

Abstract
Background
Although the elderly represents a rapidly growing population among transplant recipients, age-specific aspects have not been considered sufficiently in clinical trials. Moreover, age-specific effects of immunosuppressive therapies remain poorly understood.
Methods
Here, we assessed the impact of Rapamycin on alloimmune responses in old recipients using a fully MHC-mismatched murine transplantation model.
Results
Old untreated recipients displayed a prolonged skin graft survival compared to their young counterparts, an observation that confirmed data of our previous experiments. Rapamycin led to a significant prolongation of graft survival in both, young and old recipients. However, graft survival was age-dependent and extended in old vs. young recipients (19 days vs. 12 days, p=0.004). This age-specific effect was not linked to changes in frequencies or subset composition of either CD8+ or CD4+ T cells. Moreover, antiproliferative effects of Rapamycin on CD8+ and CD4+ T cells as assessed by in vivo BrdU-incorporation were comparable and age-independent. In contrast, the systemic production of IL-10 was markedly elevated in old recipients treated with Rapamycin. In parallel to this shift in cytokine balance, IFN-γ/IL-10 double-positive regulatory type 1 cells emerged during Th1-differentiation of old T helper cells in presence of Rapamycin. Similarly, CD4+IFN-γ+IL-10+ cells expanded among Foxp3-negative cells after in vivo treatment of old recipients with Rapamycin.
Conclusions
Our results highlight novel aspects of age-dependent immunosuppressive effects of Rapamycin, with relevance for age-specific immunosuppressive regimens.
Introduction
The clinical success of solid organ transplantation together with shifting demographics has encouraged the successful transplantation of older transplant recipients. In fact, recipients ≥ 65 years have tripled within the last 15 years, representing the fastest growing age segment.1 Currently, more than 50% of transplant recipients are ≥ 50 years old. While the consequences of immunosenescence are widely recognized, increased recipient age has been linked to less acute rejections and improved death-censored graft survival.2,3 Of additional relevance, elderly recipients carry higher risks for infections, cardiovascular diseases and malignancies, conditions that represent the 3 most common causes of death in transplant recipients ≥ 70 years.4,5 Clearly, immunosuppressants exacerbate age-related comorbidities,6 thus stressing the need for a more detailed understanding of the consequences of immunosenescence and alloimmunity with the aim to define key parameters and principles of an age-adapted immunosuppression.
Aging can affect all compartments of innate and adaptive immunity and does not necessarily represent a uniform deterioration of immunological capacity but rather individual shifts in function and regulation. On a cellular level, aging has been shown to result in a plethora of functional and phenotypic changes. For example, older graft recipients displayed significantly higher frequencies of T cells with an effector/memory phenotype, while in vitro capacities of proliferation and proinflammatory cytokine production were markedly reduced with increasing age.7–9 Experimentally, delayed rejection in older recipients was linked to fewer graft-infiltrating CD4+ T cells.10
Nevertheless, age-specific effects of current immunosuppressive drugs remain ill-understood. Moreover, older recipients have largely been excluded from clinical trials. Indeed, a large meta-analysis demonstrated that participants in clinical transplant trials had been significantly younger compared to the overall transplant recipient population in the US.11 Recognizing these shortcomings, the Food and Drug Administration (FDA) has recently encouraged clinical trials in the elderly to explore age-specific aspects of immunosuppressants.12
Rapamycin is a macrolide compound that has been established as a maintenance immunosuppressant in solid organ transplantation. The agent binds to the cytosolic protein FK-binding protein 12 (FKBP12) which inhibits the mTOR (mechanistic target of rapamycin) pathway by directly binding to mTOR Complex 1 (mTORC1).13 Rapamycin is a particularly relevant immunosuppressant in the elderly as it avoids the nephrotoxic side effects of calcineurin inhibitors such as Tacrolimus and Cyclosporine that may be more detrimental in older recipients frequently transplanted with older and/or extended criteria donor organs. Most recently, we were able to delineate age-specific metabolic aspects and age-specific modes of action when utilizing Tacrolimus.14
Here, we demonstrate in a fully MHC mismatched murine skin transplantation model that Rapamycin displays distinct age-specific mechanisms and immunosuppressive effectiveness through the emergence of IFN-γ/IL-10 double-positive regulatory type 1 cells.
Materials and Methods
Animals
Wild-type DBA/2 (H2d; 8-12 weeks) mice were purchased from Charles River Laboratory (Wilmington, MA USA). Young (male, 8-12 weeks) and old (male, 18 month) mice C57BL/6 (H2b) were obtained from the National Institute of Aging (NIA, Bethesda, MD, USA). Animals were allowed free access to water and standard chow. Experiments were approved by the Institutional Ethical Committee for Research on Animals. Use and care of animals were in accordance with National Institutes of Health and Institutional Animal Care and Use Committee guidelines.
Skin transplantation
Fully mismatched skin transplantations were performed; in detail, full-thickness skin grafts (1cm2) were obtained from tails of young DBA/2 mice and engrafted onto the dorsolateral thoracic walls of young and old C57BL/6 mice. Graft rejection was monitored daily and defined as necrosis exceeding 90%.
Rapamycin
Rapamycin was obtained from LC Laboratories, MA, USA. Rapamycin was dissolved in Dimethyl sulfoxide (DMSO; Sigma-Aldrich, MO, USA) and diluted with PBS. Young and old C57BL/6 skin transplant recipients were treated with daily intraperitoneal injections of control solution (PBS) or 1 mg/kg Rapamycin as indicated starting at the day of transplantation. For in vitro experiments, Rapamycin (500nM) was diluted in PBS and added to cell culture during the differentiation of naive CD4+ T cells.
Cell isolation and T cell differentiation
Single cell suspensions were obtained from spleens of young or old C57BL/6 mice; naive CD4+ T cells were isolated by negative selection (Stemcell Technologies, Vancouver, BC, Canada) using biotinylated antibodies directed against non-naive CD4+ T cells (CD8, CD11b, CD11c, CD19, CD24, CD25, CD44, CD45R, CD49b, TCRγ/δ, TER119) and streptavidin-coated magnetic particles. Cells were used at a purity > 95%.
Naive CD4+ T cells were cultured in 48-well plates (0.5 × 106 cells per well) suspended with 0.5ml of RPMI 1640 media supplemented with 10% fetal calf serum, 200mM L-glutamine, 100U ml-1 penicillin/streptomycin and 5 × 10-5M 2-mercaptoethanol at 37°C under 5% CO2 and 95% air atmosphere with saturated humidity. Wells were coated with 10 μg/ml antimouse α-CD3 and 2 μg/ml soluble antimouse α-CD28 was added to media. Cells were cultured in polarizing Th1 (50 ng/ml recombinant IL-12, 50 ng/ml recombinant IL-2, 100μg/ml anti-IL-6 and 100μg/ml anti-IL-4) and Treg (100ug/ml α-IL-12, 100ug/ml α-IL-4, 50ng/ml recombinant IL-2, 50ng/ml recombinant TGF-β) conditions. For Th0 conditions, naive CD4+ T cells were cultured with recombinant mouse IL-2 (50ng/ml) and in presence of α-CD3 and α-CD28 to maintain T cell survival. Rapamycin (500nM) diluted in PBS were added to wells as indicated. All recombinant cytokines and antibodies were purchased from eBioscience. After 96 hrs of culture, supernatants and cells were collected and analyzed by ELISA or flow cytometry, respectively. To determine absolute numbers of viable lymphocytes, Trypan staining was performed and live cells were counted using a Neubauer-improved counting chamber (Marienfeld Superior, Germany).
