

Abstract
Angiogenesis is a complex, highly regulated process responsible for providing and maintaining adequate tissue perfusion. Insufficient vasculature maintenance and pathological malformations can result in severe ischemic diseases, while overly abundant vascular development is associated with cancer and inflammatory disorders. A promising form of pro-angiogenic therapy is the use of angiogenic cell sources, which can provide regulatory factors as well as physical support for newly developing vasculature.
Mesenchymal Stromal Cells (MSCs) are extensively investigated candidates for vascular regeneration due to their paracrine effects and their ability to detect and home to ischemic or inflamed tissues. In particular, first trimester human umbilical cord perivascular cells (FTM HUCPVCs) are a highly promising candidate due to their pericyte-like properties, high proliferative and multilineage potential, immune-privileged properties, and robust paracrine profile. To effectively evaluate potentially angiogenic regenerative cells, it is a requisite to test them in reliable and "translatable" pre-clinical assays. The aortic ring assay is an ex vivo angiogenesis model that allows for easy quantification of tubular endothelial structures, provides accessory supportive cells and extracellular matrix (ECM) from the host, excludes inflammatory components, and is fast and inexpensive to set up. This is advantageous when compared to in vivo models (e.g., corneal assay, Matrigel plug assay); the aortic ring assay can track the administered cells and observe intercellular interactions while avoiding xeno-immune rejection.
We present a protocol for a novel application of the aortic ring assay, which includes human MSCs in co-cultures with developing rat aortic endothelial networks. This assay allows for the analysis of the MSC contribution to tube formation and development through physical pericyte-like interactions and of their potency for actively migrating to sites of angiogenesis, and for evaluating their ability to perform and mediate ECM processing. This protocol provides further information on changes in MSC phenotype and gene expression following co-culture.
Keywords: Developmental Biology, Issue 127, Angiogenesis, aortic ring assay, cell migration, endothelial networks, mesenchymal stromal cells, physical interactions, paracrine factors, perivascular cells, umbilical cord
Introduction
The complex process of angiogenesis improves and maintains tissue perfusion by promoting new blood vessel development from pre-existing vasculature1. It is a tightly regulated, balanced process by pro-angiogenic and anti-angiogenic factors. Any deficiency in this system may lead to insufficient vessel maintenance or growth, causing severe ischemic diseases including myocardial disease, stroke, and neurodegenerative disorders. However, exaggerated vascular development is characteristic for conditions including cancer and inflammatory disorders2.
Developing therapies that aim to control angiogenesis to achieve favorable tissue regeneration is of key importance. Despite extensive preclinical and clinical investigations, attempts to stimulate angiogenesis using pro-angiogenic factors and microRNAs have failed to achieve desired outcomes3,4,5. Possible reasons for the transient effects include: limited longevity of angiogenic proteins and nucleic acids, and the finite number of targeted growth factors6,7. Although soluble angiogenic factors are essential for initiating angiogenesis, the maintenance and functionality of vasculature depend on supporting cell types including pericytes and smooth muscle cells8. The field of pro-angiogenic therapies is now exploring potential stem cell and progenitor cell sources that may provide angiogenic factors locally, while physically supporting newly developed vasculature, self-renewing or even differentiating into endothelial-like cells9,10. Finding the optimal angiogenic cell types with the capability to fulfill these functional requirements holds a great promise for ischemic tissue regeneration.
In order to successfully translate potential cell-based therapies into clinical trials, pre-clinical studies need to demonstrate their efficacy and highlight the underlying angiogenic mechanisms. Despite the high number of established angiogenesis assays, the field lacks a "gold-standard" in vitro assay that could reliably evaluate the efficacy of potential candidate cell types11,12,13. Most in vitro angiogenesis assays (including the endothelial proliferation, migration and tube formation assays) typically assess the effects of cells or compounds on endothelial cells' phenotypical changes or differentiation into tubular and network structures14,15. While these features are critical for angiogenesis, a "translatable" assay should also evaluate: 1) the augmentation or replacement of the supporting cell types including pericytes or smooth muscle cells, 2) the processing of ECM and/or basement membrane, and 3) the efficiency to promote the formation of functional microvasculature. In vivo angiogenesis models, including the corneal assay and Matrigel plug assay, recapitulate the unique in vivo microenvironment but are challenged by the difficulty of tracking administered cells to observe physical interactions. Furthermore, in in vivo models, xeno-immune rejection can occur while testing potential allogeneic cell therapy candidates16. Ex vivo angiogenesis models, particularly the aortic ring assay can provide: 1) easy observation and quantification of tubular structures, 2) accessory supportive cells, 3) ECM from host and artificial supplies, 4) exclusion of inflammatory components, and 5) quick and inexpensive setup17,18. Typically, the aortic ring assay can test the angiogenic potential of small secretory proteins, pharmacological agents, and transgenic rodent models19,20,21.
MSCs are promising candidates for vascular regeneration primarily through their paracrine-mediated effects22,23,24. MSCs have been shown to secrete key angiogenic factors including Vascular Endothelial Growth Factor (VEGF), Hepatocyte Growth Factor (HGF), Insulin-like Growth Factor-1 (IGF-1), basic Fibroblast Growth Factor (bFGF), and angiopoeitin-1 (Ang-1)25,26. MSCs can also detect and home to ischemic or inflamed tissues, however, the exact mechanisms are still under investigation. Increasingly, the literature supports the hypothesis that most MSCs arise from perivascular cells, co-express pericyte markers, and can behave like pericytes27. HUCPVCs are a young source of MSCs derived from the perivascular region of the human umbilical cord. They represent a population of MSCs with pericyte-like properties and have been characterized from both FTM and term umbilical cords. FTM HUCPVCs demonstrate a high expression of pericyte markers including CD146 and NG2, high proliferative and multilineage potential, immune-privileged properties, and display a robust paracrine profile28. FTM HUCPVCs are an ideal candidate cell type to promote regeneration of injured tissue through the promotion of new vasculature via their pericyte-like properties.
To test the angiogenic potential and pericyte-like properties of human MSCs, a very limited number of angiogenesis assays are available where positive angiotropic migration (hereafter referred to as "homing"), ECM processing, and development of physical interactions between cell types can be investigated, while obtaining quantitative data on microvasculature development.
Hereby we present a protocol that describes a novel application of the aortic ring assay. Human MSCs were co-cultured with developing rat-derived aortic endothelial networks to assess their contribution to tube formation, maturation, and homeostasis. This version of the aortic ring assay assesses the ability and potency of cell therapy candidates to home to sites of angiogenesis, perform and mediate ECM processing, and contribute to endothelial tubular development through establishing pericyte-like physical interactions. In addition to quantifying the net effect of MSCs on in vitro endothelial network formation and observing intercellular interactions, we also optimized a protocol to isolate MSCs from co-cultures. By performing flow cytometry and qPCR, it is possible to characterize changes in MSC phenotype and gene expression following co-culture. As model cell types, we compared ontogenetically early (prenatal) and late (adult) sources of human MSCs: FTM HUCPVCs and human bone marrow-derived MSCs (BMSC), respectively, in the aortic ring assay. We propose that the aortic ring assay can be used to study the angiogenic potential of any physically supporting cell type when under investigation for angiogenic regenerative applications.
