

Abstract
Background
Currently, some studies have demonstrated that miR-34a could serve as a suppressor of several cancers including hepatocellular carcinoma (HCC). Previously, we discovered that miR-34a was downregulated in HCC and involved in the tumorigenesis and progression of HCC; however, the mechanism remains unclear. The purpose of this study was to estimate the expression of miR-34a in HCC by applying the microarray profiles and analyzing the predicted targets of miR-34a and their related biological pathways of HCC.
Methods
Gene expression omnibus (GEO) datasets were conducted to identify the difference of miR-34a expression between HCC and corresponding normal tissues and to explore its relationship with HCC clinicopathologic features. The natural language processing (NLP), gene ontology (GO), pathway and network analyses were performed to analyze the genes associated with the carcinogenesis and progression of HCC and the targets of miR-34a predicted in silico. In addition, the integrative analysis was performed to explore the targets of miR-34a which were also relevant to HCC.
Results
The analysis of GEO datasets demonstrated that miR-34a was downregulated in HCC tissues, and no heterogeneity was observed (Std. Mean Difference(SMD) = 0.63, 95% confidence intervals(95%CI):[0.38, 0.88], P < 0.00001; Pheterogeneity = 0.08 I2 = 41%). However, no association was found between the expression value of miR-34a and any clinicopathologic characteristics. In the NLP analysis of HCC, we obtained 25 significant HCC-associated signaling pathways. Besides, we explored 1000 miR-34a-related genes and 5 significant signaling pathways in which CCND1 and Bcl-2 served as necessary hub genes. In the integrative analysis, we found 61 hub genes and 5 significant pathways, including cell cycle, cytokine-cytokine receptor interaction, notching pathway, p53 pathway and focal adhesion, which proposed the relevant functions of miR-34a in HCC.
Conclusion
Our results may lead researchers to understand the molecular mechanism of miR-34a in the diagnosis, prognosis and therapy of HCC. Therefore, the interaction between miR-34a and its targets may promise better prediction and treatment for HCC. And the experiments in vivo and vitro will be conducted by our group to identify the specific mechanism of miR-34a in the progress and deterioration of HCC.
Electronic supplementary material
The online version of this article (10.1186/s12885-017-3941-x) contains supplementary material, which is available to authorized users.
Keywords: Hepatocellular carcinoma, miRNA-34a, Gene expression omnibus, Gene ontology, Network analysis
Background
Liver cancer ranks as the second leading cause of cancer death in men in less developed countries and sixth in more developed countries [1]. During 2012, 745,500 deaths occurred out of estimated 782,500 new HCC cases worldwide, with nearly 50% happening in China [1].High prevalence is observed in parts of East and South-East Asia where chronic hepatitis B virus (HBV) and hepatitis C virus (HCV) infection are epidemic [2]. Hepatocellular carcinoma (HCC) is recognized as the most common type of liver cancer, accounting for 90% of primary liver cancer. There exist difficulties in treating HCC due to a series of sequential and complex processes involved in the carcinogenesis and progression of HCC. Although radiation, surgery, liver transplantation or chemotherapy are widely used in the therapy of HCC, the survival rate of HCC patients is still less than 5% [3]. Therefore, it is urgent to enhance our knowledge of molecular pathogenesis of HCC and explore novel biomarkers in favor of therapy of HCC. Gene signatures would provide efficient molecular basis of the clinicopathological features for characterizing the heterogeneity of HCC. In addition, the regulatory pathways and networks involved in the mechanism of HCC would lead to the identification of molecular fingerprints for directing therapeutic strategies.
