

Abstract
Oxidative stress is known as the leading factor responsible for varicocele-related infertility and for that reason, many antioxidant therapies have been proposed. Considering that, we evaluated the reproductive outcomes and fertility of varicocelized rats and the impact of chrysin within these parameters. The animals were allocated into three groups: sham (control), varicocele treated via gavage with 50 mg/kg/day of chrysin (V1), or vehicle (V2) for 56 days. Chrysin treatment prevented oxidative damage resulting from varicocele by decreasing testicular concentrations of malondialdehyde and sperm DNA fragmentation. It also improved histological aspect of the testis and maintained morphometric parameters similar to the sham group. Furthermore, there were no differences in body and reproductive organ weights, histopathological analysis of epididymis, sperm counts and morphology, testosterone levels, sexual behavior, and fertility parameters among experimental groups. Our results reinforce the idea that injuries provoked by experimental varicocele are related, at least in part, to oxidative stress. Moreover, varicocele showed bilateral deleterious effects without interfering with fertility. Chrysin administration significantly ameliorated sperm parameters, protecting the reproductive system against varicocele damages. For that reason, chrysin might be an alternative adjuvant therapy to improve sperm quality in men presenting this condition.
1. Introduction
Infertility is one of the main public health problems, and in more than 40% of these cases, male disorders are involved. In this context, varicocele is considered the leading cause of infertility once it is present in approximately 15% of the male population, 35% of men with primary infertility and 70–80% with secondary infertility [1, 2]. In addition, varicocele is a bilateral [3], progressive [4], and hereditary condition [5] characterized by abnormal dilatation and tortuosity of the pampiniform plexus.
Even though varicocele is known and studied for many years, the physiopathological mechanisms underlying the varicocele have not yet been fully elucidated. However, some theories that include venous stasis leading to hypoxia and testicular hyperthermia, reflux of toxic adrenal and renal metabolites, and hormonal dysfunction and increased oxidative stress (OS) have been proposed [6].
Since men with varicocele present high levels of ROS, nitric oxide (NO), and lipid peroxidation products like malondialdehyde (MDA), as well as decrease in antioxidant defenses, it is believed that OS is a key element in the varicocele-related infertility [7].
Usually, men are advised to submit to a surgical procedure known as varicocelectomy, which is considered efficient once it improves seminal and hormonal parameters [8–11]. However, the effects on fertility potential are controversial due to lack of pre- and postoperative data, adequate methodology, and criteria applied in the studies [12]. For that reason and because they offer lower risks than the surgery, several works were exploring the benefits of antioxidant adjuvant therapy [13–15]. The protective activity against oxidative stress damages and the improvement of sperm parameters make these compounds a considerable solution.
Chrysin, a flavonoid found in honey, propolis, and plant extracts, is a potent antioxidant that has been widely studied due to its mechanism of action that prevents conversion of testosterone to estrogen through inhibition of enzyme CYP19 [16, 17]. This compound is commonly used by bodybuilding athletes for being a testosterone booster, but it is also known for its antineoplastic [18], anti-inflammatory [19, 20], antihypertensive [21], anti-aging [22] activities. Several studies using animal models reported that chrysin improved sperm parameters [23, 24], increased testosterone levels [25], and prevented the oxidative damage caused by exposure to toxic chemicals [26, 27]. Thus, because this flavonoid has a positive impact on sperm quality and a protective activity against the damage caused by OS, the aim of our work was to evaluate reproductive and fertility outcomes of varicocele and the impact of chrysin in these parameters after 56 days of treatment.
2. Material and Methods
2.1. Animals
Wistar male (78–80 days old, weighing 300–400 g) and female rats (85 days old, weighing 200–315 g) were supplied by the Central Biotherium of São Paulo State University (UNESP), Botucatu, and maintained under controlled environment with 23°C, 12/12 h light-dark cycle with free access to food and tap water. The experimental procedures were approved by the local Ethics Committee for the Use of Experimental Animals of the University of São Paulo State (protocol number 772-CEUA) and are in accordance with the Guide for the Care and Use of Laboratory Animals (National Institutes of Health).
2.2. Experimental Groups and Treatment
In all experiments, male rats were allocated into three experimental groups: sham-operate (sham; n = 5–10) and varicocele groups received daily oral gavage dose of 50 mg/kg/day of chrysin (V1; n = 7-8) diluted in vehicle (corn oil and DMSO) or only vehicle solution (V2; n = 7–10) for 56 days. The treatment period was chosen based on the duration of spermatogenesis in rats, which is approximately 56 days, since it requires 4.5 cycles within 13 days each, of the seminiferous epithelium in order for the spermatogonia to become spermatozoa [28]. Chrysin (5,7-dihydroxyflavone) and dimethyl sulphoxide (DMSO) were purchased from Sigma-Aldrich, St. Louis MO, USA, and the dosage used was based in previous reports that 50 mg/kg/day of chrysin had a positive impact in sperm quality [23, 24] and a protective role against oxidative stress damages [19, 29, 30].
2.3. Surgical Procedure
Experimental left varicocele was induced on postnatal day (PND) 78–80 by partial occlusion of the left renal vein using the method described by Turner [31] with modifications. The animals were anesthetized with an intramuscular injection of ketamine 100 mg/kg and xylazine 5 mg/kg. The upper left abdominal quadrant was accessed through a midline laparotomy incision. The abdominal content was displaced to the right antimer, and the insertion of the left internal spermatic vein into the left renal vein was identified. The fat and connective tissue surrounding the renal vein was dissected, creating a tunnel underneath the vein and in a medial position to the insertion of the left spermatic vein. A 4-0 silk suture was placed around the renal vein and tied over a 0.80 mm needle. The needle was then removed, allowing the vein to expand to the limit of the ligation. Further, all abnormal collateral vessels were fully ligated once they would provide an effluent rout and release the pressure in the left internal spermatic vein. This procedure resulted in an increase in the intravenous pressure, which was transmitted to the left spermatic vein to promote varicocele development. Then, the abdominal content was replaced and the midline incision was closed in two layers with 4-0 silk suture. The animals from the sham group underwent the same procedure, but the partial ligation of the renal vein was not performed. After one week of recovery, the animals either were treated daily with chrysin/vehicle (V1/V2) or did not received any treatment (sham).
2.3.1. Experiment 1. Assessment of Body and Organ Weights and Histological and Morphometric Analysis
In this experiment, Twenty-eight adult male rats (sham n = 10; V1 n = 8; V2 n = 10) were euthanized by decapitation at the 57th day of treatment in order to assess body and reproductive organ weights, to evaluate the histological aspect of testis and epididymis, and to perform the morphometric analysis of the testis.
2.4. Body and Reproductive Organ Weights
The animals were weighed daily and at the end of the treatment the testes, epididymides, prostate, and seminal vesicle were removed and had their weights recorded.