Flow cytometry
Single cell suspensions were obtained from spleens of recipient mice. Cells were labeled for surface and intracellular antigens with fluorescence-labeled α-CD4, α-CD8, α-CD62L, α-CD44, α-CD25, α-Foxp3, α-IFN-γ and α-IL-10 antimouse antibodies (eBioscience, CA). Intracellular staining for Foxp3 was performed with a commercially available staining kit, including permeabilization solution and buffer (eBioscience, CA). For intracellular cytokine staining, total splenocytes were seeded on 48-well plates and stimulated in complete media for 4 hrs at 37°C with phorbol 12-myristate 13-acetate (PMA; 50 ng/ml; Sigma-Aldrich), Ionomycin (500ng/ml; Sigma-Aldrich) and Brefeldin A (eBioscience). Subsequently, cells were fixed, permeabilized (kit by BD Biosciences, CA), and stained with respective antibodies. Intracellular cytokine staining after in vitro T cell differentiation was performed without restimulation. Flow cytometry measurements were performed on a FACS Canto II (BD Bioscience, CA, USA) and data were analyzed using FlowJo (FlowJo Software, OR, USA). For proper setting of gates, apoptotic cells were excluded and samples were compared with isotype controls, fluorescence minus 1 (FMO) stainings, permeabilized and unpermeabilized unstained cells. Isotype controls shown in Figures 4, 5, 6 were obtained from cells of young, untreated animals not subjected to restimulation and stained with the entire panel (replacing the cytokine in question with respective isotype control antibodies). For in vitro studies (Figure 6), young untreated cells were used after differentiation.
Figure 4.
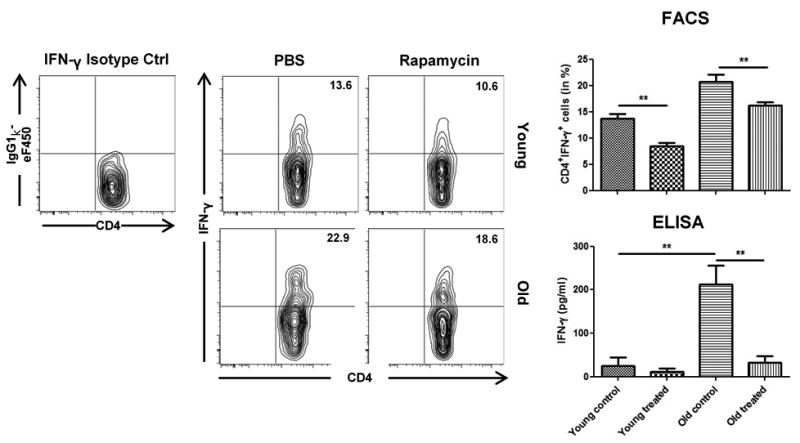
Rapamycin decreases expression of IFN-γ in old CD4+ T cells following skin transplantation. Skin graft recipients were treated daily with either PBS or Rapamycin. By day 7, splenic frequencies of CD4+IFN-γ+ T cells were assessed by flow cytometry using isotype controls. Representative plots are given for each group. Furthermore, cell culture supernatants after restimulation of recipient splenocytes with donor-type splenocytes for 72 hrs were analyzed for IFN-γ by ELISA. (n=5-6 animals/group; data represent mean ± SD; *, p<0.05; **, p<0.01; ***, p<0.001)
Figure 5.
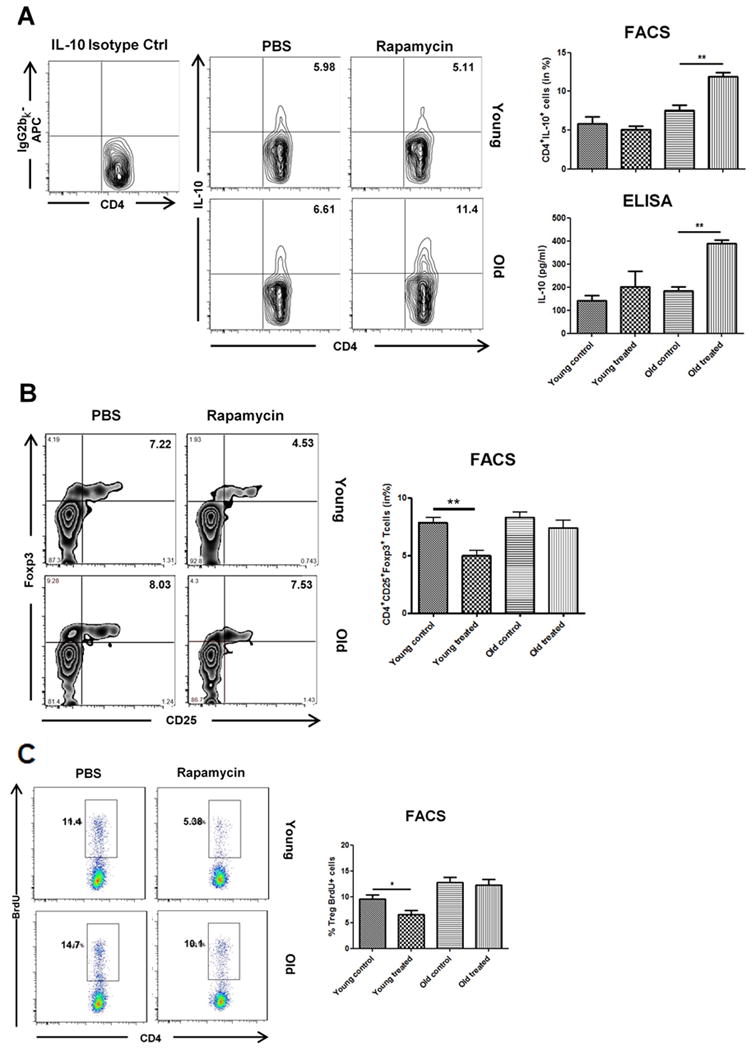
Impact of Rapamycin on the expression of IL-10 and frequencies and proliferation of regulatory T cells. Allograft recipients were treated daily with either PBS or Rapamycin. By day 7, splenic frequencies of (A) CD4+IL-10+ were then assessed by flow cytometry using isotype controls and cell culture supernatants after restimulation of recipient splenocytes with donor-type splenocytes for 72 hrs were analyzed for IL-10 using ELISA assays. (B) Spleic frequencies of CD4+CD25+Foxp3+ T cells were assessed by flow cytometry by day 7. (C) Animals received a single intraperitoneal injection of BrdU immediately prior to skin transplantation and subsequent treatment with Rapamycin or PBS. Splenic frequencies of BrdU+ cells were analyzed at day 6 by flow cytometry within CD4+CD25+Foxp3+ Tregs. Representative plots are given for each group. (n=5-6 animals/group; data represent mean ± SD; *, p<0.05; **, p<0.01; ***, p<0.001)
Figure 6.
