

Abstract
Traumatic brain injury (TBI) is characterized by primary damage to the brain from the external mechanical force and by subsequent secondary injury due to various molecular and pathophysiological responses that eventually lead to neuronal cell death. Secondary brain injury events may occur minutes, hours, or even days after the trauma, and provide valuable therapeutic targets to prevent further neuronal degeneration. At the present time, there is no effective treatment for TBI due, in part, to the widespread impact of numerous complex secondary biochemical and pathophysiological events occurring at different time points following the initial injury. MicroRNAs control a range of physiological and pathological functions such as development, differentiation, apoptosis and metabolism, and may serve as potential targets for progress assessment and intervention against TBI to mitigate secondary damage to the brain. This has implications regarding improving the diagnostic accuracy of brain impairment and long-term outcomes as well as potential novel treatments. Recent human studies have identified specific microRNAs in serum/plasma (miR-425-p, -21, -93, -191 and -499) and cerebro-spinal fluid (CSF) (miR-328, -362-3p, -451, -486a) as possible indicators of the diagnosis, severity, and prognosis of TBI. Experimental animal studies have examined specific microRNAs as biomarkers and therapeutic targets for moderate and mild TBI (e.g., miR-21, miR-23b). MicroRNA profiling was altered by voluntary exercise. Differences in basal microRNA expression in the brain of adult and aged animals and alterations in response to TBI (e.g., miR-21) have also been reported. Further large-scale studies with TBI patients are needed to provide more information on the changes in microRNA profiles in different age groups (children, adults, and elderly).
Keywords: traumatic brain injury, microRNAs, diagnostic markers, therapeutic targets, humans, animal models
Introduction
Traumatic brain injury (TBI) due to a violent blow to the head is the leading cause of death and disability in adults under the age of 45 in Western countries (Jennett, 1998; Maas et al., 2008; Ribbers, 2010) with a very high number of brain injuries resulting from motor vehicle accidents and sporting activities. There has also been a marked increase in the incidence of fall-related TBI in persons aged ≥ 75 (Adekoya et al., 2002). The severity of TBI is determined by the nature, speed and location of the impact, and by complications such as hypotension, intracranial hemorrhage or increased intracranial pressure. TBI is characterized by primary damage to the brain from the external mechanical force and by subsequent secondary injury due to various molecular and pathophysiological responses that eventually lead to neuronal cell death. These responses include brain edema, intracranial hypertension, and subsequent neurological dysfunction (Zweckberger et al., 2006). Secondary brain injury events may occur minutes, hours, or even days after the trauma (Ribbers, 2010), and provide valuable therapeutic targets to prevent further neuronal degeneration. However, treatment options for secondary brain damage have failed to show efficacy in clinical trials (Maas et al., 1999), thereby emphasizing a need for new therapeutic targets.
MicroRNAs are short non-coding RNAs (20–24 nucleotides) that negatively regulate gene expression at the post-transcriptional level by binding to the 3′-untranslated region (UTR) of target mRNAs which leads to degradation of the mRNA or direct inhibition of mRNA translation (Baek et al., 2008). They control a range of physiological and pathological functions such as development, differentiation, apoptosis and metabolism (Ambros, 2004). We have searched the PubMed database for recently published studies of microRNAs during the first few days or weeks after TBI in humans and experimental animals, and whether they can serve as potential targets for progress assessment and intervention against TBI to mitigate secondary damage to the brain. This has implications regarding improving the diagnostic accuracy of brain impairment and long-term outcomes as well as potential novel treatments.
MicroRNAs in TBI
The initial severity and prognosis of TBI in humans is classified on the Glasgow Coma Scale (GCS) that is based on three tests: ocular, verbal, and motor responses. The sum of the three values is the GCS score. The injury is classified as severe with GCS 3–8, moderate GCS 9–12, or mild GCS 13–15 (Bilgin et al., 2012). The GCS covers a broad severity of symptoms from subtle to more serious conditions in some cases accompanied with temporary loss of consciousness (Gennarelli, 2014). Sports-related and other minor traumatic brain injuries with GCS scores in the 13–15 range occur in 1.6 to 3.8 million young healthy individuals annually (Leo and McCrea, 2016). Mild TBI is characterized by loss of consciousness for < 30 minutes and post-traumatic amnesia for < 24 hours and accounts for approximately 75–90% of the total TBI cases (Fischer, 2013). Computed tomography and magnetic resonance imaging have limited sensitivity to detect mild TBI (Kesler et al., 2000; Yuh et al., 2012). Mild TBI is often unnoticed or misdiagnosed due to the absence of specific diagnostic markers.
In experimental animal models, the extent of brain injury is classified according to the parameters used, together with behavioral testing and histological examination of brain sections. For example, in the controlled cortical impact (CCI) procedure which uses a pneumatic controlled cortical impact device to deliver mechanical energy to the intact dura, the amount of brain deformation is a function of the impactor tip size, contact velocity and duration time (Lighthall, 1988; Davis et al., 2008; Gilmer et al., 2009). By varying these parameters different severity grades of TBI injury have been produced in mice and rats. Another procedure is the weight drop injury (WDI) method in which a metal rod is dropped from a known height above the exposed skull of the animal to create a brain injury (Marmarou et al., 1994). Different severity grades of TBI injury have been produced in mice by varying the weight, diameter of tip, and fall height of the impactor rod. The weight drop method was found to recreate the cognitive and behavioral deficits seen in human patients with mild TBI (Yang et al., 2013; Sharma et al., 2014). A third method is the fluid percussion injury (FPI) method, which involves craniotomy to expose the dura and placement of a female Luer lock or plastic cap that is cemented in place over the craniotomy site; this is followed by transient compression and/or deformation of the underlying brain using a fluid-mediated pressure pulse, with a pulse duration time of approximately 20 ms (Kabadi et al., 2010; Alder et al., 2011).
