, activity and efficiency across all tasks combined to provide metrics of overall match performance; total successes, total activity, and total efficiency. Since our initial analyses revealed that the execution of skilled tasks in matches varies significantly across 5 min intervals but not 15 min intervals, players that participated in a match for less than 15 min were excluded from analyses. For measures of match success and activity, each of the metrics analysed were divided by the number of minutes that an individual played in each game. An average of a player's successes and activity across their matches was then calculated and used in subsequent analyses. As measures of efficiency were proportions and not sensitive to the number of minutes played in a game, a player's average efficiency across matches was calculated and used in subsequent analyses.
(g). Player connectedness within matches
Social network analysis was used to quantify individual player performance and connectedness within a team. To do this, we used the igraph package [36] within the R software environment [37] to construct a complete network [38] for every match in the tournament, of all players within one team. Each player formed a node, and directed edges [39] linked players in the direction they passed to one another, weighted according to the number of passes. So if player A had successfully passed to player B five times in a game, a directed edge with a weight of five would link from player A to player B. There would be no reciprocal edge from player B to player A unless the former had also successfully passed to the latter, in which case this directed edge would be weighted according to the number of such passes.
From these match networks, seven measures of connectedness were calculated for each player (node) based on the successful passes executed between pairs of players (edges): out-degree (number of players one successfully passes to), in-degree (number of players one receives passes from), overall-degree (total number of players one is connected to via passing, sum of out- and in-degree), out-strength (number of passes one successfully makes), in-strength (number of passes one successfully receives) and overall-strength (total number of successful passing interactions one is involved in (sum of out- and in-strength). In addition, we calculated the weighted undirected closeness centrality, hereafter referred to as closeness—the inverse of the average weighted number of steps from a player to each other player in the network. Closeness treats ties as undirected, equating passes sent or received. In this case, it reflects the ease with which a player could send or receive the ball to or from any other player through the network [38,40]. The complete network for each game is presented in the electronic supplementary material, figure S1.
(h). Statistical analyses
We used a path model to estimate the relationships among morphology and performance in soccer tennis. The first set of paths related morphology to balance, skill and athleticism. The second set of paths related each of these variables to the player's score. Scores of the first and second principal components for morphology, athletic ability or skill were used as variables in the path models. All analyses were performed with the R statistical package [37].
We used a multivariate analysis of variance to see how a player's balance, athleticism and skill affected his scores in three aspects of 11-a-side soccer: successes, activity and efficiency. Following this analysis, we used generalized linear modelling to estimate the most likely effect of each independent variable on each dependent variable. We used Akaike's information criterion (AIC) to compare a model with a Gaussian distribution of error to one with a gamma distribution of error [41]. To estimate the fixed effects, we used multimodel averaging as described by Burnham and Anderson [42]. First, we used the MuMIn library [43] to fit all possible models to the data. Then, we calculated the AIC and Akaike weight of each model, the latter variable being the probability that the model best describes the data. Finally, we calculated the weighted average of each parameter including estimates from all models. The resulting values of parameters were used to calculate the most likely mean for each group.
To investigate the relationship between balance, athleticism and skill with network performance, we fitted mixed effects models using the lme4 package in the software package R, which account for the dependence of some players appearing in multiple games. Random intercepts were fit for game ID, player ID and player's stated position for each game. Player time on the field in each game, and the league of the game, were modelled as fixed effects and included as controls. Analyses predicting outcomes from each ability indicator separately indicated that only PCSK1 and PCATH2 reliably predicted any network outcomes. Because of this, these two predictors were included together as fixed predictors of each outcome. Significance tests were modelled using the lmerTest package in R, based on the Satterthwaite approximation, which results in fractional degrees of freedom.
3. Results
Path analysis revealed a link between morphology and total points in the soccer tennis tournament, mediated primarily by skill (figure 1d). Both greater balance and greater skill (PCSK1) increased a player's score (figure 1a,b). However, only PCSK1 was affected by morphology (PCM1), and based on its loadings (electronic supplementary material, table S1), this shows that players with greater mass or girth were more skilled (figure 1d).
Figure 1.