MicroRNAs(miRNAs) are composed of a family of endogenous, noncoding small RNA molecules(∼18–25 nucleotides long), which serve as post-transcriptional gene expression regulators via binding to the 3′--untranslated regions(3’-UTRs) region of their target messenger RNAs(mRNA) [4]. Besides, miRNAs participated in multiple biological processes in oncology via inhibiting translation of mRNA or degrading mRNA [5–8]. Several studies have identified that ectopic expression of miR-34a is related to cell cycle, proliferation, migration, invasion,apoptosis and prognosis of cancers by targeting AXL/ SIRT1/Yin Yang-1 [9–12]. Several studies have shown that miR-34a regulates the carcinogenesis and progression of cancers via modulation of Notch1 Pathway, SIRT1/p53 pathway, WNT/TCF7 signaling [13–15]. Kang et al. and Zhou et al. have reported that miR-34a is associated with chemo-resistance [13, 16]. One study conducted by Cao et al. has found that miR-34a could influence the impact of lincRNA-UFC1 on proliferation and apoptosis in HCC cells [17]. In our previous study, we demonstrated that miR-34a decreased in HCC, and in vitro experiment has also identified that miR-34a could inhibit cell proliferation, invasion and migration, and increase caspase activity and cellular apoptosis by modulating phospho-ERK1/2 and phospho-stat5 signaling, as well as the level of c-MET [18]. However, the molecular mechanism of miR-34a in HCC tumorigenesis and development remains unclear. Therefore, a systemic and comprehensive understanding of miR-34a-target genes and relevant signaling pathways is important for diagnosis, therapy and prognosis of HCC.
In this study, GEO datasets were applied to verify the associations between miR-34a expression and HCC. Additionally, we performed a series of analyses to assess the miR-34a-predicted genes which are associated with carcinogenesis, progression and chemo-resistance in HCC. We then evaluated the potential value of miR-34a in HCC diagnosis, prognosis and therapy with network and pathway analyses.
Methods
Comprehensive analysis of miR-34a expression in HCC based on GEO datasets
The expression data of miR-34a was collected from the GEO(http://www.ncbi.nlm.nih.gov/geo/) databases up to January 2016 with the following keywords: (“HCC” OR “hepatocellular carcinoma” OR “liver cancer” OR “liver carcinoma” OR “liver malignan*” OR “liver neoplasm”) and (“miRNA” OR “microRNA”). Inclusion criteria were as follows: (1) both HCC tissues and adjacent HCC tissues (or healthy liver tissues) or only HCC tissues were included in each dataset with each group containing more than two samples; (2) the dataset sample organism was Homo sapiens; (3) the expression data of miR-34a (has-miR-34a or has-miR-34a-5p) from the experimental and control groups could be acquired or calculated. Expression values of miR-34a and sample size in both test and control groups were calculated. Moreover, means and standard deviations of these values were extracted to estimate the different levels of miR-34a in case and control groups by Review Manager (Revman Version 5.3, Copenhagen, Denmark) with random-effects model. The chi-square test and the I2 statistic were applied to evaluate the heterogeneity across studies. Heterogeneity was considered to exist when the P value < 0.05 or I2 > 50%. Further, SMD and its 95%CI were pooled to evaluate the stability of the analysis. It was considered to be statistically significant if the corresponding 95%CI for the pooled SMD did not overlap 1 or −1. Additionally, for sensitivity analysis, we eliminated every study to evaluate the source of heterogeneity.
NLP analysis of HCC
Extraction and filtering of data
We searched PubMed in an attempt to identify all relevant studies published between January 1980 and May 2015. Publications were retrieved with the following key words: (hepatocellular carcinoma) and (resistance or prognosis or metastasis or recurrence or survival or carcinogenesis or sorafenib or bevacizumab) and (“1980/01/01” [PDAT]: “2015/05/25” [PDAT]). All genes and proteins related to the key words were extracted and gathered in a list with the following of gene mention tagging by applying A Biomedical Named Entity Recognizer (ABNER, an open source tool for automatically tagging genes, proteins and other entity names in text, http://pages.cs.wisc.edu/bsettles/abner/) [19] and conjunction resolution. The flow chart of NLP analysis was shown in Fig. 1.
Fig. 1.