Human studies
Six studies were found and mostly comprised both male and female patients with ages ranging from 14 to 65 years (Table 1). The two largest studies were performed with 76 patients (59 males/17 females, mean age 49 years; Yang et al., 2016) and 70 patients (51 males/19 females, mean age 41 years; Di Petro et al., 2017). Of the six studies, five had patients with mild TBI and severe TBI classified on the GCS and two of these also included patients with moderate TBI. The remaining study comprised only patients with severe TBI. These studies showed that serum or plasma levels of specific microRNAs could serve as potentially valuable indicators of the diagnosis, severity, and prognosis of TBI. Serum/plasma levels of miR-93, -191, -499, -23b, -21, -335, -425, -502, -16, -92a, -765 could be used to distinguish between patients with severe, moderate or mild TBI (Redell et al., 2010; Sun et al., 2016; Yang et al., 2016; Di Petro et al., 2017). Moreover, cerebro-spinal fluid (CSF) levels of miR-141, -257, -1297, -228, -362-3p, -451, -486 were identified as distinguishing severe TBI patients from controls (Bhomia et al., 2016; You et al., 2016). In addition, serum levels of miR-93, -191, -499, -425-p were indicative of outcome in TBI patients (Yang et al., 2016; Di Petro et al., 2017). MicroRNAs with upregulated and downregulated expression in patients with mild and severe TBI are shown in Figure 1.
Table 1.
MicroRNAs in human patients with traumatic brain injury (TBI)
Figure 1.
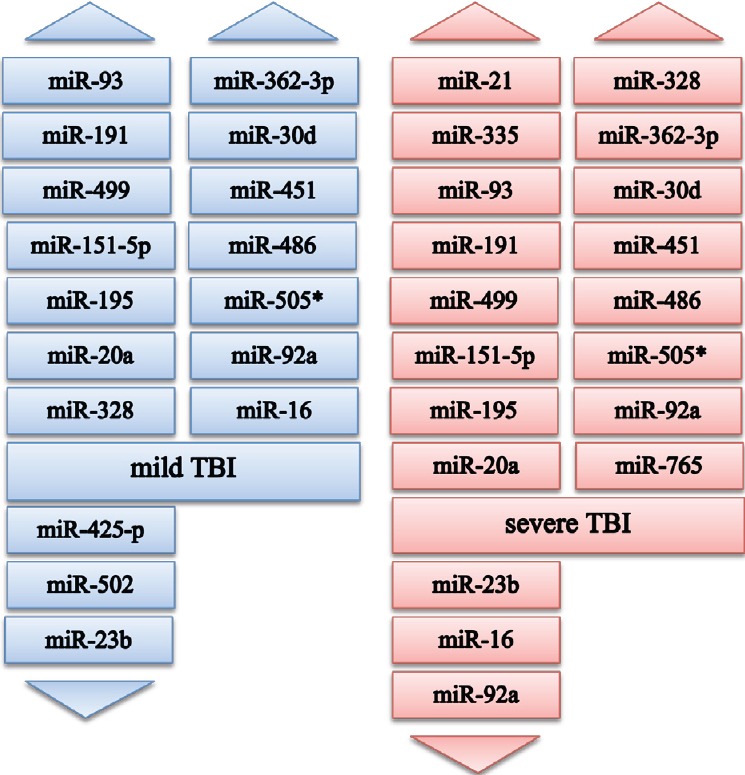
Altered expression of circulating microRNAs in human patients with mild and severe TBI (based on the studies reviewed).
For mild TBI: Increased levels of miR-93, -191, -499 at 0–24 hours, days 2–7 post-injury; miR-151-5p, -195, -20a, -328, -362-3p, -30d, -451, -486, -505*, -92a at 0–24 hours post-injury; miR-16 at 0-10 hours post-injury; decreased levels of miR-425-p, -502 at 0–1 hour, 4–12 hours post-injury, miR-23b at 0–24 hours post-injury. For severe TBI: Increased levels of miR-21 at 4–12 hours, 48–72 hours post-injury; miR-335 at 0–1 hour, 4–12 hours, 48–72 hours post-injury; miR-93, -191, -499 at 0–24 hours, days 2–7 post-injury; miR-151-5p, -195, -20a, -328, -362-3p, -30d, -451, -486, -505* at 0–48 hours post-injury; miR-92a at 0-48 hours (mean 26 hours) post-injury; miR-765 mean 68 hours post-injury; decreased levels of miR-23b at 0-24 hours post-injury; miR-16, -92a at mean 68 hours post-injury.
Animal studies
Fifteen studies were found which had mostly used male mice and rats (Table 2). Ten of these studies had used the CCI method, three the WDI method, and two the FPI method to create the brain injury. The ages of the mice ranged from 8 to 24 weeks (adult) and 22–24 months (aged), and for the rats the ages ranged from 7 to 12 weeks (adult).
Table 2.
