

Summary
Membraneless RNA granules originate via phase separation events driven by multivalent interactions. As RNA is the defining component of such granules, we examined how RNA contributes to granule assembly. Expansion of hexanucleotide GGGGCC (G4C2) repeats in the first intron of C9ORF72 is the most common genetic cause of amyotrophic lateral sclerosis and frontotemporal dementia (C9-ALS/FTD). We describe a biophysical phenomenon whereby G4C2 RNA (rG4C2) promotes the phase separation of RNA granule proteins in vitro and in cells. The ability of rG4C2 to promote phase separation is dependent on repeat length and RNA structure, as rG4C2 must assume a G-quadruplex conformation to promote granule assembly. We demonstrate a central role for RNA in promoting phase separations and implicate rG4C2 G-quadruplex structures in the pathogenesis of C9-ALS/FTD.
Keywords: Phase separation, G-quadruplex, C9ORF72, amyotrophic lateral sclerosis, frontotemporal dementia, RNA, stress granule
eTOC Blurb
Fay el al. report that rG4C2 G-quadruplexes cause the assembly of RNA granules in vitro and in cells. RNA structure impacts RNA granule assembly and increased GGGGCC length as seen in patients with C9ORF72-ALS/FTD causes more condensation of RNA granules, indicating RNAG-quadruplexes is a plausible C9ORF72-ALS/FTD drug target.
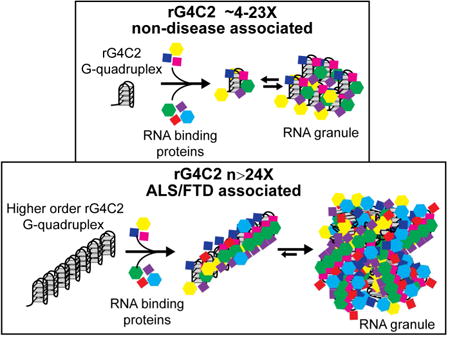
Introduction
Protein-protein interactions are proposed to play a dominant role in RNA granule dynamics (Mitrea and Kriwacki, 2016); changes affecting such interactions (e.g., mutations in selected protein factors) are linked to the formation of protein aggregates and defects in RNA granule homeostasis are implicated in neurodegenerative disease pathogenesis (Bosco et al., 2011; Burke et al., 2015; Conicella et al., 2016; Kim et al., 2013; Molliex et al., 2015; Murakami et al., 2015; Patel et al., 2015). A common feature of non-coding repeat expansion disorders is the accumulation of repeat-bearing RNA foci in the nucleus and/or cytoplasm of affected cells (La Spada and Taylor, 2010). How these RNA foci form and how they impact biological processes and contribute to disease is largely unknown.
A hexanucleotide expansion GGGGCC (G4C2), in the first intron of the chromosome 9 open reading frame 72 (C9ORF72) gene, is the most common mutation associated with amyotrophic lateral sclerosis and frontotemporal dementia (C9-ALS/FTD) (DeJesus-Hernandez et al., 2011; Renton et al., 2011). G4C2 repeats exceeding a defined threshold (typically ≥24 repeats) show association with ALS/FTD (Van Mossevelde et al., 2017). RNA is transcribed from these repeats in the sense (rG4C2) and antisense (CCCCGG: rC4G2) directions. Dipeptide repeat proteins (DRP) are translated from rG4C2 and rC4G2 repeats via Repeat Associated non-ATG (RAN) translation. Both rG4C2 and DRP form nuclear and cytoplasmic foci when transfected into cells as well as in C9-ALS/FTD patient samples. These foci disrupt RNA metabolism by a variety of mechanisms (Gitler and Tsuiji, 2016). Some rG4C2 cytoplasmic foci resemble stress granules (SGs), which are membraneless, microscopically-visible cytoplasmic foci induced by some stresses and containing translationally stalled mRNAs, 40S ribosomal subunits, RNA binding proteins (RBPs), and signaling molecules (Kedersha et al., 2013). Both rG4C2 and DRP are known to disrupt RNA metabolism, alter SG dynamics and may directly drive disease pathogenesis (Gitler and Tsuiji, 2016).
Recent reports suggest that RNA granule formation is dependent on weak interactions between prion-like, low complexity (including DRP) and intrinsically disordered regions in proteins that facilitate phase separation, a biophysical phenomenon that allows concentration of proteins into discrete non-membranous subcellular compartments (reviewed in (Hyman et al., 2014; Mitrea and Kriwacki, 2016)). The chemical biotinylated isoxazole (b-isox) can promote phase separation in vitro by concentrating and condensing low complexity domain- and intrinsically disordered region-containing proteins into structures that resemble RNA-granules (Han et al., 2012; Kato et al., 2012). Multiple RBPs contain distinct RNA-binding domains and intrinsically disordered regions, and these proteins can undergo phase transitions such as formation of liquid droplets, hydrogels or even insoluble amyloids in vitro (Burke et al., 2015; Conicella et al., 2016; Lin et al., 2015; Molliex et al., 2015; Murakami et al., 2015; Patel et al., 2015; Zhang et al., 2015). Addition of RNA to such protein-driven phase separations influences their ability to separate (Elbaum-Garfinkle et al., 2015; Lin et al., 2015; Molliex et al., 2015; Zhang et al., 2015). Additionally, repeat expansion RNA can promote an in vitro phase separation in isolation (Jain and Vale, 2017).
rG4C2 forms higher order structures including G-quadruplexes and hairpins in vitro (Fratta et al., 2012; Haeusler et al., 2014; Reddy et al., 2013; Su et al., 2014). G-quadruplexes are stable structures formed by stacks of four guanosine residues hydrogen bonded via Hoogsteen base pairing and stabilized by specific metal cations (Neidle, 2012). The availability of structural data combined with the link to neurodegenerative disease makes rG4C2 an ideal candidate to study the mechanisms by which specific RNAs modulate RNA granule assembly. In analogy to SG nucleator proteins that initiate SG formation when over-expressed in the absence of stress (Kedersha et al., 2013), we hypothesized that rG4C2 G-quadruplexes work as RNA templates that create a local microenvironment that drives RNA granule formation.
We show here that rG4C2 in a G-quadruplex conformation drives the assembly of RNA granules both in vitro and in cells. In vitro, rG4C2 condenses a cohort of proteins that is similar but not identical those condensed by b-isox. Importantly, rG4C2 repeat length correlates with the degree of protein condensation in vitro and in cells, mirroring the relationship between C9ORF72 repeat length and ALS/FTD pathogenesis. Our studies offer a framework for understanding how RNA participates in RNA granule assembly, and suggest ways in which targeting the rG4C2 G-quadruplex might be beneficial in the treatment of patients with ALS/FTD.
Results
rG4C2 promotes SGs in cells
We have previously shown that transfection of various synthetic and natural RNA species promotes SG formation (Emara et al., 2010; Ivanov et al., 2014). Interestingly, transfection of r(G4C2)4 but not r(C4G2)4 also promotes formation of cytoplasmic foci positive for G3BP1, a standard SG marker (Fig S1A-C). These r(G4C2)4-induced cytoplasmic foci also contain the SG markers eIF3b, eIF4G, and poly(A) mRNA (Fig S1A-B). As the length of G4C2 repeats is linked to C9-ALS/FTD pathogenesis (DeJesus-Hernandez et al., 2011; Renton et al., 2011) and some C9-ALS/FTD patient cells are positive for rG4C2 cytoplasmic foci (Donnelly et al., 2013), we tested whether variable length rG4C2 repeats differently affect granule formation. Transfection of in vitro transcribed rG4C2 into U2OS cells promotes G3BP1-positive granules in a repeat length dependent manner (Fig 1A-B). To determine whether repeat length alters SG properties, we assessed the number of SGs per cell and found that cells transfected with longer repeats showed a decrease in SGs per cell (Fig 1C). In assessing SG size, we observed no difference (Fig 1D).
Figure 1. rG4C2 promotes SG formation.
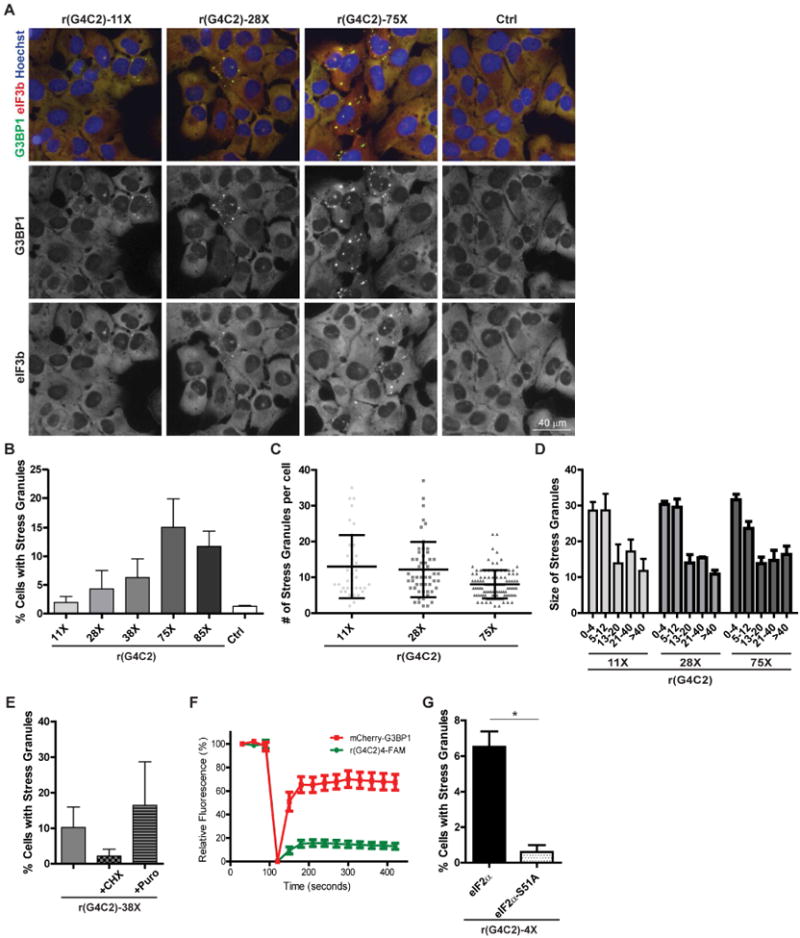
(A) U2OS cells were transiently transfected with equimolar in vitro transcribed rG4C2 of indicated length or vehicle control (Ctrl) for 6 hours then fixed, permeabilized and assayed by immunofluorescence detecting G3BP1 (green) and eIF3b (red) and counterstained with Hoechst (blue) to visualize nuclei. Scale bar is 40 μm. (B-D) Quantification of (A) as the percentage of cells with SGs, (C) number of SGs per cell, and (D) SG size. (E) U2OS cells transiently transfected with rG4C2-38× then treated with puromycin (puro, 5 μg/mL) or cycloheximide (CHX,10 μg/mL) for 30 min prior to fixation. Quantification of the percentage of cells with SG (cells with ≥2 G3BP1 positive foci). (F) FRAP analysis of rG4C2-induced SGs in U2OS cells stably expressing mCherry-G3BP1 and transiently transfected with 3′FAM labeled r(G4C2)4. Graphs represent relative fluorescence of mCherry-G3BP1 (red) and 3′FAM labeled r(G4C2)4 (green) before (first 3 data points) and after photobleaching (last 10 data points). (G) Quantification of the percentage of SG positive wildtype (black bars) or eIF2α-S51A mutant (white with black dots) MEFs transfected with r(G4C2)4. * denotes statistical significance, p<0.05. (B, E-G) Data are represented as mean ± standard deviation (SD), n≥3. (C) >37 cells and (D) >50 foci were quantified per condition across 3 biological repeats. See also Fig S1, S2.