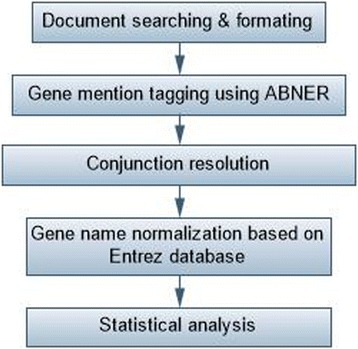
Flow chart of the natural language processing (NLP) analysis of hepatocellular carcinoma
Statistical analysis
The frequency was calculated for each gene occurrence in the NLP analysis. The higher frequency implies the stronger association between HCC and certain genes. The number of all eligible literatures in PubMed database was recorded as N. The frequency of certain gene and HCC in PubMed was marked as “m” and “n”, respectively. It was denoted as “k” when a gene and HCC occurred simultaneously. Then we calculated the possibility of frequency greater than “k” co-citation via hypergeometric distribution in completely random conditions.
The comprehensive analysis of HCC- related genes
We conducted GO analysis on the functions of differently expressed genes in HCC and classified the related genes into three major groups: biological processes, cellular component and molecular functions. Pathway analysis was applied in GenMAPP v2.1 for Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment and the value of P was calculated to select the significantly enriched pathway. We also combined three different interactions: 1) protein interaction, gene regulation, protein; 2) modification of the existing high-throughput experiments; 3) interactions between genes mentioned previously. In addition, the pathway data was downloaded from KEGG database and the interactions between genes were analyzed by KEGGSOA (http://www.bioconductor.org/packages/2.4/bioc/html/ KEGGSOAP.html) package from R (http://www.r-project.org/), including enzyme–enzyme relation, protein–protein interaction and gene expression interaction [20].
We downloaded the data of protein interactions from MIPS database (http://mips.helmholtz-muenchen.de/proj/ppi/) [21]. For the interaction mentioned previously, algorithm co-citation in the abstracts in PubMed was used to analyze gene term and all term variants co-occurring with the certain gene. And we also calculated the frequency of the cocitation genes. Then we conducted statistical analyses according to the description in NLP analysis. Finally, medusa software was used to perform the network.
Prediction of miRNA-34a target genes
We analyzed the predicted targets of miR-34a by applying 11 independent software: DIANA-microT, MicroInspector, miRanda, MirTarget2, miTarget, NBmiRTar, PicTar, PITA, RNA22, RNAhybrid and TargetScan. The results were considered reliable when the results were calculated by four or more software. GO analysis, pathway analysis and network analysis of miR-34a target genes were performed in accordance with the same principles of genes from NLP analysis.
Integrative analysis of miR-34a targets and the natural language processing
We calculated the overlap of miR-34a targets predicted in silico and HCC related-genes acquired from NLP analysis. And the ingenuity pathway analysis was carried out to show the relationships between miR-34a and its target genes associated with HCC. The overlap of the miR-34a target genes predicted by in-silico bioinformatics tools and HCC-related genes was performed subsequently.
Results
Comprehensive analysis of GEO datasets
Fourteen eligible datasets were involved in this study, which included 3 datasets containing HCC tissues and 11 datasets containing both HCC and adjacent tumor (or healthy) tissues (Table 1). The result showed that the expression level of miR-34a in HCC tissues was statistically lower than that in normal tissues with no heterogeneity (SMD = 0.63, 95%CI [0.38, 0.88], P < 0.00001; P heterogeneity = 0.08 I2 = 41% Figure 2).The three studies which contained HCC tissues only were used to evaluate the difference of miR-34a expression in HCC patients with or without vascular invasion. The comparison identified that miR-34a expression was not associated with vascular invasion in HCC patients (SMD = −1.44, 95%CI: [−0.16,-2.71], P = 0.03; Pheterogeneity < 0.00001 I2 = 95%, Figure 3a).Besides, there existed significant heterogeneity after we had reviewed every study. Among the 11 datasets mentioned above, two datasets (GSE69580, GSE10694) were applied to estimate the relationship of miR-34a with cirrhosis in HCC patients. Another two datasets (GSE10694, GSE41874) were performed to analyze whether the expression of miR-34a was related to metastasis in patients with HCC. Finally, no relationship was observed between miR-34a expression and cirrhosis or metastasis (Pcirrhosis = 0.79, Fig. 3b; Pmetastasis = 0.77, Figure 3c).
Table 1.