Protocol
All studies involving animals were conducted and reported according to ARRIVE guidelines29. All studies were performed with institutional research ethics board approval (REB number 4276). All animal procedures were approved by the Animal Care Committee of the University Health Network (Toronto, Canada), and all animals received humane care in compliance with the Guide for the Care and Use of Laboratory Animals, 8th edition (National institutes of Health 2011).
1. Aortic Ring Assay Setup
- Isolate the rat aorta.
- Euthanize 8 - 10-week-old Sprague-Dawley rats using CO2 asphyxiation: Set CO2 chambers to 20% gas replacement (flow rate = 0.2 x chamber volume/min). Confirm by the absence of pinch reflex and heart beats by palpating chest.
- Wet the fur on chest area using sterilized gauze with 70% ethanol. Use sterilized forceps and scissors to make an incision in the chest midline and cut through skin and muscle layers.
- Once the rib cage is exposed, cut the rib cage on each side and fold upwards about 5 mm from the sternum on each side. Some bleeding is expected from intercostal arteries in fresh cadavers.
- Push the lung tissue and heart to the anatomical right side to expose the aorta. The aorta is the white colored vessel located adjacent to the spinal column.
- Use forceps to pinch the aorta (after the aortic arch) and use surgical scissors to make the first incision while holding the aorta with forceps. Upon cutting the aorta, the chest cavity will quickly fill with blood, therefore it is critical to work quickly to obtain the aorta prior to blood coagulation.
- Use forceps to hold the aorta and gently peel the aorta downwards, away from the spinal column. The pull will sever the intercostal arteries without further incisions. Obtain approximately 10 cm of tissue and/or before the aorta divides into two branches in the abdominal cavity and cut the aorta out of the carcass. Push the diaphragm if needed to the caudal direction to gain access to lower sections of the aorta. NOTE: Peel the aorta gently because intense force can cause it to tear. If the aorta tears, allow blood coagulation for a few seconds and use a paper towel to remove the coagulated blood, allowing visibility of the remaining aorta.
- Place the aorta in a 15 mL tube with 10 mL of ice-cold Hank's Balanced Salt Solution (HBSS) supplemented with 1% penicillin/streptomycin (P/S). Keep the tubes on ice. NOTE: The aortic tissue can last for 1-2 h on ice but it is best to process it as quickly as possible.
- Prepare the aortic ring assay culture media in a sterile biosafety cabinet (BSC).
- Prepare the Endothelial Growth Medium (EGM) by using a filtration unit to filter 500 mL of Endothelial Basal Medium (EBM) supplemented with 2% Fetal Bovine Serum (FBS), 1% gentamicin and growth factors (see Table of Materials). Place 50 mL of the filtered media into a bead or water bath at 37 °C.
- Use another filtration unit to filter 100 mL of EBM supplemented with 2% FBS and 1% P/S to prepared the EBM 2% FBS (EBM-FBS) and store at 4 °C.
- Coat 12-well tissue culture plates with Basal Membrane Extract (BME) while working on ice.
- Place a 12-well plate on a cold surface (large ice pack or tray). Add 200 µL/well of freshly thawed BME using refrigerated pipette tips and swirl the BME to ensure even coating. Work quickly to avoid uneven polymerization of the BME. NOTE: A typical aorta excision yields 20 aortic rings. To account for variability between the aortic ring assays, setup at least three aortic ring assays per treatment group and include a control. If the source permits, 6 rings per treatment group is recommended.
- Place the coated plates into humidified incubators (95% relative humidity, 37 °C, 5% CO2; these conditions apply to all humidified incubator steps hereafter) for 30 min. Place 1,000 µL pipette tips back into -20 °C for step 1.5.3. NOTE: While the basal membrane extract is used in stock concentration, its consistency and stiffness is strictly controlled by the polymerization time. Make sure to keep the exact same polymerization times for all parallels and independent experiments.
- Section the aorta into uniform rings.
- Take the aorta from the 15-mL tube and place it into a 10 cm dish with 10 mL of fresh HBSS supplemented with 1% P/S.
- Use two sets of forceps to carefully separate the excess connective tissue, adipose tissue, and use a scalpel for the remaining branches of the intercostal arteries connected to the aorta. This reduces variability between ring units based on differing levels of the residual tissue. Remove any residual coagulated blood from inside of the aorta.
- Place a ruler or grid under the 10 cm dish to precisely cut 1 - 2 mm wide sections of the aorta using sterile scalpel and forceps. Cut the sections as exact as possible to reduce variability between units.
- Embed the aortic rings into BME. NOTE: If the sectioning of aortic rings is not completed before BME polymerization incubation, remove the coated plates from humidified incubators and add 100 µL of EBM to each well to terminate the polymerization. Exact timing is critical as over-polymerization of BME may interfere with the endothelial network development and cell migration. Once the aortic rings are prepared, remove the EBM from the wells and continue from step 1.5.2.
- Remove the BME coated wells from the humidified incubators and return to the BSC.
- Carefully pick up individual aortic rings with forceps and place one in the middle of each BME-coated well. Repeat for the remaining aortic rings. NOTE: Media transferred along with the aortic ring should be kept minimal in order to avoid polymerization irregularities.
- Use cooled (-20 °C) 1,000 µL pipette tips to add 300 µL of BME on top of each aortic ring and evenly distribute the BME around the well.
- Return the embedded aortic rings to the humidified incubators for 30 min.
- Remove the plates with the embedded aortic ring from the humidified incubators and slowly add 1,000 µL of EGM (prepared and warmed at step 1.2.1) by placing the pipette tip against the wall of the culture well. NOTE: Directly pipetting the media onto the BME can disrupt the polymerized BME and hinder continuous endothelial network development. Use an appropriate multichannel pipette if available.
- Following 24 h, remove 1,000 µL of EGM and replace with 1,000 µL of EBM-FBS prepared in step 1.2.2, again by slowly pipetting against the side of the culture well.
Maintain the aortic ring assay by replacing 500 µL of EBM-FBS with 500 µL of fresh EBM-FBS (37 °C) every 48 h. NOTE: See section 2 to start the MSC culture expansion on the same day as aortic ring assay establishment. This will ensure that MSCs are ready for co-culture when the endothelial networks are ready.
Use the bright field microscopy to follow aortic ring assay endothelial network development (see Figure 1 as reference for optimal network development). Once the endothelial networks aspect is as shown in Figure 2, proceed with setting up the aortic ring assay-MSC co-cultures (section 3). Refer to step 4.1 for further details of imaging the endothelial networks.
2. Tissue Culture
- Prepare stock solutions of tissue culture media.
- Culture FTM HUCPVCs (previously established, n ≥ 3 independent lines for each)30 and commercially available BMSCs in alpha-MEM supplemented with 10% FBS and 1% P/S.
- Sterilize the media using 0.2 µm pore size filter bottles.
- Store the prepared media solutions at 4 °C for up to 3 weeks.