Relationship between morphology, maximum athletic capacity, motor skill function and balance with performance in the soccer-tennis tournament. We found (a) overall motor skill function (PCSK1) and (b) balance were highly positively correlated with cumulative soccer-tennis score. (c) Overall athletic ability (PCATH1) was not associated with soccer-tennis score. (d) The best predictive path model that describes the relationship between morphology, balance, maximum athletic capacity and motor skill function with overall success in cumulative soccer-tennis score. Morphology 1 is PCM1 and morphology 2 is PCM2. Athleticism 1 is PCATH1 and athleticism 2 is PCATH2. Skill 1 is PCSK1 and Skill 2 is PCSK2.
Skill was the only factor that was strongly associated with a player's performance in 11-a-side soccer matches (figure 2). Our multivariate analysis revealed a significant effect of scores for PCSK1, but no other significant effects (table 2). Our univariate analyses confirmed that skill was associated with activity (electronic supplementary material, table S7) and successes (electronic supplementary material, table S9), but not efficiency (electronic supplementary material, table S11). PCSK1 was included in the most likely models according to the AIC (see the electronic supplementary material, tables S6 and S8). The likelihood that the best model of activity or performance included skill (PCSK1) equalled or exceeded 95% (see estimates of importance in the electronic supplementary material, tables S7 and S9). Specifically, players with higher PCSK1 were more likely to have higher activity and successes in the match.
Figure 2.

Relationship between maximum athletic capacity (PCATH1), motor skill function (PCSK1) and balance with three different measures of player performance from 11-a-side soccer matches. Overall match success was significantly associated with (a) motor skill (PCSK1) but not (b) balance or (c) overall athletic ability (PCATH1). Total individual match activity was significantly associated with (d) PCSK1 but not (e) balance or (f) PCATH1. Individual match efficiency was not significantly associated with (g) PCSK1, (h) balance or (i) PCATH1.
Table 2.
Multivariate analysis of variance (MANOVA) for activity, successes, and efficiency in the 11-a-side soccer matches and their relationship with balance, skill and athleticism. (Parameter estimates are provided in the electronic supplementary material.)
| effect | Pillai's trace | F3,16 | P |
|---|---|---|---|
| balance | 0.074 | 0.424 | 0.738 |
| skill 1 (PCSK1) | 0.503 | 5.390 | 0.009 |
| skill 2 (PCSK2) | 0.155 | 0.976 | 0.428 |
| athleticism 1 (PCATH1) | 0.126 | 0.768 | 0.529 |
| athleticism 2 (PCATH2) | 0.209 | 1.408 | 0.277 |
Based on comparisons between individuals playing in the different levels of competition, we found premier league players had greater overall athletic ability (PCATH1) than reserve players (F1,22 = 5.37, p = 0.030) (table 3). However, there was no difference in either balance (F1,22 = 0.45, p = 0.51) or overall motor skill (PCSK1) (F1,22 = 0.001, p = 0.98) between the premier and reserve players. Player position only had a significant association with balance (F4,19 = 3.85, p = 0.019), with forwards possessing greater balance than central-defenders (p = 0.020) and wide-midfielders (p = 0.039).
Table 3.
Average differences in skill, athleticism and balance between players from the higher premier league grade and the lower reserve team grade. (The only difference between the grades was in overall athleticism (PCATH1). League means with s.e.m.; ANOVA; MS, mean squares value; F, F ratio. Italic value indicates statistical significance.)
| mean |
league effect |
|||||
|---|---|---|---|---|---|---|
| premier | reserve | d.f. | MS | F | P | |
| motor skill 1 (PCSK1) | 0.00 ± 0.44 | 0.00 ± 0.43 | 1,22 | 0.00 | 0.00 | 0.991 |
| motor skill 2 (PCSK2) | 0.14 ± 0.31 | −0.14 ± 0.29 | 1,22 | 0.49 | 0.46 | 0.503 |
| athletic ability 1 (PCATH1) | 0.67 ± 0.46 | −0.67 ± 0.35 | 1,22 | 10.85 | 5.37 | 0.030 |
| athletic ability 2 (PCATH2) | 0.02 ± 0.33 | −0.02 ± 0.30 | 1,22 | 0.01 | 0.01 | 0.916 |
| balance | 0.14 ± 0.30 | −0.14 ± 0.28 | 1,22 | 0.46 | 0.45 | 0.509 |
For each of the network outcomes, total time on the field was the strongest predictor and was significant in each analysis. Players' league was not significantly associated with any network outcomes (table 4). Maximum athletic abilities (PCATH2) predicted in-degree (the number of different players from which an individual received the ball) and total degree, but no other network outcomes. By contrast, PCSK1 significantly predicted all social network metrics except for out-degree, which was the number of different players to which an individual successfully passed the ball (B = 0.06, t15.55 = 0.449, p = 0.660). For in-degree, total degree, weighted in-degree, weighted out-degree, weighted degree, and closeness, PCSK1 significantly predicted higher scores (all p's less than 0.04).