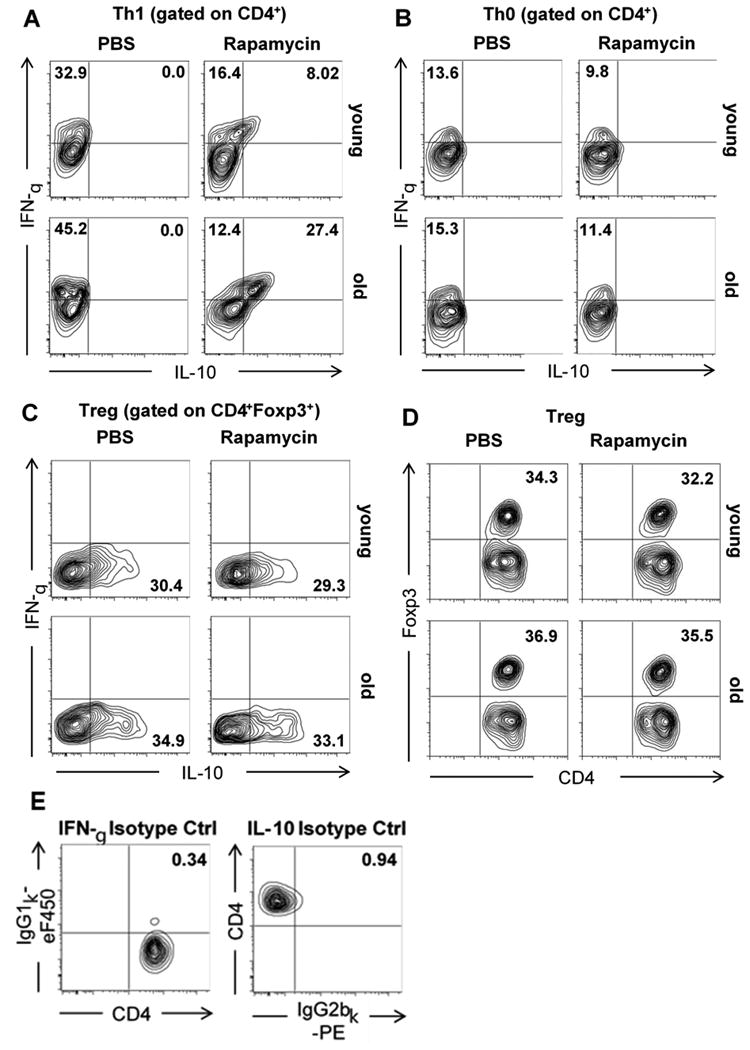
Presence of Rapamycin leads to expansion of double-positive IFN-γ+IL-10+ cells in naive old CD4+ T cells cultured under Th1 polarizing conditions. Single cell suspensions were obtained from spleens of young and old C57BL/6 mice and naive CD4+ T cells were isolated by negative selection. Naive T cells were then cultured at 500.000/well under (A) Th1, (B) Th0, and (C, D) Treg polarizing conditions in the presence or absence of Rapamycin. After 96h, cells were stained for CD4 and intracellular expression of (A-C) IFN-γ and IL-10 or (D) Foxp3, respectively. (E) Isotype controls for intracellular cytokines were used for proper setting of gates. Representative plots are given for each group (n=5-6 animals/group).
Bromdesoxyuridine (BrdU) proliferation assay
To assess T cell proliferation in vivo, recipient mice treated with control solution (PBS) or Rapamycin received an intraperitoneal injection of 2 mg of BrdU solution (BD Biosciences, CA) at the day of transplantation. Animals were euthanized by day 6 after transplantation; single cell suspensions of splenocytes were obtained and stained with APC-labeled anti-BrdU antibody per the manufacturer's protocol (BD Biosciences, CA) and subsequently analyzed by FACS.
ELISA
INF-γ and IL-10 were measured in supernatants after T helper cell differentiation or after restimulation of recipient splenocytes with donor-type splenocytes for 72 hrs. Commercial ELISA kits (eBioscience) were used according to the manufacturer's instructions. Briefly, ELISA plates were coated with 100 μL anticytokine capture antibody at 4°C for 12 hrs; plates were then washed ×5 with 0.05% PBS-Tween and coated for 1 hr with the blocking buffer provided by the manufacturer. Samples or cytokine standards were added in duplicates (100 μL per well) and incubated at 4°C for 12 hrs. Subsequently, wells were washed ×5 with PBS-Tween and incubated with 100 μL anticytokine detection antibody at 4°C overnight; wells were then washed again ×5 with PBS-Tween and incubated with 100 μL avidin-horseradish peroxidase at room temperature for 30 min. Thereafter, wells were washed ×7 with PBS-Tween and then incubated with 100 μL per well of substrate. The reaction was stopped after 15 minutes with 1 mol/L H2SO4, and absorbance was measured with a multiplate microplate fluorescence reader (Synergy HT, Biotek, Winooski, VT) at 405 nm.
Statistics
Differences between groups were analyzed using unpaired Student's t test. Graft survival was compared by Log-rank test. A p value < 0.05 was considered significant (*p<0.05; **p<0.01; ***p<0.001).
Results
Rapamycin impacts graft survival in an age-specific fashion
Immunosenescence has been linked in studies by ourselves and others with compromised alloimmune responses and prolonged allograft survival in old recipients.15 Here, we transplanted fully MHC-mismatched skin grafts to assess age-specific effects on allograft survival after treatment with Rapamycin (Figure 1A). In line with previous observations, old untreated mice displayed a prolonged allograft survival (median survival time 9 vs. 7 days in untreated old vs. young recipients, p=0.006). Treatment with Rapamycin extended graft survival in both, young and old animals. Of note, prolongation of graft survival was particularly pronounced and highly significant in old recipients (from 9 to 19 days, p<0.0001) compared to a prolongation of 7 to 12 days in young recipients. These data indicated age-specific immunosuppressive effects of Rapamycin on allograft survival in old recipients.
Figure 1.