Characteristics of miR-34a gene expression used in the analysis of GEO datasets
| GEO accession | Author | Sample size | Country | Platform | |
|---|---|---|---|---|---|
| HCC patients | Healthy controls | ||||
| GSE10694 | Gu et al. | 78 | 88 | China | GPL6542 |
| GSE12717 | Su et al. | 10 | 6 | China | GPL7274 |
| GSE22058 | Burchard | 73 | 73 | USA | GPL10457 |
| GSE21362 | Sato et al. | 96 | 96 | Japan | GPL10312 |
| GSE31383 | Hoshida et al. | 9 | 10 | USA | GPL10122 |
| GSE36915 | Lee et al. | 68 | 21 | Taiwan | GPL8179 |
| GSE40744 | Farci et al. | 14 | 12 | USA | GPL14613 |
| GSE41874 | Okano et al. | 6 | 4 | Japan | GPL7722 |
| GSE64632 | Selaru et al. | 6 | 6 | USA | GPL18116 |
| GSE67882 | Banerjee et al. | 4 | 8 | India | GPL10850 |
| GSE69580 | Hung et al. | 5 | 5 | Taiwan | GPL10850 |
| GSE20594 | Hoshida et al. | 45 | 32 | USA | GPL10122 |
| GSE67138 | Chuang et al. | 23 | 34 | USA | GPL8786 |
| GSE67139 | Chuang et al. | 63 | 57 | USA | GPL8786 |
Fig. 2.
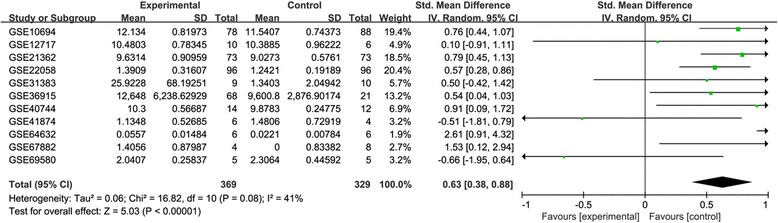
Forest plot showing SMD of miR-34a expression between HCC tissues and corresponding normal tissues
Fig. 3.
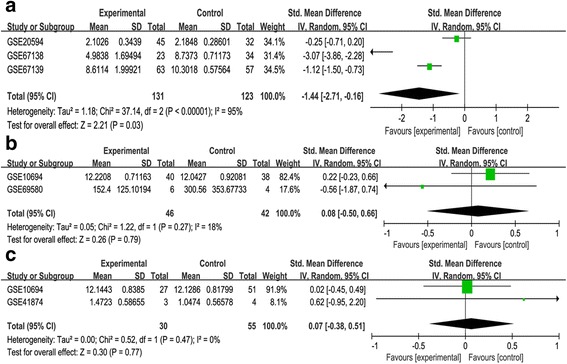
Forest plot showing the association between miR-34a expression and HCC clinocopathologic characteristics. a Forest plot showing SMD of miR-34a expression in HCC tissues with or without vascular invasion. b Forest plot showing SMD of miR-34a expression in HCC tissues with or without cirrhosis. c Forest plot showing SMD of miR-34a expression in HCC tissues with or without metastasis
NLP analysis of HCC
The analysis of HCC-related genes
A total of 64,577 articles for HCC-related molecules were identified for initial browse after primary search in PubMed. A total of 1800 HCC-relevant genes were obtained. All of these cancer-related genes were categorized into different biological processes, cellular components, molecular functions according to GO analysis (Additional file 1: Table S1). In the pathway analysis, we found 25 signaling pathways were significant (P < 0.005), which has been discussed in our previously published work [20]. By constructing gene network, we found the hub genes functioned as a key factor in regulating the stability of the network. In our previous study, the gene network of those 1800 genes were performed [20].The published article also showed that several hub genes were highly related to other genes: PIK3CA, PIK3R2, MAPK1, MAPk3, JAK2, EGFR, KRAS, NRAS [20].