Table 4.
The effects of motor skill (PCSK1), overall athletic ability (PCATH2) and time on field on metrics of social network connectedness calculated on the movement of the ball through 11-a-side soccer matches. (None of the other predictors were found to have a significant effect on the social network traits. Game league was included as a control but was never significant and is not shown here. Significance is taken at the level of p < 0.05 (using Wald tests) and denoted by text in italics. Standard errors are provided.)
| Social network metric | PCSK1 | PCATH2 | field time |
|---|---|---|---|
| number of players passed to (out-degree) | 0.03 ± 0.07 | 0.11 ± 0.06 | 0.69 ± 0.07 |
| number of players received from (in-degree) | 0.17 ± 0.07 | 0.19 ± 0.07 | 0.52 ± 0.06 |
| number of passing partners (overall-degree) | 0.18 ± 0.07 | 0.17 ± 0.07 | 0.70 ± 0.06 |
| number of successful passes made (out-strength) | 0.24 ± 0.08 | 0.07 ± 0.08 | 0.73 ± 0.06 |
| number of successful passes received (in-strength) | 0.32 ± 0.08 | 0.16 ± 0.08 | 0.75 ± 0.06 |
| number of successful passes (overall-strength) | 0.32 ± 0.08 | 0.12 ± 0.08 | 0.79 ± 0.06 |
| closeness (network centrality) | 0.22 ± 0.05 | 0.07 ± 0.05 | 0.72 ± 0.05 |
4. Discussion
Our study is, to our knowledge, the first to quantify how skill, balance and athleticism drive success among individuals in a complex physical activity in any species, and does so by viewing soccer players through the analytical lens of evolutionary biology. We found that individuals with greater skill were more likely to perform well in soccer-tennis games and 11-a-side matches. Furthermore, skill was also the best predictor of an individual's contribution to the success of a team, based on a social network analysis of ball movement.
Although our study shows skill was the best predictor of success in soccer matches, at least in theory this could be because our specific measures of athleticism do not capture variation in overall athleticism and/or are irrelevant to soccer match performance [44,45]. However, this seems unlikely, as the athletic traits we quantified included metrics of agility, speed, acceleration, power and endurance—all of which are generally considered to be relevant to soccer [4,46]. Furthermore, the first principal component explains most of the variation in athleticism and all measures were strongly positively correlated, suggesting it captures meaningful variation.
Most quantitative protocols for talent identification in soccer focus more on the generic athletic attributes of team sports, such as speed, strength, agility and endurance, rather than skill [5,46,47]. In fact, the vast majority of science that occurs on soccer players focuses on how to improve speed, strength and fitness, and how to facilitate rapid recovery after matches or following injury. Quantitative assessments of soccer-specific skill and experimental testing of programmes designed to improve the uptake of skill are almost non-existent in football academies and professional clubs. This is surprising given that soccer is primarily a game that rewards high technical skill and the game's most skilful players attract the highest salaries and are the most revered and coveted. So why is there still no detailed and widely used quantitative metric for soccer-specific skill? Tests of soccer-specific skill are usually more time-intensive and can offer lower repeatabilities than measures of athletic performance [23]. Furthermore, there are also disagreements over which skills are the most relevant [10]. Finally, many coaches still feel that assessment of skill is their domain, and not that of scientists. But despite these potential barriers, the development of a comprehensive protocol for soccer-specific skill could markedly improve both talent identification and development [34]. Our study has shown that the best players in matches also have the higher skill, yet the importance of skill to match success is currently not reflected in soccer science.