MicroRNAs in animal models of traumatic brain injury (TBI)
CCI studies
The CCI parameters ranged from 3 to 5 mm for diameter of impactor tip, 0.8 to 6 m/s for impact velocity, 85 to 200 ms for duration time, and deformation depth of cortex 1 to 2 mm. By comparison with the parameters used by Liu et al. (2014) to induce a moderate TBI by CCI (equivalent to a GCS score of 9–12 for humans), most of the studies had used parameters to create a moderate TBI. In two of the ten studies (Sandhir et al., 2014; Meissner et al., 2016), the parameters chosen would more likely be associated with a mild-moderate brain injury. Many of the studies identified specific microRNAs as possible biomarkers and therapeutic targets. Meissner et al. (2016) reported the upregulated expression in cortex of miR-2137, -451, -21*, -144, -184, and the downregulated expression of miR-190, -137, -541, -107 at 1, 6 and 12 hours after TBI in mice compared to control. Similarly, miR-23a and miR-27a were downregulated in cortex of TBI mice as early as 1 hour and lasting up to 24 hours post-injury compared to control (Sabirzhanov et al., 2014). Liu et al. (2014) found that miR-142-3p, -144, -340-5p, -674-5p, -153, -186, -190, -132*, -138-1* were upregulated whereas let-7b was downregulated in hippocampus of TBI rats compared to controls. Also, miR-155 and miR-223 levels were significantly increased in mitochondrial fraction of hippocampus of TBI rats compared to controls (Wang et al., 2015). Interestingly, two studies examined microRNA profiles in mice given free access to a running wheel to exercise. The cortical levels of miR-21, -92a and -874 decreased while those of miR-138, -124 and let-7c increased compared with the TBI-non-runner group (Miao et al., 2015). Hu et al. (2015) observed the level of miR-21 was significantly elevated in hippocampus of TBI-non-runners compared with that of sham animals and the increase in miR-21 expression in hippocampus was reversed in TBI-runners. Also Sandhir et al. (2014) showed that basal expression of miR-21 was higher in the aged mice brain compared to adult brain. While miR-21 levels increased following TBI in adult mice with maximum increase at 24 hours post-injury, in the aged mice miR-21 expression in the brain was decreased after TBI. Strategies using microRNA mimics or agomirs to increase the levels of downregulated microRNAs or microRNA antagomirs to decrease the levels of upregulated microRNAs were also examined as possible ways to improve outcomes in TBI (e.g. Sabirzhanov et al., 2014, 2016).
WDI studies
The parameters used ranged from 246 g/333 g (mice) to 450 g (rats) for weight of rod 1 mm diameter, and 2 cm/3 cm (mice) to 150 cm (rats) for fall height. No lesions were observed in stained sections of injured mouse brains, consistent with creating a mild TBI in these animals (Sharma et al., 2014; Chandran et al., 2017). A modified neurological severity score (mNSS) (Zhang et al., 2013) of 10 in injured rats indicated a moderate TBI (Sun et al., 2016). Varying intensities of mild TBI induced a differential microRNA expression profile in the mouse brain post-injury (Sharma et al., 2014; Chandran et al., 2017). A set of 13 microRNAs was found to identify mild TBI regardless of its severity: mmu-miR-376a, hsa-214, mmu-214, mmu-337-5p, mmu-574-3p, mmu-434-3p, mmu-671-3p, mmu-218, mmu-199a-3p were all upregulated, whereas hsa-miR-106b*, mmu-106b, mmu-31, mmu-196c were all downregulated (Sharma et al., 2014). In the rat study, levels of miR-23b were decreased in the plasma, hippocampus and cortex of TBI animals post-injury compared to sham. Pretreatment of TBI animals with lentivirus-miR-23b significantly reduced the lesion volume of the contused hemisphere and brain edema compared with TBI group. Also mNSS score at 3, 7 and 14 days post-injury was decreased in TBI animals pretreated with lentivirus-miR-23b (Sun et al., 2016).
FPI studies
In one of the rat studies the pressure applied was 1.8–2.0 atm while in the other it was 1.8–2.2 atm. The duration of pulse was not reported in either study. Based on mNSS score of 8 on day 1 post-injury (Ge et al., 2014) and histological sections of cerebral cortex after injury (Lei et al., 2009) both studies had utilized animals with moderate TBI. Expression of miR-21 was increased in the impacted brain area (Lei et al., 2009; Ge et al., 2014) and was significantly increased or decreased in the agomir- or antagomir-injected animals post-injury compared to controls (Ge et al., 2014). Upregulation and downregulation of specific microRNAs in brains of TBI animals occurred at different time points post-injury (Lei et al., 2009). MiR-21 could be a potential therapeutic target for intervention after TBI.
The alteration of microRNAs in different body fluids and tissues from the described human and animal studies is summarized in Table 3.
Table 3.
Summary of microRNA alterations in traumatic brain injury (TBI) of human patients and animal models

MicroRNA Diagnostic and Therapeutic Strategies for TBI
The human studies have identified specific microRNAs in serum/plasma (miR-425-p, -21, -93, -191 and -499) and CSF (miR-328, -362-3p, -451, -486a) as possible indicators of the diagnosis, severity, and prognosis of TBI (Table 1). To date, there have been only a limited number of studies involving patients with TBI. These studies have included patients with mild, moderate, and severe TBI. However, such studies are important to increase the accuracy of diagnosis and outcomes, and refine treatments of injured patients. The early identification of biomarkers measured in samples collected within 1 hour from injury by the first responders (ambulance crew) at the injury scene before patients were admitted to hospital would allow clinicians to recognize and treat patients at risk of secondary brain injury and optimize early management. While the levels of specific microRNAs in serum or plasma may be developed into biomarkers for TBI, it is likely that other organs contribute to microRNAs in the blood (Sierzega et al., 2017) so that the circulating levels may not accurately reflect the levels of specific microRNAs in the brain itself. The microRNA profile in CSF would more reliably indicate alterations in the brain caused by TBI, since it is in direct contact with the extracellular matrix in the brain and its composition better reflects biochemical changes that occur in this organ. However, collection of CSF involves an invasive procedure and may not be easily accomplished in the elderly due to narrowing of the lumbar intervertebral space due to disc degeneration. Measurement of specific microRNAs in the serum/plasma or CSF of human patients enables mild TBI to be recognized and distinguished from severe TBI. At present, many cases of mild TBI go unnoticed or misdiagnosed, as the current diagnostic tests are neither sensitive nor specific enough to identify individuals who have sustained a mild TBI (Belanger et al., 2007; Dash et al., 2010).
