

Abstract
The crosstalk between tumor cells and bone cells in the bone microenvironment is crucial to understanding the mechanism of bone metastasis formation. We developed an in vitro fully human preclinical model of a co-culture of breast cancer cells and monocytes undergoing differentiation towards osteoclasts. We optimized a model of osteoclastogenesis starting from a sample of peripheral blood collected from healthy donors. Peripheral blood mononuclear cells (PBMCs) were first separated by density gradient centrifugation, seeded at a high density and induced to differentiate by adding two growth factors (GFs): receptor activator of nuclear factor-κB ligand (RANKL) and macrophage colony-stimulating factor(MCSF). The cells were left in culture for 14 days and then fixed and analyzed by downstream analysis. In osteolytic bone metastases, one of the effects of cancer cell arrival in bone is the induction of osteoclastogenesis. We thus challenged our model with co-cultures of breast cancer cells to study the differentiation power of cancer cells with respect to GFs. A straightforward way of studying cancer cell-osteoclast interaction is to perform indirect co-cultures based on the use of conditioned medium collected from breast cancer cell cultures and mixed with fresh medium. This mixture is then used to induce osteoclast differentiation. We also optimized a method of direct co-culture in which cancer cells and monocytes undergoing differentiation share the medium and exchange secreted factors. This is a significant improvement over the original indirect co-culture method as researchers can observe the reciprocal interactions of the two cell types and perform downstream analyses for both cancer cells and osteoclasts. This method enables us to study the effect of drugs on the metastatic bone microenvironment and to seed cell lines other than those derived from breast cancer. The model can also be used to study other diseases such as osteoporosis or other bone conditions.
Keywords: Cancer Research, Issue 127, osteoclastogenesis, cancer cells, co-cultures, RANKL, MCSF, bone metastasis, in vitro, human preclinical model
Introduction
Bone is a common site of metastasis for different types of primary tumors such as prostate, lung and breast cancer, with 20 - 25% of patients developing bone metastases during the course of disease1,2,3. In particular, 70% of breast cancer patients carry evidence of bone metastasis at death4. Tumor and stromal cell interaction is essential for cancer progression in both primary cancer and secondary lesions. In the bone microenvironment, osteolytic bone metastases from breast cancer depend on the establishment of a pathological vicious cycle occurring between cancer cells, bone cells, and the bone microenvironment. Cancer cells disrupt bone balance, increasing bone resorption5,6,7.
In normal and pathological conditions, osteoclasts are the cells responsible for bone resorption, whereas osteoblasts, in depositing new matrix, are responsible for new bone formation8. Osteoclast activity is regulated by osteoblasts through the expression of RANKL, which binds to its receptor RANK on the pre-osteoclast surface, inducing pre-osteoclast fusion, a necessary process for differentiation into mature osteoclasts. The induction of osteoclastogenesis increases bone resorption. A large number of in vivo studies have significantly improved our understanding of bone metastasis formation9,10,11. Breast cancer cells from the primary tumor and in the bone microenvironment perturb bone homeostasis, promoting osteoclastogenesis and bone resorption8. In this scenario, all the molecular interactions that occur between cancer cells and osteoclasts are of crucial importance. As already mentioned, the mechanism of bone metastasis formation has been elucidated in in vivo mice models. However, in addition to the need for the approval of all in vivo animal experiments by the Ethics Committee, there are several other drawbacks to performing in vivo experiments including high costs and time-consuming methods. Several authors have combined preclinical in vivo and in vitro models of osteoclastogenesis using a murine line of pre-osteoclasts called RAW246.79,10,11. The drawbacks of this model stem from the fact that the cells are already committed to becoming pre-osteoclasts and are not of human origin. For these reasons, translational research could greatly benefit from the availability of in vitro fully human preclinical models to study bone cancer cell interactions.