Since rG4C2-induced granules contain known SG markers, we asked whether these granules are in dynamic equilibrium with polysomes by treating with cycloheximide and puromycin, drugs that inhibit or promote SG assembly, respectively. rG4C2-38X (38 repeats of the G4C2) induced granules decrease when treated with cycloheximide and increase when treated with puromycin (Fig 1E), consistent with these granules being in dynamic equilibrium with polysomes (Kedersha et al., 2000). The presence of SG markers and their dynamic behavior indicates rG4C2-induced foci can be classified as bona fide SGs (Aulas et al., 2017). Fluorescence recovery after photobleaching (FRAP) was employed to assess the dynamics of both the r(G4C2)4 and mCherry-tagged G3BP1. The fluorescent signal from transiently transfected 3′FAM labeled r(G4C2)4 is static and does not return after bleaching as indicated by only 15% recovery after photobleaching, whereas mCherry-tagged G3BP1 exhibits dynamic behavior quickly returning to the bleached region with 70% of the signal recovering (Fig 1F, S2).
SGs form due to a block in translation initiation which can occur though several mechanisms. One such mechanism is via phosphorylation of eIF2α on serine 51. eIF2α brings the initiator methionine tRNA to the 43S initiation complex, and phosphorylation effectively prevents initiator tRNA recruitment and, consequently, inhibits translation. To assess whether eIF2α phosphorylation is required for rG4C2 mediated SG assembly, we utilized a mouse embryonic fibroblasts (MEFs) with a serine 51 to alanine point mutation in eIF2α (S51A). Transfection of r(G4C2)4 promotes SG assembly in wildtype MEFs but not in MEFs containing the eIF2α-S51A mutation (Fig 1G), indicating eIF2α phosphorylation is required for rG4C2 induced SG assembly.
Together, these data indicate rG4C2 promotes SGs in a length dependent manner, and rG4C2 is a stable component of these granules. Importantly, transfection of plasmids used for in vitro transcription encoding variable-length G4C2 repeats also promotes formation of rG4C2-containing nuclear foci (Fig S1D), consistent with other investigations (DeJesus-Hernandez et al., 2011). These data also indicate that rG4C2 is sufficient to promote RNA granules both in nuclear and cytoplasmic compartments.
rG4C2 mediated condensation promotes in vitro RNA granule assembly
rG4C2 can promote SGs and nuclear foci (Fig 1, S1). Formation of RNA granules has been mechanistically linked to phase separation, a process whereby demixing high concentrations of macromolecules in crowded microenvironments results in the assembly of non-membrane-bound entities (Hyman et al., 2014; Mitrea and Kriwacki, 2016). As b-isox assembles RNA granule-like structures in cell lysates (Kato et al., 2012), we hypothesized that rG4C2 might promote a phase separation in vitro. To test this, we developed a visual assay to assess phase separation. Antisense r(C4G2)4 oligo, an oligo substituting the purine for purine (adenosine for guanosine) r(A4C2)4 oligo, and a scrambled control (rScram) oligo that is a scrambled version of r(G4C2)4 with the same guanosine content as r(G4C2)4, were used as controls. Briefly, 3′ fluorescently labeled r(G4C2)4, r(C4G2)4, r(A4C2)4, or rScram were incubated with U2OS lysate, concentrated by centrifugation, spotted on a slide and then examined using fluorescence microscopy. Fluorescently-labeled r(G4C2)4 promotes microscopically visible foci in cell lysates, but not in buffer controls (Fig 2A, left panel). These foci form in a concentration-dependent manner (Fig 2B). rScram also forms microscopically visible foci in cell lysate yet to a lesser extent than r(G4C2)4 (Fig 2A-B). Neither r(A4C2)4 nor r(C4G2)4 form foci even at higher concentrations (Fig 2A-B).
Figure 2. rG4C2 promotes foci formation in vitro.
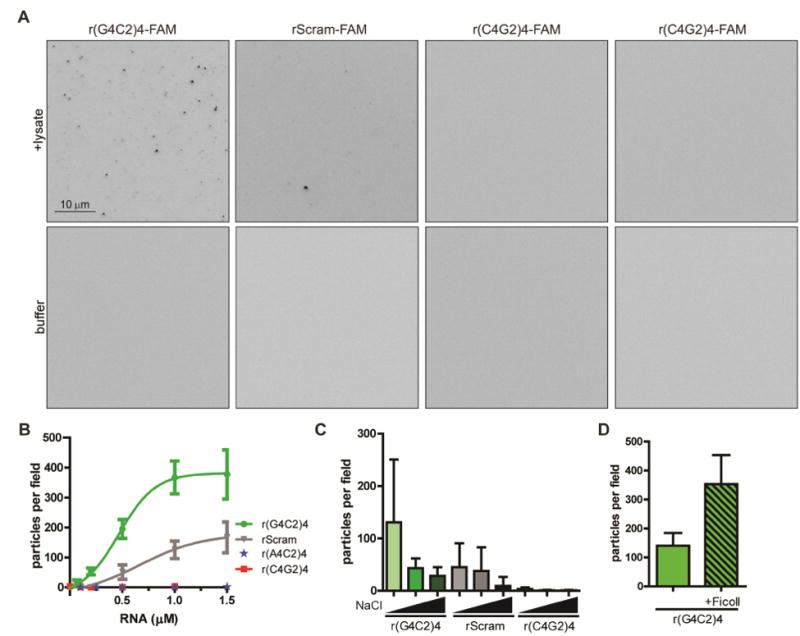
(A) Images detecting pelleted 3′FAM labeled r(G4C2)4, r(C4G2)4, r(A4C2)4, or scrambled control (rScram) (0.5 μM) after incubation with U2OS lysate (upper panels) or buffer (lower panels), concentrated by centrifugation, then assayed by fluorescence microscopy. Scale bar is 10 μm. (B-D) Quantification of assay described in A using r(G4C2)4 (green), r(C4G2)4 (red), r(A4C2)4 (yellow), or rScram (grey) at indicated RNA concentration, (C) 0.25 μM RNA with 75, 150, 200 mM NaCl (labeled left to right), or (D) r(G4C2)4 (0.5 μM) with or without 2.5% Ficoll. (B-D) Data are represented as mean ± standard deviation, n≥3. Also see Fig S3.
To assess the properties of these foci, we assess whether electrostatic interactions play a role in their assembly. Under hypotonic conditions, more r(G4C2)4 foci form while hypertonic conditions prevent foci formation (Fig 2C). Similar trends were observed with rScram and r(C4G2)4 (Fig 2C), indicating that RNA foci formation is more prominent under hypotonic conditions yet the extent is RNA specific . We also assessed whether molecular crowding affects r(G4C2)4 foci formation, and found that molecular crowding using Ficoll promotes r(G4C2)4 foci formation (Fig 2D). Together these data indicate that molecular crowding and electrostatic interactions drive RNA foci formation.
At high concentrations, poly-guanosine, but not other poly-nucleosides, can assume a gel-like state (Bang, 1910). Visual inspection of the r(G4C2)4 condensed material revealed a transparent pellet, in contrast to b-isox, which forms a white opaque pellet of similar size. In contrast to b-isox, r(G4C2)4 requires cell lysate in order to assume a sedimentable state as it does not sediment in buffer alone (Fig S3), suggesting that cellular components are required for in vitro r(G4C2)4 foci formation.
Material condensed by r(G4C2)4 appears to be specific, as shown by differences in condensed versus total banding pattern identified on a silver stained SDS-polyacrylamide gel (Fig 3A, compare lane 1 and 6). Proteins condensed by b-isox and r(G4C2)4 were extracted and assayed by mass spectrometry. Using a cutoff of ≥10 peptides, 333 proteins were condensed by r(G4C2)4 as compared with 196 condensed by b-isox (Fig 3B). 19 of the top 25 r(G4C2)4 identified proteins/protein complexes have been previously identified as RNA granule components (Fig 3C, (Kato et al., 2012; Lessel et al., 2017; Wen et al., 2010)). Gene ontology term analysis of r(G4C2)4 condensed proteins indicates an enrichment in RNA processing, (Fig 3D). Together, this supports that we are observing an in vitro formed granule promoted by RNA. Western blotting was used to confirm the mass spectrometry identified proteins. In common with b-isox, r(G4C2)4 condenses the large and small ribosomal subunits, select RBPs including YB-1 and HuR, some translation initiation factors, and components that regulate SG assembly including G3BP1, Caprin1, and USP10 (Fig 3E). Consistent with the mass spectroscopy results, and in contrast to b-isox, RNA helicases including DDX3 and DDX6, the RNA binding protein TIAR, and the ALS-associated proteins TDP-43 and Ataxin-2 are largely not condensed by r(G4C2)4 (Fig 3E). These data indicate that r(G4C2)4 and b-isox condense both overlapping and unique subsets of proteins.