2.5. Histological Analysis
Testes and epididymides after weighted were fixed in Bouin's solution (25% formaldehyde, 75% saturated solution of picric acid, and 5% glacial acetic acid) for 24 h. The organs were immersed in paraffin wax and sectioned in 5 μm. These sections (cross-section for testis and longitudinal sections for epididymis) were stained with hematoxylin and eosin (HE) and examined under light microscopy. Testicular evaluation was conducted by analyzing a hundred seminiferous tubules per animal, randomly chosen, and classified as normal (organized germ cell layers in the seminiferous epithelium) or abnormal (presence of acidophilic and multinucleated cells, retained spermatids, degeneration of a specific cell type, and vacuolization of the epithelium or exfoliation of cells in the lumen). The testicular interstitium was also evaluated but in a qualitative manner. Regarding the epididymis, the histological examination was qualitative. The entire longitudinal section was evaluated for the structure and aspect of epithelium, lumen, and interstitium.
2.6. Morphometric Analysis of Seminiferous Tubules
The diameter of the seminiferous tubules and height of germinal epithelium (stage IX of spermatogenesis) were evaluated in 10 sections of seminiferous tubules per animal.
2.6.1. Experiment 2. Testosterone Levels, Sperm Parameters, and Comet Assay
Following the same experimental protocol as the previous step, twenty-four adult male rats (n = 8 per group) were euthanized and had their blood collected for hormonal measurements, the testis was excised for sperm count and epididymal portions were used for evaluation of sperm motility, morphology, count, and comet assay.
2.7. Testosterone Levels
Blood collection was performed between 8:00 and 12:00 AM. Serum was obtained by centrifugation of the samples (2500 rpm, for 20 min at 4°C) and subsequently was stored at −20°C. The concentration of testosterone (serum and intratesticular) was accessed using ImmuChem™ Double Antibody Testosterone 125I RIA Kit, and intra-assay variability for this hormone was 4.1%.
2.8. Sperm Motility
The epididymis was placed in a small petri dish containing 1 mL of modified human tubal fluid (HTF) medium (Irvine Scientific) prewarmed to 34°C. The proximal cauda was perforated and sperm suspension was transferred to a Makler chamber maintained at 34°C. Through a phase-contrast microscope at 400x magnification, one hundred sperm were classified as mobile and immobile.
2.9. Sperm Morphology
For sperm morphology analysis, 100 μL of sperm solution (the same used for motility assay) was aliquoted and added to 900 μL of 10% formol saline. Then, smears were prepared on histological slides, which were left to air dry for 40 minutes and observed under a phase-contrast microscope at 400x magnification. A total of 200 spermatozoa per animal were evaluated and abnormalities were classified in two main categories: head abnormalities (without characteristic curvature, pin head, and isolated head) and tail abnormalities (rolled, broken, and bended) [32, 33].
2.10. Sperm DNA Fragmentation (Comet Assay)
Sperm DNA fragmentation was evaluated by the comet assay as previously described [34] with the following modifications: frozen remaining sperm solution from the epididymis cauda was thawed and diluted in low melting point agarose (0.5% at 37°C; LGC Laboratories, Sao Paulo, Brazil), at a concentration of 106 cells/mL. An aliquot (75 μL) of the solution was placed on precoated slides with 1% normal melting point agarose (LGC Laboratories, Middlesex, UK). The slide was first immersed in lysis buffer (100 mM Na2-EDTA, 10 mM Tris-HCl, 2,5 M NaCl, and pH 11.0) containing 40 mM DTT and 2% Triton X-100 for 1 h at 4°C and subsequently in lysis buffer containing proteinase K (0.1 mg/mL), for 2.5 hrs at 37°C. The DNA was fractionated by alkaline electrophoresis (300 mM NaOH, 1 mM Na2 EDTA, and pH 13.0) at 3 V/cm and 270 mAmps for 45 mins. Finally, the slides were washed with water, fixed in absolute ethanol, and air-dried and stored in the dark until analysis. For microscopy, the slides were stained with SYBR® Gold (1 : 10,000; Invitrogen, Waltham, MA) and analyzed using the Comet Assay IV software (Comet Assay IV, Perceptive Instruments, Wiltshire, UK). A total of 50 cells were analyzed per slide (2 slides/animal) and DNA damage was measured by analyzing the tail intensity (% of migrated DNA).
2.11. Sperm Count
After collecting the testis from both sides, these organs were decapsulated and then frozen at −20°C until the homogenization previously described by [35], with adaptations [36, 37]. The remaining parenchyma was thawed over ice and homogenized in 5 mL of NaCl 0.9% containing Triton X-100 0.5% followed by sonication for 30 seconds. After a 10-fold dilution, an aliquot was transferred to Newbauer's chamber (4 fields per animal) and mature spermatids (stage 19 of the spermatogenic cycle) were counted. Daily sperm production (DSP) was calculated dividing the number of mature spermatids by 6.1, which is the number of days that these spermatids are present in the seminiferous epithelium. Regarding the epididymis, caput/corpus and cauda were cut in small fragments with scissors and homogenized and the counting was performed as described for the testis.
2.11.1. Experiment 3. Sexual Behavior, Reproductive Performance, and Lipid Peroxidation
On the 57th day of treatment, twenty-six male rats were paired with adult females in order to evaluate sexual behavior and reproductive performance. After analyzing these parameters, the animals were euthanized by decapitation and had both testes and epididymides weighted and stored at −80°C for posterior lipid peroxidation assay.
2.12. Sexual Behavior and Reproductive Performance
Sexual behavior was performed on the 57th day of treatment for 4 hours in the dark phase of the cycle under red illumination. For this, 26 male rats (sham, n = 9; V1, n = 7; V2, n = 10) were placed in boxes of polycarbonate crystal, for 5 minutes before the introduction of one sexually receptive adult female. The behavior assay started at the time that female was placed, and the following measures were recorded for the next 40 minutes: latency of the first mount, number of mounts, latency of the first intromission, number of intromissions preceding the first ejaculation, latency of first ejaculation, and number of ejaculations [38, 39]. In case the male did not mount or intromit in the first 10 minutes after the introduction of the adult female, it was considered sexually inactive. Following the evaluation of sexual behavior, the rats (female and male) stayed in the same box for additional 4 hours, permitting a greater number of ejaculations. After this period, vaginal smears were collected for sperm detection and establishment of the day 0 of gestation (GD0). On GD21, females were weighed and killed by decapitation to enable fertility evaluation. During the laparotomy, the uterus and ovaries were collected and the number of corpora lutea, implantation sites, reabsorptions, live fetuses, and fetal weights was determined. From these results, the following endpoints were determined: pregnancy rate = number of pregnant female/number of inseminated female × 100; rate of preimplantation loss = (number of corpora lutea − number of implantations/number of corpora lutea) × 100; and rate of postimplantation loss = (number of implantations − number of live fetuses)/number of implantations × 100.
2.13. Lipid Peroxidation
2.13.1. Homogenate Protein Determination
The testes from both sides were homogenized separately in 50 mM Tris-HCl buffer solution (pH 7.4). The protein content in the homogenate was determined by Bradford assay [40] using bovine serum albumin as a standard.
2.13.2. Thiobarbituric Acid Reactive Substances (TBARS) Assay
The level of lipid peroxidation was assessed using the TBARS assay by Draper and Hadley [41]. Briefly, 200 μL of the obtained homogenate was added to 500 μL thiobarbituric acid (0.8%), 500 μL acetic acid buffer, 200 μL of SDS (8.1%), and 100 μL of water. These samples were then incubated for 2 hours at 95°C and allowed to cool to room temperature. The absorbance was measured at 532 nm. All of the assays were performed in duplicate, and the amount of lipid peroxidation was expressed as nmol TBARS/g protein.