The animal studies have examined specific microRNAs as biomarkers and therapeutic targets for moderate and mild TBI (e.g., miR-21, miR-23b) (Table 2). No studies were found on microRNA expression levels in animals subjected to severe TBI, presumably because of the likely high mortality rate and ethical concerns. Several of the studies had examined the effect of voluntary exercise on microRNA profiling and shown to be altered by exercise. In addition, differences in basal microRNA expression in the brain of adult and aged animals and alterations in response to TBI (e.g., miR-21) have been reported. The latter is very relevant as persons aged ≥ 75 have an increasing incidence of fall-related TBI.
Downstream targets of several important microRNAs have been indicated in the studies reviewed. For example, miR-23b specifically targeted to 3′-UTR of autophagy-related protein 12 (ATG12) mRNA in human TBI patients (Sun et al., 2016). Also upregulation of miR-711 inhibited Akt expression in the injured mouse cortex as early as 1 hour post-injury. TBI resulted in increased levels of pro-apoptotic proteins PUMA and two isoforms of Bim large and small, which are regulators of apoptosis upstream of mitochondrial release of allograft inflammatory factor 1 (AIF-1) and cytochrome c (Sabirzhanov et al., 2016). MiR-21 regulated cell survival, invasiveness and apoptosis through specific mRNA targets that included PTEN, which was upregulated by 3.8-fold in the mouse adult brain 24 hours post-injury (Sandhir et al., 2014). Post-injury decreases in miR-23a and miR-27a contributed to neuronal cell death in mouse brain after TBI by upregulating proapoptotic Bcl-2 family members (Sabirzhanov et al., 2014). In rat hippocampus, upregulated miR-221 may regulate cell proliferation and angiogenesis by PDGF signaling pathway at the early stages after TBI (Sun et al., 2014). A decrease was detected in expression of NRF2, the protein of Nfe212 gene that is a putative target of both the upregulated microRNAs, miR-144 and miR-153, in rat hippocampus post-TBI injury. SNCA, one of the target proteins of miR-153, was also significantly suppressed in the hippocampus after TBI (Liu et al. 2014). In injured rat cerebral cortex and hippocampus, upregulated miR-21 modulated the expression of apoptotic and angiogenesis-related molecules in brain after TBI. The expression of Pten, a miR-21 target gene, was inhibited and Akt signaling was activated by miR-21 after TBI (Ge et al., 2014). The mature mRNAs predicted to be targeted by the upregulated microRNAs in human patients with TBI are involved in major TBI related pathways such as erythropoietin signaling, G protein coupled receptor signaling, GABA receptor signaling and neuropathic pain signaling in dorsal horn neurons (Bhomia et al., 2016). Several brain function related pathways such as axon guidance, gap junction, dopaminergic synapse, cholinergic synapse, long-term potentiation, tight junction, adherens junction, and long-term depression were targeted by the modulated microRNAs in injured mouse brain (Sharma et al., 2014).
Wang et al. (2015) examined miR-155, -223, -146a, -142-3p, -142-5p, -150, -124a expression in enriched cortical neuron, astrocyte and microglial cultures of injured rats. MiR-155, -223, -146a, -142-3p, -142-5p, -150 were detected in all three cell types but showed the highest expression in microglia and astrocytes, whereas levels of miR-124a were highest in cortical neurons. While not included in this review, other studies have examined the actions of specific microRNAs in cultures of cortical neurons (Han et al., 2014; Ma et al., 2016).
Future Perspectives
At the present time, there is no effective treatment for TBI due, in part, to the widespread impact of numerous complex secondary biochemical and pathophysiological events occurring at different time points following the initial injury (Rosenfeld et al., 2012). These secondary injury events include edema, glutamate excitotoxicity, inflammation, oxidative stress damage, activation of apoptosis, necrosis and autophagy, and impaired mitochondrial function (Raghupathi, 2004; Singh et al., 2006; Ziebell and Morganti-Kossmann, 2010). The earliest and most severe neuropathological changes occur in the hippocampus after TBI (Orrison et al., 2009; Fanselow and Dong, 2010). The hippocampus, a key brain structure for learning and memory, is particularly vulnerable to TBI. The molecular mechanisms underlying hippocampal alterations and cognitive impairments following TBI are unclear and further studies are warranted.
A single microRNA can regulate the expression of hundreds of target genes, so alterations in a panel of microRNAs could greatly affect the pathophysiology and outcome of TBI. Identification of specific genes and signal transduction pathways directly involved in TBI is essential for development of novel therapeutic strategies.
Evidences have also shown that several aspects of the response to brain injury are altered in aged animals. These include worsening of behavioral outcomes, increase of inflammatory responses to injury, and decrease of neuroprotective responses (Sandhir et al., 2004, 2008; Anderson et al., 2009; Sandhir and Berman, 2010). In the aged brain following injury there is increased blood-brain barrier (BBB) permeability accompanied by increased matrix metalloproteinase-9 activity and decreased BBB repair processes (Lee et al., 2012). These changes are all likely to result from altered gene and protein expressions after injury. Voluntary exercise by individuals with TBI (e.g., treadmill, exercycle) could aid recovery of TBI patients.