Figure 3. Selective condensation of RNA granule-related proteins by rG4C2.
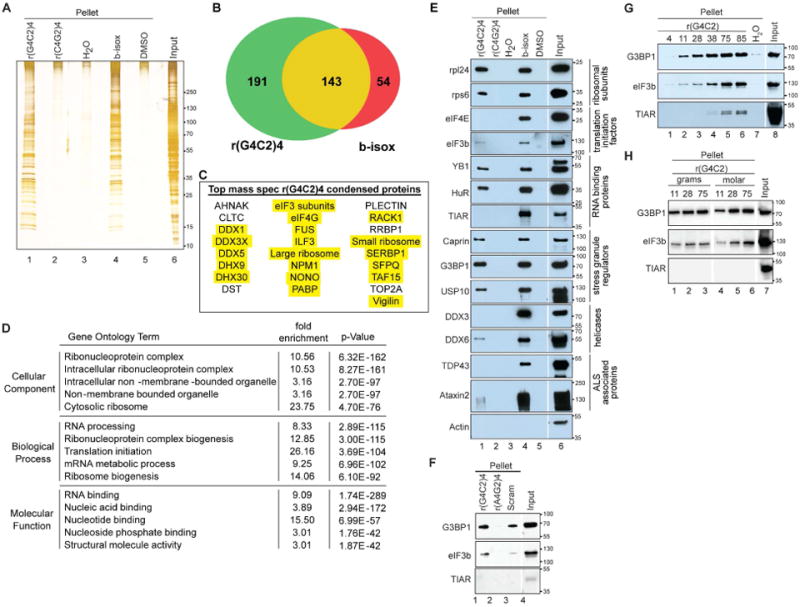
(A) U2OS lysate incubated with indicated RNAs, b-isox, or vehicle controls (H2O and DMSO), pelleted, resuspended and resolved by 4-20% SDS-PAGE then silver stained. (B) Venn diagram of proteins identified by mass spectroscopy that were condensed by r(G4C2)4 and b-isox. Number of proteins identified to be unique for rG4C2 (green), b-isox (red) or overlapping (yellow) is shown. Cut offs set at ≥10 total peptides. (C) Top r(G4C2)4 condensed targets identified by mass spectroscopy, proteins previously identified as components of RNA granules are highlighted in yellow. (D) Gene ontology (GO) analysis of proteins identified by mass spectroscopy that were pelleted by r(G4C2)4, cut offs were set at ≥10 total peptides. Top hits sorted by p-value with a fold enrichment of >3. (E) Proteins pelleted by indicated RNAs, b-isox, or vehicle controls (H2O, DMSO) after incubation with U2OS lysate for 60 min at 4°C, washed twice, and resolved on 4-20% SDS-PAGE gels. Western blots probed for indicated antibodies. (F-H) Western blots probed for G3BP1, eIF3b, and TIAR, same reaction as (E) except (F) using indicated RNAs, (G) equimolar amounts of in vitro transcribed RNAs of indicated length, (H) equal μg (lanes 1-3) compared to equimolar (lanes 4-6) amounts of in vitro transcribed RNAs of indicated length. Cropped blots are indicated by white separation. Also see Fig S4 and Table
As r(G4C2)4 forms RNA granules in vitro, we sought to confirm this phenomenon is universal and not cell type specific. r(G4C2)4-induced condensation of eIF3b, G3BP1, and/or HuR occurred in lysates generated from several different cell lines, as well as from mouse brain (Fig S4). SG components reliably condense G3BP1 and eIF3b with r(G4C2)4 and b-isox, whereas TIAR reliably condenses with b-isox and under some specific conditions with r(G4C2)4 (discussed below). We therefore tracked these proteins for the remainder of our studies. To further validate our condensation assay, we assessed r(A4C2)4 and rScram controls. We found r(A4C2)4 does not promote condensation while rScram partially promotes condensation yet not to the same degree as r(G4C2)4 (Fig 3F).
G4C2 repeat length is a defining feature of C9-ALS/FTD pathogenesis (DeJesus-Hernandez et al., 2011; Renton et al., 2011). To determine whether rG4C2-mediated in vitro condensation demonstrates repeat length-dependent variations, a range of G4C2 repeat lengths were in vitro transcribed and used in equal molar amounts in our condensation assay (Fig 3G). Generally, longer rG4C2 repeats promote more efficient condensation as judged by G3BP1 and eIF3b. Importantly, longer (but not shorter) repeats recruit TIAR, an important SG nucleator (Gilks et al., 2004), suggesting that repeat length also influences the repertoire of proteins condensed. Importantly, when this assay is performed with equal grams of G4C2 11×, 28× and 75× in vitro transcribed RNA, similar amounts of condensation are observed (Fig 3H). This experiment shows that the number of repeats is important yet these repeats can be in cis or in trans to promote this effect.
rG4C2-mediated phase separations
Phase transitions are reversible and influenced by temperature and concentration. Increased temperature causes an increase in free energy that promotes mixing and prevents phase separation. Conversely, increased concentration of initiating factors enhances the transition and thus condensation (Hyman et al., 2014; Mitrea and Kriwacki, 2016). To assess whether temperature affects rG4C2-mediated condensation, r(G4C2)4 condensation reactions were pelleted then washed at 4°C, room temperature (∼20°C), or 37°C. Increased temperature prevents both r(G4C2)4 and b-isox-mediated condensation (Fig 4A, compare lanes 1-2 to 3-4 to 5-6). Conversely, increasing rG4C2 or b-isox concentration causes increased condensation of G3BP1, eIF3b, and eventually condenses TIAR at higher concentrations of r(G4C2)4 (Fig 4B). These data are consistent with the visual assay which shows that assembly of r(G4C2)4-induced foci is concentration dependent (Fig 2B). Kinetically, G3BP1 and eIF3b are quickly condensed by r(G4C2)4, and are unchanged over one hour (Fig 4C). This is in contrast to b-isox which shows increased protein condensation over a one hour time course (Fig 4C, compare lanes 8 and 9).
Figure 4. rG4C2 promotes in vitro phase separations.
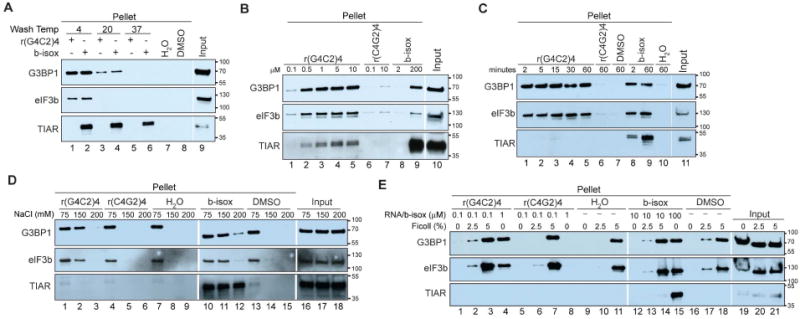
(A-E) Western blots detecting G3BP1, eIF3b, and TIAR from material condensed from U2OS lysate by r(G4C2)4, r(C4G2)4, or b-isox. Condensation was varied by: (A) temperature of wash steps; (B) concentrations of RNA or b-isox; (C) incubation time; (D) NaCl concentration in the buffer (for lysis and the reaction); (E) addition of the crowding agent Ficoll. Cropped blots are indicated by white separation.
RNA granule formation is attributed to phase separation, resulting from multiple low affinity electrostatic and other weak non-covalent interactions between different molecules, that are enhanced by molecular crowding. Thus ionic conditions play a critical role in phase separation (Mitrea and Kriwacki, 2016). At physiologic salt concentration (150 mM NaCl), rG4C2 and b-isox selectively condense specific proteins including G3BP1 and eIF3b (Fig 4D, lanes 2, 5, 8, 11, 14). At hypotonic salt concentration (75 mM NaCl), G3BP1 and eIF3b condense irrespective of whether RNAs, b-isox, or vehicle controls is added (Fig 4D, lanes 1, 4, 7, 10, 13). Under these conditions, TIAR remains specific as it only condenses with b-isox (Fig 4D, lanes 1, 4, 7, 10, 13 bottom panel). At hypertonic salt concentration (200 mM NaCl) rG4C2- and b-isox-mediated condensation is greatly reduced (Fig 4D, lanes 2, 5, 8, 11, 14).
Molecular crowding affects phase separation and RNA granule assembly both in vitro and in cells (Boeynaems et al., 2017; Bounedjah et al., 2012; Kedersha et al., 2016; Molliex et al., 2015). The molecular crowding agent Ficoll causes a ten-fold reduction in r(G4C2)4 and b-isox required to promote condensation (Fig 4E, lanes 2, 6, 10, 13, 17). Higher concentrations of Ficoll induce G3BP1 and eIF3b condensation independent of adding RNA, b-isox, or vehicle control, whereas TIAR condensation is specific to high concentrations of b-isox (Fig 4E). Thus electrostatic interactions and molecular crowding regulate rG4C2-mediated condensation, and such condensation occurs at physiologic salt concentrations and is promoted by molecular crowding.
Cellular RNA is essential for rG4C2-mediated condensation
To assess whether cellular RNA is necessary for rG4C2-mediated condensation, RNase A was added during the incubation or wash steps of the condensation reaction. Addition of RNase A efficiently degrades any RNA (data not shown) and as a result little to no G3BP1 or eIF3b is pelleted. This indicates that RNA (including r(G4C2)4) is necessary both for the initial condensation (Fig 5A, lane 2) and for maintaining the phase separation as the condensed material is partially eliminated by addition of RNase A during the washes (Fig 5A, lane 3). To determine whether cellular mRNAs are required for rG4C2-mediated condensation, lysates were treated with micrococcal nuclease (MNase) prior to the condensation assay. MNase is commonly used to prepare in vitro translation-competent lysates where endogenous mRNAs are eliminated. MNase treatment prevents rG4C2-mediated condensation and only mildly affects b-isox mediated condensation (Fig 5B, (Han et al., 2012)). Analysis of RNA that was harvested prior to centrifugation but after incubation with r(G4C2)4 shows neither major cellular RNA species (e.g., 18S/28S/5S rRNA and tRNAs), nor r(G4C2)4 or r(C4G2)4 are degraded under these conditions (Fig S5), consistent with MNase only affecting mRNAs. Together these data indicate that cellular RNAs are necessary, but not sufficient, to mediate rG4C2-induced condensation.