Maintain FTM HUCPVC and BMSC cultures in humidified incubators and passage at 70 - 80% confluency determined by phase contrast microscopy. Use appropriate volumes of media for the size of tissue culture dish used (i.e. 10 mL in a 10 cm dish). Use these culture conditions for maintaining and passaging the MSCs.
Dissociate the MSC monolayers for passaging or aortic ring assay-MSC co-cultures using a dissociation enzyme solution (4 mL/10 cm dish) and incubate in the humidified incubators for 3 min. Ensure detachment of the cells using bright field microscopy; incubate for an additional 1 - 2 min if required.
Transfer the dissociated cells into a 15 mL tube and centrifuge at 400 x g for 5 min.
Aspirate the supernatant without disturbing the cell pellet and resuspend the cells in 1 mL of an appropriate culture media (alpha-MEM complete media for passaging cells or EBM-FBS for aortic ring assay/MSC co-cultures) for counting using a cell counter.
3. Preparation of Aortic Ring Assay/MSC Co-cultures
- Seed 104 MSCs on each embedded aortic ring (9,000 cells/cm2/12-well plate). Based on the number of aortic ring assays, pre-stain the MSCs with viable, nontransferable fluorescent dye. Maintain at least three wells of aortic rings without MSCs for a control group.
- To stain 105 MSCs/mL EBM-FBS media, add 2.5 µL of viable, non-transferable fluorescent dye/mL media (5 µM) into a 15 mL tube and place in the humidified incubator for 30 min. Gently mix the tube halfway through the incubation.
- Following the 30 min incubation, add 1 volume of fresh EBM-FBS to the stained cells and spin at 400 x g for 5 min.
- Remove the supernatant and re-suspend the cell pellet in 1 mL of EBM-FBS media. Confirm successful staining of the MSCs using fluorescence microscopy.
Remove the aortic ring-containing plates from the humidified incubator and remove 500 µL of EBM-FBS from each well.
- Carefully seed 104 MSCs in 500 µL of EBM-FBS on each embedded aortic ring by evenly pipetting around the endothelial networks. Ensure even distribution by gently shaking the culture plate.
- Add additional EBM-FBS to ensure that the total volume of media on the aortic ring assay/MSC co-cultures is 1,000 µL.
Use fluorescence microscopy to visualize the fluorescently labelled MSCs in the aortic ring assay.
4. Microscopy
- Use bright-field microscopy to image the rat endothelial networks prior to the aortic ring assay/MSC co-cultures for baseline (day 0) measurements of endothelial network properties (e.g., network length, network loops). Image 4 quadrants per well from the aortic ring to the furthest section of the endothelial network within that quadrant.
- Image the aortic ring networks following the aortic ring assay/MSC co-cultures at days 3, 5 and 7. Use these images to quantify the MSC effect on the endothelial network development.
- Use fluorescence microscopy to image pre-labelled MSCs in the aortic ring assay.
- Following 24 h of aortic ring assay/MSC co-cultures, use fluorescence microscopy to visualize MSC homing, elongation and integration with the endothelial networks. Overlap the fluorescent images of MSCs with the bright field images of endothelial cells to observe the colocalization of both cell types.
- Image the MSCs localized within the developed endothelial networks proximal to the aortic ring tissue, and the MSCs localized within newly developing networks distal to the aortic ring tissue (Figure 2).
5. Flow Cytometry and qPCR
One day prior to dissociating the aortic ring endothelial networks, thaw frozen dispase at 4 °C O/N. Section 5.3 is identical for both flow cytometry and qPCR.
Warm 50 mL of dispase and 50 mL of 0.5% trypsin in 37 °C bead or water bath.
- Retrieve the cells for analysis from the aortic ring assay/MSC co-cultures.
- After 1 week of aortic ring assay/MSC co-culture, remove the culture media and add 1 mL of Phosphate-Buffered Saline (PBS) slowly to each well for 3 min and remove. Repeat this wash twice more.
- Add 800 µL of pre-warmed dispase to each well and incubate in the humidified incubators for 15 min.
- Remove the plates from the humidified incubators and suspend dispase by pipetting (5 - 10x) to break up the BME, and transfer the contents of each well into separate 15 mL tubes.
- Repeat step 5.3.3 again with fresh pre-warmed dispase until no residual BME is observed in the culture well. Add 1 volume of PBS supplemented with 3% FBS to the dispase-cell suspension to inactivate the dispase. Remove the aortic rings floating in the cell suspension using pipette tips.
- Spin the cell suspensions at 400 x g for 5 min. Remove the supernatant carefully, leaving 3 mL of cell suspension in the 15 mL tube. NOTE: An obvious, solid pellet may not be visible but cells and BME are present.
- Add 3 mL of prewarmed 0.5% trypsin to the cell suspension and vigorously resuspend (5 - 10x by pipetting) cells. Incubate the trypsinized cell solutions in the humidified incubators for 10 min.
- Remove the trypsinized cell suspensions from humidified incubators and add 6 mL of PBS supplemented with 3% FBS and resuspend a few times. Spin the cell suspensions at 400 x g for 5 min.
- Carefully remove all the supernatant, leaving the cell pellet in the tube and resuspend cells in 1 mL PBS supplemented with 3% FBS in each tube.
- Filter the 1 mL cell suspension using a 70 µm cell strainer to remove aggregated cells and residual BME from the cell suspension. Count the cells in 1 mL of cell suspension using a cell counter.
- Prepare the cells for flow cytometry.
- Incubate the cell suspensions (105 cells in 200 µL PBS supplemented with 3% FBS) with fluorophore-conjugated (FITC or APC) primary antibodies (TRA-1-85-, CD146, CD31) concentration = 1:40) at 4 °C for 30 min, protected from light. NOTE: Flow cytometry for MSCs was optimized by Hong et al., 201328. A minimum of 10,000 cells is required for accurate readings.
- Following the 30 min incubation, resuspend the cells in 2 mL of PBS supplemented with 3% FBS and centrifuge (400 x g, 5 min).
- Keep cells at 4 °C protected from light until analyzing by flow cytometry (at least 1 x 104 events) within 1 h. For the gating strategy, see Supplementary Figure 1. Sort human MSCs using anti-TRA-1-85 (APC) (concentration: 1:40 and in 0.5% FBS/PBS) and sort cells into 350 µL of cell lysis buffer for qPCR analysis. NOTE: Lysed cells can be stored at -80 °C for up to 3 months for future qPCR analysis.
- Prepare the cells for qPCR analysis.
- Isolate the RNA from lysed cells using column-based RNA isolation kits and determine the RNA concentration and quality.
- Prepare cDNA from up to 5 µg of RNA per 100 µL reverse transcriptase reaction. Perform a pre-amplification step if the RNA yield is low (< 10 ng/µL). Use of less than 100 ng of RNA will result in a high rate of false negatives. NOTE: The cDNA templates can be stored at -20 °C for further analysis for up to 4 months.
- Perform the qPCR using angiogenesis expression profiler arrays to detect changes in gene expression. Use 5 ng of cDNA per reaction (40 cycles, 60 °C annealing/extending temperature). Express fold change of expression compared to undifferentiated MSC derived cDNA samples. Run the appropriate negative controls (aortic ring without human MSCs).