We optimized a method of osteoclastogenesis in vitro starting from human peripheral blood samples12,13. Osteoclasts derive from monocytes, which are present, albeit to a small degree, in peripheral blood samples. Mononuclear cells are first separated from the erythrocytes and granulocytes present in whole blood by Ficoll density gradient; they are then selected thanks to their ability to adhere to plastic substrate, unlike lymphocytes. After seeding, cells are cultured for 14 days. MCSF and RANKL are the GFs required by monocytes to differentiate first into macrophages and then into osteoclasts14,15. MCSF is needed for the entire duration of the assay, whereas RANKL is used to induce the differentiation process in the late stages of osteoclastogenesis. In the early phase of differentiation, MCSF helps monocytes proliferate and survive14,15. During the second part of osteoclastogenesis, cells fuse together and mature as osteoclasts, showing the characteristic distribution of Actin F in rings and expressing specific markers such as tartrate-resistant acid phosphatase (TRAP) and calcitonin receptor (CTR)14,15. Our method consists of adding MCSF to the monocyte culture for the first 7 days of the experiment and a combination of MCSF and RANKL from days 7 to 14. At the end of the experiment, osteoclastogenesis is analyzed by counting the differentiated cells, as detailed below.
The monocyte cultures induced to differentiate by GFs form the basis of our preclinical model. We optimized a co-culture system without GFs to better understand the osteoclastogenic power of breast cancer cells. We first developed a model of indirect co-cultures by adding a medium (80% α -Minimal Essential Medium (α-MEM) and 20% conditioned medium collected from a culture of breast cancer cells that were about 90% confluent to cells undergoing differentiation12. The conditioned medium (not collected under serum deprivation conditions) was collected after 24 hours and mixed with fresh medium at a proportion of 1:4. The conditioned medium induced significant osteoclast differentiation with respect to the negative control. However, as the information on the reciprocal interaction between cancer cells and bone cells is lost when using indirect co-cultures, we improved our system by carrying out direct co-cultures. We seeded cancer cells in 0.4 µM inserts and placed them in wells where mononuclear cells were plated. Using this method, cells share the same medium and exchange secreted proteins. We thus created a fully human preclinical model of osteoclastogenesis induced by cancer cells13.
This system is extremely versatile and can be used for different research purposes, e.g., in pharmacological studies investigating the role of drugs in bone metastasis. Our model makes it possible to study the efficacy and mechanisms of action of bone-targeted therapies and/or antitumor drugs in the bone microenvironment in the presence of cancer cells13. Designing the experiments with the correct controls, i.e., cancer cells and osteoclasts cultured individually, makes it easier to understand the impact of the co-culture on drug activity. This approach becomes even more interesting when the drug being studied targets both cancer cells and osteoclasts, e.g., everolimus16. This model can also be used to identify new pathways of interaction between cancer cells and bone cells.
Protocol
Human osteoclasts were differentiated from PBMCs of healthy donors who gave written informed consent to take part in the study. The study protocol was approved by the local Ethics Committee, in accordance with the ethical standards laid down in the 1964 Declaration of Helsinki.
1. Osteoclast Differentiation
NOTE: Collect peripheral blood or buffy coats from healthy human donors in EDTA. Do not use less than 20 mL of peripheral blood. Buffy coats are preferable because they have a greater abundance of mononuclear cells. Perform all the following steps in a sterile tissue culture hood.
Dilute EDTA whole blood 1:1 with 1x phosphate-buffered saline (PBS). Re-suspend well.
- Layer on lymphocyte separation media (blood PBS: lymphocyte separation media, 2:1) in 50 mL tubes.
- Pipette 30 mL of blood diluted in PBS into a 50 mL tube.
- Gently underlay 15 mL of lymphocyte separation media into the bottom of the 50-mL tube.
Centrifuge at 757 × g for 10 min at room temperature (no brakes).
- Collecting the mononuclear cells:
- Collect the white layer of mononuclear cells with a 5 mL pipette without agitating and place it in a new 50 mL tube.
- Dilute the collected PBMCs with 20 mL of PBS.
- Washing the PBMCs:
- Centrifuge at 225 x g for 5 min at room temperature.
- Discard the supernatant by inverting the tube. Repeat once.
- RBC lysis: NOTE: If the cell pellet is red, treat cells with Red Blood cell lysing buffer solution (see Table of Materials).
- Put the 50 mL tube on ice.
- Add 5 mL of Red Blood cell lysing buffer.
- Wait 3 - 5 min to complete the erythrocyte lysis. The time varies according to the degree of erythrocyte contamination.
- Stop the reaction by adding 20 mL of PBS.
- Centrifuge at 225 x g for 5 min at room temperature. Discard the supernatant by inverting the tube. NOTE: Optional: If the RBC lysis is performed, wash the PBMCs as described below.