2.14. Statistical Analysis
Data were expressed as mean ± standard error of mean (SEM) or median and interquartile range. Parametric variables were compared by ANOVA followed by Tukey's test, nonparametric variables were compared by Kruskal-Wallis followed by Dunn's test, and χ2 test was used for categorical dependent variables. Differences were considered significant when p ≤ 0.05. The statistical analyses were performed by the software GraphPad Prism (version 5.0).
3. Results
3.1. Body and Organ Weights Were Not Altered by Varicocele
Final body weight and both relative and absolute reproductive organ weights were similar among experimental groups (Table 1).
Table 1.
Body and reproductive organ weights.
| Parameters | Experimental groups | ||
|---|---|---|---|
| Sham (n = 10) | V1 (n = 8) | V2 (n = 10) | |
| Final body weight (g) | 476.2 ± 14.96 | 453.1 ± 17.27 | 472.9 ± 16.45 |
| Testis (g) | 1.83 ± 0.08 | 1.81 ± 0.02 | 1.77 ± 0.04 |
| Testis (g/100 g) | 0.38 ± 0.01 | 0.41 ± 0.01 | 0.38 ± 0.01 |
| Epididymis (mg) | 647 ± 26.74 | 626.1 ± 14.99 | 619 ± 21.08 |
| Epididymis (mg/100 g) | 135.7 ± 2.99 | 143.1 ± 3.12 | 131.5 ± 4.00 |
| Ventral prostate (mg) | 546.3 ± 38.02 | 612.6 ± 31.50 | 514.6 ± 44.68 |
| Ventral prostate (mg/100 g) | 114.1 ± 5.71 | 135 ± 7.11 | 113 ± 6.71 |
| Seminal vesicle (g) | 1.18 ± 0.12 | 1.25 ± 0.07 | 1.08 ± 0.05 |
| Seminal vesicle (g/100 g) | 0.25 ± 0.02 | 0.26 ± 0.02 | 0.23 ± 0.01 |
Values are expressed as mean ± SEM, p > 0.05. ANOVA followed by Tukey's test.
3.2. Chrysin Improved Histological Aspect of the Testis and Maintained Morphometric Parameters Similar to the Sham Group
Histopathological analysis of epididymis did not reveale any alterations (data not shown) among experimental groups. However, varicocele induction significantly increased the number of seminiferous tubules with abnormalities in ipsilateral testis and the treatment with chrysin decreased this parameter to similar values to sham (Figures 1(a)–1(g)). Despite tubules from the contralateral side also presented this alteration, there were no differences compared to V1 and sham. The morphometric measurements showed that varicocele promoted a reduction in the diameter of seminiferous tubules in the ipsilateral side, and chrysin treatment maintained this parameter similar to the sham group. Additionally, the epithelial height had a significant decrease in comparison to sham. The contralateral seminiferous tubules of V2 had similar diameter and epithelium height to other groups (Figures 1(h) and 1(i)).
Figure 1.

Representative histological testicular aspect in ipsilateral (a, c, e) and contralateral (b, d, f) sides. L: lumen; Ep: epithelium; In: interstitial tissue. Note, in the V2 group, there are tubules with depletion of germ cells (∗). Final magnification: 200x. Percentage of normal seminiferous tubules (g), tubule diameter (h), and epithelium height (i) of animals from sham, V1, and V2 groups. Values expressed as median and interquartile range. Kruskal-Wallis analysis of variance test, followed by Dunn's test (g). Values expressed as mean ± SEM, p ≤ 0.05. ANOVA test followed by Tukey's test (h and i). Different letters indicate p ≤ 0.05.
3.3. Daily Sperm Production and Epididymal Sperm Counts, Serum and Intratesticular Testosterone Levels, and Sperm Morphology Were Not Altered by Varicocele
Sperm counts in the testis and epididymis of the V2 group (Table 2) as well as serum and intratesticular testosterone concentrations were very similar among experimental groups (Figure 2). Regarding sperm morphology, even though V2 group presented low percentages of normal sperm when compared to V1 and sham groups, the differences were not significant (Figure 3).
Table 2.
Adult male rats spermatic parameters.
| Parameters | Experimental groups | ||
|---|---|---|---|
| Sham (n = 8) | V1 (n = 8) | V2 (n = 6) | |
| Sperm count in the testis | |||
| Daily sperm production (×106/testis/day) | 45.79 ± 1.38 | 39.93 ± 0.67 | 42.64 ± 1.73 |
| Relative daily sperm production (×106/testis/g/day) | 27.78 ± 0.63 | 28.21 ± 0.75 | 25.96 ± 1.47 |
| Sperm count in the epididymis | |||
| Caput/corpus | |||
| Sperm number (×106/organ) | 197.4 ± 9.77 | 188.6 ± 4.42 | 207.2 ± 5.37 |
| Relative sperm number (×106/g/organ) | 465.4 ± 24.10 | 500.3 ± 13.43 | 463 ± 28.09 |
| Sperm transit time (days) | 4.25 ± 0.27 | 4.62 ± 0.13 | 4.89 ± 0.16 |
| Cauda | |||
| Sperm number (×106/organ) | 396.0 ± 30.45 | 357.6 ± 18.11 | 382.8 ± 18.91 |
| Relative sperm number (×106/g/organ) | 1458 ± 70.61 | 1447 ± 52.32 | 1451 ± 50.07 |
| Sperm transit time (days) | 6.69 ± 0.21 | 6.64 ± 0.25 | 6.96 ± 0.22 |
Values are expressed as mean ± SEM, p > 0.05. ANOVA followed by Tukey's test.
Figure 2.

Serum testosterone levels (a) and intratesticular testosterone levels (b) in the ipsilateral (sham i, V1 i, and V2 i) and contralateral side (Sham c, V1 c, and V2 c). Values are expressed as mean ± SEM, p > 0.05. ANOVA followed by Tukey's test.
Figure 3.
Sperm morphology. Percentage of spermatozoa with the presence of cytoplasmatic droplet in the ipsilateral (sham i, V1 i, and V2 i) and contralateral side (sham c, V1 c, and V2 c). Values are expressed as median and interquartile range. Kruskal-Wallis analysis of variance test, followed by Dunn's test (p > 0.05).
3.4. The Impairment of Sperm Motility Caused by Varicocele Was Not Significantly Improved by Chrysin Treatment
V2 group presented a significant decrease in mobile sperm only when compared to the ipsilateral side of sham (p < 0.02), and despite the treatment with chrysin had improved this parameter, no differences were documented (Figure 4).
Figure 4.

Sperm motility. Percentage of spermatozoa with type A (mobile with progression), type B (mobile without progression), and type C (immobile) in the ipsilateral (sham i, V1 i, and V2 i) and contralateral side (sham c, V1 c, and V2 c). Values expressed as median and interquartile range. Kruskal-Wallis analysis of variance test, followed by Dunn's test. Different letters indicate p ≤ 0.05.
3.5. Chrysin Reduced DNA Fragmentation Caused by Varicocele
The comet assay showed that varicocelized rats (V2) had an extremely significant increase in sperm DNA fragmentation (p ≤ 0.0001) in ipsilateral and contralateral sides, and the treatment with chrysin improved this parameter, decreasing the percentage of tail intensity to sham levels (Figure 5).