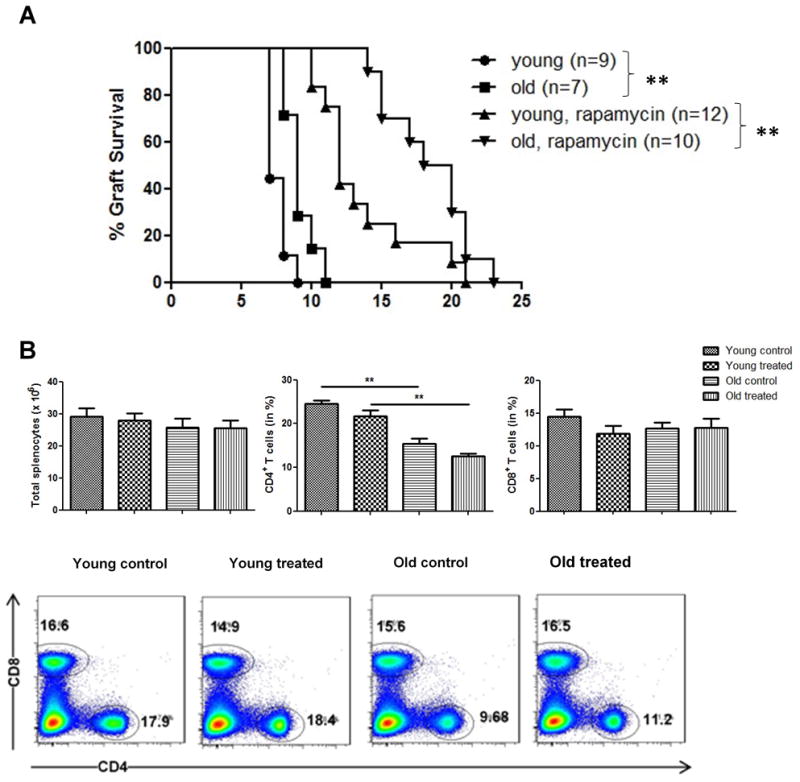
Survival of fully mismatched skin grafts following administration of Rapamycin; recipient splenic cell counts and CD4+/CD8+ T cell frequencies. (A) Skin allografts from young DBA mice were transplanted onto young or old C57BL/6 mice. Recipients received daily intraperitoneal injections of either control solution (PBS) or Rapamycin and graft survival was monitored daily (n=7-12 animals/group). (B) Spleens were collected by day 7 after transplantation and single cell suspensions of splenocytes were obtained. Total cell count was determined and frequencies of CD4+ and CD8+ T cells were assessed using flow cytometry (n=5-6 animals/group). Log-rank test was used to compare graft survival; data represent mean ± SD; *, p<0.05; **, p<0.01; ***, p<0.001.
Rapamycin does not alter T cell subsets in old recipients
It is well established that the broad changes linked to aging affect mechanisms of allorecognition and rejection.3 To dissect the effects of Rapamycin on adaptive T cell responses, we assessed total splenocyte counts and characterized the T cell compartment in allograft recipients treated with Rapamycin (Figure 1B). While all groups showed comparable and age-independent total splenic cell counts, frequencies of CD4+ T cells declined with aging, an observation that confirmed data of our own previous studies.15 More importantly, treatment with Rapamycin did not impact frequencies of CD4+ and CD8+ T cells in either young or old recipients.
Next, we performed a detailed analysis of CD4+ and CD8+ T cell subsets and observed that Rapamycin could reduce frequencies of central memory (CD62L+CD44+) CD4+ T cells in young but not old recipients (Figure 2A). Moreover, CD8+ T cell subsets remained unchanged regardless of treatment and recipient age (Figure 2B). Of note, old recipients in both, untreated and Rapamycin-treated animals had lower frequencies of naive (CD62L+CD44-) CD4+ and CD8+ T cells in accordance with previous observations.15–17
Figure 2.
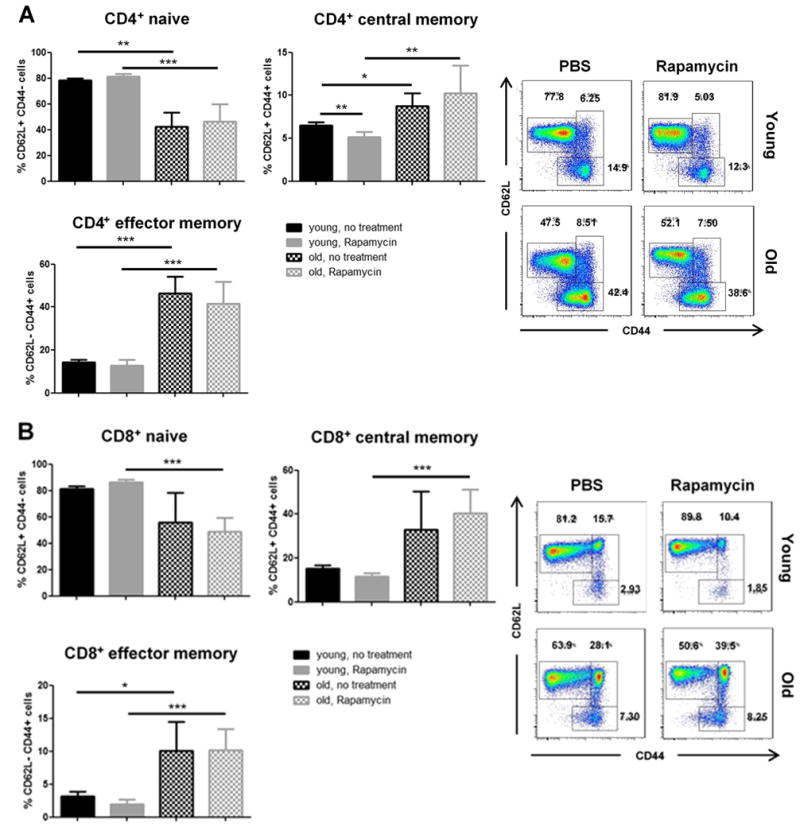
T cell subsets in young and old graft recipients treated with Rapamycin. Allograft recipients were treated daily with PBS or Rapamycin. By day 7, splenic frequencies of (A) CD4+CD62L+CD44- (naïve), CD4+CD62L+CD44+ (central memory) and CD4+CD62L-CD44+ (effector memory) T cells in addition to (B) CD8+CD62L+CD44-, CD8+CD62L+CD44+ and CD8+CD62L-CD44+ T cells were assessed by flow cytometry. Representative plots are given for each group. (n=5-6 animals/group; data represent mean ± SD; *, <0.05; **, p<0.01; ***, p<0.001)
Taken together, despite conferring a marked graft survival benefit in old recipients, treatment with Rapamycin did not affect the composition of T cell subsets in old recipients.