- Wash PBMCs:
- Add 20 mL of PBS and centrifuge at 225 x g for 5 min at room temperature.
- Discard the supernatant by inverting the tube.
Re-suspend the cells in complete α-MEM (supplemented with 1% glutamine (initial concentration 100X) and 10% fetal bovine serum).
- Count the cells with a Neubauer chamber.
- Dilute the sample 1:100 in glacial acetic acid and use 10 µL of the dilution to count cells.
- Count at least twice and calculate the mean of replicates to get the total mean number of cells collected.
- Plate cells:
- Plate cells at a concentration of 750,000 PBMCs per cm2.
- Store plated cells in an incubator at 37 °C in a 5% CO2 atmosphere. NOTE: Design the experiment to include negative controls, which are cells that are treated with normal complete α-MEM without MCSF or RANKL GFs. Remember to perform each condition in at least 3 technical replicates. Perform at least 3 biological replicates.
- Change the medium supplemented with GFs:
- Wait about 3 h after the cell seeding and then remove the medium by pipette (200 - 1,000 µL) to discard debris, unattached cells, and erythrocytes. NOTE: Be careful when changing the medium to avoid cell detachment.
- Add new complete α-MEM supplemented with 20 ng/mL of MCSF (500 µL/well). NOTE: When co-culturing with cancer cells, seed the cancer cells the day after the PBMC seeding.
- Change the media every 2 - 3 days (α-MEM supplemented with 20 ng/mL of MCSF):
- Discard the medium by a 200 - 1,000 µL pipette.
- Add new fresh medium by a 200 - 1,000 µL pipette.
6-7 days after cell seeding, add 20 ng/mL of RANKL to complete α-MEM+MCSF.
- Change the medium (complete α-MEM+MCSF+RANKL) every 2-3 days:
- Discard the medium with a 200 - 1,000 µL pipette.
- Add new medium with a 200 - 1,000 µL pipette. Stop induction of differentiation 14 days after cell seeding.
- Discard medium and wash twice with PBS.
- Fix cells with paraformaldehyde:
- Leave cells to dry.
- Add 4% paraformaldehyde (see Table of Materials).
- Incubate for 20 min at room temperature.
Discard paraformaldehyde and wash twice with 500 µL of PBS. CAUTION: Paraformaldehyde is toxic; please wear gloves, mask and eyeglasses.
Allow cells to dry.
Perform TRAP staining according to the manufacturer's instructions (see Table of Materials).
Count osteoclast-like cells manually under the microscope at magnification 10X; NA 0.30. NOTE: Osteoclast-like cells are defined as TRAP+ polycarion cells with at least 4 nuclei.
2. Cancer Cell Cultures
NOTE: The experiments can be performed with different types of breast adenocarcinoma cell lines such as SCP217, an osteotropic cell line originating from the triple negative human breast cancer cell line (MDA-MB-231), and the hormonal receptor-positive cell line MCF7.
- Culture the cells as a monolayer in 75-cm2 flasks.
- Seed 1,000,000 cancer cells in a 75-cm2 flask.
- Store the cells at 37 °C in complete α-MEM medium supplemented with 1% glutamine and 10% fetal bovine serum in a 5% CO2 atmosphere.
- Detach the cells at a confluency of about 90%:
- Discard the medium, wash with PBS, and add 2 - 3 mL of trypsin 1X in PBS.
- Incubate cells for 3-5 min at 37 °C.
- Collect the detached cells from the 75-cm2 flask by a 10 mL pipette, adding 8 mL of complete medium to stop the enzymatic reaction.
- Centrifuge at 225 x g for 5 min.
- Wash the cancer cells:
- Add 10 mL of PBS.
- Centrifuge at 600 x g for 5 min.
- Count cancer cells with a Neubauer chamber.
- Count cancer cells at least twice and calculate the mean of replicates to get the total mean number of cells collected.
3. Direct Co-Cultures of Cancer Cells a nd Monocytes Undergoing Differentiation towards Osteoclasts
- Seed the cancer cells on inserts (pore diameter 0.4 µm):
- Seed the cancer cells at a concentration of 4,000 cells per cm2.
- Incubate at 37 °C and 5% CO2. NOTE: Seed the cancer cells the day after PBMC seeding for the co-culture experiment in 0.4 µm inserts.