6. Network Quantification Following Aortic Ring Assay/MSC Co-cultures
Download imaging software such as ImageJ along with Angiogenesis Analyzer plugin31.
Import the endothelial network images taken and open using the software.
Convert the image scales based on the specifications of the microscope used.
Use a straight-line tool to measure the radial network growth.
Count the endothelial network loops using the cell counter tool (loops with at least 4 sides should be quantified).
For additional quantification of network properties, use the Angiogenesis Analyzer plugin or other similar plugins to analyze master segments, total segment length, total network areas, and number of junctions. Use the blurred mask tool to blur aortic ring tissue and empty spaces to avoid false positive calculations.
Transfer the values to a statistics program and graph endothelial network properties following the aortic ring assay/MSC co-cultures.
Representative Results
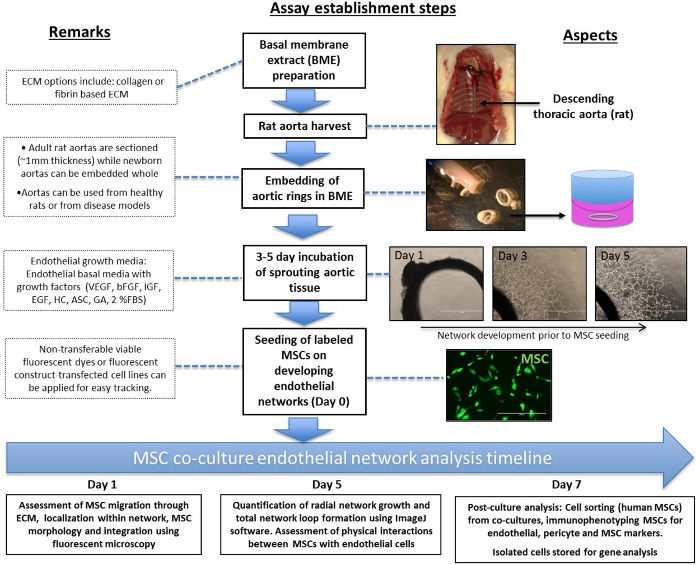
The schematic work-flow for establishing the aortic ring/MSC co-culture assay is demonstrated in Figure 1. The main steps include: rat aorta isolation, sectioning and embedding of the aortic rings, monitoring the endothelial sprouting and network development, and finally labelling and administering the MSCs. The timeline of the endothelial network analysis outlines the window for the analyses feasible for each period of co-culturing: day 1, 5 & 7. Additional notes are highlighted by dotted boxes.
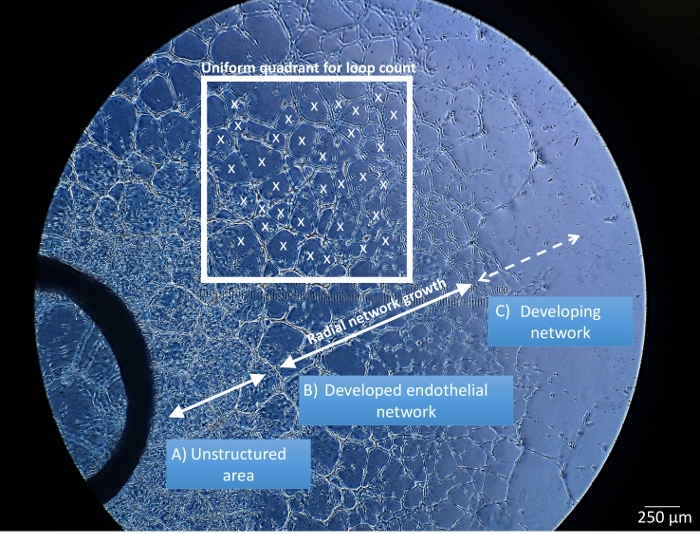
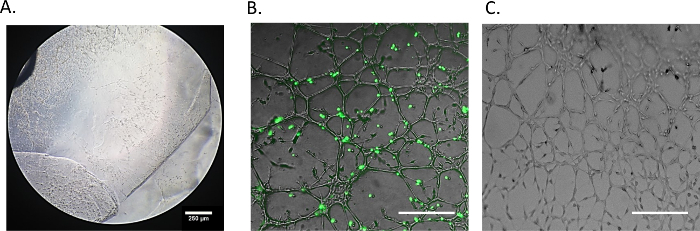
The identification of structurally distinct regions in the aortic ring endothelial cell cultures is performed by bright field microscopy, 3 - 5 d after the aortic rings are embedded in ECM (Figure 2). The unstructured area (Figure 2A) is characterized as a region of high cell proliferation but low structural organization in the direct vicinity of the aortic tissue. The developed endothelial networks (Figure 2B) refer to regions with high structural organization, where networks are fully established and predominantly composed of closed loops, even prior to the addition of MSCs. The developing networks (Figure 2C) are located at the distal areas of the endothelial cultures. These are the sites of endothelial cell migration, and elongation as the new network structures form. The above mentioned 3 regions are easily identifiable throughout the experiments and are used when referencing where the MSCs home to in the co-cultures. The prelabelled MSCs should be co-cultured with the aortic ring assay when all three network segments have developed. The quantification of endothelial networks following MSC co-cultures include regions of developed and developing network (Figure 2, white frame).
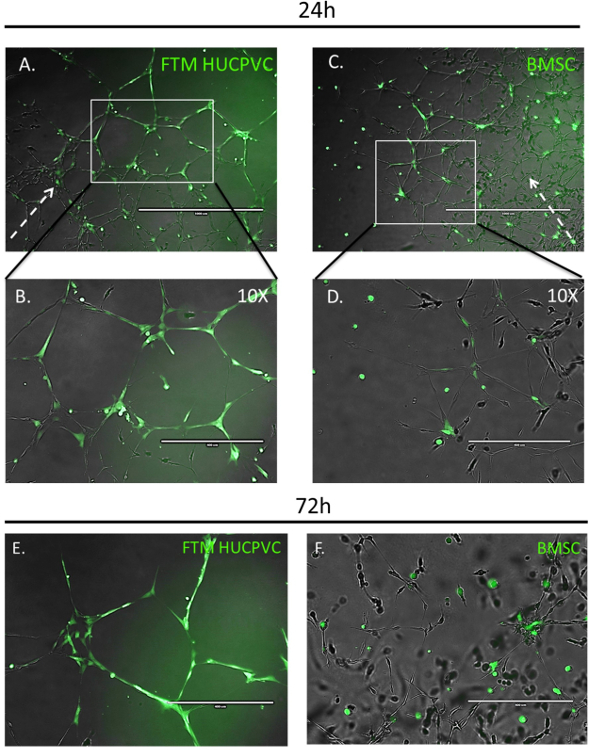
Fluorescence microscopy allows for qualitative measurements of MSC migration, integration and morphology in conjunction with developing endothelial networks in the aortic ring-MSC co-culture assay (Figure 3). The MSCs labelled with viable, cytoplasmic fluorophore were imaged 24 h after administration to co-cultures. FTM HUCPVCs were found at the periphery of the developing endothelial networks, displayed elongated cell morphologies, and contributed to the further development of endothelial networks (Figure 3A, B). For comparison, BMSCs homed to the developing networks while displaying less interaction with endothelial cells (Figure 3C, D). High magnification images at 72 h showed that FTM HUCPVCs maintained high coverage and stabilization of endothelial networks while BMSCs in co-cultures presented with spherical morphologies with limited integration into endothelial networks (Figure 3E, F). The observed differences clearly demonstrate the qualitative, functional differences between human MSC types. A therapeutic candidate cell type that has significant homing and endothelial integration properties (Figure 3A, B & E) stands out when compared to a cell type with limited endothelial network integration and augmentation capabilities (Figure 3C, D & F).