Antiproliferative effects of Rapamycin are age-independent
Antiproliferative effects on T cells through mTOR inhibition represent a major aspect of immunosuppressive capacities of Rapamycin. We thus set out to assess the impact of Rapamycin on the proliferation of young and old T cells in vivo after allogenic stimuli. Here, transplant recipients received an intraperitoneal dose of Bromdesoxyuridine (BrdU) immediately prior to skin grafting and BrdU incorpration was subsequently measured at day 6 by flow cytometry. Extensive analysis of CD4+ T cell subsets revealed that Rapamycin reduced the proliferation of CD4+ T cells in an age-independent fashion throughout all subsets (Figure 3A). Moreover, proliferation of CD8+ T cells was also uniformly suppressed in an age-dependent pattern (Figure 3B). Thus, although we observed particularly pronounced effects of Rapamycin in older recipients, our data indicated that effects were not linked to age-dependent effects on either the composition or the proliferation of specific T cell subsets.
Figure 3.
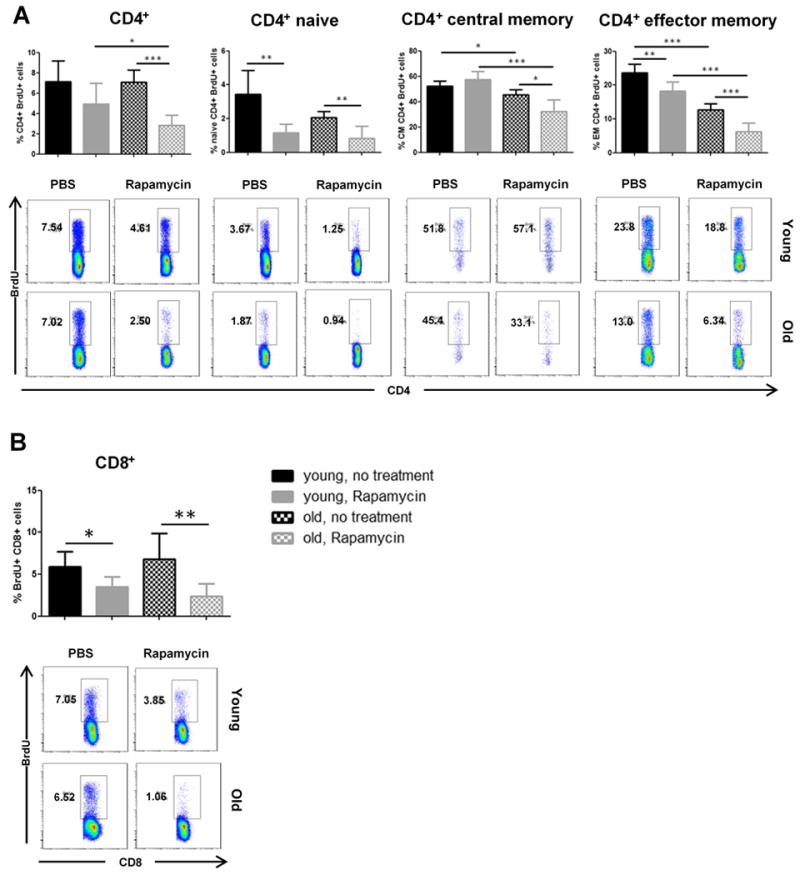
Impact of Rapamycin on in vivo proliferation of young and old T cells. Animals received a single intraperitoneal injection of BrdU immediately prior to skin transplantation and were then treated daily with either PBS or Rapamycin. Six days after transplantation, splenic frequencies of BrdU+ cells were analyzed by flow cytometry within (A) total CD4+ and CD4+CD62L+CD44- (naïve), CD4+CD62L+CD44+ (central memory) and CD4+CD62L-CD44+ (effector memory) as well as (B) total CD8+ and CD8+CD62L+CD44-, CD8+CD62L+CD44+ and CD8+CD62L-CD44+ T cell populations. Representative plots are given for each group. (n=5-6 animals/group; data represent mean ± SD; *, p<0.05; **, p<0.01; ***, p<0.001)
Rapamycin affects systemic cytokine balances
CD4+ T cell differentiation and cytokine profiles play a critical role in regulating immune responses. IFN-γ and IL-10, cytokines with pro and antiinflammatory capacities, respectively, have been identified as key cytokines that regulate alloimmunity.18 We therefore tested whether the effects of Rapamycin on graft survival were linked to a modified balance of systemic IFN-γ or IL-10 produced by CD4+ T cells.
Treatment with Rapamycin was linked to a reduction in systemic IFN-γ production in both, young and old recipients as assessed by frequencies of CD4+IFN-γ+ T helper cells (Figure 4). These findings were confirmed by an analysis of cell culture supernatants using ELISA assays after the restimulation of recipient splenocytes with donor type antigen. Strikingly, systemic IL-10 production was markedly increased in old recipients treated with Rapamycin (Figure 5A); this observation was also confirmed by ELISA. As CD4+CD25+Foxp3+ regulatory T cells (Tregs) represent a major T helper cell subset contributing to systemic IL-10 production, we assessed systemic frequencies of Tregs after skin transplantation and Rapamycin treatment. Surprisingly, Rapamycin treatment did not impact frequencies and proliferative responses of regulatory T cells in old animals (Figure 5B-C).
Taken together, these results demonstrated a shift in systemic cytokine balances in old, but not young transplant recipients with a reduction in systemic IFN-γ levels and a marked systemic increase in IL-10 production after the treatment with Rapamycin.
Rapamycin favors expansion of CD4+IFN-γ+IL-10+ cells during T helper cell differentiation
Next, we tested whether age-dependent in vivo shifts in the balance of systemic levels of IFN-γ and IL-10 could be traced back to effects of Rapamycin on T helper cell differentiation. Naive CD4+ T cells were isolated from young and old recipient-type C57BL/6 mice and cultured in Th1 and Treg polarizing conditions as well as Th0 control conditions in the presence of Rapamycin, compared to untreated controls. Supernatants of cell cultures collected by 96 hours demonstrated a significant suppression of IFN-γ production in both, young and old T cells in presence of Rapamycin under Th1 conditions (Figure S1A). Moreover, we detected a marked increase in IL-10 production of old, Rapamycin-treated T cells in Th1 compared to Th0 polarizing conditions (Figure S1A). Analysis of IFN-γ+ and/or IL-10+ T helper cells by flow cytometry revealed a parallel increase in CD4+IFN-γ+IL-10+ double-positive cells in old recipients treated with Rapamycin in Th1 but not Th0 conditions (Figure 6A and B). Of note, double-positive CD4+IFN-γ+IL-10+ cells, recently characterized as regulatory type 1 cells, display potent immunosuppressive properties and are able to promote allograft survival independently of CD4+CD25+Foxp3+ regulatory T cells.19,20 Calculating absolute numbers of CD4+IFN-γ+IL-10+ double-positive cells using the total cell count of wells and frequencies obtained by flow cytometry confirmed the specific expansion of this population among old T helper cells cultured in presence of Rapamycin (Figure S1B and C). In addition, Rapamycin did not significantly affect expression of IL-10 or Foxp3 in naive CD4+ T cells cultured under Treg polarizing conditions (Figure 6C and D). Thus, these results suggest that Rapamycin exerts age-specific immunosuppressive effects in old T cells that are linked to promotion of regulatory type 1 cells during T helper cell type 1 differentiation, and independent of CD4+CD25+Foxp3+ regulatory T cells.