Wait for 24 h to allow the cells to adhere.
Place the seeded insert culture over the mononuclear cultures to start the co-culture (see section 1.10 of the protocol).
- Change the media every 2 - 3 days with complete α-MEM.
- Discard the medium with a 200 - 1,000 µL pipette.
- Add 500 µL of fresh medium. NOTE: The primary aim of the co-culture is to understand the osteoclastogenic power of cancer cells, therefore, the GFs are not added to α-MEM. In each experiment of co-culture, include negative controls (see note in step 1.10 of the protocol) and positive controls of osteoclasts differentiated by RANKL and MCSF (no cancer cells). Cancer cells seeded in the inserts not placed over mononuclear cells are used as a negative control of the co-culture.
- Stop the co-culture and use cancer cells for downstream analysis:
- 13 days after PBMC seeding and 12 days after cancer cell seeding, place the insert on a new plate.
- Stop cancer cell culture on the basis of which downstream analyses are to be performed (see Discussion).
On day 14, stop osteoclast differentiation and fix cells, as reported in step 1.15.
Perform TRAP staining immediately or within 14 - 30 days. NOTE: Optional staining: To confirm that the TRAP+ polycarion cells are osteoclasts, researchers can perform additional staining to detect CTR, an osteoclast-specific marker. The detection of F-actin rings is another osteoclast-specific staining assay12,13,14.
Representative Results
A method was optimized to easily differentiate osteoclasts from human peripheral blood monocytes. The monocyte culture was cultured with cancer cells, confirming (as described in the literature18) that cancer cells are capable of sustaining osteoclastogenesis in bone metastases. Osteoclasts differentiated from cancer cells and GFs are shown in Figure 1. An osteoclast-like cell is a cell with 4 or more nuclei and is positive for TRAP staining (purple cells). The accepted cut-off for the number of nuclei used to define an osteoclast is 318, but we decided to use 4 to reduce the risk of false positives to a minimum.
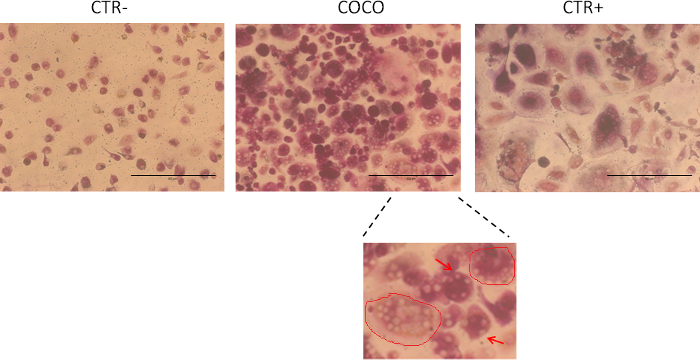
The model shows that breast cancer cells sustained osteoclastogenesis because the number of osteoclasts in the wells induced by cancer cells was similar to that obtained in the positive control (Figure 2). As shown in Figure 2A, a number of cells also differentiated spontaneously into osteoclasts in the negative control. In order to confirm the usefulness of the model, the system was challenged with an antitumor drug12,13. Osteoclast numbers were higher in co-culture conditions and positive control (CTR+) samples than in negative control (CTR-) samples. Osteoclast numbers significantly decreased in the presence of the drug (everolimus) in both co-culture conditions and positive controls (CTR+). The t-test was used for statistical analyses. Cells were counted in each well. The surface areaof the osteoclast is a characteristic of maturation and can be analyzed using open source software such as ImageJ. Osteoclasts derived from GFs were larger than those induced by cancer cells (Figure 2B). Treatment with an antitumor drug (everolimus) turned off the effects of GFs. The model provided useful information on the role of the drug in osteoclastogenesis inhibition. Figure 2C shows that osteoclastogenesis decreased in the presence of the drug. These results were obtained by co-culture-induced osteoclastogenesis. As explained in the protocol section, a number of other downstream analyses can be performed to investigate the mechanisms of the crosstalk and/or of the analyzed drugs. The model summarized in Figure 3 is highly versatileand can be used to study the effect of drugs on cancer cells, in particular, osteoclasts.