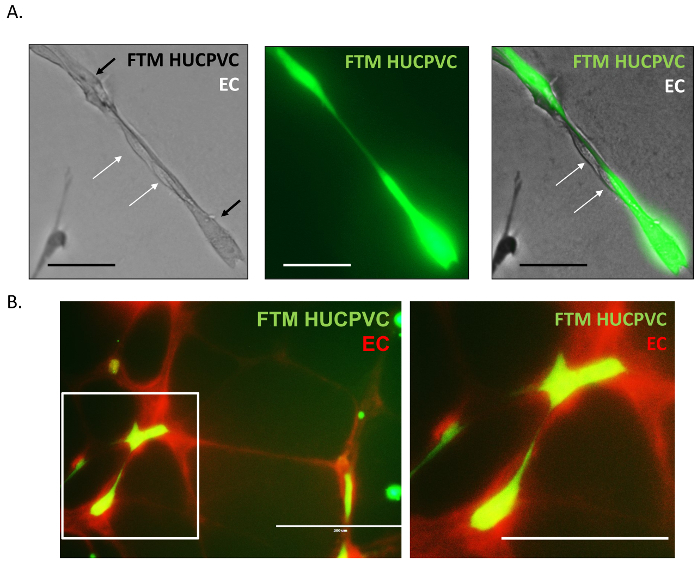
High magnification fluorescence microscopy images were acquired to further dissect physical interactions between endothelial cells and FTM HUCPVCs in tubular networks. The pre-labelled FTM HUCPVCs (Figure 4A, green) are shown adhering with unstained endothelial cells (Figure 4A,white arrows). FTM HUCPVCs were found to provide structural support to the endothelial cells by serving as an axis and attachment surface between nodes. The aortic ring endothelial networks can also be pre-labelled using a viable fluorophore similarly to the MSC staining (Figure 4B, red). In double stained co-cultures, fluorescence microscopy revealed elongated adhesions between the two cell types, fortifying the observation of cooperation and supportive cell behavior.
The quantification of endothelial network structural properties was performed at day 5 of MSC co-cultures (Figure 5). Uniform quadrants were defined for measurement throughout the entire aortic ring endothelial network (Figure 2, white frame). The mean network growth was calculated as the distance from the first proximal closed network loops by aortic ring to the furthest distal closed loops (radius size) (Figure 5A). The mean network growth of aortic ring-MSC co-cultures were as follows: FTM HUCPVCs (2.98 ±0.3 mm) and BMSCs (1.5 ±0.15 mm). The untreated endothelial networks developed a mean radius of 1.9 ±0.1mm. The statistical comparison between the MSC treatment groups demonstrated that FTM HUCPVCs contributed to significantly greater network growth when compared to BMSCs and untreated networks (p ≤0.001). The untreated endothelial networks developed significantly more than BMSC containing co-cultures (p ≤0.01) (Figure 5B).
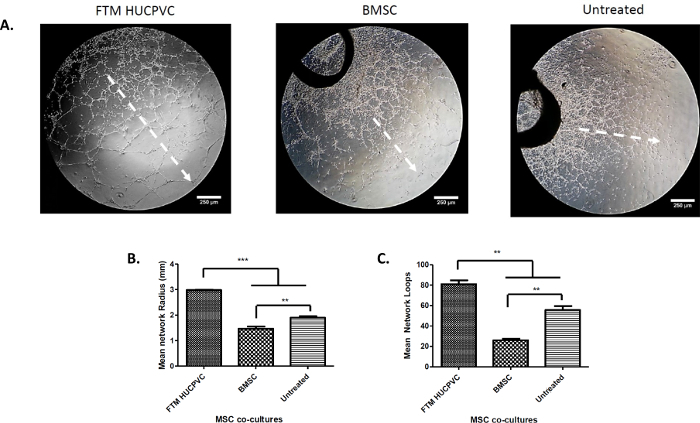
The total numbers of loops were calculated in each quadrant and expressed as average total loop formation (Figure 5C). The average number of total closed loops were as follows: FTM HUCPVC treated co-cultures (81 ±7), BMSCs (26 ±3) and untreated rings (55 ±7). FTM HUCPVCs contributed to significantly greater endothelial loop formation when compared to BMSCs (p ≤0.01) and untreated networks (p ≤0.01). The BMSC co-cultures resulted in fewer endothelial network loops when compared to untreated networks (p ≤0.01). This quantification identifies a cell type that has a significantly positive effect on endothelial network development (FTM HUCPVC) and a cell type that may even impair endothelial networks. It is to be noted that the significantly increased network development was observed to correlate well with the MSC coverage on the endothelial networks. It suggests that the direct interactions are crucial for the MSCs to execute their supportive function.
Possible pitfalls of the aortic ring co-culture establishments are represented in Figure 6. Inadequate or uneven polymerization of BME is a problem that can interfere with successful endothelial network development. Timing the BME polymerization for exactly 30 minutes and transferring the media without disrupting the polymer will ensure an intact, functional BME phase (Figure 6A). Staining the MSCs for prolonged incubation times and allowing the MSCs to pellet for extended periods may interfere with cell morphology and phenotype on co-cultures. Figure 5B shows FTM HUCPVCs stained for 1 h and allowed to pellet for 1 h prior to co-culture, both being sub-optimal timings. FTM HUCPVCs do not show preference to sites of endothelial networks and are scattered throughout the network. Comparing properly treated cells (Figure 3) to mishandled FTM HUCPVCs, the mishandled cells display rounded cell morphology and limited cell-to-cell interactions with the endothelial cells (Figure 6B). Lastly, if the aortic ring assay cannot be established on the day of the dissection, the aortas can be stored at -80 °C in EGM-C media supplemented with 10% dimethyl sulfoxide (DMSO) and 10% FBS. The endothelial network development may take 1 - 3 d longer when applying thawed aortic rings compared to fresh tissue, but the endothelial networks will be sufficient for MSC co-culture establishment (Figure 6C).