Figure 5.
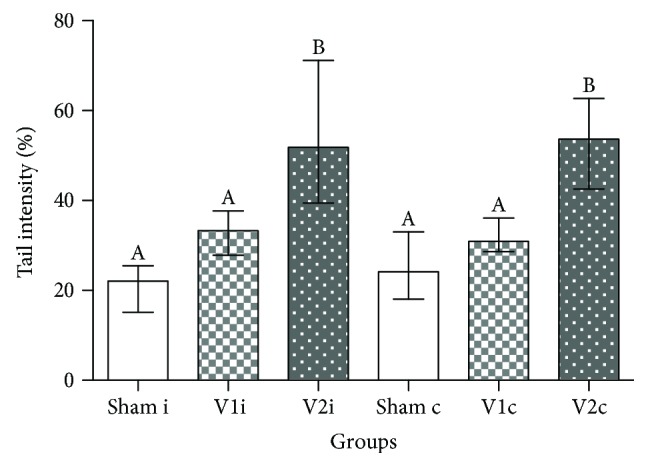
Comet Assay. Percentage of sperm DNA fragmentation through tail intensity in the ipsilateral (sham i, V1 i, and V2 i) and contralateral side (sham c, V1 c, and V2 c). Values expressed as median and interquartile range. Kruskal-Wallis analysis of variance test, followed by Dunn's test. Different letters indicate p ≤ 0.05.
3.6. Chrysin Reduced the Levels of Testicular Malondialdehyde
The determination of testicular malondialdehyde revealed that varicocelized animals presented high concentrations of this LPO product in ipsilateral (p < 0.0005) and contralateral (p < 0.01) sides when compared to chrysin and sham groups (Figure 6).
Figure 6.
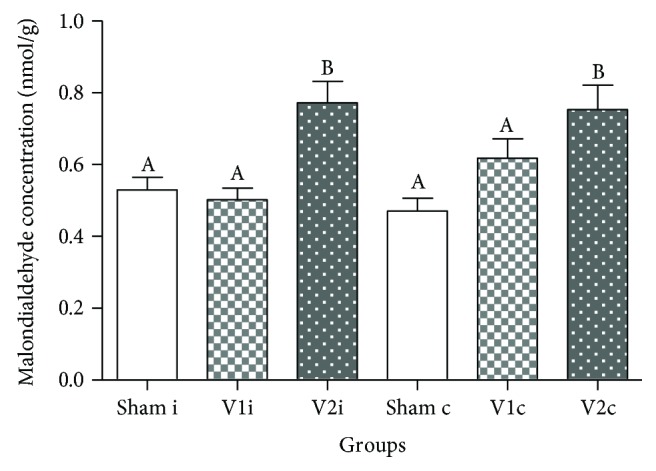
Malondialdehyde concentration in ipsilateral and contralateral testis of animals from sham, V1, and V2 groups. Values are expressed as mean ± SEM. ANOVA followed by Tukey's test was performed. Different letters indicate p ≤ 0.01.
3.7. Sexual Behavior or Reproductive Performance Was Not Altered by Varicocele
When sexual behavior was evaluated, there were no significant differences among experimental groups regarding the number and latency of mounts and intromissions, as well as ejaculation (Table 3). In the same manner, the reproductive performance was not significantly altered concerning pregnancy rate, pre- and postimplantation lost, weight of uterus plus fetuses, numbers of corpora lutea, implantation sites, reabsorptions, live fetuses, and fetal and dams weights (Table 4).
Table 3.
Sexual behavior.
| Parameters | Experimental groups | ||
|---|---|---|---|
| Sham (n = 9) | V1 (n = 7) | V2 (n = 10) | |
| Latency of 1st mount (s) | 66.5 ± 16.05 (9) | 36.71 ± 11.47 (7) | 53.50 ± 14.61 (10) |
| Number of mounts (n) | 11.67 ± 2.76 (9) | 15.43 ± 6.70 (7) | 10 ± 1.52 (10) |
| Latency of 1st intromission (s) | 196.7 ± 45.24 (9) | 374.7 ± 127.6 (7) | 328.8 ± 96.85 (9) |
| Number of intromissions until 1st ejaculation (n) | 22.33 ± 4.71 (9) | 24.43 ± 3.22 (7) | 22.33 ± 4.49 (9) |
| Latency of 1st ejaculation (s) | 899.1 ± 151.1 (8) | 1212 ± 206.7 (5) | 1050 ± 270.2 (5) |
| Number of ejaculations (n) | 2.43 ± 0.43 (7) | 1.33 ± 0.21 (5) | 2.0 ± 0.55 (5) |
Values are expressed as mean ± SEM, p > 0.05. ANOVA followed by Tukey's test.
Table 4.
Fertility test after natural mating.
| Parameters | Experimental groups | ||
|---|---|---|---|
| Sham (n = 7) | V1 (n = 8) | V2 (n = 7) | |
| 1Pregnancy rate (%) | 66.66 | 100 | 70 |
| 2Preimplantation loss (%) | 0.00 (0.00–12.50) (6) | 8.33 (7.69–25.00) (7) | 0.00 (0.00–7.69) (7) |
| 2Postimplantation loss (%) | 0.00 (0.00–2.27) (6) | 0.00 (0.00–8.33) (7) | 0.00 (0.00–9.09) (7) |
| 3Body weight of dams (g) | 364 ± 11.54 (7) | 365 ± 15.78 (8) | 379.9 ± 9.92 (7) |
| 3Uterus weight with fetuses (g) | 69.16 ± 6.89 (7) | 74.36 ± 4.32 (8) | 76.05 ± 4.2 (7) |
| 3Corpora lutea number | 12.57 ± 0.72 (7) | 13.25 ± 0.75 (8) | 12.71 ± 0.56 (7) |
| 3Implantation number | 12.00 ± 0.62 (7) | 11.5 ± 0.63 (8) | 12.29 ± 0.42 (7) |
| 3Number of live fetuses | 10.86 ± 1.32 (7) | 11.25 ± 0.70 (8) | 11.71 ± 0.64 (7) |
| 3Fetus weight (g) | 4.61 ± 0.20 (7) | 4.71 ± 0.07 (8) | 4.74 ± 0.13 (7) |
1 χ 2 test with results expressed in percentage. 2Values are expressed as median and interquartile intervals (Kruskal-Wallis test followed by Dunn's test, p > 0.05). 3Values are expressed as mean ± SEM, p > 0.05 ANOVA followed by Tukey's test.
4. Discussion
Our work is the first to report the protective effects of chrysin on reproductive disorders induced by experimental varicocele in rats. It also demonstrate the deleterious effects associated with this condition besides providing, for the first time, a more comprehensive evaluation of sexual behavior and reproductive performance in this experimental model and to evaluate intratesticular testosterone concentrations after an antioxidant treatment.