Rapamycin treatment induces increased frequencies of Tr1 cells in old recipients
To test whether the effects of Rapamycin on T helper cell differentiation in vitro also lead to the emergence of Tr1 cells, we treated young and old skin recipients with daily injections of Rapamycin and performed an extensive cytokine analysis in CD4+ T cells by 7 days. While frequencies of CD4+CD25+Foxp3+ regulatory T cells remained stable in both age groups under Rapamycin treatment (Figure 7A), we detected a distinct population of CD4+IFN-γ+IL-10+ cells among Foxp3-negative cells in old recipients (Figure 7B). Additional flow cytometric analysis revealed that this double-positive population did not express significant intracellular amounts of IL-2 or IL-4 (Figure 7C), thus meeting the critical phenotypic criteria of regulatory type 1 cells.21 Taken together, these results suggest that treatment of old skin transplant recipients with Rapamycin leads to pronounced prolongation of graft survival associated with the induction of CD4+IFN-γ+IL-10+ Tr1 cells and independently of regulatory T cells.
Figure 7.
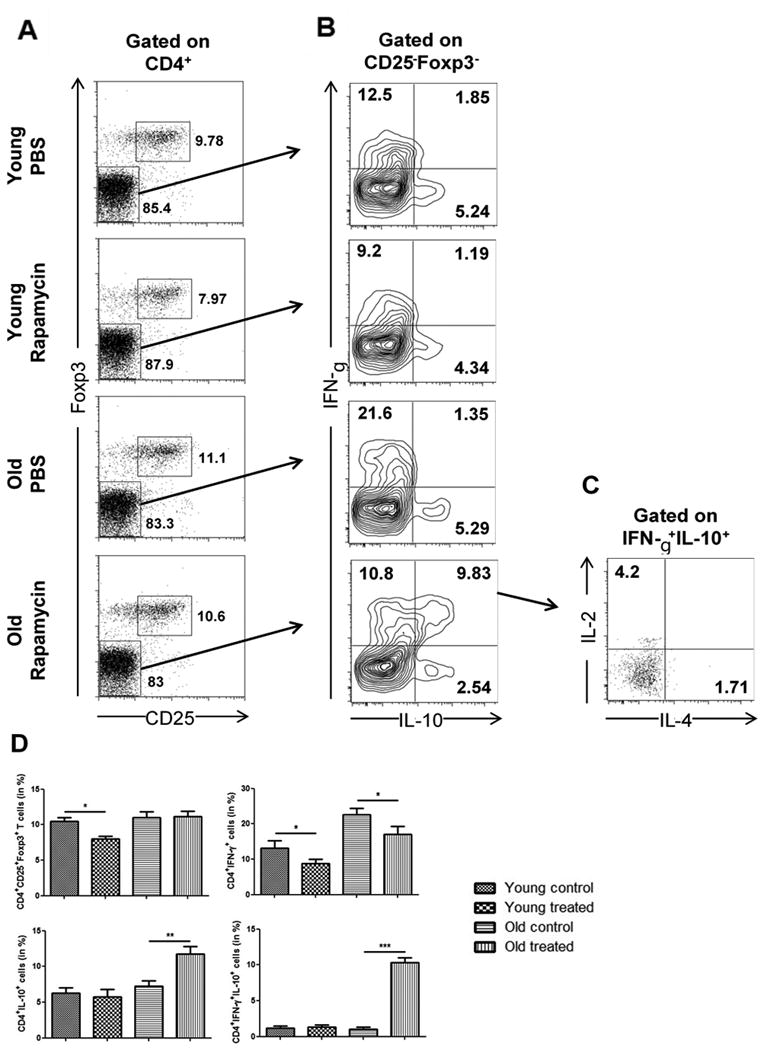
Emergence of CD4+IFN-γ+IL-10+ Tr1 cells in vivo during treatment of old allograft recipients with Rapamycin. Recipients received daily treatments with PBS or Rapamycin following skin grafting. By day 7, single cell suspensions were obtained from spleens and (A) frequencies of CD4+CD25+Foxp3+ regulatory T cells were analyzed. (B) Gated on CD25-Foxp3- non-Treg cells, intracellular expression of IL-10 and IFN-γ was assessed after restimulation with PMA/Ionomycin for 4 hrs. Finally, (C) intracellular expression of IL-2 (4.075 ± 0.67) and IL-4 (1.2025 ± 0.54) was analyzed among CD4+IFN-γ+IL-10+ cells in old treated animals. (D) Statistical analyses of cell frequencies as determined by flow cytometry. Representative plots are given for each group (n=6 animals/group; data represent mean ± SD; *, p<0.05; ***, p<0.001).
Discussion
The recent 2 decades have seen a significant shift towards increasing numbers of older solid organ transplant recipients, emphasizing the significance of understanding the consequences of immunosenescence on alloimmunity and subsequent effects on transplant outcomes. Clinically, we have demonstrated that increasing recipient age is linked to reduced acute rejection rates.22 In previous studies, we have dissected age-specific effects on alloimmune responses facilitated by CD4+ and CD8+ T cells.15,16 Most recently, we have also been able to show age-specific effects when applying Tacrolimus.14 Although the majority of transplant recipients is > 50 years and over 20% patients on waiting lists are older than 65 years, clinical trials have mostly excluded older patients.2 In addition to showing less acute rejections, side-effects of immunosuppressants appear to be exacerbated in older recipients.4,5
In the current study, we were able to delineate age-specific effects of Rapamycin, a commonly used immunosuppressant, on alloimmune responses and graft survival that represent critical contributions when caring for an overall aging recipient population.