As an alternative proceeding of the aortic ring co-culture assessment, the cellular fraction of BME embedded aortic rings can be extracted and analyzed by flow cytometry (Supplementary Figure 1). The human specific marker TRA-1-85 (Supplementary Figure 1A, B, y axis) positive cell population from FTM HUCPVC containing co-cultures was identified as human MSCs. The co-labelling showed low expression of the endothelial cell marker CD31 (Supplementary Figure 1A) and high expression of the pericyte marker CD146 (Supplementary Figure 1B) within the human specific marker positive cell population. Such evaluation can decipher eventual immunophenotypic and lineage commitment changes of the MSCs induced by the interactions with endothelial cells, and provide further valuable information about the tested cell type. In order to obtain a sufficient number of cells for FC analysis, the cells extracted from parallel wells of the same experimental group can be combined. It is important to consider using non-pre-stained MSC containing the aortic rings for flow cytometry analysis so that the reminiscent fluorescence of the viable dye does not interfere with the signal of the antibody-related fluorophores. Otherwise the negative gating should take the pre-stained cell fluorescence into consideration.
We sorted human MSCs extracted form aortic ring co-cultures (day 7 of co-culture) using the human cell surface marker specific antibody (TRA-1-85). FTM HUCPVCs and BMSCs recovered from the aortic ring assay were processed for qPCR analysis to test the expression of angiogenic genes. Using a commercially available qPCR array, three aortic ring co-cultures provided sufficient amounts of genetic material to quantify the expression of 84 human growth factor genes (Supplementary Figure 2A). The preliminary results indicate that FTM HUCPVCs and BMSCs express key secreted angiogenic factors at comparable levels (Supplementary Figure 2B). Besides comparing expression levels of different cell types in the assay, the effect of transferring cells to EBM based assay media from their expansion culture media should be considered. When using cells of higher phenotypic or genetic plasticity, a comparator control group of human MSCs in EBM media - without aortic tissue - should be included in the evaluation.
The similarities in angiogenic factor expression of MSCs suggests that the significant difference in their network augmentation is not a result of different paracrine activities. This is in concordance with the earlier observation that increased direct intercellular interaction appears to promote endothelial network development.
Figure 1: Schematic Diagram of a Novel Application of the Aortic Ring Assay Setup. The main steps of setup and analysis of MSC co-cultures with the aortic ring assay are outlined in solid boxes and additional notes are outlined in dotted boxes. Scale bar = 1,000 µm. Please click here to view a larger version of this figure.
Figure 2: Representative Image of the Aortic Ring Network Analysis. Endothelial networks are divided into three concentric regions based on structural differences: unstructured area in close proximity to aortic ring tissue (A), developed/structured endothelial networks (B), and developing networks located in the periphery of the ex vivo tissue culture (C). Radial network growth and uniform quadrant for the loop count are defined within the developed endothelial network (B). x: closed endothelial loop counted in uniform quadrant. Scale bar = 250 µm. Please click here to view a larger version of this figure.
Figure 3: Fluorescent Imaging of Network Region-dependent Integration of Human MSCs in the Aortic Ring Assay following 24 & 72 h. Prestained (green) FTM HUCPVCs and BMSCs were added to the developing aortic ring endothelial tube networks. Fluorescence microscopy images taken 24 h after establishing MSC co-cultures display FTM HUCPVCs that migrate through the ECM and home to peripheral developing endothelial networks (A). Higher magnification images display elongated morphologies of FTM HUCPVCs while in close contact with the endothelial networks (B). Fewer BMSCs process the ECM and home to endothelial networks with no observable preference to peripheral developing networks (C). BMSCs display spherical cell morphologies (D). High magnification fluorescence microscopy images of prestained MSCs in rat aortic ring assay following 72 h of co-culture (E, F). FTM HUCPVCS present elongated morphologies while displaying endothelial coverage through direct cell-to-cell interactions with endothelial cells (solid white arrows) both in network nodes and tubules (E). BMSCs maintain spherical cell morphologies clustered in the endothelial network nodes (F). Broken arrow represents the direction of the endothelial network growth from the aortic ring tissue. A, C: Scale bar = 1,000 µm B, D: Scale bar = 400 µm. Please click here to view a larger version of this figure.
Figure 4: High Magnification Fluorescence Microscopy Images of Pericyte-endothelial Cell Physical Interactions. Unstained endothelial cells were found associated to continuous protrusions connecting pre-stained FTM HUCPVCs in the tubular network (A). Fluorescent images of pre-stained endothelial networks (EC, red) with pre-stained FTM HUCPVCs (FTM, green) demonstrate interactions recapitulating endothelial cell, pericyte interactions (B). A: Scale bar = 100 µm B: Scale bar = 200 µm. Please click here to view a larger version of this figure.
Figure 5: Quantification of Network Growth in the MSC Treatment Groups 5 D after Co-culture Establishment. Low magnification phase contrast microscopy images were taken from the aortic ring treatment groups to measure network growth and development (A). Scattered arrow shows the direction of network growth. Scale bar = 250 µm. Microscopy images were used to quantify network properties, including mean network growth and mean network loop formation in one assay quadrant. FTM HUCPVCs contributed to greater network growth when compared to BMSC co-cultures and untreated networks (p ≤0.001) (B). The p value was calculated using a one-way ANOVA to be p <0.0001 using Tukey's Post test (N = 3). Network loops with at least four closed sides were quantified (C). FTM HUCPVC co-cultures developed greater network loops when compared to BMSC co-cultures (p ≤0.001) and untreated networks (p ≤0.01). The p value was calculated using a one-way ANOVA to be p <0 .0001 using Tukey's Post test (N = 3). The average of 4 fields of endothelial networks were quantified. Scale bar = 250 µm. For pairwise comparison, * = p ≤0.05, ** = p ≤0.01, *** = p ≤0.001. Please click here to view a larger version of this figure.
Figure 6: Possible Errors in the Aortic Ring Assay Establishment. Aortic rings embedded in incomplete BME polymerization may lead to inconsistent and broken endothelial networks (A). FTM HUCPVCs stained for long periods with large time gaps between staining and establishing co-cultures, yield minimal homing and endothelial cell interactions (B). The endothelial network from rat aorta stored in -80 °C and thawed for aortic ring assay (C). A: Scale bar = 250 µm B, C: Scale bar = 400 µm. Please click here to view a larger version of this figure.
Supplementary Figure 1: Flow Cytometry Analysis of Human FTM HUCPVCs Extracted from Aortic Ring Co-cultures. Cellular fractions of aortic ring cultures were isolated and processed for flow cytometry analysis. Fluorophore conjugated antibody against human specific cell surface marker (TRA-1-85, APC) was applied in combination with endothelial marker (CD31, FITC, (A)) or pericyte marker (CD146, FITC, (B)) specific antibodies. The cell population positive for the human marker (y axis) tested low positivity for endothelial marker CD31 (A, Q2) and high positive for pericyte marker CD146 (B, Q6). This suggests that FTM HUCPVCs maintained their perivascular cell properties in the aortic ring co-cultures and did not develop an endothelial phenotype. The quadrants on plots were defined using isotype controls matching the applied primary antibodies. Please click here to download this file.
Supplementary Figure 2: Quantitative PCR Analysis of Key Angiogenic Genes. The similar gene expression of key angiogenic genes by FTM HUCPVCs and BMSCs following 1-week co-culture in the aortic ring assay (A). Representative CT values are shown in table (B). Please click here to download this file.