Although players from the premier team were more athletic than those in the reserve team, athleticism failed to predict which players performed best within each grade, suggesting that grades may reflect only a minimal level of athleticism required for entry or differences in training load. Traditionally, scouting and training programmes have relied heavily on those athletic traits that differ between elite and sub-elite grades, while overlooking variation within each grade. These same programmes ignore the complexity of performance in dynamic environments, including competitors, teammates and a ball. This may also help to explain why there has been a limited uptake of quantitative protocols for talent identification across most professional team sports, especially soccer [5,12]. Wild animals modulate their movements to account for their environment—for example, slowing along thinner branches [48] or when turning sharply [25]. Similarly, in a soccer game, players must respond to a variety of dynamic environmental conditions through motor performance [30], such as sprinting, dribbling, heading, passing, pivoting, feinting, shooting and even diving [32]; these performances may interact with each other in complex ways. Advances in sports science require that performances are compared among individuals within a grade in similar ways as when evolutionary biologists compare performance among individuals in a natural population.
Evolutionary biologists also have much to gain by crossing taxonomic divides. Although it's easier to measure sprint speeds than motor skill in almost any animal species, including humans, such measures of maximal performance alone are simply not enough to predict success during a complex activity. For example, sprint speed is widely cited as important for escaping predators [16,22], but there are as many studies that report no relationship between speed and survival as there are that report a positive relationship [22]. Apparently, speed is just one trait that can determine escape from predators and in certain environments or situations, other traits will be more important. Because almost all predators are faster than their prey, escape probably depends on acceleration and agility, as well as speed. An adaptive landscape may have multiple optima for escape strategies, based on different combinations of phenotypes. For example, an individual that is fast but not agile may be as successful as one that is slow but agile [49]. Hence, the future of performance research relies on integrating biomechanics, performance, and behaviour in a multi-dimensional phenotypic space. By quantifying multiple performances simultaneously, one can see whether multiple phenotypes can lead to similar success. The benefit of working with humans is that they (often) behave as instructed, which helps researchers test theoretical models and refine experimental protocols (e.g. [49]). The knowledge gained from studies of humans can subsequently be extended to non-human animals in uncontrolled environments.
Here we have argued that analyses of sporting data and human physical function can be used to address an enormous range of evolutionary topics that are difficult to address in nonhuman models. Such topics include deception [32], senescence [33] and functional trade-offs [29,30]. Similarly, approaches to data analysis used by evolutionary biologists to study adaptation, multivariate trait evolution and adaptive landscapes can also contribute to advances in sports science. Sports scientists might better accomplish their aims—identifying talented players, nurturing young athletes, and managing and estimating the rates of senescence of individual performance [50]—using methods from evolutionary biology. Both fields would benefit from changing current mindsets that hold back collaborations between disciplines.
Humans are governed by the same musculoskeletal constraints as other terrestrial vertebrates; therefore, analyses of human function and behaviour can address fundamental questions in biology. Although sports reflect cultural effects imposed by social environments, they also reflect the human desire to compete in ritualized contests that advertise the quality of a genotype. Thus, our path analysis of morphology, skill and performance could be extended to examine the effects on mating success. Such an extension would bring the power of Arnold's paradigm for studying selection gradients to bear on competitive behaviours that occur throughout human populations [9]. In light of this potential, future collaborations between evolutionary biologists and sports scientists should promote an exciting new field of evolutionary sports science.
Supplementary Material
Supplementary Material
Acknowledgements
We thank anonymous reviewers for helpful comments on the manuscript. We thank players of University of Queensland Football Club and Glen Volker.
Ethics
All data were collected in accordance with the University of Queensland Ethics committee for experimental research.
Data accessibility
The data used for this study have been deposited in Dryad http://dx.doi.org/10.5061/dryad.16vd4 [51].
Authors' contributions
R.S.W., G.K.D., A.H.H. and M.D.S. conceived study and collected data. R.S.W., G.K.D., S.C.M., M.J.A. and A.C.N. analysed data. All authors were involved in interpreting the results and contributed to writing the manuscript.
Competing interests
We have no competing interests.
Funding
R.S.W. is supported by an Australian Research Council fellowship (FT150100492).