As previously reported, the principal mechanism responsible for deleterious effects of varicocele involves OS. The reactive molecules originated from OS, such as ROS, promote oxidative degradation of lipids in a process known as lipid peroxidation (LPO) [42]. Considering that sperm plasma membrane is mainly constituted by polyunsaturated fatty acids [43], these specialized cells became potential targets. Several authors have reported high levels of malondialdehyde (MDA), a LPO product in the semen [8, 44, 45] and testis [46], and showed a negative correlation with sperm parameters. In accordance with these studies [47, 48], our results revealed that varicocelized rats presented a significant bilateral increase of testicular MDA and the treatment with chrysin decreased these levels. Since several works have showed that chrysin was capable of enhancing levels of superoxide dismutase, catalase, and glutathione peroxidase in the testis [24, 49], liver [29, 50], kidney [50, 51], and colon [30], we strongly believe that this chemical probably reinforced the cell antioxidant defense preventing the increase of MDA [24, 26].
Besides compromising the integrity of the sperm plasma membrane, ROS may also interfere in cellular mechanisms through degradation of proteins. For that reason, nuclear and mitochondrial DNA of germ cells is highly susceptible to double-strand breaks [52]. Considering that the spermatozoa do not have the necessary antioxidant machinery to repair this damage, and the fact that high levels of DNA fragmentation are highly correlated with infertility [53], it is extremely important to protect this cell against ROS attack. In our experiments, chrysin treatment maintained DNA integrity similar to sham levels and varicocelized animals, in accordance with previous works [54–56], presented a significant bilateral increase in sperm DNA fragmentation. The positive impact of chrysin within this parameter might be due to its ability to increase levels of oxidative enzymes.
Moreover, another factor that is involved in the impairment of sperm quality is hypoxia. Studies have demonstrated that, along with OS [57], testis deoxygenation caused by venous stasis [58] increased the expression of pro-apoptotic proteins such as caspase [59], Bax, and Bad [60]. This combination of factors might explain the significant increase in abnormalities seen in testis histopathology of the V2 group, once the most frequent alteration observed was the depletion of germ cells. On the other hand, varicocelized rats treated with chrysin had similar percentage of normal seminiferous tubules compared to the sham group. Knowing that chrysin treatment decreased the expression of these proteins in the testis [61] and liver [26], we can conclude that this mechanism might be responsible, at least in part, for preventing cell apoptosis.
Similar to previous reports [59], another alteration verified within the testis of the V2 group was the diminution of seminiferous tubule diameter when compared to sham and V1 groups. In addition, the seminiferous epithelium in ipsilateral testis of varicocelized rats also had lower measures than the sham group. In this study, testicular temperature was not determined; however, we believe that hyperthermia, caused by venous stasis, is the probable cause of these alterations since long-term exposure to high temperatures can impair testicular morphometry [62].
Although varicocele is often associated with impaired spermatogenesis and hormonal dysfunction [63–66], our work showed that serum and intratesticular testosterone concentrations as well as daily sperm production in the testis and sperm number in epididymis were similar among experimental groups. This data contrasts with previous works that reported a decrease in serum testosterone and sperm reserves of varicocelized rats [67, 68]. Based on these observations, we can conclude that varicocele, at least in this experimental conditions, was not related to Leydig cell dysfunction which contrasts with previous reports [55]. Additionally, even though chrysin is known to prevent conversion of testosterone to estrogen, it did not influence androgen levels. We were the first to evaluate serum and intratesticular testosterone at the same time after antioxidant treatment, demonstrating that it was not altered by varicocele or chrysin treatment.
Regarding sperm motility, varicocele caused a significant decrease in mobile sperm in the ipsilateral side when compared to sham. This effect might be explained by hypoxia and by its sensitivity to oxidative stress [67]. Varicocele venous stasis decreases the supply of oxygen, which in turn increases the production of ROS. These molecules, besides interfering in the fluidity and integrity of the sperm membrane through LPO [8], can impair the synthesis of proteins involved in energy metabolism pathways, transport, and proton pumps causing energy deprivation [68]. Even though the treatment with chrysin improved this parameter, there were no differences in comparison with other groups. In the same manner and according to a study conducted by the World Health Organization [2], sperm morphology remained unaltered among experimental groups even though varicocelized rats had a lower percentage of normal sperm when compared to sham and V1.
Previous works documented, through artificial insemination [69] and natural mating [70], respectively, that pregnancy rate was significantly reduced after experimental varicocele induction. These were the only works that actually performed a fertility test on varicocelized rats, but the evidences were not sufficient to comprehend the impact that this condition has on fertility. For that reason, our work showed remarkable data regarding post- and preimplantation loss, besides giving information about dams and offspring. Furthermore, we were the first to evaluate the sexual behavior of these animals. Despite that our findings have shown that varicocele caused sperm quality impairment, there were no differences in reproductive performance and sexual behavior among experimental groups. We recognize that performing the fertility test after 56 days of chrysin treatment did not cover the whole time taken for the sperm production and transit through the epididymis and genital ducts until ejaculation. Also, natural mating may not be sensible enough to test fertility in rats. However, considering that men have lower reproductive efficiency than rats [71, 72], these results raise concern about men's reproductive health and infertility.
In conclusion, our data showed that a varicocele had a bilateral character, despite causing more severe damages in ipsilateral testis and reinforcing that OS is a key element in its pathophysiology once several testicular and sperm alterations, associated with high concentrations of MDA, were observed in varicocelized rats. It also contributed with new information regarding the sexual behavior and reproductive performance in this experimental model. Furthermore, chrysin proved to be a potent antioxidant agent, capable of protecting germ cells against the deleterious effects of varicocele, by decreasing testicular MDA concentrations and sperm DNA fragmentation and improving sperm quality. For that reason, the use of this compound as an adjuvant therapy should be considered in order to improve sperm quality of men presenting this condition.
Acknowledgments
The authors would like to acknowledge Dr. André Luis Philadelpho, Department of Anatomy, Dr. Maria José Queiroz de Freitas Alves, Department of Physiology, José Eduardo Bozano, Department of Morphology, from the Institute of Biosciences, São Paulo State University (UNESP), Dr. Daisy Maria Favero Salvadori and Dr. Fábio Henrique Fernandes, Department of Pathology, Medical School, Botucatu, and Dr. Janete Aparecida Anselmo Franci and Dr. Ruither de Oliveira Gomes Carolino, Department of Morphology, Stomatology and Physiology, School of Dentistry, University of São Paulo, Ribeirão Preto, for the excellent technical assistance. The authors are also grateful to the State of São Paulo Research Foundation (FAPESP) (Grant no. 2015/26309-3), Coordinator for Improvement of Higher Education Personnel (CAPES), and National Council for Scientific and Technological Development (CNPq) (Grant no. 308842/2013-8).
Disclosure
Dr. Wilma De Grava Kempinas is a recipient of a senior research fellowship from CNPq (National Council for Scientific and Technological Development).
Conflicts of Interest
The authors declare that there are no conflicts of interests.