Figure 5. Cellular RNA is required for rG4C2-mediated phase separation.
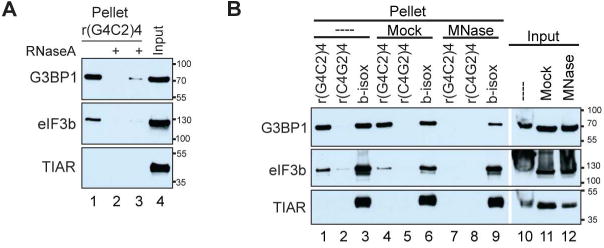
(A) Western blots detecting G3BP1, eIF3b, and TIAR from material condensed after r(G4C2)4 (lanes 1-3) was incubated with U2OS lysate (lanes 1,3) or lysate supplemented with RNase A (lane 2), or washed with buffer with RNase A (lane 2-3). (B) Western blots detecting G3BP1, eIF3b, and TIAR from material condensed after r(G4C2)4, r(C4G2)4, or b-isox were incubated with untreated (lanes 1-3, 10), mock treated (lanes 4-6, 11), or MNase treated (lanes 7-9, 12) U2OS lysate. See also Fig S5. Cropped blots are indicated by white separation.
G-quadruplexes promote rG4C2-mediated condensation
rG4C2 has been reported to assume hairpin and G-quadruplex structures (Fratta et al., 2012; Haeusler et al., 2014; Reddy et al., 2013; Su et al., 2014). Expanded repeats found in other neurological diseases including Huntington's disease (CAGn), and myotonic dystrophy type 1 and 2 (CUGn, CCUGn) can also adopt hairpin conformations (Cleary and Ranum, 2013; Krzyzosiak et al., 2012). We compared the ability of several G-quadruplex-containing oligos, other neurological disease repeat RNAs that assume hairpin structures, and unstructured control RNAs to condense G3BP1 and eIF3b from cell lysates. Whereas G-quadruplex-containing oligos (rG4C2, rG3U3, 5′Ala tiRNA, rUUAGGG) condense G3BP1 and eIF3b, other neurological disease-associated RNA repeats do not (Fig 6A), suggesting G-quadruplex structures may promote this phase separation event.
Figure 6. RNA G-quadruplexes promote phase transitions in vitro and in cells.
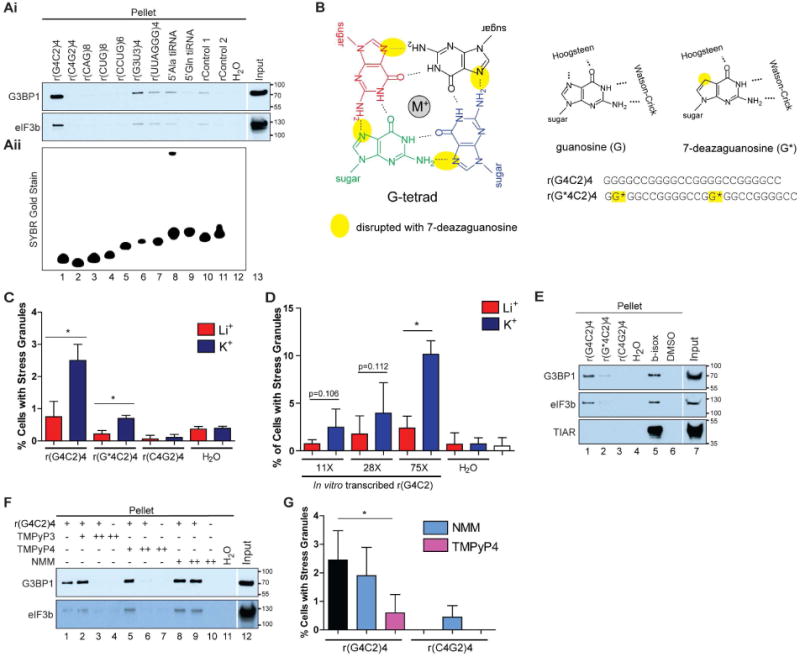
(Ai) Western blot analysis of pelleted material after indicated RNAs were incubated with U2OS cell lysate. (Aii) SYBR Gold stained 15% TBE-Urea gel detecting RNA loaded prior to lysate addition (Ai). (B) Four guanosine residues form a G-tetrad via hydrogen bonding using the Watson-Crick and Hoogsteen interfaces. Substitution of 7-deazaguanosine (denoted as G*) for guanosine prevents a Hoogsteen base pair (yellow ovals). The G-tetrad is coordinated with a metal ion (denoted M+) in the central channel. (C-D) SG quantification after transfection with (C) r(G4C2)4, r(G*4C2)4, or r(C4G2)4 or (D) in vitro transcribed r(G4C2)-11×, 28×, or 75× equilibrated with Li+ or K+. (E-F) Western blots detecting G3BP1, eIF3b, and TIAR condensed from U2OS lysate by (E) r(G4C2)4, r(G*4C2)4, or r(C4G2)4 or (F) r(G4C2)4 preincubated with TMPyP3, TMPyP4, or NMM (+ for 2 μM, ++ for 10 μM) for 30 min. (G) SG quantification of U2OS cells transfected with r(G4C2)4 or r(C4G2)4 preincubated with of TMPyP4 or NMM (100 μM) for 30 min prior to transfection. * denotes statistical significance with p<0.05. Data are represented as mean ± SD, n≥3. See also Fig S6. Cropped blots are indicated by white separation.
G-quadruplexes are composed of stacked G-tetrads, four guanosine residues hydrogen bonded by Hoogsteen base pairs, coordinated by a central monovalent cation (Fig 6B). The size of the cation is critical, as potassium ions support G-quadruplex assembly whereas smaller lithium ions do not (Neidle, 2012). Potassium equilibration reduces the mobility of r(G4C2)4 in native gels, a phenomenon that is not observed with lithium equilibration (Fig S6A), consistent with rG4C2 forming a G-quadruplex (Fratta et al., 2012; Haeusler et al., 2014; Reddy et al., 2013; Su et al., 2014). These slower-migrating species are also observed under denaturing gel conditions (urea within the gel and formamide in the loading dye) (Fig S6B), indicative of the remarkable stability of G-quadruplexes. Similar shifts in mobility were also observed with longer in vitro transcribed rG4C2 repeats (Fig S6C). Additionally, r(G4C2)4 in the G-quadruplex conformation shows specificity in binding partners as lithium-equilibrated and potassium-equilibrated 3′biotinylated r(G4C2)4 have different binding partners from cell lysate (Fig S6E-F, compare lanes 1 and 2). Together these data are consistent with rG4C2 forming G-quadruplex structures that have a specific subset of binding partners.
Transfection of potassium-equilibrated rG4C2, including both r(G4C2)4 and in vitro transcribed r(G4C2) 11×, 28×, 75× RNA into U2OS cells promotes more SG assembly whereas lithium-equilibrated RNAs promote fewer SGs (Fig 6C,D). r(C4G2)4 equilibrated with either potassium or lithium, or potassium or lithium alone do not induce SG assembly (Fig 6C,D).
rScram is able to promote visual foci and to condense G3BP1 and eIF3b (Fig 2A,B, 3F). Because of its high G-content, this control RNA may form G-quadruplex structures (intra- or inter-molecularly) that partially mimic rG4C2. We assessed lithium and potassium equilibrated rScram by gel electrophoresis and found rScram forms higher molecular weight species that are sensitive to potassium concentrations, suggesting rScram can form G-quadruplex-like structures (Fig S6D).
Substitution of guanosine for its analogue 7-deazaguanosine maintains Watson-Crick base pairing but does not support Hoogsteen hydrogen bonding, thereby destabilizing G-quadruplex structures (Fig 6B; (Murchie and Lilley, 1992)). Introduction of two 7-deazaguanosines into r(G4C2)4 (referred to as r(G*4C2)4) affects its mobility on native and denaturing gels (Fig S6A-B), decreases SG formation compared to r(G4C2)4 (Fig 6C), and impairs condensation of G3BP1 and eIF3b (Fig 6E, compare lanes 1 and 2). Moreover, compounds that disrupt G-quadruplexes tetra-(N-methyl-3-pyridyl)porphyrin (TMPyP3) and tetra-(N-methyl-4-pyridyl)porphyrin (TMPyP4)) (Han et al., 2001) inhibit r(G4C2)4-mediated condensation of G3BP1 or eIF3b, whereas N-methylmorpholine (NMM), a G-quadruplex stabilizer (Nicoludis et al., 2012), slightly enhances condensation of G3BP1 and eIF3b (Fig 6F). r(G4C2)4-induced SG assembly is significantly decreased when pretreated with TMPyP4, yet no change is observed with NMM (Fig 6G). These data are consistent with a model whereby rG4C2 G-quadruplexes phase separate in vitro by condensing RNA granule components including G3BP1 and eIF3b, which in cells promotes SG formation.
Discussion
The mechanisms of RNA granule assembly are extremely complex due to overlapping contributions from multiple protein:protein and protein:RNA interactions. Recent findings highlight fundamental roles for protein-protein interactions and phase transitions in granule formation (Mitrea and Kriwacki, 2016). Paradoxically, although RNA granules are RNA-containing entities by definition, the role(s) of RNA in this process is largely unknown. Recent work from the Vale lab indicate that RNA associated with repeat expansions can phase separate in vitro and in cells (Jain and Vale, 2017). Using rG4C2 as a model RNA, here, we demonstrated that RNA G-quadruplexes nucleate RNA granule assembly in cells and promote phase transitions in vitro.
Our study is unique as the in vitro phase separation uses lysate under physiological ionic conditions, better recapitulating the cellular environment. We found rG4C2 promotes condensation of a specific subset of proteins from cell lysates (Fig 3). The majority of these proteins are also condensed by b-isox, a chemical that induces phase transitions in vitro. rG4C2 and b-isox condensed proteins are enriched for low complexity, prion-like and intrinsically disordered domains that contribute to RNA granule assembly both in vitro and in cells. However, the modes of rG4C2 and b-isox-mediated condensation differ. In the absence of lysate, b-isox forms a white pellet (data not shown) while rG4C2 fails to condense (Fig 2, S3). b-isox forms a structural lattice, independent of other factors, that then templates low complexity domains into a specific state (Kato et al., 2012). In contrast, rG4C2-mediated condensation requires factors within the lysate, including cellular RNA (Fig 2, 5, S3). Importantly, rG4C2 promotes RNA granule formation in cells, both cytoplasmic SGs and nuclear RNA foci (Fig 1, S1), mirroring its ability to promote RNA-granule-like condensation in vitro.