References
- 1.Balish SM, Eys MA, Schulte-Hostedde AI. 2013. Evolutionary sport and exercise psychology: integrating proximate and ultimate explanations. Psychol. Sport Exerc. 14, 413–422. ( 10.1016/j.psychsport.2012.12.006) [DOI] [Google Scholar]
- 2.Deaner RO, Smith BA. 2012. Sex differences in sport across 50 societies. Cross Cult. Res. 47, 268–309. ( 10.1177/1069397112463687) [DOI] [Google Scholar]
- 3.De Block A, Dewitte S. 2009. Darwinism and the cultural evolution of sports. Perspect. Biol. Med. 52, 1–16. ( 10.1353/pbm.0.0063) [DOI] [PubMed] [Google Scholar]
- 4.Vaeyens R, Malina RM, Janssens M, Van Renterghem B, Bourgois J, Vrijens J, Philippaerts RM. 2006. A multidisciplinary selection model for youth soccer: the Ghent Youth Soccer Project. Br. J. Sports Med. 40, 928–934. ( 10.1136/Bjsm.2006.029652) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Reilly T, Williams AM, Nevill A, Frank A. 2000. A multidisciplinary approach to talent identification in soccer. J. Sports Sci. 18, 695–702. ( 10.1080/02640410050120078) [DOI] [PubMed] [Google Scholar]
- 6.Bartholomew GA. 1966. Interaction of physiology and behavior under natural conditions. In The Galapagos (ed. Bowman RI.), pp. 39–45. Berkeley, CA: University of California Press. [Google Scholar]
- 7.Bennett AF, Huey RB. 1990. Studying the evolution of physical performance. Oxf. Surv. Evol. Biol. 7, 251–284. [Google Scholar]
- 8.Garland T Jr, Hankins E, Huey RB. 1990. Locomotor capacity and social dominance in male lizards. Funct. Ecol. 4, 243–250. ( 10.2307/2389343) [DOI] [Google Scholar]
- 9.Arnold SJ. 1983. Morphology, performance and fitness. Am. Zool. 23, 347–361. ( 10.1093/icb/23.2.347) [DOI] [Google Scholar]
- 10.Ali A. 2011. Measuring soccer skill performance: a review. Scand. J. Med. Sci. Sports 21, 170–183. ( 10.1111/J.1600-0838.2010.01256.X) [DOI] [PubMed] [Google Scholar]
- 11.Unnithan V, White J, Georgiou A, Iga J, Drust B. 2012. Talent identification in youth soccer. J. Sports Sci. 30, 1719–1726. ( 10.1080/02640414.2012.731515) [DOI] [PubMed] [Google Scholar]
- 12.Carling C, le Gall F, Reilly T, Williams AM. 2009. Do anthropometric and fitness characteristics vary according to birth date distribution in elite youth academy soccer players? Scand. J. Med. Sci. Sports 19, 3–9. ( 10.1111/J.1600-0838.2008.00867.X) [DOI] [PubMed] [Google Scholar]
- 13.Wilson RS, James RS, David G, Hermann E, Morgan OJ, Niehaus AC, Hunter A, Smith M. 2016. Multivariate analyses of individual variation in soccer skill as a tool for talent identification and development: utilizing evolutionary theory in sports science. J. Sports Sci. 34, 2074–2086. ( 10.1080/02640414.2016.1151544) [DOI] [PubMed] [Google Scholar]
- 14.Endler JA. 1986. Natural selection in the wild. Princeton, NJ: Princeton University Press. [Google Scholar]
- 15.Kingsolver JG, Hoekstra HE, Hoekstra JM, Berrigan D, Vignieri SN, Hill CE, Hoang A, Gibert P, Beerli P. 2001. The strength of phenotypic selection in natural populations. Am. Nat. 157, 245–261. ( 10.1086/319193) [DOI] [PubMed] [Google Scholar]
- 16.Jayne BC, Bennet AF. 1990. Selection on locomotor performance capacity in a natural population of garter snakes. Evolution 44, 1204–1229. ( 10.2307/2409283) [DOI] [PubMed] [Google Scholar]
- 17.Arnott G, Elwood RW. 2010. Signal residuals and hermit crab displays: flaunt it if you have it! Anim. Behav. 79, 137–143. ( 10.1016/j.anbehav.2009.10.011) [DOI] [Google Scholar]
- 18.Huyghe K, Vanhooydonck B, Scheers H, Molina-Borja M, Van Damme R. 2005. Morphology, performance and fighting capacity in male lizards, Gallotia galloti. Funct. Ecol. 19, 800–807. ( 10.1111/j.1365-2435.2005.01038.x) [DOI] [Google Scholar]
- 19.Wilson RS, Angilletta MJ, James RS, Navas C, Seebacher F. 2007. Dishonest signals of strength in male slender crayfish (Cherax dispar) during agonistic interactions. Am. Nat. 170, 284–291. ( 10.1086/519399) [DOI] [PubMed] [Google Scholar]