References
- 1.Smith R., Kaune H., Parodi D., et al. Increased sperm DNA damage in patients with varicocele: relationship with seminal oxidative stress. Human Reproduction. 2006;21(4):986–993. doi: 10.1093/humrep/dei429. [DOI] [PubMed] [Google Scholar]
- 2.World Health Organization. The influence of varicocele on parameters of fertility in a large group of men presenting to infertility clinics. Fertility and Sterility. 1992;57(6):1289–1293. doi: 10.1016/S0015-0282(16)55089-4. [DOI] [PubMed] [Google Scholar]
- 3.Gat Y., Bachar G. N., Zukerman Z., Belenky A., Gornish M. Varicocele: a bilateral disease. Fertility and Sterility. 2004;81(2):424–429. doi: 10.1016/j.fertnstert.2003.08.010. [DOI] [PubMed] [Google Scholar]
- 4.Witt M. A., Lipshultz L. I. Varicocele: a progressive or static lesion? Urology. 1993;42(5):541–543. doi: 10.1016/0090-4295(93)90268-F. [DOI] [PubMed] [Google Scholar]
- 5.Gokce A., Davarci M., Yalcinkaya F. R., et al. Hereditary behavior of varicocele. Journal of Andrology. 2010;31(3):288–290. doi: 10.2164/jandrol.109.008698. [DOI] [PubMed] [Google Scholar]
- 6.Marmar J. L. Varicocele and male infertility: part II: the pathophysiology of varicoceles in the light of current molecular and genetic information. Human Reproduction Update. 2001;7(5):461–472. doi: 10.1093/humupd/7.5.461. [DOI] [PubMed] [Google Scholar]
- 7.Cho C. L., Esteves S. C., Agarwal A. Novel insights into the pathophysiology of varicocele and its association with reactive oxygen species and sperm DNA fragmentation. Asian Journal of Andrology. 2016;18(2):186–193. doi: 10.4103/1008-682X.170441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Mostafa T., Anis T. H., El-Nashar A., Imam H., Othman I. A. Varicocelectomy reduces reactive oxygen species levels and increases antioxidant activity of seminal plasma from infertile men with varicocele. International Journal of Andrology. 2001;24(5):261–265. doi: 10.1046/j.1365-2605.2001.00296.x. [DOI] [PubMed] [Google Scholar]
- 9.Hsiao W., Rosoff J. S., Pale J. R., Powell J. L., Goldstein M. Varicocelectomy is associated with increases in serum testosterone independent of clinical grade. Urology. 2013;81(6):1213–1218. doi: 10.1016/j.urology.2013.01.060. [DOI] [PubMed] [Google Scholar]
- 10.Abdel-Meguid T. A., Farsi H. M., Al-Sayyad A., Tayib A., Mosli H. A., Halawani A. H. Effects of varicocele on serum testosterone and changes of testosterone after varicocelectomy: a prospective controlled study. Urology. 2014;84(5):1081–1087. doi: 10.1016/j.urology.2014.05.029. [DOI] [PubMed] [Google Scholar]
- 11.Schatte E. C., Hirshberg S. J., Fallick M. L., Lipschultz L. I., Kim E. D. Varicocelectomy improves sperm strict morphology and motility. The Journal of Urology. 1998;160(4):1338–1340. doi: 10.1016/S0022-5347(01)62531-X. [DOI] [PubMed] [Google Scholar]
- 12.Masson P., Brannigan R. E. The varicocele. The Urologic Clinics of North America. 2014;41(1):129–144. doi: 10.1016/j.ucl.2013.08.001. [DOI] [PubMed] [Google Scholar]
- 13.Al-Rubiey F. K. Effect of L-carnitine and meloxicam treatment on testicular Leydig cell numbers of varicocelized rats. Middle East Fertility Society Journal. 2012;17(1):47–53. doi: 10.1016/j.mefs.2011.08.009. [DOI] [Google Scholar]
- 14.Sohrabipour S., Jafari A., Kamalinejad M., Sarrafnejd A., Shahrestany T., Sadeghipour H. R. The role of flaxseed and vitamin E on oxidative stress in prepubertal rats with experimental varicocele: an experimental study. Iranian Journal of Reproductive Medicine. 2013;11(6):459–466. [PMC free article] [PubMed] [Google Scholar]
- 15.Mendes T. B., Paccola C. C., de Oliveira Neves F. M., et al. Resveratrol improves reproductive parameters of adult rats varicocelized in peripuberty. Reproduction. 2016;152(1):23–35. doi: 10.1530/REP-16-0025. [DOI] [PubMed] [Google Scholar]
- 16.Kellis J., Vickery L. Inhibition of human estrogen synthetase (aromatase) by flavones. Science. 1984;225(4666):1032–1034. doi: 10.1126/science.6474163. [DOI] [PubMed] [Google Scholar]
- 17.Sanderson J. T., Hordijk J., Denison M. S., Springsteel M. F., Nantz M. H., van den Berg M. Induction and inhibition of aromatase (CYP19) activity by natural and synthetic flavonoid compounds in H295R human adrenocortical carcinoma cells. Toxicological Sciences. 2004;82(1):70–79. doi: 10.1093/toxsci/kfh257. [DOI] [PubMed] [Google Scholar]
- 18.Kasala E. R., Bodduluru L. N., Madana R. M., V A. K., Gogoi R., Barua C. C. Chemopreventive and therapeutic potential of chrysin in cancer: mechanistic perspectives. Toxicology Letters. 2015;233(2):214–225. doi: 10.1016/j.toxlet.2015.01.008. [DOI] [PubMed] [Google Scholar]
- 19.Darwish H. A., Arab H. H., Abdelsalam R. M. Chrysin alleviates testicular dysfunction in adjuvant arthritic rats via suppression of inflammation and apoptosis: comparison with celecoxib. Toxicology and Applied Pharmacology. 2014;279(2):129–140. doi: 10.1016/j.taap.2014.05.018. [DOI] [PubMed] [Google Scholar]
- 20.Rauf A., Khan R., Raza M., et al. Suppression of inflammatory response by chrysin, a flavone isolated from Potentilla evestita Th. Wolf. In silico predictive study on its mechanistic effect. Fitoterapia. 2015;103:129–135. doi: 10.1016/j.fitote.2015.03.019. [DOI] [PubMed] [Google Scholar]
- 21.Villar I. C., Jimenez R., Galisteo M., Garcia-Saura M. F., Zarzuelo A., Duarte J. Effects of chronic chrysin treatment in spontaneously hypertensive rats. Planta Medica. 2002;68(9):847–850. doi: 10.1055/s-2002-34400. [DOI] [PubMed] [Google Scholar]
- 22.Souza L. C., Antunes M. S., Filho C. B., Del Fabbro L., de Gomes M. G. Flavonoid chrysin prevents age-related cognitive decline via attenuation of oxidative stress and modulation of BDNF levels in aged mouse brain. Pharmacology, Biochemistry, and Behavior. 2015;134:22–30. doi: 10.1016/j.pbb.2015.04.010. [DOI] [PubMed] [Google Scholar]
- 23.Dhawan K., Kumar S., Sharma A. Beneficial effects of chrysin and benzoflavone on virility in 2-year-old male rats. Journal of Medicinal Food. 2002;5(1):43–48. doi: 10.1089/109662002753723214. [DOI] [PubMed] [Google Scholar]
- 24.Ciftci O., Ozdemir I., Aydin M., Beytur A. Beneficial effects of chrysin on the reproductive system of adult male rats. Andrologia. 2012;44(3):181–186. doi: 10.1111/j.1439-0272.2010.01127.x. [DOI] [PubMed] [Google Scholar]