Plasma microRNAs have the potential to improve TBI patient classification and possibly clinical management. Combining plasma levels of miR-16, -92a and -765, 100% sensitivity and 100% accuracy could be achieved in identifying severe TBI patients (Redell et al., 2010). It is possible that a panel of specific circulating microRNAs could achieve 100% sensitivity and 100% accuracy in identifying mild TBI. Bhomia et al. (2016) identified a panel of 10 unique microRNAs for the diagnosis of mild/moderate TBI and severe TBI. Temporal expression of specific microRNAs may be used as a molecular marker for assessment of TBI progression and inference of damage time. At the present time there are concerns about the specificity of human biofluid biomarkers such as S100β, glial fibrillary acid protein, neuron-specific enolase, and tau protein for TBI. Many investigators have failed to detect associations between serum levels of S100β and CT abnormalities. The utiity of S100β in the setting of multiple trauma remains controversial because it is also elevated in trauma patients without head injuries (Papa et al., 2015). A study by Papa et al. (2012) indicated that serum glial fibrillary acid protein had good specificity for brain injury acutely after injury, although it is also detected in patients with non-TBI injuries. Studies with the serum biomarker neuron-specific enolase concluded that it had limited utiliiy as a marker of neuronal damage (Fridriksson et al., 2000). A proteolytically cleaved form of tau protein, c-tau, was significantly elevated in CSF in TBI patients and the levels correlated with clinical outcome (Shaw et al., 2002). Although levels of c-tau were also elevated in plasma from patients with severe TBI, there was no correlation between plasma levels and clinical outcome (Chatfield et al., 2002). There clearly is a need for identifying and innovating sensitive and reliable new TBI biomarkers.
A strong association has been shown between TBI and later development of ischemic stroke that remained significant even after several potential confounders, such as vascular risk factors and comorbidities, were taken into consideration. Specific biomarkers for ischemic stroke have been reported (Martinez and Peplow, 2016). Further large-scale studies with TBI patients are needed to provide more information on the changes in microRNA profiles in different age groups (children, adults, and elderly). Future studies should assess the values of the altered microRNAs in TBI to other measures of injury severity according to assessments other than the GCS such as duration of coma and the duration of post-traumatic amnesia. Also as hypertension is a common comorbidity in persons with TBI (Holcomb et al., 2012), and a risk factor for ischemic stroke, studies with hypertensive animals subjected to a brain contusion injury should be performed.
Footnotes
Conflicts of interest: None declared.
Plagiarism check: Checked twice by iThenticate.
Peer review: Externally peer reviewed.
Open peer reviewer: Dzung H Dinh, Illinois Neurological Institute, USA.
References
- 1.Adekoya N, Thurman DJ, White DD, Webb KW. Surveillance for traumatic brain injury deaths--United States, 1989-1998. MMWR Surveill Summ. 2002;51:1–14. [PubMed] [Google Scholar]
- 2.Alder J, Fujioka W, Lifshitz J, Crockett DP, Thakker-Varia S. Lateral fluid percussion: model of traumatic brain injury in mice. J Vis Exp. 2011;54:3063. doi: 10.3791/3063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ambros V. The functions of animal microRNAs. Nature. 2004;431:350–355. doi: 10.1038/nature02871. [DOI] [PubMed] [Google Scholar]
- 4.Anderson J, Sandhir R, Hamilton ES, Berman NE. Impaired expression of neuroprotective molecules in the HIF-1 pathway following traumatic brain injury in aged mice. J Neurotrauma. 2009;26:1557–1566. doi: 10.1089/neu.2008.0765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Baek D, Villén J, Shin C, Camargo FD, Gygi SP, Bartel DP. The impact of microRNAs on protein output. Nature. 2008;455:64–71. doi: 10.1038/nature07242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Belanger H, Vanderploeg R, Curtiss G, Warden DL. Recent neuroimaging techniques in mild traumatic brain injury. J Neuropsychiatry Clin Neurosci. 2007;19:5–20. doi: 10.1176/jnp.2007.19.1.5. [DOI] [PubMed] [Google Scholar]