- 20.Wilson RS, Hammill E, Johnston IA. 2007. Competition moderates the benefits of thermal acclimation to reproductive performance in male eastern mosquitofish. Proc. R. Soc. B 274, 1199–1204. ( 10.1098/Rspb.2006.0401) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Seebacher F, Wilson RS. 2006. Fighting fit: thermal plasticity of metabolic function and fighting success in the crayfish Cherax destructor. Funct. Ecol. 20, 1045–1053. ( 10.1111/j.1365-2435.2006.01194.x) [DOI] [Google Scholar]
- 22.Irschick DJ, Meyers JJ, Husak JF, Le Galliard JF. 2008. How does selection operate on whole-organism functional performance capacities? A review and synthesis. Evol. Ecol. Res. 10, 177–196. [Google Scholar]
- 23.Wilson RS, Husak JF, Halsey L, Clemente CJ. 2015. Predicting the movement speeds of animals in natural environments. Integr. Comp. Biol. 55, 1125–1141 ( 10.1093/icb/icv106) [DOI] [PubMed] [Google Scholar]
- 24.Barske J, Schlinger BA, Wikelski M, Fusani L. 2011. Female choice for male motor skills. Proc. R. Soc. B 278, 3523–3528. ( 10.1098/rspb.2011.0382) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wynn ML, Clemente C, Amir Abdul Nasir AF, Wilson RS. 2015. Running faster causes disaster: trade-offs between speed, manoeuvrability and motor control when running around corners in northern quolls (Dasyurus hallucatus). J. Exp. Biol. 218, 433–439. [DOI] [PubMed] [Google Scholar]
- 26.Amir Abdul Nasir AA, Clemente C, Wynn M, Wilson RS. 2017. Optimal running speeds when there is a trade-off between speed and the probability of mistakes. Funct. Ecol. 31, 1941–1949. ( 10.1111/1365-2435.12902) [DOI] [Google Scholar]
- 27.Briffa M, Lane SM. 2017. The role of skill in animal contests: a neglected component of fighting ability. Proc. R. Soc. B 284, 20171596 ( 10.1098/rspb.2017.1596) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Clemente CJ, Wilson RS. 2015. Balancing biomechanical constraints: optimal escape speeds when there is a trade-off between speed and manoeuvrability. Integr. Comp. Biol. 55, 1142–1154. ( 10.1093/icb/icv103) [DOI] [PubMed] [Google Scholar]
- 29.Van Damme R, Wilson RS, Van Hooydonck B, Aerts P. 2002. Performance constraints in decathletes. Nature 415, 755–756. ( 10.1038/415755b) [DOI] [PubMed] [Google Scholar]
- 30.Wilson RS, Niehaus AC, David G, Hunter A, Smith M. 2014. Does individual quality mask the detection of performance trade-offs? A test using analyses of human physical performance. J. Exp. Biol. 217, 545–551. ( 10.1242/Jeb.092056) [DOI] [PubMed] [Google Scholar]
- 31.Careau V, Wilson RS. 2017. Performance trade-offs and ageing in the “world's greatest athletes”. Proc. R. Soc. B 284, 20171048 ( 10.1098/rspb.2017.1048) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.David GK, Condon CK, Bywater CL, Ortiz-Barrientos D, Wilson RS. 2012. Receivers limit the prevalence of deception in humans: evidence from diving behaviour in soccer players. PLoS ONE 6, e26017 ( 10.1371/journal.pone.0026017) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lailvaux S, Wilson RS, Kasumovic M. 2014. Trait compensation and sex-specific aging of performance in male and female professional basketball players. Evolution 68, 1523–1532. ( 10.1111/evo.12375) [DOI] [PubMed] [Google Scholar]
- 34.Honer O, Votteler A, Schmid M, Schultz F, Roth K. 2015. Psychometric properties of the motor diagnostics in the German football talent identification and development programme. J. Sports Sci. 33, 145–159. ( 10.1080/02640414.2014.928416) [DOI] [PubMed] [Google Scholar]
- 35.Stuart-Fox D. 2006. Testing game theory models: fighting ability and decision rules in chameleon contests. Proc. R. Soc. B 273, 1555–1561. ( 10.1098/rspb.2006.3468) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Csardi G, Nepusz T.2006. The igraph software package for complex network research, inter journal complex systems 1695. http://igraph.org .