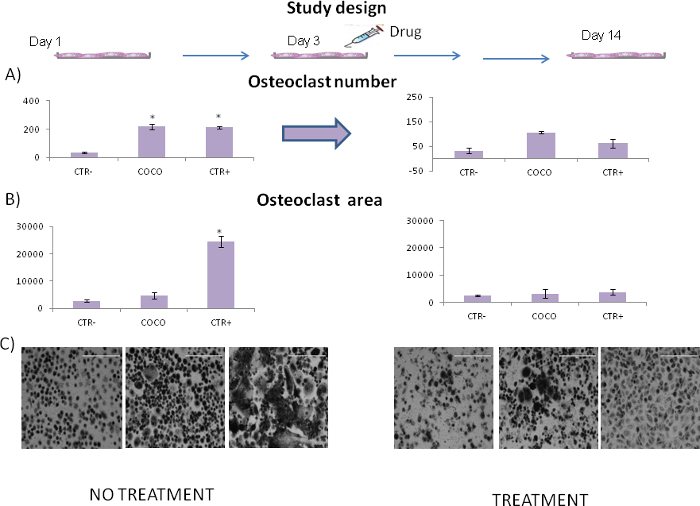
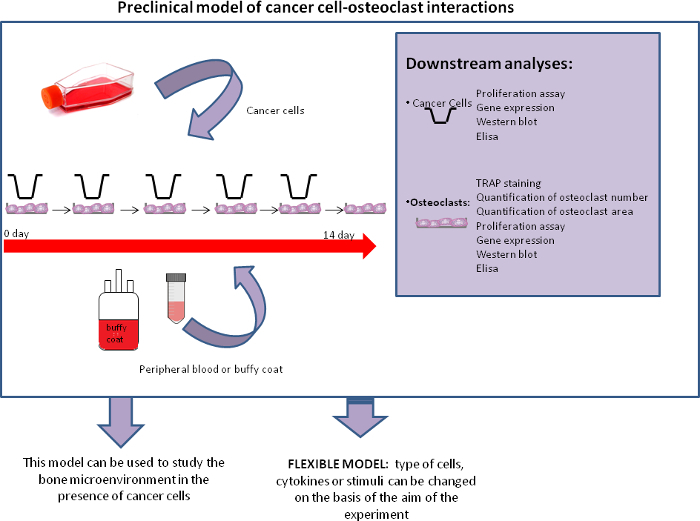
Figure 1. Osteoclastogenesis assay. Cells stained after 14 days in culture. CTR- monocytes cultured in the absence of growth factors; CTR+ monocytes cultured with RANKL and MCSF growth factors; COCO, osteoclasts obtained by co-culturing blood cells with cancer cells. 10X magnification. Osteoclast-like cells were cells with at least 4 nuclei that were positive to TRAP (purple cells). Cells were quantified by counting the cells in each well; 4 replicates for each condition were performed and the average was calculated. The scale bar is 400 µm. The image zoom in the lower panel (100X) illustrates the surface area of 2 osteoclasts (red circles). The arrows indicate osteoclast nuclei. Please click here to view a larger version of this figure.
Figure 2. Effect of everolimus on osteoclastogenesis treatment. (A) Number of osteoclasts according to the presence/absence of a drug; 4 replicates for each condition were performed. CTR- monocytes cultured in the absence of growth factors; CTR+ monocytes cultured with RANKL and MCSF growth factors; COCO, osteoclasts obtained by co-culturing blood cells with cancer cells. Data are expressed as mean ± SE.Significance was evaluated by the t-test; *p<0.05. (B) Osteoclast surface area was calculated by ImageJ open source software. At least 50 osteoclasts per condition were measured. CTR- monocytes cultured in the absence of growth factors; CTR+ monocytes cultured with RANKL and MCSF growth factors; COCO, osteoclasts obtained by co-culturing blood cells with cancer cells. (C) Images of the cells after 14 days in culture in the absence or presence of an antitumor drug (everolimus). 10X magnification; scale bar is 400 µm. Please click here to view a larger version of this figure.
Figure 3. Co-culture model. Scheme of the model and of its possible downstream analyses and future applications. Please click here to view a larger version of this figure.