- 25.Jana K., Yin X., Schiffer R. B., et al. Chrysin, a natural flavonoid enhances steroidogenesis and steroidogenic acute regulatory protein gene expression in mouse Leydig cells. The Journal of Endocrinology. 2008;197(2):315–323. doi: 10.1677/JOE-07-0282. [DOI] [PubMed] [Google Scholar]
- 26.Ali N., Rashid S., Nafees S., Hasan S. K., Sultana S. Beneficial effects of chrysin against methotrexate-induced hepatotoxicity via attenuation of oxidative stress and apoptosis. Molecular and Cellular Biochemistry. 2014;385(1-2):215–223. doi: 10.1007/s11010-013-1830-4. [DOI] [PubMed] [Google Scholar]
- 27.Manzolli E. S., Serpeloni J. M., Grotto D., et al. Protective effects of the flavonoid chrysin against methylmercury-induced genotoxicity and alterations of antioxidant status, in vivo. Oxidative Medicine and Cellular Longevity. 2015;2015:7. doi: 10.1155/2015/602360.602360 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Russell L. D., Ettlin R. A., Sinha Hikim A. P., Clegg E. D. Histological and Histopathological Evaluation of the Testis. Montreal: Cache River Press; 1990. [Google Scholar]
- 29.Rehman M. U., Ali N., Rashid S., et al. Alleviation of hepatic injury by chrysin in cisplatin administered rats: probable role of oxidative and inflammatory markers. Pharmacological Reports. 2014;66(6):1050–1059. doi: 10.1016/j.pharep.2014.06.004. [DOI] [PubMed] [Google Scholar]
- 30.Khan R., Khan A. Q., Qamar W., et al. Chrysin protects against cisplatin-induced colon. toxicity via amelioration of oxidative stress and apoptosis: probable role of p38MAPK and p53. Toxicology and Applied Pharmacology. 2012;258(3):315–329. doi: 10.1016/j.taap.2011.11.013. [DOI] [PubMed] [Google Scholar]
- 31.Turner T. T. The study of varicocele through the use of animal models. Human Reproduction Update. 2001;7(1):78–84. doi: 10.1093/humupd/7.1.78. [DOI] [PubMed] [Google Scholar]
- 32.Seed J., Chapin R. E., Clegg E. D., et al. Methods for assessing sperm motility, morphology, and counts in the rat, rabbit, and dog: a consensus report. Reproductive Toxicology. 1996;10(3):237–244. doi: 10.1016/0890-6238(96)00028-7. [DOI] [PubMed] [Google Scholar]
- 33.Filler R. Methods for evaluation of rats epididymal sperm morphology. In: Chapin R. E., Heindel J. J., editors. Male Reproductive Toxicology. San Diego, CA, USA: Academic Press; 1993. pp. 334–343. [Google Scholar]
- 34.Codrington A. M., Hales B. F., Robaire B. Spermiogenic germ cell phase—specific DNA damage following cyclophosphamide exposure. Journal of Andrology. 2004;25(3):354–362. doi: 10.1002/j.1939-4640.2004.tb02800.x. [DOI] [PubMed] [Google Scholar]
- 35.Robb G. W., Amann R. P., Killian G. J. Daily sperm production and epididymal sperm reserves of pubertal and adult rats. Journal of Reproduction and Fertility. 1978;54(1):103–107. doi: 10.1530/jrf.0.0540103. [DOI] [PubMed] [Google Scholar]
- 36.Fernandez C. D. B., Porto E. M., Arena A. C., Kempinas W. G. Effects of altered epididymal sperm transit time on sperm quality. International Journal of Andrology. 2008;31(4):427–437. doi: 10.1111/j.1365-2605.2007.00788.x. [DOI] [PubMed] [Google Scholar]
- 37.Fernandes G. S. A., Arena A. C., Fernandez C. D. B., Mercadante A., Barbisan L. F., Kempinas W. G. Reproductive effects in male rats exposed to diuron. Reproductive Toxicology. 2007;23(1):106–112. doi: 10.1016/j.reprotox.2006.09.002. [DOI] [PubMed] [Google Scholar]
- 38.Ahlenius S., Larsson K. Apomorphine and haloperidol-induced effects on male rat sexual behavior: no evidence for actions due to stimulation of central dopamine autoreceptors. Pharmacology, Biochemistry, and Behavior. 1984;21(3):463–466. doi: 10.1016/S0091-3057(84)80111-2. [DOI] [PubMed] [Google Scholar]
- 39.Agmo A. Male rat sexual behavior. Brain Research Protocols. 1997;1(2):203–209. doi: 10.1016/S1385-299X(96)00036-0. [DOI] [PubMed] [Google Scholar]
- 40.Bradford M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry. 1976;72(1-2):248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- 41.Draper H. H., Hadley M. [43] Malondialdehyde determination as index of lipid peroxidation. Methods in Enzymology. 1990;186:421–431. doi: 10.1016/0076-6879(90)86135-I. [DOI] [PubMed] [Google Scholar]
- 42.Ayala A., Munoz M. F., Arguelles S. Lipid peroxidation: production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxidative Medicine and Cellular Longevity. 2014;2014:31. doi: 10.1155/2014/360438.360438 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Agarwal A., Sharma R. K., Desai N. R., Prabakaran S., Tavares A., Sabanegh E. Role of oxidative stress in pathogenesis of varicocele and infertility. Urology. 2009;73(3):461–469. doi: 10.1016/j.urology.2008.07.053. [DOI] [PubMed] [Google Scholar]
- 44.Rao B., Soufir J. C., Martin M., David G. Lipid peroxidation in human spermatozoa as relatd to midpiece abnormalities and motility. Gamete Research. 1989;24(2):127–134. doi: 10.1002/mrd.1120240202. [DOI] [PubMed] [Google Scholar]
- 45.Abd-Elmoaty M. A., Saleh R., Sharma R., Agarwal A. Increased levels of oxidants and reduced antioxidants in semen of infertile men with varicocele. Fertility and Sterility. 2010;94(4):1531–1534. doi: 10.1016/j.fertnstert.2009.12.039. [DOI] [PubMed] [Google Scholar]
- 46.Koksal I. T., Tefekli A., Usta M., Erol H., Abbasoglu S., Kadioglu A. The role of reactive oxygen species in testicular dysfunction associated with varicocele. BJU International. 2000;86(4):549–552. doi: 10.1046/j.1464-410X.2000.00755.x. [DOI] [PubMed] [Google Scholar]
- 47.Gholirad S., Razi M., Hassani Bafrani H. Tracing of zinc and iron in experimentally induced varicocele: correlation with oxidative, nitrosative and carbonyl stress. Andrologia. 2017;49(6, article e12687) doi: 10.1111/and.12687. [DOI] [PubMed] [Google Scholar]
- 48.Ozturk H., Tander B., Aydin A., Okumus Z., Cetinkursun S. The effects of chemical sympathectomy on testicular injury in varicocele. BJU International. 2001;87(3):232–234. doi: 10.1046/j.1464-410x.2001.01987.x. [DOI] [PubMed] [Google Scholar]
- 49.Aksu E. H., Ozkaraca M., Kandemir F. M., et al. Mitigation of paracetamol-induced reproductive damage by chrysin in male rats via reducing oxidative stress. Andrologia. 2016;48(10):1145–1154. doi: 10.1111/and.12553. [DOI] [PubMed] [Google Scholar]