Discussion
There are several critical stages in setting up a successful aortic ring assay MSC co-culture experiment. First, the most important steps when isolating and sectioning aorta are: 1) obtaining exclusively the thoracic segment of aorta; 2) carefully removing branching blood vessels, connective and adipose tissue and; 3) cutting even sections of the aorta (~1 mm) to limit variability between each assay. Second, the successful embedding of aortic rings into BME is critical for this assay. If the BME is not completely polymerized or polymerized unevenly, the aortic rings embedded in the BME will not be able to initiate endothelial sprouting or they may develop discontinuous endothelial networks. If the BME polymerization is insufficient, assays are not reliable for quantification. Third, the complete factor-enriched endothelial media is necessary to provide nourishment for the aortic rings to initiate sprouting. Once the aortic ring rings have initiated vessel sprouting, the next critical upcoming steps involve the cell type to be tested. To this end, MSCs need to be successfully pre-stained and administered to the sprouting aortic rings. There are two critical procedures for this section. 1) Selecting the appropriate day for seeding: the networks should be in the developing stage and not be reaching high coverage of the culture well. If the MSCs are not administered in a timely manner, it becomes challenging to quantify the net effects of the MSCs on network development. 2) Prestaining the MSCs prior to addition: if the MSCs are over-stained or remain in pellet for an extensive period of time, clumping will occur that impairs homing and integration within endothelial networks. Lastly, quantifying the networks can easily be achieved if appropriate controls are prepared. Aortic rings without MSCs will develop endothelial networks because the aorta itself supports sprouting, but the MSCs may promote greater network development with different network structures as described in this manuscript. The quantification of endothelial networks should be performed at least 5 d following establishing co-cultures. Due to the closed system, angiogenic response is transient and endothelial networks start degenerating following approximately 7 d. If the purpose of the observation is to evaluate the tested cell type effect on the development of endothelial networks, the images should be acquired within the first week of co-culturing.
A possible modification of the current assay can be the application of primary human tissue. As a 1 mm section of rat aorta makes one unit for co-culture, human aortas can provide with a high number of sections of highly consistent measurements. Despite the highly-restricted availability of viable human primary tissue, the high output suggests potential feasibility. When developing an assay for large scale evaluations, pre-forming the assay and storing its components until cells for testing become available can be considered. The BME coating can be prepared on tissue culture plates and stored in a frozen, dehydrated form for longer periods of time. Also, as we tested in our laboratory, rat aortic tissue can be frozen and stored for later application, thus making work with the most crucial element of the assay more convenient.
Despite the many advantages of the aortic ring assay, there are a few limitations to be listed. First, the establishment of the assay cultures can be challenging. In its present form, the routine application requires sufficient manual skills. Operator errors can introduce variability. It can be mirrored in network growth and can make it difficult to reliably evaluate the effect of administered cells and consequently the important angiogenic properties. However, these challenges are similar if not smaller when compared to those of other methods available, and the required training period can be conveniently short and economic as opposed to in vivo experiments. Second, a lag period between embedding of the ex vivo aortic tissue and the initial network development that marks the time of cell administration needs to be accounted for by the user. Third, the quantification of endothelial network properties can be time consuming but can be resolved by assigning hallmark parameters, including radial growth, tube length and network mesh dimensions. This can be performed by using computer aided analysis of images (Image J), which can significantly decrease the time required for consistent and reliable quantification. Fourth, the variability between each assay can occur as a result of slight inconsistencies in animal tissue source and handling by the operator. We found that this challenge can be effectively overcome and the quantification becomes statistically reliable by setting up triplicates for each treatment group. Lastly, extracting and sorting the cells of human and rat origin for post-assay analysis can be challenging. However, specific proteinases, including dispase, and cell recovery solution can be applied to recover cells for immunophenotypic or gene expression analysis.
Compared to existing methods that either culture a single endothelial cell type or live animal systems, this presented assay uses the combination of ex vivo aortic tissue and BME. This setup enables the user to gain both qualitative and quantitative data elucidating the angiogenic properties of cell therapy candidates. Combining BME and multi-cellular ex vivo tissue provides properties closely mimicking the microenvironment that the therapeutic cell type may encounter after local administration. Because of the possibility of a high number of parallels and the close control over culture conditions, the operator is provided with a system that contains necessary elements for reliable evaluation, while eliminating variables and inconsistencies found in animal models. Furthermore, assessments are more feasible because it reduces cost and the need of repeated animal procedures. Single cell assays and most animal models lack the factors and variables introduced by immune responses that would otherwise occur in the clinical application of cell therapy candidates. The inflammatory response alters angiogenesis and therefore, the therapeutic effect of implanted substances or cells cannot be accurately quantified. The aortic ring assay excludes many inflammatory components that would disturb assay measurements. However, it contains resident macrophages that are important players in the microvasculature development32. With the easy identification of both the endothelial network and prestained human cells, the effect of the MSC secreted inflammatory cytokines, and even cellular elements of the immune system, can be easily observed using this assay.
High control by the user over the assay conditions and the reproducible nature of the experimental setup allows for the introduction of further elements to the system. Firstly, the aortic ring assay can be used as an injury model. Following the embedding of the aortic rings and endothelial sprouting, assays can be introduced to ischemic injury, inflammatory factors (TNF-α) or bacterial lipopolysaccharide (LPS), followed by MSC addition to determine possible rescue of angiogenic response following injury. Secondly, to elucidate the possible mechanisms of how the MSCs contribute to the angiogenic response, neutralizing antibodies can be utilized to silence potential receptors critical for cell-to-cell interactions. Lastly, to investigate the paracrine properties of MSCs, the aortic ring assay can be setup in a transwell system to exclude the effect of cell-to-cell interactions of the angiogenic response but focus on paracrine properties.
In summary, the aortic ring assay can assess the ability and potency of cell therapy candidates to mediate ECM processing, migrate to areas of angiogenesis, and contribute to vessel development through physical contact. The aortic ring ex vivo angiogenesis assay could be developed as a valuable, quantitative pre-screening tool for candidate cell lines for regenerative therapy.
Disclosures
Dr. Clifford L. Librach is joint holder of the patent: Methods of isolation and use of cells derived from first trimester umbilical cord tissue. Granted in Canada and Australia.
Acknowledgments
The authors thank the following staff members and research personnel: Andrée Gauthier-Fisher, Matthew Librach, Tanya Barretto, Tharsan Velauthapillai, and Sarah Laronde.