- 7.Bhomia M, Balakarthiresan NS, Wang KK, Papas L, Maheshwari BR. A panel of serum miRNA biomarkers for the diagnosis of severe to mild traumatic brain injury in humans. Sci Rep. 2016;6:28148. doi: 10.1038/srep28148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bilgin S, Guclu-Gunduz A, Oruckaptan H, Kose N, Celik B. Gait and Glasgow Coma Scale scores can predict functional recovery in patients with traumatic brain injury. Neural Regen Res. 2012;7:1978–1984. doi: 10.3969/j.issn.1673-5374.2012.25.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Chandran R, Sharma A, Bhomia M, Balakathiresan NS, Knollmann-Ritschel BE, Maheshwari RK. Differential expression of microRNAs in the brains of mice subjected to increasing grade of mild traumatic brain injury. Brain Inj. 2017;31:106–119. doi: 10.1080/02699052.2016.1213420. [DOI] [PubMed] [Google Scholar]
- 10.Chatfield DA, Zemlan FP, Day DJ, Menon DK. Discordant temporal patterns of S100beta and cleaved tau protein elevation after head injury: a pilot study. Br J Neurosurg. 2002;16:471–476. doi: 10.1080/0268869021000030285. [DOI] [PubMed] [Google Scholar]
- 11.Dash PK, Zhao J, Hergenroeder G, Moore AN. Biomarkers for the diagnosis, prognosis, and evaluation of treatment efficacy for traumatic brain injury. Neurotherapeutics. 2010;7:100–114. doi: 10.1016/j.nurt.2009.10.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Davis LM, Pauly JR, Readnower RD, Rho JM, Sullivan PG. Fasting is neuroprotective following traumatic brain injury. J Neurosci Res. 2008;86:1812–1822. doi: 10.1002/jnr.21628. [DOI] [PubMed] [Google Scholar]
- 13.Di Pietro V, Ragusa M, Davies D, Su Z, Hazeldine J, Lazzarino G, Hill LJ, Crombie N, Foster M, Purrello M, Logan A, Belli A. MicroRNAs as novel biomarkers for the diagnosis and prognosis of mild and severe traumatic brain injury. J Neurotrauma. 2017;34:1948–1956. doi: 10.1089/neu.2016.4857. [DOI] [PubMed] [Google Scholar]
- 14.Fanselow MS, Dong HW. Are the dorsal and ventral hippocampus functionally distinct structures? Neuron. 2010;65:7–19. doi: 10.1016/j.neuron.2009.11.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Fischer H. U.S. Military Casualty Statistics: Operation New Dawn, Operation Iraqi Freedom, and Operation Enduring Freedom. CRS Report for Congress Prepared for Members and Committees of Congress: Congressional Research Service. 2013:1–12. [Google Scholar]
- 16.Fridriksson T, Kini N, Walsh-Kelly C, Hennes H. Serum neuron-specific enolase as a predictor of intracranial lesions in children with head trauma: A pilot study. Acad Emerg Med. 2000;7:816–820. doi: 10.1111/j.1553-2712.2000.tb02276.x. [DOI] [PubMed] [Google Scholar]
- 17.Ge XT, Lei P, Wang HC, Zhang AL, Han ZL, Chen X, Li SH, Jiang RC, Kang CS, Zhang JN. miR-21 improves the neurological outcome after traumatic brain injury in rats. Sci Rep. 2014;4:6718. doi: 10.1038/srep06718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gennarelli TA. Future directions in brain injury research. Prog Neurol Surg. 2014;28:243–250. doi: 10.1159/000358788. [DOI] [PubMed] [Google Scholar]
- 19.Gilmer LK, Roberts KN, Joy K, Sullivan PG, Scheff SW. Early mitochondrial dysfunction after cortical contusion injury. J Neurotrauma. 2009;26:1271–1280. doi: 10.1089/neu.2008.0857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Han Z, Chen F, Ge X, Tan J, Lei P, Zhang J. miR-21 alleviated apoptosis of cortical neurons through promoting PTEN-Akt signaling pathway in vitro after experimental traumatic brain injury. Brain Res. 2014;1582:12–20. doi: 10.1016/j.brainres.2014.07.045. [DOI] [PubMed] [Google Scholar]
- 21.Holcomb EM, Millis SR, Hanks RA. Comorbid diseases in persons with traumatic brain injury: descriptive findings using the modified cumulative illness rating scale. Arch Phys Med Rehabil. 2012;93:1338–1342. doi: 10.1016/j.apmr.2012.04.029. [DOI] [PubMed] [Google Scholar]
- 22.Hu T, Zhou FJ, Chang YF, Li YS, Liu GC, Hong Y, Chen HL, Xiyang YB, Bao TH. miR21 is associated with the cognitive improvement following voluntary running wheel exercise in TBI mice. J Mol Neurosci. 2015;57:114–122. doi: 10.1007/s12031-015-0584-8. [DOI] [PubMed] [Google Scholar]
- 23.Hu Z, Yu D, Almeida-Suhett C, Tu K, Marini AM, Eiden L, Braga MF, Zhu J, Li Z. Expression of miRNAs and their cooperative regulation of the pathophysiology in traumatic brain injury. PLoS One. 2012;7:e39357. doi: 10.1371/journal.pone.0039357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Jennett B. Epidemiology of head injury. Arch Dis Child. 1998;78:403–406. doi: 10.1136/adc.78.5.403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kabadi SV, Hilton GD, Stoica BA, Zapple DN, Faden AI. Fluid-percussion-induced traumatic brain injury model in rats. Nat Protoc. 2010;5:1552–1563. doi: 10.1038/nprot.2010.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kesler SR, Adams HF, Bigler ED. SPECT, MR and quantitative MR imaging: correlates with neuropsychological and psychological outcome in traumatic brain injury. Brain Inj. 2000;14:851–857. doi: 10.1080/026990500445682. [DOI] [PubMed] [Google Scholar]