The analysis of miR-34a predicted targets
In the present study, we analyzed the potential targets of miR-34a to explore the biological function of miR-34a relying on its target-protein-coding genes. One thousand potential genes were identified and categorized in GO analysis (Additional file 2: Table S2). Subsequent to the pathway analysis,five pathways were discovered to be statistically significant: Focal adhesion, p53 signaling pathway, Cell cycle, Cytokine-cytokine receptor interaction, Notch signaling pathway (P < 0.05, Table 2).
Table 2.
Pathway analysis of miR-34a-related genes
| term | count | P value | genes |
|---|---|---|---|
| hsa04510:Focal adhesion | 8 | 5.09E-04 | CCND1, MAP2K1, PGF, BCL2, MET, VEGFA, PDGFRA, RELN |
| hsa04115:p53 signaling pathway | 5 | 0.001385856 | CCNE2, CCND1, SERPINE1, CDK6, IGFBP3 |
| hsa04110:Cell cycle | 6 | 0.001921952 | CCNE2, E2F3, CCND1, E2F5, CDK6, CDC25A |
| hsa04060:Cytokine-cytokine receptor interaction | 6 | 0.039495189 | CCL22, MET, VEGFA, PDGFRA, KITLG, KIT |
| hsa04330:Notch signaling pathway | 3 | 0.045372273 | NOTCH2, NOTCH1, JAG1 |
The genes were obtained from the natural language processing (NLP) analysis and 5 signaling pathways were significant (P < = 0.05)
In the network analysis, only two genes are statistically significant and both of them were the highest hub genes, including CCND1 and BCL2 (Fig. 4c, Figure 4d). CCND1 participates in the regulation of the three pathways mentioned in Table 2 (focal adhesion, p53 signaling pathway, cell cycle) and BCL-2 acts as a member of focal adhesion. In addition, the expression of CCND1 and BCL-2 were both associated with the biological processes described in GO analysis: cell cycle and proliferation, stress response, developmental processes, protein metabolism.
Fig. 4.
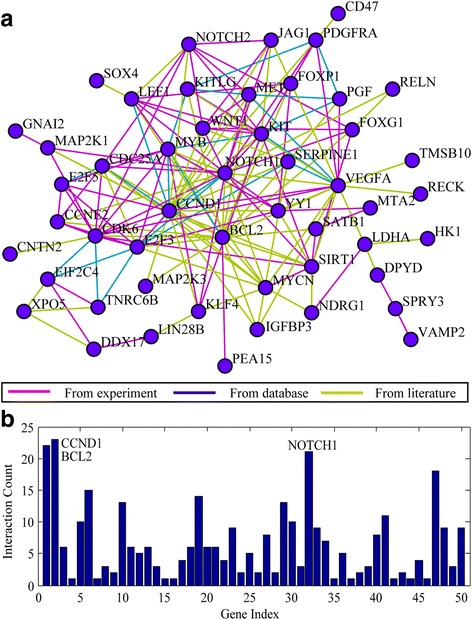
Network analysis and connectivity analysis of miR-34a targets. a Network analysis of miR-34a targets. Brown represents association, green represents inhibition and blue represents activation. b Connectivity analysis of miR-34a targets. The connectivity of Bcl-2 is the highest one which has a total of twenty-two related-genes (z-test, P = 0.0037)
Integrative analysis of miR-34a target genes and the NLP results
In the integrative analysis, we calculated the overlap of miR-34a targets and HCC-related genes obtained from the NLP analysis. As a result, 61 HCC-related genes, such as VEGFA, Bcl-2, CCND1, MET, KIT, Notch1, DPYD, SERPINE1 and CDK6 were potentially modulated by miR-34a, were summarized in Additional file 3: Table S3. Besides, the relationship among these genes was shown in Fig. 5, including five significant pathways: cell cycle (P = 0.001921952); cytokine-cytokine receptor interaction (P = 0.039495189); notching signaling pathway (P = 0.045372273); p53 pathway (P = 0.001385856) and focal adhesion (P = 5.09E-04). Among the five pathways mentioned above, the three genes (E2F3, E2F5, CDC25A) were involved in the cell cycle pathway, and KIT/CCL22 were found in the cytokine-cytokine receptor interaction pathway. Besides, Notch1/Notch2/JAG1 and CDK6/IGFPB3/CCNE2 were significantly related to the notching signaling pathway and p53 pathway, respectively. Additionally, we concluded that VEGFA, Bcl-2, CCND1, MET, PDGFRA were related to the functions of focal adhesion in HCC patients.