In our analysis, we tracked G3BP1, eIF3b, and TIAR due to their importance in RNA metabolism and RNA granule assembly. Whereas G3BP1 and eIF3b reliably condense with both rG4C2 and b-isox, TIAR consistently condensed with b-isox and only under some conditions with rG4C2. Interestingly, TIAR condenses with longer rG4C2 repeats or under high concentrations of short repeats (Fig 3G, 4B). These results suggest that condensation of some protein factors requires rG4C2 to reach a certain threshold, either by increasing the length of repeats or by increasing the concentration of shorter repeats. Differential condensation of protein factors may be relevant to ALS/FTD where longer repeat-containing rG4C2 transcripts form visible foci that recruit RNA binding proteins and cause their mislocalization (Gitler and Tsuiji, 2016).
Like b-isox induced condensation, rG4C2-mediated condensation is reversible, fast, concentration- and temperature-dependent, and sensitive to molecular crowding. It is highly responsive to ionic conditions, which both modulate electrostatic interactions between rG4C2 and other molecules during condensation and directly contribute to stabilization of G-quadruplexes. Additionally our experiments were conducted under physiological ionic conditions. The rG4C2 G-quadruplex structure is required for granule formation (Fig 6), and our model is that rG4C2 G-quadruplexes seed granule formation. rG4C2 appears to take on a solidlike state due to its lack of dynamic properties (Fig 1F, S2), consistent with rG4C2 being a core granule component (Jain et al., 2016; Kedersha et al., 2005; Wheeler et al., 2016). Increasing the number of rG4C2 G-quadruplex units by increasing rG4C2 repeat length or rG4C2 concentration increases nucleation of microscopically visible granules and RNA mediated condensation in vitro. In this manner, RNA structure drives granule formation. Most, if not all, RNA within the cell is thought to be coated with proteins. It is likely that a critical concentration of free RNA or disruption of RNA-protein interactions is required to allow selected RNAs to assemble G-quadruplex structures. Currently, it remains unknown whether and to what extent RNA G-quadruplexes fold in vivo. Algorithms predict that a high percentage of the transcriptome is capable of forming G-quadruplexes (Huppert and Balasubramanian, 2005; Todd et al., 2005), yet recent genome-wide sequencing methodologies suggest G-quadruplexes are not formed in cells (Guo and Bartel, 2016; Kwok et al., 2016), indicating a clear need for further investigation. Our data indicate that when folded into a G-quadruplex rG4C2 has differential binding partners (Fig S6E-F) and nucleates RNA granules (Fig 6), and that this occurs to a greater extent with more RNA repeats. We observed no change in SG formation when we transfected r(G4C2)4 and NMM treated r(G4C2)4 (Fig 6G). This suggests that we cannot further promote G-quadruplex folding with NMM suggesting that RNA G-quadruplexes can exist in cells.
For C9-ALS/FTD, we hypothesize that by increasing the length of rG4C2, more repeats allow more templating, increasing the recruitment of proteins/RNA to allow RNA granule assembly. In turn, nucleation of granules may affect RNA metabolism in neurons and trigger pathological cascades culminating in neuron loss. These events may be more detrimental in neurons due to the necessity of localized translation, which requires transport of RNAs and translational machinery down long axonal processes.
We propose that other RNA structures may also template or affect RNA granules in cells. Consistent with this hypothesis, SGs form in response to stress that causes translating polysomes to disassemble producing a sudden excess of free mRNAs in the cytoplasm. It is hypothesized that this mRNA pool nucleates the formation of RNA granules by binding a large set of RBPs (Bounedjah et al., 2014; Wheeler et al., 2016). It will be important to determine whether other mRNA structures regulate SG formation.
Proteins participating in physiological phase transitions are prone to assemble protein aggregates when they possess disease-associated mutations (Bosco et al., 2011; Burke et al., 2015; Conicella et al., 2016; Kim et al., 2013; Molliex et al., 2015; Murakami et al., 2015; Patel et al., 2015). Similarly, various RNAs acquire repeat expansion mutations that increase the structural complexity and/or sequence units (Cleary and Ranum, 2014; Krzyzosiak et al., 2012). We propose that RNA plays an active role in RNA granule assembly and that RNA-mediated perturbations in granule dynamics may underline cellular toxicity and contribute to disease.
Experimental Procedures
Cell culture and cell treatments
Cell lines (U2OS, U251, LN229, and MEFs) were maintained in 5-7% CO2 and cultured in DMEM supplemented with 20 mM Hepes, 10% FBS, 100 U/mL penicillin and 100 μg/mL streptomycin. Stable U2OS cells expressing mCherry-tagged G3BP1 were previously described (Kedersha et al., 2016).
Reagents
Synthetic RNA oligos were purchased from Integrated DNA Technologies (Table S1) and reagents and antibodies are indicated in Tables S2 and S3.
RNA and b-isox precipitation
3×106 U2OS cells or comparable density for other cells were plated on 10 cm dishes. The next day, cells were washed with HBSS, scraped with 500 μL condensation buffer (CB, 50 mM Hepes, 10% glycerol, 150 mM NaCl, 5 mM MgCl2, 0.5% NP40, Halt Protease and Phosphatase Inhibitors, 1 mM DTT, RNaseIN). For mouse brain, the brain was harvested, flash frozen and stored at -80°C. Samples were dounce homogenized, c entrifuged at 14,000 × g for 10 min at 4°C, and supernatant was used as lysate. RNA (1 μM, unless otherwise indicated), b-isox (100 μM) or vehicle controls (H2O or DMSO) were added to lysate and incubated for 1 hr at 4°C. Samples were centrifuged at 14,000 × g for 15 min at 4°C, supernatant was discarded and resulting pellet was washed twice with CB. The pellet was resuspended in 1× SDS loading dye for western blot or silver stain analysis, trichloroacetic acid (TCA) for mass spectrometry, or Trizol for RNA analysis.
Modifications to the procedure are indicated. For MNase reactions, lysates were made as above then treated with MNase (0.06 U/mL) supplemented with 1 mM CaCl2 for 10 min at 37°C then quenched with 2 mM EGTA. Mock treatment included all steps but lacked MNase. For RNase A experiments, RNase A (1 μg/mL) was added to the reaction or wash steps. For salt-adjusted experiments, cells were lysed and washed in CB with indicated NaCl. For Ficoll addition, Ficoll was supplemented to lysate prior to reaction and to wash buffer.
Western blot/silver stain
Samples were generated as described above and input controls were diluted in 1× SDS sample buffer, heated at 95°C for 5 min and run on Mini-Pr otean TGX gels. Proteins were transferred to nitrocellulose using Trans-Blot Turbo (Bio-Rad), and blocked in 5% non-fat milk in TBST for ≥20 min. Primary antibodies were diluted in 5% normal horse serum (NHS) and incubated overnight at 4 °C or 1 hr at ∼20°C. Blots were washed ≥3 times for 10 min at ∼20°C with TBST. Secondary antibodies were incubated for ≥30 min at ∼20°C. Blots were washed, ECL was applied, and signals detected using film or Chemidoc Imaging System (Bio-Rad).
For silver stain analysis, samples were run on 4-20% Tris-Glycine gels then stained using the Pierce Silver Stain Kit and manufacturer's recommendations.
Visualization and quantification of in vitro precipitates
Lysates were generated as described above, then mixed with RNA as indicated. Samples were centrifuged at 14,000 × g for 15 min, and pellets resuspended in 10 μL CB. 4 μL was placed on a glass slide and 22×22 mm coverslip on top. 10 min post-mounting, 3-5 random fields were imaged with a 60× oil objective on the Nikon microscope described elsewhere. Images were quantified using ImageJ by Thresholding with the Triangle algorithm, then “Analyze Particles…” function (particle = 5 to ■ pixels2).
Quantification of SGs
ImageJ was used to quantify the number of SGs per cell and the size of SG. 8 bit grey scale G3BP1 images were thresholded, the cell was outlined via freehand selection and “Analyze Particles” assessed both size and number of particles per cell. Data was exported to Excel and graphs were generated with Prism 5.0. To assess SG size approximately the same minimum Threshold was used and the size noted is in pixels where ∼3.2 pixels = 1 μm.
In vitro transcription
Gel purified pEF6-(G4C2) repeat plasmids (May et al., 2014; Mori et al., 2013) cleaved with XbaI were used in standard reaction HiScribe T7 polymerase RNA synthesis or T7-Riboprobe System following manufacturer's recommendations. After 3 hr incubation, 1 μL DNase I was added and incubated for 30 min. RNA was extract with Trizol.
RNA gel analysis
500 μL HBSS and 700 μL Trizol were added to the pellet of the condensation reaction then RNA extracted. For denaturing gels, RNA was heated in denaturing loading dye, cooled and analyzed on 15% TBE-Urea gel. For native gels, RNA was diluted in 1× sample buffer and run on 10% TBE gel (10% acrylamide:bis-acrylamide (19:1), 0.5× TBE, 0.01% APS, 0.1% TEMED). To visualize RNA, gels were stained with SYBR Gold (1:10,000 in 0.5× TBE) for 10 min and visualized by UV light.