- 27.Lee P, Kim J, Williams R, Sandhir R, Gregory E, Brooks WM, Berman NE. Effects of aging on blood brain barrier and matrix metalloproteases following controlled cortical impact in mice. Exp Neurol. 2012;234:50–61. doi: 10.1016/j.expneurol.2011.12.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lei P, Li Y, Chen X, Yang S, Zhang J. Microarray based analysis of microRNA expression in rat cerebral cortex after traumatic brain injury. Brain Res. 2009;12841:191–201. doi: 10.1016/j.brainres.2009.05.074. [DOI] [PubMed] [Google Scholar]
- 29.Leo P, McCrea M. Epidemiology. In: Laskowitz D, Grant G, editors. Translational Research in Traumatic Brain Injury. Boca Raton: CRC Press/Taylor and Francis Group; 2016. [PubMed] [Google Scholar]
- 30.Lighthall JW. Controlled cortical impact: a new experimental brain injury model. J Neurotrauma. 1988;5:1–15. doi: 10.1089/neu.1988.5.1. [DOI] [PubMed] [Google Scholar]
- 31.Liu L, Sun T, Liu Z, Chen X, Zhao L, Qu G, Li Q. Traumatic brain injury dysregulates microRNAs to modulate cell signaling in rat hippocampus. PLoS One. 2014;9:e103948. doi: 10.1371/journal.pone.0103948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ma J, Shui S, Han X, Guo D, Li T, Yan L. microRNA-22 attenuates neuronal cell apoptosis in a cell model of traumatic brain injury. Am J Transl Res. 2016;8:1895–1902. [PMC free article] [PubMed] [Google Scholar]
- 33.Maas AI, Steyerberg EW, Murray GD, Bullock R, Baethmann A, Marshall LF, Teasdale GM. Why have recent trials of neuroprotective agents in head injury failed to show convincing efficacy? A pragmatic analysis and theoretical considerations. Neurosurgery. 1999;44:1286–1298. [PubMed] [Google Scholar]
- 34.Maas AI, Stocchetti N, Bullock R. Moderate and severe traumatic brain injury in adults. Lancet Neurol. 2008;7:728–741. doi: 10.1016/S1474-4422(08)70164-9. [DOI] [PubMed] [Google Scholar]
- 35.Marmarou A, Foda MA, van den Brink W, Campbell J, Kita H, Demetriadou K. A new model of diffuse brain injury in rats. Part I: Pathophysiology and biomechanics. J Neurosurg. 1994;80:2910–2300. doi: 10.3171/jns.1994.80.2.0291. [DOI] [PubMed] [Google Scholar]
- 36.Martinez B, Peplow PV. Blood microRNAs as potential diagnostic and prognostic markers in cerebral ischemic injury. Neural Regen Res. 2016;11:1375–1378. doi: 10.4103/1673-5374.191196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Meissner L, Gallozzi M, Balbi M, Schwarzmaier S, Tiedt S, Terpolilli NA, Plesnila N. Temporal profile of microRNA expression in contused cortex after traumatic brain injury in mice. J Neurotrauma. 2016;33:713–720. doi: 10.1089/neu.2015.4077. [DOI] [PubMed] [Google Scholar]
- 38.Miao W, Bao TH, Han JH, Yin M, Yan Y, Wang WW, Zhu YH. Voluntary exercise prior to traumatic brain injury alters miRNA expression in the injured mouse cerebral cortex. Braz J Med Biol Res. 2015;48:433–439. doi: 10.1590/1414-431X20144012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Orrison WW, Hanson EH, Alamo T, Watson D, Sharma M, Perkins TG, Tandy RD. Traumatic brain injury: a review and high-field MRI findings in 100 unarmed combatants using a literature-based checklist. J Neurotrauma. 2009;26:689–701. doi: 10.1089/neu.2008.0636. [DOI] [PubMed] [Google Scholar]
- 40.Papa L, Edwards D, Ramia M. Exploring serum biomarkers for mild traumatic brain injury. In: Kobeissy FH, editor. Brain Neurotrauma. Molecular, Neuropsychological, and Rehabilitation Aspects. Boca Raton: CRC Press/Taylor & Francis; 2015. [PubMed] [Google Scholar]
- 41.Papa L, Lewis LM, Falk JL, Zhang Z, Silvestri S, Giordano P, Demery JA, Dixit NK, Ferguson I, Liu MC, Mo J, Akinyi L, Schmid K, Mondello S, Robertson CS, Tortella FC, Hayes RL, Wang KK. Elevated levels of serum glial fibrillary acidic protein breakdown products in mild and moderate traumatic brain injury are associated with intracranial lesions and neurosurgical intervention. Ann Emerg Med. 2012;59:471–483. doi: 10.1016/j.annemergmed.2011.08.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Raghupathi R. Cell death mechanisms following traumatic brain injury. Brain Pathol. 2004;14:215–222. doi: 10.1111/j.1750-3639.2004.tb00056.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Redell JB, Moore AN, Ward NH, Hergenroader GW, Dash RK. Human traumatic brain injury alters plasma microRNA levels. J Neurotrauma. 2010;27:2147–2156. doi: 10.1089/neu.2010.1481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Ribbers GM. Stone JH, Blouin M, editors. Brain Injury: Long term outcome after traumatic brain injury. International Encyclopedia of Rehabilitation. 2010. [accessed14 July 2017]. http://cirrie.buffalo.edu/encyclopedia/en/article/338 .