Fig. 5.
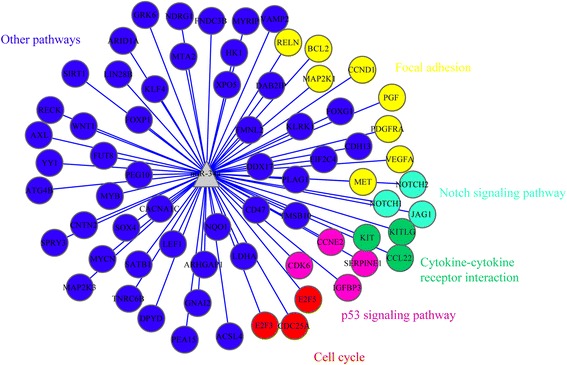
Integrative -analysis of miR-34a target genes and the NLP results. Sixty-nine overlapping genes and their functional pathway that are not only associated with the molecular mechanism of HCC but also are the potential miR-34a target genes were obtained in this final integrative-analysis
Discussion
Nowadays, in order to prolong the survival time of HCC patients, more and more therapies are applied in the clinical stage, including radiotherapy, interventional operation, combined therapy. However, the survival rate is still unsatisfactory. In recent years, more and more researches have focused on the molecular targeted therapy and made exciting progress. Liu et al. have concluded that miR-222 and miR-494 could enhance HCC patients’ resistance to sorafenib by activating the PI3K/AKT signaling pathway [22, 23]. The study reported by Lin et al. has also identified that miR-21 partially reduced the cytotoxic effects of sorafenib in combination with matrine against HCC [24]. However, another two reports have verified that increased expression of miRNAs could assist the efficiency of sorafenib treatment for HCC patients [25, 26]. In our previous study, we identified that increased expression of miR-34a might help the diagnosis and prognosis of HCC by regulating c-MET [18]. Although numerous articles have verified the functions of miR-34a in the carcinogenesis and progression of HCC, the molecular mechanism of miR-34a related to HCC still remains unclear. Therefore, this article was the first to identify the relationship of miR-34a expression with HCC based on microarray data. In addition, we separately analyzed the related pathways of HCC-relate genes and miR-34a targets, and further explored the potential molecular mechanism of miR-34a by overlapping genes associated with HCC and miR-34a.
The consequences of GEO analysis testified that miR-34a expression was downregulated in HCC tissues, which was consistent with our previous study [18]. On the contrary, another two articles have demonstrated that upregulation of miR-34a could promote the proliferation of HCC [27, 28]. The research conducted by Gougelet et al. has identified that the higher expression of miR-34a induced by activation of β-catenin could enhance the risk of HCC by targeting CCND1 [27]. However, the other study has testified that Aflatoxin-B1 (AFB1) might contribute to the progression of HCC by upregulating miR-34a, which may down-regulate Wnt/β-catenin signaling pathway [28]. Besides, due to the limitation of sample size, no statistical significance was found in the associations of miR-34a expression value with HCC clinicopathological features. Nevertheless, some other studies have shown that the low expression of miR-34a might promote the progression of HCC.
Two researchers have suggested that ectopic expression of miR-34a was related to tumor metastasis and invasion via modulating c-MET signaling pathway [18, 29]. Cheng et al. have also provided evidence that abrogation functions of miR-34a could contribute to HCC development through cell cycle pathway and p53 pathway [30]. Moreover, it has been demonstrated that miR-34a/ toll-like receptor 4(TLR4) axis may function as a key regulator in increasing the risk of HCC [31]. Besides, another article has verified that overexpression of miR-34a could improve the effect of cisplatin treatment by enhancing cytotoxicity of NK cells for HCC patients [32]. In addition, Lou et al. have shown that decreased expression of miR-34a may reduce the sensitivity of HCC cells to quercetin by upregulating SIRT1 and downregulating p53 [11]. Therefore, the functions of miR-34a in HCC are complex and still needed further investigation.