Immunofluorescence
1×105 U2OS cells were plated on 12 mm glass coverslips. For RNA transfections, the following day cells were transfected with indicated RNAs using Lipofectamine 2000. Cycloheximide (100 μg/mL) and puromycin (5 μg/mL) treatment was carried out 30 min prior to fixation as indicated. 6-8 hr post transfection cells were washed with PBS, fixed with 4% paraformaldehyde at ∼20°C for 15 min, permeabilized with methanol (-20°C) for 5 min and blocked (5% NHS) for ≥20 min. Primary and secondary antibody incubations were carried out for ≥1 hr at ∼20°C or overnight at 4°C. Between antibody incubations coverslips were w ashed 3 times for 5 min with PBS. Hoechst 33258 was added during secondary antibody incubation to visualize nuclei. Coverslips were mounted with Vinol and visualized using a Nikon Eclipse E800 microscope with a mercury lamp and standard filters (UV-2A 360/40; 420/LP), Cy2 (FITC HQ 480/40; 535/50), Cy3 (Cy 3 HQ 545/30; 610/75), and Cy5 (Cy 5 HQ 620/60; 700/75) equip with a SPOT Pursuit digital Camera (Diagnostics Instruments) with the manufacturer's software. Cells were counted as positive for SGs if ≥2 G3BP1 foci were present and nuclei were counted for total number of cells. Graphs were generated with GraphPad Prism 5 and represent compiled data from 3-5 (20× or 40× magnification) images per experiment across ≥3 independent experiments.
Fluorescence recovery after photobleach (FRAP)
U2OS cells expressing mCherry tagged G3BP1 were plated onto 35-mm FluoroDishes. The following day cells were transfected with 3′FAM labeled r(G4C2)4 using Lipofectamine 2000. Cells were imaged on a Nikon Eclipse TE2000U Inverted Microscope using an Eclipse EZ-C1 system (v. 3.90; Nikon) and a Plan Apo 60× Pan Apo (NA 1.40) objective lens or the Zeiss LSM 800 with Airyscan confocal system on a Zeiss Axio Observer Z1 Inverted Microscope. FAM fluorescence was excited with the 488-nm line from a Melles Griot 488 Ion Laser and detected with a 515/30 emission filter; mCherry fluorescence was obtained using Melles Griot 543 laser excitation and a 590/50 emission filter or best settings as selected by the Zeiss software. Three images were collected prior to photo bleaching a selected region (SG) using 100% (5% for the Zeiss system) laser power, and ten images collected post-photobleaching. Data points include >5 SGs per experiment across 3 experiments. Graphs were generated using GraphPad Prism 5.
Mass spectrometry
For mass spectrometry, b-isox and RNA precipitation assays were scaled up, completed as above, and pellets were resuspended in 20% TCA. Samples were incubated on ice for 20 min, spun down in at 14,000 × g for 20 min at 4°C. The resulting pellet was washed with 10% TCA, then with ice cold acetone. The resulting pellet was air dried for 5 min and stored at -80°C until analyzed by the Taplin Mass Spectrometry facility (Harvard Medical School, Boston) and can be found in Table S4.
Supplementary Material
Figure S1: rG4C2 forms cytoplasmic stress granules and nuclear foci, Related to Figure 1 (A) Immunofluorescence detecting G3BP1 (green), eIF4G (red), and eIF3b (blue) in U2OS cells transfected with r(G4C2)4 and r(C4G2)4. Scale bar is 20 μm. (B) FISH detecting poly(A) (red) coupled with immunofluorescence detecting G3BP1 (green) then stained with Hoechst (blue) in U2OS cells transfected with r(G4C2)4 and r(C4G2)4. Scale bar is 20 μm. (C) Graph indicating percentage of stress granule positive cells (cells with 2 or more G3BP1 positive foci) from part A. (D) FISH detecting G4C2 (red) and poly(A) (green) then stained with Hoechst (blue) in U2OS cells transfected with plasmids (pEF6) encoding indicated number of G4C2 repeats. * denotes statistical significance, p<0.05, n=3. Scale bar is 40 μm.
Figure S2: rG4C2 is stable while G3BP1 exhibits dynamic behavior shuttling in and out of stress granules, Related to Figure 1: FRAP analysis of r(G4C2)4–induced stress granules in U2OS cells stably expressing mCherry-G3BP1 and transiently transfected with 3′FAM labeled r(G4C2)4. (A) Graph represents relative fluorescence of mCherry-G3BP1 (red) and 3′FAM labeled r(G4C2)4 (green) with images taken every 2 seconds and bleaching occurred around ∼6-8 seconds. (B) Representative fluorescent images of mCherry-G3BP1 (red) and 3′FAM labeled r(G4C2)4 (green) FRAP taken before (2 seconds) and after photobleaching (8, 20,38 seconds). Bleached region is indicated by grey circle. Enlarged region is indicated by white box and represents 1.75× of original image.
Figure S3: rG4C2 condensation requires cell lysate, Related to Figure 2: r(G4C2)4 (0.1-10 μM) was incubated with buffer (-Lysate, lanes 1-5) or U2OS lysate (+Lysate, lanes 6-10) and pelleted by centrifugation. RNA was isolated from the pellet and resolved on 15% TBE-Urea gel then stained with SYBR Gold. Input is r(G4C2)4 (lanes 10-13) or lysate (lane 14).
Figure S4: rG4C2 mediated condensation occurs in lysates from other cells and tissues, Related to Figure 3: (A-D) Western blots detecting eIF3b, HuR, TIAR, and/or G3BP1 from pellets formed after RNA (1 μM), b-isox (100 μM), or vehicle controls was incubated for 60 minutes at 4 °C with (A) MEF (mouse embryonic fibroblast) lysate, (B) homogenized mouse brain lysate, (C) U251 (neuroblastoma) cell lysate, (D) LN229 (neuroblastoma) cell lysate. Cropped blots are indicated by white separation.
Figure S5: MNase does not degrade rG4C2, Related to Figure 5: RNA harvested from r(G4C2)4 (1 μM), r(C4G2)4 (1 μM), or b-isox (100 μM) condensation reactions with lysates that were untreated (lanes 1-3, 10), mock treated (treated with 1 mM CaCl2 (1 mM) then quenched with 2 mM EGTA (lanes 4-6, 11) or treated with MNase (0.04 units/mL) and 1 mM CaCl2 then quenched with 2 mM EGTA (lanes 7-9, 12). RNA was run on 15% TBE-Urea gel and stained with SYBR Gold. Inputs include lysates (lanes 10-12) prior to addition of RNA or b-isox, and RNA (lanes 13-14) or b-isox (lane 15) prior to addition of RNA.
Figure S6: rG4C2 G-quadruplex structures show potassium dependence and have specific binding partners, Related to Figure 6: (A) 10% native TBE gel stained with SYBR Gold to visualize RNAs that were equilibrated with 100 mM LiCl or KCl overnight prior to gel analysis. (B) 15% TBE-Urea gel stained with SYBR Gold to visualize RNAs as described in (A). (C) 6% native TBE gel stained with SYBR Gold to visualize in vitro transcribed RNAs that were equilibrated with 100 mM LiCl or KCl overnight prior to gel analysis. (D) 15% TBE-Urea gel stained with SYBR Gold to visualize RNA oligos equilibrated with 100 mM LiCl or KCl overnight prior to gel analysis. (E-F) Material pulldown with streptavidin beads after incubation of 3′ biotin labeled r(G4C2)4 (lanes 1-2) or r(G*4C2)4 (lanes 3-4) equilibrated in 100 mM LiCl (lanes 1, 3) or KCl (2, 4) prior to incubation in U2OS cell lysate. Control reactions lack biotin tagged RNA but were supplemented with LiCl (lane 5) or KCl (lane 6). Material pulled down was analyzed by (E) silver stain of a 4-20% SDS Page gel and (F) western blot probed for CNBP and nucleolin. Cropped blots are indicated by white separation.
Table S1: used in this study, Related to Experimental Procedures
Table S2: Antibodies used in this study, Related to Experimental Procedures
Table S3: Reagents used in this study, Related to Experimental Procedures
Table S4: r(G4C2)4 and b-isox condensed material, Related to Figure 3: Raw mass spectrometry data of proteins condensed by r(G4C2)4 and b-isox divided into three separate tabs, r(G4C2)4 for r(G4C2)4 condensed proteins, b-isox for b-isox condensed proteins, and comparison for a comparison of r(G4C2)4 and b-isox condensed proteins. For r(G4C2)4 and b-isox tabs, column C is the gene symbol, column D is the unique number of reads, column E is total number of reads. For comparison tab, column B is the gene symbol, column C is the unique number of reads for r(G4C2)4, column D is the total number of reads for r(G4C2)4, column E is the unique number of reads for b-isox, column F is the total number of reads for b-isox, and column G is the difference between the total number of r(G4C2)4 and b-isox reads.
Highlights.
GGGGCC RNA (rG4C2) promote assembly of visible foci in vitro
rG4C2 promotes reversible in vitro phase separations similar to b-isox
G-quadruplex structure of rG4C2 promotes phase separations in vitro and in cells
Acknowledgments
We thank Anderson and Ivanov lab members for discussion and Victoria Ivanova for technical support, Christian Haass, Dieter Edbauer, and Kohji Mori (Ludwig-Maximilians University, Munich, Germany) for contributing the pEF6-G4C2-repeat constructs, and Katia Urso from Antonios Aliprantis’ lab (Brigham and Women's Hospital, Harvard Medical School, Boston, USA) for the mice used to harvest brain tissue. This work was supported by the NIH (GM111700 and CA168872 to PA, and NS094918 to PI).