Rapamycin has previously been highlighted as a favorable component of immunosuppressive regimens in the elderly due to its well-established antitumor and antiproliferative capacities. Indeed, clinical data demonstrate that de novo posttransplant malignancies were significantly lower with mTOR-inhibitors compared to calcineurin inhibitor-based immunosuppression.23 Rapamycin has furthermore recently been reported to reduce markers of cellular senescence and systemic inflammation in elderly patients with improvements in physical performance, effects that may also be mediated through the induction of autophagy.24,25 Moreover, Rapamycin is the first pharmacological agent that has been shown to extend lifespan in several experimental animal models.26,27 In our present study, Rapamycin more than doubled median graft survival in old recipients (Figure 1A), demonstrating that Rapamycin favors allograft survival in old recipients. These results add new insights to the unique role of Rapamycin in aging. On a molecular level, our results were associated with a significant reduction of IFN-γ production by CD4+ T cells after Rapamycin treatment (Figure 4). More importantly, old graft recipients also exhibited a significant increase in CD4+IL-10+ T cells (Figure 5A). In support of the specific effects of Rapamycin during immunosenescence, recent reports have shown even inhibitory effects on IL-10 production and signaling in young mice.13,28
Of note, Rapamycin did not promote CD4+CD25+Foxp3+ T cells in our model and even led to decreased frequencies in young animals (Figure 5B). So far, both Treg expansion or lack of Treg expansion have been reported upon treatment with Rapamycin.29–31 Interestingly, a detailed study by Valmori et al, demonstrated the capacity of Rapamycin to induce regulatory functions in conventional CD4+ T cells independent of Tregs, however without providing mechanistic insights into how these effects are mediated.32 Here, we show that the presence of Rapamycin during differentiation of old naive T cells in Th1-polarizing conditions gave rise to a distinct population of CD4+IFN-γ+IL-10+ regulatory type 1 cells (Figure 6), findings that we could also confirm in vivo (Figure 7). Battaglia et al, have previously reported that Rapamycin in presence of IL-10 can favor tolerance through the induction of Tr1 cells in a model of islet cell transplantation.30 Of note, the same group also successfully generated donor-specific Tr1 cells for cell therapy after kidney transplantation by coculturing interleukin-10–producing dendritic cells obtained from healthy donors with peripheral blood mononuclear cells of patients on dialysis.33 With increased systemic levels of IL-10 in aging reported both clinically and in murine models,9,34 our data thus point towards a novel pathway linking changes in cytokine balances to age-specific immunosuppressive aspects of Rapamycin.
As old IL-10 knockout mice show an altered lymphocyte and myeloid profiles and are not available in aged colonies,35 we have been unable to confirm the causal role of Tr1 cells in old transplant recipients treated with Rapamycin. Moreover, comparing data in IL-10 knockout mice with those when applying a purified anti-IL-10 monoclonal antibody 20,36 suggested that antibody treatment does not provide a sufficient neutralization of IL-10, possibly related to high systemic cytokine levels in addition to a redundant production of IL-10 by various different cell types in the context of transplantation.37 In addition, reliable surface markers for flow sorting of Tr1 cells apart from intracellular expression of cytokines and/or transcription factors remain to be established.21
Taken together, our results demonstrate that Rapamycin favors allograft survival in old transplant recipients independent of effects on frequencies or proliferative rates of T cell subsets. Moreover, Rapamycin led to a systemic increase in IL-10 production in old recipients associated with an age-specific induction of regulatory type 1 cells, independent of regulatory T cells. Rapamycin may therefore serve as an immunosuppressant with specific advantages for the treatment of elderly transplant recipients.
Supplementary Material
Acknowledgments
Funding: S.G.T. was supported by grants from the National Institutes of Health (RO1AG039449). M.Q. (QU 420/1-1), T.H. (HE 7457/1-1) and K.E. (KFO243/1) were supported by the German Research Foundation (DFG). K.M. was supported by Osaka Medical College Foundation.
Abbreviations
- BrdU
Bromdesoxyuridine
- CD
cluster of differentiation
- ELISA
enzyme-linked immunosorbent assay
- IFN
interferon
- IL
interleukin
- MHC
major histocompatibility complex
- mTOR
mechanistic target of rapamycin
- Th1
T helper type 1
- Treg
regulatory T cell
- PBS
phosphate buffered saline
Footnotes
Authorship: M.Q. and T.H. performed experiments, analyzed data and wrote the manuscript. K.E. performed experiments and analyzed data. K.M., H.U, and Y.N. helped performing experiments. A.E. designed and performed experiments. R.A. and H.A. helped with preparation of the manuscript. S.G.T. designed experiments and wrote the manuscript.
Disclosures: The authors declare no conflicts of interest.
References
- 1.US Department of Health and Human Services Human Respirces and Services Administration. OPTN/SRTR 2012 Annual Data Report. https://srtr.transplant.hrsa.gov/annual_reports/2012/pdf/2012_SRTR_ADR.pdf. Published June 2014.
- 2.Wolfe RA, Roys EC, Merion RM. Trends in organ donation and transplantation in the United States, 1999-2008. Am J Transplant. 2010;10(4 Pt 2):961–972. doi: 10.1111/j.1600-6143.2010.03021.x. [DOI] [PubMed] [Google Scholar]
- 3.Heinbokel T, Elkhal A, Liu G, Edtinger K, Tullius SG. Immunosenescence and organ transplantation. Transplant Rev (Orlando) 2013;27(3):65–75. doi: 10.1016/j.trre.2013.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Karim A, Farrugia D, Cheshire J, et al. Recipient age and risk for mortality after kidney transplantation in England. Transplantation. 2014;97(8):832–838. doi: 10.1097/01.TP.0000438026.03958.7b. [DOI] [PubMed] [Google Scholar]
- 5.Bradley BA. Rejection and recipient age. Transpl Immunol. 2002;10(2-3):125–132. doi: 10.1016/s0966-3274(02)00058-8. [DOI] [PubMed] [Google Scholar]
- 6.Taylor AL, Watson CJE, Bradley JA. Immunosuppressive agents in solid organ transplantation: mechanisms of action and therapeutic efficacy. Crit Rev Oncol Hematol. 2005;56(1):23–46. doi: 10.1016/j.critrevonc.2005.03.012. [DOI] [PubMed] [Google Scholar]
- 7.Saule P, Trauet J, Dutriez V, Lekeux V, Dessaint JP, Labalette M. Accumulation of memory T cells from childhood to old age: central and effector memory cells in CD4(+) versus effector memory and terminally differentiated memory cells in CD8(+) compartment. Mech Ageing Dev. 2006;127(3):274–281. doi: 10.1016/j.mad.2005.11.001. [DOI] [PubMed] [Google Scholar]
- 8.Murasko DM, Weiner P, Kaye D. Decline in mitogen induced proliferation of lymphocytes with increasing age. Clin Exp Immunol. 1987;70(2):440–448. [PMC free article] [PubMed] [Google Scholar]
- 9.Bernstein ED, Murasko DM. Effect of age on cytokine production in humans. Age (Omaha) 1998;21(4):137–151. doi: 10.1007/s11357-998-0024-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Tullius SG, Tran H, Guleria I, Malek SK, Tilney NL, Milford E. The combination of donor and recipient age is critical in determining host immunoresponsiveness and renal transplant outcome. Ann Surg. 2010;252(4):662–674. doi: 10.1097/SLA.0b013e3181f65c7d. [DOI] [PubMed] [Google Scholar]
- 11.Blosser CD, Huverserian A, Bloom RD, et al. Age, exclusion criteria, and generalizability of randomized trials enrolling kidney transplant recipients. Transplantation. 2011;91(8):858–863. doi: 10.1097/TP.0b013e31820f42d9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.U.S. Government Accountability Office. Prescription drugs: FDA guidance and regulations related to data on elderly persons in clinical drug trials. http://www.gao.gov/assets/100/95182.pdf. Published September 28, 2007.