Discussion
Preclinical in vitro models studying the mechanisms of crosstalk between cancer cells and bone cells are needed to identify mechanisms of bone metastasis that can be used to create new therapeutic strategies. We developed a fully human in vitro model of osteoclastogenesis from human peripheral blood (Figure 3). During optimization of the methodology, a number of critical points were identified and resolved. The first concerned the quantity of mononuclear cells to seed. The number of cells obtained by the microscope count is only a rough quantification as it is difficult to discriminate between lymphocytes and monocytes, the latter being the target cells to seed for the osteoclastogenesis experiment. We were able to identify monocytes because lymphocytes do not adhere to plastic substrate and thus the majority was eliminated at the first change of medium. The cells were seeded at a high density, as it is essential for the cells to be able to interact with each other for differentiation, especially in the second phase of osteoclastogenesis in which pre-osteoclasts need to be close together in order to fuse and become mature osteoclasts. A further difficulty arises when there are multiple experimental conditions because these require a high number of cells and a large amount of blood. One possible solution is to use buffy coats that have a high concentration of PBMCs, e.g., 4 x 105-6 x 106 mononuclear cells are normally collected from a 50 mL buffy coat sample compared to fewer than 1 x 105 PBMCs collected from a 20 mL blood sample.
As blood samples were collected from different volunteers, there was some degree of variability in the osteoclastogenesis assay. It was thus important to perform each assay with at least 3-4 technical replicates. Biological replicates were also needed and as different healthy donors were used, we did not normally calculate the mean of values obtained by the different experiments. We ensured that the trends of the different tests were equivalent and then chose one experiment from one of the healthy donors to be included in the manuscript.
Our preclinical model involves the co-culturing of monocytes undergoing differentiation with cancer cells. As we previously showed, cancer cells can also sustain in vitro osteoclastogenesis12,13. This system can be used to test drugs or to investigate mechanisms of interaction, as it is possible to perform several downstream analyses on both the cancer cells and osteoclasts. In fact, it is possible to perform proliferation assays on the cancer cells and gene expression analyses/Western blot assays on both cancer cells and osteoclasts13. It would be of interest to understand how secreted proteins differ in the different conditions. Our model has a number of advantages, i.e., it is a fully human model that is not dependent on murine cell lines, unlike several models reported in the literature. The model is also fairly robust and reproducible, and can be used for other purposes. For example, other cancer cell lines can be co-cultured with osteoclasts and the difference in their osteoclastogenic power studied. Moreover, this system can be used to evaluate other physiological or pathological mechanisms to improve our understanding of bone biology and osteoclast-mediated diseases, such as osteoporosis. Other cytokines and cell types can also be co-cultured together with osteoclasts depending on the aim of the experiment.
Several downstream analyses are possible with this model. It is important to decide during the design phase of the study what analyses will be performed so that an adequate number of wells can be seeded.
Downstream Assays with Osteoclasts:
(i) Gene expression analyses. Plan additional wells of osteoclasts to extract total RNA, as the wells stained for TRAP evaluation are not available for RNA extraction. (ii) Western blot analyses. (iii) Quantification of osteoclast surface area. Osteoclast size is a parameter for osteoclast maturation and can be quantified using open source software such as ImageJ.
Downstream Assays with Cancer Cells:
(i) Proliferation assays such as MTT 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide. (ii) Gene expression analyses. (iii) Western blot analyses.
Downstream Assays with Osteoclasts and Cancer Cells (Co-culture):
As cancer cells and osteoclasts share the same culture medium, their interaction can be studied with the ELISA assay which analyzesthe secretion of specific cytokines, GFs and other secreted proteins.
The development of this model was crucial for our project and we plan to further improve it in the near future. A major step forward would be to differentiate monocytes in 3D mineralized collagen scaffolds to mimic bone matrix. It would then be possible to directly evaluate osteoclast activity with a bone resorption assay. As we continue to learn more about the diverse roles played by osteoclasts in physiological and pathological processing ranging from bone remodeling to regulating bone metastases, our model will enable researchers to isolate and study osteoclasts in vitro.
Disclosures
The authors have no conflicts of interest to disclose.
Acknowledgments
We would like to thank Yibin Kang for providing the SCP2 cell line and Cristiano Verna for editorial assistance.