Footnotes
Author Contributions: Investigation, MF; Conceptualization, MF, PI; Supervision, PI, PA; Writing-Original Draft, MF; Writing-Review and Editing, MF, PA, PI; Funding Acquisition, PI, PA
Declaration of Interests: The authors declare no competing interests.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Aulas A, Fay MM, Lyons SM, Achorn CA, Kedersha N, Anderson P, Ivanov P. Stress-specific differences in assembly and composition of stress granules and related foci. J Cell Sci. 2017 doi: 10.1242/jcs.199240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bang I. Examination of the guanyle acid. Biochemische Zeitschrift. 1910;26:293–311. [Google Scholar]
- Boeynaems S, Bogaert E, Kovacs D, Konijnenberg A, Timmerman E, Volkov A, Guharoy M, De Decker M, Jaspers T, Ryan VH, et al. Phase Separation of C9orf72 Dipeptide Repeats Perturbs Stress Granule Dynamics. Molecular cell. 2017;65:1044–1055. doi: 10.1016/j.molcel.2017.02.013. e1045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bosco DA, Lemay N, Ko HK, Zhou H, Burke C, Kwiatkowski TJ, Jr, Sapp P, McKenna-Yasek D, Brown RH, Jr, Hayward LJ. Mutant FUS proteins that cause amyotrophic lateral sclerosis incorporate into stress granules. Hum Mol Genet. 2011;19:4160–4175. doi: 10.1093/hmg/ddq335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bounedjah O, Desforges B, Wu TD, Pioche-Durieu C, Marco S, Hamon L, Curmi PA, Guerquin-Kern JL, Pietrement O, Pastre D. Free mRNA in excess upon polysome dissociation is a scaffold for protein multimerization to form stress granules. Nucleic acids research. 2014;42:8678–8691. doi: 10.1093/nar/gku582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bounedjah O, Hamon L, Savarin P, Desforges B, Curmi PA, Pastre D. Macromolecular crowding regulates assembly of mRNA stress granules after osmotic stress: new role for compatible osmolytes. The Journal of biological chemistry. 2012;287:2446–2458. doi: 10.1074/jbc.M111.292748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burke KA, Janke AM, Rhine CL, Fawzi NL. Residue-by-Residue View of In Vitro FUS Granules that Bind the C-Terminal Domain of RNA Polymerase II. Molecular cell. 2015;60:231–241. doi: 10.1016/j.molcel.2015.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cleary JD, Ranum LP. Repeat-associated non-ATG (RAN) translation in neurological disease. Human molecular genetics. 2013;22:R45–51. doi: 10.1093/hmg/ddt371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cleary JD, Ranum LP. Repeat associated non-ATG (RAN) translation: new starts in microsatellite expansion disorders. Curr Opin Genet Dev. 2014;26:6–15. doi: 10.1016/j.gde.2014.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conicella AE, Zerze GH, Mittal J, Fawzi NL. ALS Mutations Disrupt Phase Separation Mediated by alpha-Helical Structure in the TDP-43 Low-Complexity C-Terminal Domain. Structure. 2016;24:1537–1549. doi: 10.1016/j.str.2016.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeJesus-Hernandez M, Mackenzie IR, Boeve BF, Boxer AL, Baker M, Rutherford NJ, Nicholson AM, Finch NA, Flynn H, Adamson J, et al. Expanded GGGGCC hexanucleotide repeat in noncoding region of C9ORF72 causes chromosome 9p-linked FTD and ALS. Neuron. 2011;72:245–256. doi: 10.1016/j.neuron.2011.09.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donnelly CJ, Zhang PW, Pham JT, Haeusler AR, Mistry NA, Vidensky S, Daley EL, Poth EM, Hoover B, Fines DM, et al. RNA toxicity from the ALS/FTD C9ORF72 expansion is mitigated by antisense intervention. Neuron. 2013;80:415–428. doi: 10.1016/j.neuron.2013.10.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elbaum-Garfinkle S, Kim Y, Szczepaniak K, Chen CC, Eckmann CR, Myong S, Brangwynne CP. The disordered P granule protein LAF-1 drives phase separation into droplets with tunable viscosity and dynamics. Proceedings of the National Academy of Sciences of the United States of America. 2015;112:7189–7194. doi: 10.1073/pnas.1504822112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Emara MM, Ivanov P, Hickman T, Dawra N, Tisdale S, Kedersha N, Hu GF, Anderson P. Angiogenin-induced tRNA-derived stress-induced RNAs promote stress-induced stress granule assembly. The Journal of biological chemistry. 2010;285:10959–10968. doi: 10.1074/jbc.M109.077560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fratta P, Mizielinska S, Nicoll AJ, Zloh M, Fisher EM, Parkinson G, Isaacs AM. C9orf72 hexanucleotide repeat associated with amyotrophic lateral sclerosis and frontotemporal dementia forms RNA G-quadruplexes. Scientific reports. 2012;2:1016. doi: 10.1038/srep01016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilks N, Kedersha N, Ayodele M, Shen L, Stoecklin G, Dember LM, Anderson P. Stress granule assembly is mediated by prion-like aggregation of TIA-1. Molecular biology of the cell. 2004;15:5383–5398. doi: 10.1091/mbc.E04-08-0715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gitler AD, Tsuiji H. There has been an awakening: Emerging mechanisms of C9orf72 mutations in FTD/ALS. Brain research. 2016;1647:19–29. doi: 10.1016/j.brainres.2016.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo JU, Bartel DP. RNA G-quadruplexes are globally unfolded in eukaryotic cells and depleted in bacteria. Science. 2016;353 doi: 10.1126/science.aaf5371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haeusler AR, Donnelly CJ, Periz G, Simko EA, Shaw PG, Kim MS, Maragakis NJ, Troncoso JC, Pandey A, Sattler R, et al. C9orf72 nucleotide repeat structures initiate molecular cascades of disease. Nature. 2014;507:195–200. doi: 10.1038/nature13124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han H, Langley DR, Rangan A, Hurley LH. Selective interactions of cationic porphyrins with G-quadruplex structures. Journal of the American Chemical Society. 2001;123:8902–8913. doi: 10.1021/ja002179j. [DOI] [PubMed] [Google Scholar]
- Han TW, Kato M, Xie S, Wu LC, Mirzaei H, Pei J, Chen M, Xie Y, Allen J, Xiao G, et al. Cell-free formation of RNA granules: bound RNAs identify features and components of cellular assemblies. Cell. 2012;149:768–779. doi: 10.1016/j.cell.2012.04.016. [DOI] [PubMed] [Google Scholar]
- Huppert JL, Balasubramanian S. Prevalence of quadruplexes in the human genome. Nucleic acids research. 2005;33:2908–2916. doi: 10.1093/nar/gki609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hyman AA, Weber CA, Julicher F. Liquid-liquid phase separation in biology. Annual review of cell and developmental biology. 2014;30:39–58. doi: 10.1146/annurev-cellbio-100913-013325. [DOI] [PubMed] [Google Scholar]
- Ivanov P, O'Day E, Emara MM, Wagner G, Lieberman J, Anderson P. G-quadruplex structures contribute to the neuroprotective effects of angiogenin-induced tRNA fragments. Proceedings of the National Academy of Sciences of the United States of America. 2014;111:18201–18206. doi: 10.1073/pnas.1407361111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jain A, Vale RD. RNA phase transitions in repeat expansion disorders. Nature. 2017;546:243–247. doi: 10.1038/nature22386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jain S, Wheeler JR, Walters RW, Agrawal A, Barsic A, Parker R. ATPase-Modulated Stress Granules Contain a Diverse Proteome and Substructure. Cell. 2016 doi: 10.1016/j.cell.2015.12.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kato M, Han TW, Xie S, Shi K, Du X, Wu LC, Mirzaei H, Goldsmith EJ, Longgood J, Pei J, et al. Cell-free formation of RNA granules: low complexity sequence domains form dynamic fibers within hydrogels. Cell. 2012;149:753–767. doi: 10.1016/j.cell.2012.04.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kedersha N, Cho MR, Li W, Yacono PW, Chen S, Gilks N, Golan DE, Anderson P. Dynamic shuttling of TIA-1 accompanies the recruitment of mRNA to mammalian stress granules. J Cell Biol. 2000;151:1257–1268. doi: 10.1083/jcb.151.6.1257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kedersha N, Ivanov P, Anderson P. Stress granules and cell signaling: more than just a passing phase? Trends in biochemical sciences. 2013;38:494–506. doi: 10.1016/j.tibs.2013.07.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kedersha N, Panas MD, Achorn CA, Lyons S, Tisdale S, Hickman T, Thomas M, Lieberman J, McInerney GM, Ivanov P, et al. G3BP-Caprin1-USP10 complexes mediate stress granule condensation and associate with 40S subunits. J Cell Biol. 2016;212:845–860. doi: 10.1083/jcb.201508028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kedersha N, Stoecklin G, Ayodele M, Yacono P, Lykke-Andersen J, Fritzler MJ, Scheuner D, Kaufman RJ, Golan DE, Anderson P. Stress granules and processing bodies are dynamically linked sites of mRNP remodeling. J Cell Biol. 2005;169:871–884. doi: 10.1083/jcb.200502088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim HJ, Kim NC, Wang YD, Scarborough EA, Moore J, Diaz Z, MacLea KS, Freibaum B, Li S, Molliex A, et al. Mutations in prion-like domains in hnRNPA2B1 and hnRNPA1 cause multisystem proteinopathy and ALS. Nature. 2013;495:467–473. doi: 10.1038/nature11922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krzyzosiak WJ, Sobczak K, Wojciechowska M, Fiszer A, Mykowska A, Kozlowski P. Triplet repeat RNA structure and its role as pathogenic agent and therapeutic target. Nucleic acids research. 2012;40:11–26. doi: 10.1093/nar/gkr729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwok CK, Marsico G, Sahakyan AB, Chambers VS, Balasubramanian S. rG4-seq reveals widespread formation of G-quadruplex structures in the human transcriptome. Nature methods. 2016;13:841–844. doi: 10.1038/nmeth.3965. [DOI] [PubMed] [Google Scholar]