- 45.Rosenfeld JV, Maas AI, Bragge P, Morganti-Kossmann MC, Manley GT, Gruen RL. Early management of severe traumatic brain injury. Lancet. 2012;380:1088–1098. doi: 10.1016/S0140-6736(12)60864-2. [DOI] [PubMed] [Google Scholar]
- 46.Sabirzhanov B, Zhao Z, Stoica BA, Loane DJ, Wu J, Borroto C, Dorsey SG, Faden AI. Downregulation of miR-23a and miR-27a following experimental traumatic brain injury induces neuronal cell death through activation of proapoptotic Bcl-2 proteins. J Neurosci. 2014;34:10055–10071. doi: 10.1523/JNEUROSCI.1260-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Sabirzhanov B, Stoica BA, Zhao Z, Loane DJ, Wu J, Dorsey SG, Faden AI. miR-711 upregulation induces neuronal cell death after traumatic brain injury. Cell Death Differ. 2016;23:654–658. doi: 10.1038/cdd.2015.132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Sandhir R, Puri V, Klein RM, Berman NE. Differential expression of cytokines and chemokines during secondary neuron death following brain injury in old and young mice. Neurosci Lett. 2004;369:28–32. doi: 10.1016/j.neulet.2004.07.032. [DOI] [PubMed] [Google Scholar]
- 49.Sandhir R, Onyszchuk G, Berman NE. Exacerbated glial response in the aged mouse hippocampus following controlled cortical impact injury. Exp Neurol. 2008;213:372–380. doi: 10.1016/j.expneurol.2008.06.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Sandhir R, Berman NE. Age-dependent response of CCAT/enhancer binding protein following traumatic brain injury in mice. Neurochem Int. 2010;56:188–193. doi: 10.1016/j.neuint.2009.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Sandhir R, Gregory E, Berman NE. Differential response of miRNA-21 and its targets after traumatic brain injury in aging mice. Neurochem Int. 2014;78:117–121. doi: 10.1016/j.neuint.2014.09.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Sharma A, Chandran R, Barry ES, Bhomia M, Hutchison MA, Balakathiresan NS, Grunberg NE, Maheshwari RK. Identification of serum microRNA signatures for diagnosis of mild traumatic brain injury in a closed head injury model. PLoS One. 2014;9:e112019. doi: 10.1371/journal.pone.0112019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Shaw GJ, Jauch EC, Zemlan FP. Serum cleaved tau protein levels and clinical outcome in adult patients with closed head injury. Ann Emerg Med. 2002;39:254–257. doi: 10.1067/mem.2002.121214. [DOI] [PubMed] [Google Scholar]
- 54.Sierzega M, Kaczor M, Kolodziejczyk P, Kulig J, Sanak M, Richter P. Evaluation of serum microRNA biomarkers for gastric cancer based on blood and tissue pools profiling: the importance of miR-21 and miR-331. Br J Cancer. 2017;117:266–273. doi: 10.1038/bjc.2017.190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Singh IN, Sullivan PG, Deng Y, Mbye LH, Hall ED. Time course of post-traumatic mitochondrial oxidative damage and dysfunction in a mouse model of focal traumatic brain injury: implications for neuroprotective therapy. J Cereb Blood Flow Metab. 2006;26:1407–1418. doi: 10.1038/sj.jcbfm.9600297. [DOI] [PubMed] [Google Scholar]
- 56.Sun L, Liu A, Zhang J, Ji W, Li Y, Yang X, Wu Z, Guo J. miR-23b improves cognitive impairments in traumatic brain injury by targeting ATG12-mediated neuronal autophagy. Behav Brain Res. 2016 doi: 10.1016/j.bbr.2016.09.020. doi: 10.1016/j.bbr.2016.09.020. [DOI] [PubMed] [Google Scholar]
- 57.Sun TY, Chen XR, Liu ZL, Zhao LL, Jiang YX, Qu GQ, Wang RS, Huang SZ, Liu L. Expression profiling of microRNAs in hippocampus of rats following traumatic brain injury. J Huarzhong Univ Sci Technolog Med Sci. 2014;34:548–553. doi: 10.1007/s11596-014-1313-1. [DOI] [PubMed] [Google Scholar]
- 58.Wang WX, Visavadiya NP, Pandya JD, Nelson PT, Sullivan PG, Springer JE. Mitochondria-associated microRNAs in rat hippocampus following traumatic brain injury. Exp Neurol. 2015;265:84–93. doi: 10.1016/j.expneurol.2014.12.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Yang SH, Gustafson J, Gangidine M, Stepien D, Schuster R, Pritts TA, Goodman MD, Remick DG, Lentsch AB. A murine model of mild traumatic brain injury exhibiting cognitive and motor deficits. J Surg Res. 2013;184:981–988. doi: 10.1016/j.jss.2013.03.075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Yang T, Song J, Bu X, Wang C, Wu J, Cai J, Wan S, Fan C, Zhang C, Wang J. Elevated serum miR-93, miR-191, and miR-499 are noninvasive biomarkers for the presence and progression of traumatic brain injury. J Neurochem. 2016;137:122–129. doi: 10.1111/jnc.13534. [DOI] [PubMed] [Google Scholar]
- 61.You WD, Tang QL, Wang L, Lei J, Feng JF, Mao Q, Gao GY, Jiang JY. Alteration of microRNA expression in cerebrospinal fluid of unconscious patients after traumatic brain injury and a bioinformatic analysis of related single nucleotide polymorphisms. Chin J Traumatol. 2016;19:11–15. doi: 10.1016/j.cjtee.2016.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Yuh EL, Cooper SR, Ferguson AR, Manley GT. Quantitative CT improves outcome prediction in acute traumatic brain injury. J Neurotrauma. 2012;29:735–746. doi: 10.1089/neu.2011.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Zhang R, Liu Y, Yan K, Chen L, Chen XR, Li P, Chen FF, Jiang XD. Anti-inflammatory and immunomodulatory mechanisms of mesenchymal stem cell transplantation in experimental traumatic brain injury. J Neuroinflammation. 2013;10:106. doi: 10.1186/1742-2094-10-106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Ziebell JM, Morganti-Kossmann MC. Involvement of pro- and anti-inflammatory cytokines and chemokines in the pathology of traumatic brain injury. Neurotherapeutics. 2010;7:22–30. doi: 10.1016/j.nurt.2009.10.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Zweckberger K, Erös C, Zimmermann R, Kim SW, Engel D, Plesnila N. Effect of early and delayed decompressive craniectomy on secondary brain damage after controlled cortical impact in mice. J Neurotrauma. 2006;23:1083–1093. doi: 10.1089/neu.2006.23.1083. [DOI] [PubMed] [Google Scholar]