References
- Ibrahim T, Mercatali L, Amadori D. Bone and cancer: the osteoncology. Clin Cases Miner Bone Metab. 2013;10(2):121–123. [PMC free article] [PubMed] [Google Scholar]
- Coleman RE, Rubens RD. The clinical course of bone metastases from breast cancer. Br J Cancer. 1987;55(1):61–66. doi: 10.1038/bjc.1987.13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ibrahim T, Mercatali L, Amadori D. A new emergency in oncology: bone metastases in breast cancer patients. Oncol Lett. 2013;6(2):306–310. doi: 10.3892/ol.2013.1372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swartz MA, et al. Tumor microenvironment complexity: emerging roles in cancer therapy. Cancer Res. 2012;72(10):2473–2480. doi: 10.1158/0008-5472.CAN-12-0122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roodman GD. Mechanisms of bone metastasis. N Eng J Med. 2004;350(12):1655–1664. doi: 10.1056/NEJMra030831. [DOI] [PubMed] [Google Scholar]
- Patel LR, Camacho DF, Shiozawa Y, Pienta KJ, Taichman RS. Mechanisms of cancer cell metastasis to the bone: a multistep process. Future Oncol. 2011;7(11):1285–1297. doi: 10.2217/fon.11.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen YC, Sosnoski DM, Mastro AM. Breast cancer metastasis to the bone: mechanisms of bone loss. Breast Cancer Res. 2010;12(6):215. doi: 10.1186/bcr2781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guise TA. Breast cancer bone metastases: it's all about the neighborhood. Cell. 2013;154(5):957–959. doi: 10.1016/j.cell.2013.08.020. [DOI] [PubMed] [Google Scholar]
- Ell B, et al. Tumor-induced osteoclast miRNA changes as regulators and biomarkers of osteolytic bonemetastasis. Cancer Cell. 2013;24(4):542–556. doi: 10.1016/j.ccr.2013.09.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu X, et al. VCAM-1 promotes osteolytic expansion of indolent bone micrometastasis of breast cancer by engaging α4β1-positive osteoclast progenitors. Cancer Cell. 2011;20(6):701–714. doi: 10.1016/j.ccr.2011.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang H, et al. The osteogenic niche promotes early-stage bone colonization of disseminated breast cancer cells. Cancer Cell. 2015;27(2):193–210. doi: 10.1016/j.ccell.2014.11.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liverani C, et al. CSF-1 blockade impairs breast cancer osteoclastogenic potential in co-culture systems. Bone. 2014;66:214–222. doi: 10.1016/j.bone.2014.06.017. [DOI] [PubMed] [Google Scholar]
- Mercatali L, et al. The effect of everolimus in an in vitro model of triple negative breast cancer and osteoclasts. Int J Mol Sci. 2016;1(11):e1827. doi: 10.3390/ijms17111827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glantschnig H, Fisher JE, Wesolowski G, Rodan GA, Reszka AA. M-CSF, TNFalpha and RANK ligand promote osteoclast survival by signaling through mTOR/S6 kinase. Cell DeathDiffer. 2003;10(10):1165–1177. doi: 10.1038/sj.cdd.4401285. [DOI] [PubMed] [Google Scholar]
- Sugatani T, Hruska KA. Akt1/Akt2 and mammalian target of rapamycin/Bim play critical roles in osteoclast differentiation and survival, respectively, whereas Akt is dispensable for cell survival in isolated osteoclast precursors. J Biol Chem. 2005;280(5):3583–3589. doi: 10.1074/jbc.M410480200. [DOI] [PubMed] [Google Scholar]
- Bertoldo F, et al. Targeting bone metastatic cancer: role of the mTOR pathway. Biochim Biophys Acta. 2014;1845(2):248–254. doi: 10.1016/j.bbcan.2014.01.009. [DOI] [PubMed] [Google Scholar]
- Kang Y, Siegel PM, Shu W, Drobnjak M, Kakonen SM, Cordón-Cardo C, et al. A multigenic program mediating breast cancer metastasis to bone. Cancer Cell. 2003;3(6):537–549. doi: 10.1016/s1535-6108(03)00132-6. [DOI] [PubMed] [Google Scholar]
- Simone V, Ciavarella S, Brunetti O, Savonarola A, Cives M, Tucci M. Everolimus restrains the paracrine pro-osteoclast activity of breast cancer cells. BMC Cancer. 2015;14(15):692. doi: 10.1186/s12885-015-1717-8. [DOI] [PMC free article] [PubMed] [Google Scholar]