- La Spada AR, Taylor JP. Repeat expansion disease: progress and puzzles in disease pathogenesis. Nature reviews Genetics. 2010;11:247–258. doi: 10.1038/nrg2748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lessel D, Schob C, Kury S, Reinders MRF, Harel T, Eldomery MK, Coban-Akdemir Z, Denecke J, Edvardson S, Colin E, et al. De Novo Missense Mutations in DHX30 Impair Global Translation and Cause a Neurodevelopmental Disorder. American journal of human genetics. 2017;101:716–724. doi: 10.1016/j.ajhg.2017.09.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin Y, Protter DS, Rosen MK, Parker R. Formation and Maturation of Phase-Separated Liquid Droplets by RNA-Binding Proteins. Molecular cell. 2015;60:208–219. doi: 10.1016/j.molcel.2015.08.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- May S, Hornburg D, Schludi MH, Arzberger T, Rentzsch K, Schwenk BM, Grasser FA, Mori K, Kremmer E, Banzhaf-Strathmann J, et al. C9orf72 FTLD/ALS-associated Gly-Ala dipeptide repeat proteins cause neuronal toxicity and Unc119 sequestration. Acta Neuropathol. 2014;128:485–503. doi: 10.1007/s00401-014-1329-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitrea DM, Kriwacki RW. Phase separation in biology; functional organization of a higher order. Cell communication and signaling: CCS. 2016;14:1. doi: 10.1186/s12964-015-0125-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Molliex A, Temirov J, Lee J, Coughlin M, Kanagaraj AP, Kim HJ, Mittag T, Taylor JP. Phase separation by low complexity domains promotes stress granule assembly and drives pathological fibrillization. Cell. 2015;163:123–133. doi: 10.1016/j.cell.2015.09.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mori K, Weng SM, Arzberger T, May S, Rentzsch K, Kremmer E, Schmid B, Kretzschmar HA, Cruts M, Van Broeckhoven C, et al. The C9orf72 GGGGCC repeat is translated into aggregating dipeptide-repeat proteins in FTLD/ALS. Science. 2013;339:1335–1338. doi: 10.1126/science.1232927. [DOI] [PubMed] [Google Scholar]
- Murakami T, Qamar S, Lin JQ, Schierle GS, Rees E, Miyashita A, Costa AR, Dodd RB, Chan FT, Michel CH, et al. ALS/FTD Mutation-Induced Phase Transition of FUS Liquid Droplets and Reversible Hydrogels into Irreversible Hydrogels Impairs RNP Granule Function. Neuron. 2015;88:678–690. doi: 10.1016/j.neuron.2015.10.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murchie AI, Lilley DM. Retinoblastoma susceptibility genes contain 5′ sequences with a high propensity to form guanine-tetrad structures. Nucleic acids research. 1992;20:49–53. doi: 10.1093/nar/20.1.49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neidle S. Therapeutic applications of quadruplex nucleic acids. 1st. London; Waltham, MA: Elsevier Academic Press; 2012. [Google Scholar]
- Nicoludis JM, Barrett SP, Mergny JL, Yatsunyk LA. Interaction of human telomeric DNA with N-methyl mesoporphyrin IX. Nucleic acids research. 2012;40:5432–5447. doi: 10.1093/nar/gks152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patel A, Lee HO, Jawerth L, Maharana S, Jahnel M, Hein MY, Stoynov S, Mahamid J, Saha S, Franzmann TM, et al. A Liquid-to-Solid Phase Transition of the ALS Protein FUS Accelerated by Disease Mutation. Cell. 2015;162:1066–1077. doi: 10.1016/j.cell.2015.07.047. [DOI] [PubMed] [Google Scholar]
- Reddy K, Zamiri B, Stanley SY, Macgregor RB, Jr, Pearson CE. The disease-associated r(GGGGCC)n repeat from the C9orf72 gene forms tract length-dependent uni- and multimolecular RNA G-quadruplex structures. The Journal of biological chemistry. 2013;288:9860–9866. doi: 10.1074/jbc.C113.452532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Renton AE, Majounie E, Waite A, Simon-Sanchez J, Rollinson S, Gibbs JR, Schymick JC, Laaksovirta H, van Swieten JC, Myllykangas L, et al. A hexanucleotide repeat expansion in C9ORF72 is the cause of chromosome 9p21-linked ALS-FTD. Neuron. 2011;72:257–268. doi: 10.1016/j.neuron.2011.09.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Su Z, Zhang Y, Gendron TF, Bauer PO, Chew J, Yang WY, Fostvedt E, Jansen-West K, Belzil VV, Desaro P, et al. Discovery of a biomarker and lead small molecules to target r(GGGGCC)-associated defects in c9FTD/ALS. Neuron. 2014;83:1043–1050. doi: 10.1016/j.neuron.2014.07.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Todd AK, Johnston M, Neidle S. Highly prevalent putative quadruplex sequence motifs in human DNA. Nucleic acids research. 2005;33:2901–2907. doi: 10.1093/nar/gki553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Mossevelde S, van der Zee J, Cruts M, Van Broeckhoven C. Relationship between C9orf72 repeat size and clinical phenotype. Current opinion in genetics & development. 2017;44:117–124. doi: 10.1016/j.gde.2017.02.008. [DOI] [PubMed] [Google Scholar]
- Wen WL, Stevenson AL, Wang CY, Chen HJ, Kearsey SE, Norbury CJ, Watt S, Bahler J, Wang SW. Vgl1, a multi-KH domain protein, is a novel component of the fission yeast stress granules required for cell survival under thermal stress. Nucleic acids research. 2010;38:6555–6566. doi: 10.1093/nar/gkq555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wheeler JR, Matheny T, Jain S, Abrisch R, Parker R. Distinct stages in stress granule assembly and disassembly. eLife. 2016:5. doi: 10.7554/eLife.18413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang H, Elbaum-Garfinkle S, Langdon EM, Taylor N, Occhipinti P, Bridges AA, Brangwynne CP, Gladfelter AS. RNA Controls PolyQ Protein Phase Transitions. Molecular cell. 2015;60:220–230. doi: 10.1016/j.molcel.2015.09.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1: rG4C2 forms cytoplasmic stress granules and nuclear foci, Related to Figure 1 (A) Immunofluorescence detecting G3BP1 (green), eIF4G (red), and eIF3b (blue) in U2OS cells transfected with r(G4C2)4 and r(C4G2)4. Scale bar is 20 μm. (B) FISH detecting poly(A) (red) coupled with immunofluorescence detecting G3BP1 (green) then stained with Hoechst (blue) in U2OS cells transfected with r(G4C2)4 and r(C4G2)4. Scale bar is 20 μm. (C) Graph indicating percentage of stress granule positive cells (cells with 2 or more G3BP1 positive foci) from part A. (D) FISH detecting G4C2 (red) and poly(A) (green) then stained with Hoechst (blue) in U2OS cells transfected with plasmids (pEF6) encoding indicated number of G4C2 repeats. * denotes statistical significance, p<0.05, n=3. Scale bar is 40 μm.
Figure S2: rG4C2 is stable while G3BP1 exhibits dynamic behavior shuttling in and out of stress granules, Related to Figure 1: FRAP analysis of r(G4C2)4–induced stress granules in U2OS cells stably expressing mCherry-G3BP1 and transiently transfected with 3′FAM labeled r(G4C2)4. (A) Graph represents relative fluorescence of mCherry-G3BP1 (red) and 3′FAM labeled r(G4C2)4 (green) with images taken every 2 seconds and bleaching occurred around ∼6-8 seconds. (B) Representative fluorescent images of mCherry-G3BP1 (red) and 3′FAM labeled r(G4C2)4 (green) FRAP taken before (2 seconds) and after photobleaching (8, 20,38 seconds). Bleached region is indicated by grey circle. Enlarged region is indicated by white box and represents 1.75× of original image.
Figure S3: rG4C2 condensation requires cell lysate, Related to Figure 2: r(G4C2)4 (0.1-10 μM) was incubated with buffer (-Lysate, lanes 1-5) or U2OS lysate (+Lysate, lanes 6-10) and pelleted by centrifugation. RNA was isolated from the pellet and resolved on 15% TBE-Urea gel then stained with SYBR Gold. Input is r(G4C2)4 (lanes 10-13) or lysate (lane 14).
Figure S4: rG4C2 mediated condensation occurs in lysates from other cells and tissues, Related to Figure 3: (A-D) Western blots detecting eIF3b, HuR, TIAR, and/or G3BP1 from pellets formed after RNA (1 μM), b-isox (100 μM), or vehicle controls was incubated for 60 minutes at 4 °C with (A) MEF (mouse embryonic fibroblast) lysate, (B) homogenized mouse brain lysate, (C) U251 (neuroblastoma) cell lysate, (D) LN229 (neuroblastoma) cell lysate. Cropped blots are indicated by white separation.
Figure S5: MNase does not degrade rG4C2, Related to Figure 5: RNA harvested from r(G4C2)4 (1 μM), r(C4G2)4 (1 μM), or b-isox (100 μM) condensation reactions with lysates that were untreated (lanes 1-3, 10), mock treated (treated with 1 mM CaCl2 (1 mM) then quenched with 2 mM EGTA (lanes 4-6, 11) or treated with MNase (0.04 units/mL) and 1 mM CaCl2 then quenched with 2 mM EGTA (lanes 7-9, 12). RNA was run on 15% TBE-Urea gel and stained with SYBR Gold. Inputs include lysates (lanes 10-12) prior to addition of RNA or b-isox, and RNA (lanes 13-14) or b-isox (lane 15) prior to addition of RNA.
Figure S6: rG4C2 G-quadruplex structures show potassium dependence and have specific binding partners, Related to Figure 6: (A) 10% native TBE gel stained with SYBR Gold to visualize RNAs that were equilibrated with 100 mM LiCl or KCl overnight prior to gel analysis. (B) 15% TBE-Urea gel stained with SYBR Gold to visualize RNAs as described in (A). (C) 6% native TBE gel stained with SYBR Gold to visualize in vitro transcribed RNAs that were equilibrated with 100 mM LiCl or KCl overnight prior to gel analysis. (D) 15% TBE-Urea gel stained with SYBR Gold to visualize RNA oligos equilibrated with 100 mM LiCl or KCl overnight prior to gel analysis. (E-F) Material pulldown with streptavidin beads after incubation of 3′ biotin labeled r(G4C2)4 (lanes 1-2) or r(G*4C2)4 (lanes 3-4) equilibrated in 100 mM LiCl (lanes 1, 3) or KCl (2, 4) prior to incubation in U2OS cell lysate. Control reactions lack biotin tagged RNA but were supplemented with LiCl (lane 5) or KCl (lane 6). Material pulled down was analyzed by (E) silver stain of a 4-20% SDS Page gel and (F) western blot probed for CNBP and nucleolin. Cropped blots are indicated by white separation.
Table S1: used in this study, Related to Experimental Procedures
Table S2: Antibodies used in this study, Related to Experimental Procedures
Table S3: Reagents used in this study, Related to Experimental Procedures
Table S4: r(G4C2)4 and b-isox condensed material, Related to Figure 3: Raw mass spectrometry data of proteins condensed by r(G4C2)4 and b-isox divided into three separate tabs, r(G4C2)4 for r(G4C2)4 condensed proteins, b-isox for b-isox condensed proteins, and comparison for a comparison of r(G4C2)4 and b-isox condensed proteins. For r(G4C2)4 and b-isox tabs, column C is the gene symbol, column D is the unique number of reads, column E is total number of reads. For comparison tab, column B is the gene symbol, column C is the unique number of reads for r(G4C2)4, column D is the total number of reads for r(G4C2)4, column E is the unique number of reads for b-isox, column F is the total number of reads for b-isox, and column G is the difference between the total number of r(G4C2)4 and b-isox reads.