- 13.Thomson AW, Turnquist HR, Raimondi G. Immunoregulatory functions of mTOR inhibition. Nat Rev Immunol. 2009;9(5):324–337. doi: 10.1038/nri2546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Krenzien F, Quante M, Heinbokel T, et al. Age-Dependent Metabolic and Immunosuppressive Effects of Tacrolimus. Am J Transplant. 2017;17(5):1242–1254. doi: 10.1111/ajt.14087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Denecke C, Bedi DS, Ge X, et al. Prolonged graft survival in older recipient mice is determined by impaired effector T-cell but intact regulatory T-cell responses. PLoS One. 2010;5(2):e9232. doi: 10.1371/journal.pone.0009232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bedi DS, Krenzien F, Quante M, et al. Defective CD8 signaling pathways delay rejection in older recipients. Transplantation. 2016;100(1):69–79. doi: 10.1097/TP.0000000000000886. [DOI] [PubMed] [Google Scholar]
- 17.Appay V, Sauce D. Naive T cells: the crux of cellular immune aging? Exp Gerontol. 2014;54:90–93. doi: 10.1016/j.exger.2014.01.003. [DOI] [PubMed] [Google Scholar]
- 18.Zhu J, Yamane H, Paul WE. Differentiation of effector CD4 T cell populations. Annu Rev Immunol. 2010;28:445–489. doi: 10.1146/annurev-immunol-030409-101212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Gagliani N, Magnani CF, Huber S, et al. Coexpression of CD49b and LAG-3 identifies human and mouse T regulatory type 1 cells. Nat Med. 2013;19(6):739–746. doi: 10.1038/nm.3179. [DOI] [PubMed] [Google Scholar]
- 20.Elkhal A, Rodriguez Cetina Biefer H, Heinbokel T, et al. NAD(+) regulates Treg cell fate and promotes allograft survival via a systemic IL-10 production that is CD4(+) CD25(+) Foxp3(+) T cells independent. Sci Rep. 2016;6:22325. doi: 10.1038/srep22325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Zeng H, Zhang R, Jin B, Chen L. Type 1 regulatory T cells: a new mechanism of peripheral immune tolerance. Cell Mol Immunol. 2015;12(5):566–571. doi: 10.1038/cmi.2015.44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Tullius SG, Milford E. Kidney allocation and the aging immune response. N Engl J Med. 2011;364(14):1369–1370. doi: 10.1056/NEJMc1103007. [DOI] [PubMed] [Google Scholar]
- 23.Kauffman HM, Cherikh WS, Cheng Y, Hanto DW, Kahan BD. Maintenance immunosuppression with target-of-rapamycin inhibitors is associated with a reduced incidence of de novo malignancies. Transplantation. 2005;80(7):883–889. doi: 10.1097/01.tp.0000184006.43152.8d. [DOI] [PubMed] [Google Scholar]
- 24.Singh M, Jensen MD, Lerman A, et al. Effect of Low-Dose Rapamycin on Senescence Markers and Physical Functioning in Older Adults with Coronary Artery Disease: Results of a Pilot Study. J frailty aging. 2016;5(4):204–207. doi: 10.14283/jfa.2016.112. [DOI] [PubMed] [Google Scholar]
- 25.Kim YC, Guan KL, Laplante M, et al. mTOR: a pharmacologic target for autophagy regulation. J Clin Invest. 2015;125(1):25–32. doi: 10.1172/JCI73939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Harrison DE, Strong R, Sharp ZD, et al. Rapamycin fed late in life extends lifespan in genetically heterogeneous mice. Nature. 2009 Jul; doi: 10.1038/nature08221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ehninger D, Neff F, Xie K. Longevity, aging and rapamycin. Cell Mol Life Sci. 2014;71(22):4325–4346. doi: 10.1007/s00018-014-1677-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Foldenauer MEB, McClellan SA, Berger EA, Hazlett LD. Mammalian target of rapamycin regulates IL-10 and resistance to pseudomonas aeruginosa corneal infection. J Immunol. 2013;190(11):5649–5658. doi: 10.4049/jimmunol.1203094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Lim DG, Joe IY, Park YH, et al. Effect of immunosuppressants on the expansion and function of naturally occurring regulatory T cells. Transpl Immunol. 2007;18(2):94–100. doi: 10.1016/j.trim.2007.05.005. [DOI] [PubMed] [Google Scholar]
- 30.Battaglia M, Stabilini A, Draghici E, et al. Rapamycin and interleukin-10 treatment induces T regulatory type 1 cells that mediate antigen-specific transplantation tolerance. Diabetes. 2006;55(1):40–49. [PubMed] [Google Scholar]
- 31.Lu L, Qian XF, Rao JH, Wang XH, Zheng SG, Zhang F. Rapamycin Promotes the Expansion of CD4+ Foxp3+ Regulatory T Cells After Liver Transplantation. Transplant Proc. 2010;42(5):1755–1757. doi: 10.1016/j.transproceed.2009.10.008. [DOI] [PubMed] [Google Scholar]
- 32.Valmori D, Tosello V, Souleimanian NE, et al. Rapamycin-mediated enrichment of T cells with regulatory activity in stimulated CD4+ T cell cultures is not due to the selective expansion of naturally occurring regulatory T cells but to the induction of regulatory functions in conventional CD4+ T cells. J Immunol. 2006;177(2):944–949. doi: 10.4049/jimmunol.177.2.944. [DOI] [PubMed] [Google Scholar]
- 33.Petrelli A, Tresoldi E, Mfarrej BG, et al. Generation of donor-specific T regulatory type 1 cells from patients on dialysis for cell therapy after kidney transplantation. Transplantation. 2015;99(8):1582–1589. doi: 10.1097/TP.0000000000000751. [DOI] [PubMed] [Google Scholar]
- 34.Hobbs MV, Weigle WO, Ernst DN. Interleukin-10 production by splenic CD4+ cells and cell subsets from young and old mice. Cell Immunol. 1994;154(1):264–272. doi: 10.1006/cimm.1994.1076. [DOI] [PubMed] [Google Scholar]
- 35.Kühn R, Löhler J, Rennick D, Rajewsky K, Müller W. Interleukin-10-deficient mice develop chronic enterocolitis. Cell. 1993;75(2):263–274. doi: 10.1016/0092-8674(93)80068-p. [DOI] [PubMed] [Google Scholar]
- 36.Tullius SG, Biefer HRC, Li S, et al. NAD+ protects against EAE by regulating CD4+ T-cell differentiation. Nat Commun. 2014;5:5101. doi: 10.1038/ncomms6101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Marlow GJ. Why interleukin-10 supplementation does not work in Crohn's disease patients. World J Gastroenterol. 2013;19(25):3931. doi: 10.3748/wjg.v19.i25.3931. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.