- 50.Pushpavalli G., Kalaiarasi P., Veeramani C., Pugalendi K. V. Effect of chrysin on hepatoprotective and antioxidant status in d-galactosamine-induced hepatitis in rats. European Journal of Pharmacology. 2010;631(1–3):36–41. doi: 10.1016/j.ejphar.2009.12.031. [DOI] [PubMed] [Google Scholar]
- 51.Sultana S., Verma K., Khan R. Nephroprotective efficacy of chrysin against cisplatin-induced toxicity via attenuation of oxidative stress. The Journal of Pharmacy and Pharmacology. 2012;64(6):872–881. doi: 10.1111/j.2042-7158.2012.01470.x. [DOI] [PubMed] [Google Scholar]
- 52.Schraufstatter I. U., Hinshaw D. B., Hyslop P. A., Spragg R. G., Cochrane C. G. Oxidant injury of cells. DNA strand-breaks activate polyadenosine diphosphate-ribose polymerase and lead to depletion of nicotinamide adenine dinucleotide. The Journal of Clinical Investigation. 1986;77(4):1312–1320. doi: 10.1172/JCI112436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Sharma R. K., Agarwal A. Role of reactive oxygen species in male infertility. Urology. 1996;48(6):835–850. doi: 10.1016/S0090-4295(96)00313-5. [DOI] [PubMed] [Google Scholar]
- 54.Aitken R. J., Smith T. B., Jobling M. S., Baker M. A., De Iuliis G. N. Oxidative stress and male reproductive health. Asian Journal of Andrology. 2014;16(1):31–38. doi: 10.4103/1008-682X.122203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Naughton C. K., Nangia A. K., Agarwal A. Varicocele and male infertility: part II: pathophysiology of varicoceles in male infertility. Human Reproduction Update. 2001;7(5):473–481. doi: 10.1093/humupd/7.5.473. [DOI] [PubMed] [Google Scholar]
- 56.Saleh R. A., Agarwal A., Sharma R. K., Said T. M., Sikka S. C., Thomas A. J., Jr. Evaluation of nuclear DNA damage in spermatozoa from infertile men with varicocele. Fertility and Sterility. 2003;80(6):1431–1436. doi: 10.1016/S0015-0282(03)02211-8. [DOI] [PubMed] [Google Scholar]
- 57.Thompson C. Apoptosis in the pathogenesis and treatment of disease. Science. 1995;267(5203):1456–1462. doi: 10.1126/science.7878464. [DOI] [PubMed] [Google Scholar]
- 58.Goren M. R., Kilinc F., Kayaselcuk F., Ozer C., Oguzulgen I., Hasirci E. Effects of experimental left varicocele repair on hypoxia-inducible factor-1α and vascular endothelial growth factor expressions and angiogenesis in rat testis. Andrologia. 2017;49(2) doi: 10.1111/and.12614. [DOI] [PubMed] [Google Scholar]
- 59.Bayomy N. A., Sarhan N. I., Abdel-Razek K. M. Effect of an experimental left varicocele on the bilateral testes of adult rats: a histological and immunohistochemical study. The Egyptian Journal of Histology. 2012;35(3):509–519. doi: 10.1097/01.EHX.0000418065.13002.11. [DOI] [Google Scholar]
- 60.Oh Y. S., Jo N. H., Park J. K., Gye M. C. Changes in inflammatory cytokines accompany deregulation of claudin-11, resulting in inter-sertoli tight junctions in varicocele rat testes. The Journal of Urology. 2016;196(4):1303–1312. doi: 10.1016/j.juro.2016.05.004. [DOI] [PubMed] [Google Scholar]
- 61.El-Sisi A. E., El-Sayad M. E., Abdelsalam N. M. Protective effects of mirtazapine and chrysin on experimentally induced testicular damage in rats. Biomedicine & Pharmacotherapy. 2017;95:1059–1066. doi: 10.1016/j.biopha.2017.09.022. [DOI] [PubMed] [Google Scholar]
- 62.Kurowicka B., Dietrich G. J., Kotwica G. Effect of neonatal or adult heat acclimation on testicular and epididymal morphometry and sperm production in rats. Reproductive Biology. 2015;15(1):1–8. doi: 10.1016/j.repbio.2015.01.001. [DOI] [PubMed] [Google Scholar]
- 63.Cayan S., Kadioglu A., Orhan I., Kandirali E., Tefekli A., Tellaloglu S. The effect of microsurgical varicocelectomy on serum follicle stimulating hormone, testosterone and free testosterone levels in infertile men with varicocele. BJU International. 1999;84(9):1046–1049. doi: 10.1046/j.1464-410x.1999.00353.x. [DOI] [PubMed] [Google Scholar]
- 64.Damsgaard J., Joensen U. N., Carlsen E., et al. Varicocele is associated with impaired semen quality and reproductive hormone levels: a study of 7035 healthy young men from six European countries. European Urology. 2016;70(6):1019–1029. doi: 10.1016/j.eururo.2016.06.044. [DOI] [PubMed] [Google Scholar]
- 65.Rajfer J., Turner T. T., Rivera F., Howards S. S., Sikka S. C. Inhibition of testicular testosterone biosynthesis following experimental varicocele in rats. Biology of Reproduction. 1987;36(4):933–937. doi: 10.1095/biolreprod36.4.933. [DOI] [PubMed] [Google Scholar]
- 66.Zheng Y., Zhang X., Zhou J., Cheng F., Zhou B. Effects on the ipsilateral testis during progression of experimental varicocele in rat. Medical Science Monitor. 2008;14(6):BR122–BR126. [PubMed] [Google Scholar]
- 67.Hamada A., Sharma R., du Plessis S. S., et al. Two-dimensional differential in-gel electrophoresis–based proteomics of male gametes in relation to oxidative stress. Fertility and Sterility. 2013;99(5):1216–1226.e2. doi: 10.1016/j.fertnstert.2012.11.046. [DOI] [PubMed] [Google Scholar]
- 68.Agarwal A., Sharma R., Samanta L., Durairajanayagam D., Sabanegh E. Proteomic signatures of infertile men with clinical varicocele and their validation studies reveal mitochondrial dysfunction leading to infertility. Asian Journal of Andrology. 2016;18(2):282–291. doi: 10.4103/1008-682X.170445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Sofikitis N., Takahashi C., Nakamura I., Hirakawa S., Miyagawa I. Surgical repair of secondary right varicocele in rats with primary left varicocele: effects on fertility, testicular temperature, spermatogenesis, and sperm maturation. Archives of Andrology. 1992;28(1):43–52. doi: 10.3109/01485019208987679. [DOI] [PubMed] [Google Scholar]
- 70.Heidari R., Alizadeh R., Abbasi N., et al. Do Pilea microphylla improve sperm DNA fragmentation and sperm parameters in varicocelized rats? Acta Medica Iranica. 2015;53(9):547–554. [PubMed] [Google Scholar]
- 71.Amann R. Use of animal models for detecting specific alterations in reproduction. Fundamental and Applied Toxicology. 1982;2(1):13–26. doi: 10.1016/S0272-0590(82)80059-6. [DOI] [PubMed] [Google Scholar]
- 72.Kempinas W. D. G., Klinefelter G. R. The epididymis as a target for toxicants. In: McQueen C. A. , editor. Comprehensive Toxicology. Oxford: Academic Press; 2014. pp. 149–166. [Google Scholar]