- 37.R Core Team 2017. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing; https://www.R-project.org/. [Google Scholar]
- 38.Marsden P. 1990. Network data and measurement. Annu. Rev. Sociol. 16, 435–463. ( 10.1146/annurev.so.16.080190.002251) [DOI] [Google Scholar]
- 39.Scott JP, Carrington PJ. 2011. The SAGE handbook of social network analysis. Thousand Oaks, CA: Sage. [Google Scholar]
- 40.Freeman LC. 1979. Centrality in social networks: conceptual clarification. Soc. Networks 1, 215–239. ( 10.1016/0378-8733(78)90021-7) [DOI] [Google Scholar]
- 41.Zuur AF, Ieno EN, Walker N, Saveliev AA, Smith GM.2009. Mixed effects models and extensions in ecology with R. New York, NY: Springer.
- 42.Burnham KP, Anderson DR.2002. Model selection and multimodel inference: a practical information–theoretic approach. New York, NY: Springer.
- 43.Bartoń K. 2013 MuMIn: multi-model inference, R package version 1.9.13. [Google Scholar]
- 44.Bangsbo J. 2006. Physical and metabolic demands of training and match-play in the elite footballer. J. Sports Sci. 24, 665–674. ( 10.1080/02640410500482529) [DOI] [PubMed] [Google Scholar]
- 45.Rampinini E, Coutts AJ, Castagna C, Sassi R, Impellizzeri FM. 2007. Variation in top level soccer match performance. Int. J. Sports Med. 28, 1018–1024. ( 10.1055/s-2007-965158) [DOI] [PubMed] [Google Scholar]
- 46.Vaeyens R, Lenoir M, Williams AM, Philippaerts RM. 2008. Talent identification and development programmes in sport—current models and future directions. Sports Med. 38, 703–714. ( 10.2165/00007256-200838090-00001) [DOI] [PubMed] [Google Scholar]
- 47.le Gall F, Carling C, Williams M, Reilly T. 2010. Anthropometric and fitness characteristics of international, professional and amateur male graduate soccer players from an elite youth academy. J. Sci. Med. Sport 13, 90–95. ( 10.1016/J.Jsams.2008.07.004) [DOI] [PubMed] [Google Scholar]
- 48.Wheatley R, Angilletta MJ, Niehaus AC, Wilson RS. 2015. How fast should an animal run when escaping? An optimality model based on the trade-off between speed and accuracy. Integr. Comp. Biol. 55, 1166–1175. [DOI] [PubMed] [Google Scholar]
- 49.Clemente CJ, Wilson RS. 2015. Balancing biomechanical constraints: optimal escape speeds when there is a trade-off between speed and manoeuvrability. Integr. Comp. Biol. 55, 1142–1154. [DOI] [PubMed] [Google Scholar]
- 50.Sewell D, Watkins P, Griffin M. 2012. Sport and exercise science. New York, NY: Routledge. [Google Scholar]
- 51.Wilson RS, David GK, Murphy SC, Angilletta MJ, Jr, Niehaus AC, Hunter AH, Smith MD.2017. Data from: Skill not athleticism predicts individual variation in match performance of soccer players. Dryad Digital Repository. ( ) [DOI] [PMC free article] [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Citations
- Wilson RS, David GK, Murphy SC, Angilletta MJ, Jr, Niehaus AC, Hunter AH, Smith MD.2017. Data from: Skill not athleticism predicts individual variation in match performance of soccer players. Dryad Digital Repository. ( ) [DOI] [PMC free article] [PubMed]
Supplementary Materials
Data Availability Statement
The data used for this study have been deposited in Dryad http://dx.doi.org/10.5061/dryad.16vd4 [51].