References
- Potente M, Gerhardt H, Carmeliet P. Basic and therapeutic aspects of angiogenesis. Cell. 2011;146(6):873–887. doi: 10.1016/j.cell.2011.08.039. [DOI] [PubMed] [Google Scholar]
- Hoeben A, Landuyt B, Highley MS, Wildiers H, Van Oosterom AT, De Bruijn EA. Vascular endothelial growth factor and angiogenesis. Pharmacol Rev. 2004;56(4):549–580. doi: 10.1124/pr.56.4.3. [DOI] [PubMed] [Google Scholar]
- Khan TA, Sellke FW, Laham RJ. Gene therapy progress and prospects: therapeutic angiogenesis for limb and myocardial ischemia. Gene Ther. 2003;10(4):285–291. doi: 10.1038/sj.gt.3301969. [DOI] [PubMed] [Google Scholar]
- Gupta R, Tongers J, Losordo DW. Human studies of angiogenic gene therapy. Circ Res. 2009;105(8):724–736. doi: 10.1161/CIRCRESAHA.109.200386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chu H, Wang Y. Therapeutic angiogenesis: controlled delivery of angiogenic factors. Ther Deliv. 2012;3(6):693–714. doi: 10.4155/tde.12.50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao Y. Therapeutic angiogenesis for ischemic disorders: what is missing for clinical benefits? Discov Med. 2010;9(46):179–184. [PubMed] [Google Scholar]
- Said SS, Pickering JG, Mequanint K. Advances in growth factor delivery for therapeutic angiogenesis. J Vasc Res. 2013;50(1):35–35. doi: 10.1159/000345108. [DOI] [PubMed] [Google Scholar]
- Bergers G, Song S. The role of pericytes in blood-vessel formation and maintenance. Neuro Oncol. 2005;7(4):452–464. doi: 10.1215/S1152851705000232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leeper NJ, Hunter AL, Cooke JP. Stem cell therapy for vascular regeneration: adult, embryonic, and induced pluripotent stem cells. Circulation. 2010;122(5):517–526. doi: 10.1161/CIRCULATIONAHA.109.881441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sieveking DP, Ng MK. Cell therapies for therapeutic angiogenesis: back to the bench. Vasc Med. 2009;14(2):153–166. doi: 10.1177/1358863X08098698. [DOI] [PubMed] [Google Scholar]
- Staton CA, Reed MW, Brown NJ. A critical analysis of current in vitro and in vivo angiogenesis assays. Int J Exp Pathol. 2009;90(3):195–221. doi: 10.1111/j.1365-2613.2008.00633.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Auerbach R, Lewis R, Shinners B, Kubai L, Akhtar N. Angiogenesis assays: a critical overview. Clin Chem. 2003;49(1):32–40. doi: 10.1373/49.1.32. [DOI] [PubMed] [Google Scholar]
- Tahergorabi Z, Khazaei M. A review on angiogenesis and its assays. Iran J Basic Med Sci. 2012;15(6):1110–1126. [PMC free article] [PubMed] [Google Scholar]
- Arnaoutova I, George J, Kleinman HK, Benton G. The endothelial cell tube formation assay on basement membrane turns 20: state of the science and the art. Angiogenesis. 2009;12(3):267–274. doi: 10.1007/s10456-009-9146-4. [DOI] [PubMed] [Google Scholar]
- Arnaoutova I, Kleinman HK. In vitro angiogenesis: endothelial cell tube formation on gelled basement membrane extract. Nat Protoc. 2010;5(4):628–635. doi: 10.1038/nprot.2010.6. [DOI] [PubMed] [Google Scholar]
- Norrby K. In vivo models of angiogenesis. J Cell Mol Med. 2006;10(3):588–612. doi: 10.1111/j.1582-4934.2006.tb00423.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicosia RF. The aortic ring model of angiogenesis: a quarter century of search and discovery. J Cell Mol Med. 2009;13(10):4113–4136. doi: 10.1111/j.1582-4934.2009.00891.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baker M, et al. Use of the mouse aortic ring assay to study angiogenesis. Nat Protoc. 2011;7(1):89–104. doi: 10.1038/nprot.2011.435. [DOI] [PubMed] [Google Scholar]
- Guo J, et al. A secreted protein (Canopy 2, CNPY2) enhances angiogenesis and promotes smooth muscle cell migration and proliferation. Cardiovasc Res. 2015;105(3):383–393. doi: 10.1093/cvr/cvv010. [DOI] [PubMed] [Google Scholar]
- Wittig C, Scheuer C, Parakenings J, Menger MD, Laschke MW. Geraniol Suppresses Angiogenesis by Downregulating Vascular Endothelial Growth Factor (VEGF)/VEGFR-2 Signaling. PLoS One. 2015;10(7):e0131946. doi: 10.1371/journal.pone.0131946. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masson VV, et al. Mouse Aortic Ring Assay: A New Approach of the Molecular Genetics of Angiogenesis. Biol Proced Online. 2002;4:24–31. doi: 10.1251/bpo30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caplan AI. Adult mesenchymal stem cells for tissue engineering versus regenerative medicine. J Cell Physiol. 2007;213(2):341–347. doi: 10.1002/jcp.21200. [DOI] [PubMed] [Google Scholar]
- Caplan AI, Dennis JE. Mesenchymal stem cells as trophic mediators. J Cell Biochem. 2006;98(5):1076–1084. doi: 10.1002/jcb.20886. [DOI] [PubMed] [Google Scholar]
- Keating A. Mesenchymal stromal cells: new directions. Cell Stem Cell. 2012;10(6):709–716. doi: 10.1016/j.stem.2012.05.015. [DOI] [PubMed] [Google Scholar]
- Kilroy GE, et al. Cytokine profile of human adipose-derived stem cells: expression of angiogenic, hematopoietic, and pro-inflammatory factors. J Cell Physiol. 2007;212(3):702–709. doi: 10.1002/jcp.21068. [DOI] [PubMed] [Google Scholar]
- Tang YL, et al. Paracrine action enhances the effects of autologous mesenchymal stem cell transplantation on vascular regeneration in rat model of myocardial infarction. Ann Thorac Surg. 2005;80(1):229–236. doi: 10.1016/j.athoracsur.2005.02.072. [DOI] [PubMed] [Google Scholar]
- Caplan AI. All MSCs are pericytes? Cell Stem Cell. 2008;3(3):229–230. doi: 10.1016/j.stem.2008.08.008. [DOI] [PubMed] [Google Scholar]
- Hong SH, et al. Ontogeny of human umbilical cord perivascular cells: molecular and fate potential changes during gestation. Stem Cells Dev. 2013;22(17):2425–2439. doi: 10.1089/scd.2012.0552. [DOI] [PubMed] [Google Scholar]
- Kilkenny C, Browne WJ, Cuthill IC, Emerson M, Altman DG. Improving bioscience research reporting: The ARRIVE guidelines for reporting animal research. J Pharmacol Pharmacother. 2010;1(2):94–99. doi: 10.4103/0976-500X.72351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sarugaser R, Ennis J, Stanford WL, Davies JE. Isolation, propagation, and characterization of human umbilical cord perivascular cells (HUCPVCs) Methods Mol Biol. 2009;482:269–279. doi: 10.1007/978-1-59745-060-7_17. [DOI] [PubMed] [Google Scholar]
- Schneider CA, Rasband WS, Eliceiri KW. NIH Image to ImageJ: 25 years of image analysis. Nat Methods. 2012;9(7):671–675. doi: 10.1038/nmeth.2089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gelati M, Aplin AC, Fogel E, Smith KD, Nicosia RF. The angiogenic response of the aorta to injury and inflammatory cytokines requires macrophages. J Immunol. 2008;181(8):5711–5719. doi: 10.4049/jimmunol.181.8.5711. [DOI] [PMC free article] [PubMed] [Google Scholar]