In order to explore the molecular mechanism of miR-34a in moderating the progression of HCC, we conducted a series of bioinformatics analyses. The results showed that among the targets of miR-34a, the most significant hub genes were CCND1 and Bcl-2, which were parts of focal adhesion, p53 signaling pathway, cell cycle pathway. Meanwhile, we also found these two genes (CCND1 and Bcl-2) were included in the hub genes which were calculated from the interactive analysis of miR-34a and the NLP analysis. MiR-195 could suppress the development of cancers by targeting CCND1 [33]. Besbes et al. have also concluded that decreased expression of Bcl-2 family members contributed to the progression of apoptosis in cancers [34]. Especially, Gougelet et al. and Zhu et al. have suggested that the feedback loop composed of miR-34a/β-catenin/CCND1 played a critical role in regulating the progression of HCC [27, 28]. Hence, miR-34a may serve as an important regulator in the development of HCC by targeting CCND1 and Bcl-2.
In the interactive analysis, we not only calculated the hub genes, but also the statistically significant pathways involved in moderating the progression of miR-34a-related HCC. The outcome showed eight genes-VEGFA, Bcl-2, CCND1, MET, NOTCH1, SERPINE1, DYPD and CDK6-had the highest connectivity. It was worth noticing that the eight genes were classified into five significantly different pathways. It suggested that miR-34a may impact the development and progression of HCC by moderating cell cycle, cytokine-cytokine receptor interaction, notching pathway, p53 pathway and focal adhesion. Thus, the functions of every genes involved in the five pathways were essential in understanding the effects of miR-34a on HCC patients.
Cell cycle
E2F3 and E2F5 are members of the E2F family which is composed of transcription factors and attributed to the cell cycle progression by regulating G1/S-phase transition [35]. Some studies have shown that miR-503 and miR-195 could inhibit the G1/S transition by suppressing the expression of E2F5 [36, 37]. And the downregulation of E2F5 has been identified to be associated with development of HBV-related HCC [38]. Several articles have suggested that miRNAs could suppress the proliferation, metastasis and invasion of HCC cells by targeting E2F3 [39–41]. Additionally, it has been verified that the inhibition of E2F3 induced by overexpression of miRNAs could enhance the sensitivity of HCC patients to anti-cancer agents and decelerate the progression of HCC [39, 42].
Cytokine-cytokine receptor interaction
C-kit is the receptor of stem cell factor (SCF), and the activation of c-kit have been suggested to be crucial for cell proliferation and migration [43]. The activation of c-kit has been suggested to be attributed to the cell proliferation and cirrhosis of HCC [43, 44]. Yang et al. have proved that the activation of TGF-β-miR-34a-CCL22 signaling could promote the progression of portal vein tumor thrombus in HCC patients [45]. Two studies have also proved that miRNAs could promote the development of HCC by blocking G1/S transition via reducing expression of CDK6 [36, 46].
Notching pathway
Notching signaling pathway composed of notch receptors(Notch1–4) and notch ligands(Jag1) is critical for determining cell fates and associated with therapy of HCC [47, 48]. The result reported by Xue et al. indicated that JAG1/Notch1 signaling is positively associated with the extrahepatic metastasis in HCC by moderating the level of osteopontin (OPN) [49]. However, Wang et al. have concluded that increased expression of Notch1/Jagged1 could promote the progression of HCC via inhibiting beta-catenin expression [50]. Another two studies have indicated that Notch1 may be a therapeutic target by downregulating Wnt/β-catenin pathway and CyclinD1/CDK4 pathway in HBV-associated HCC [51, 52]. In addition, it has been demonstrated that inhibition of Notch2 regulated by C8orf4 could suppress the self-renewal of liver cancer stem cells(CScs) and Notch2 and Jag1 may function as novel therapeutic targets for HCC treatment [53, 54].
P53 pathway
P53 is widely recognized as a tumour suppressor in regulating cell cycle, apoptosis, metabolism and DNA repair [55]. Giacoia et al. have concluded that wide-type p53 could upregulate the expression of miR-107 and then reduce the level of CDK6 and Notch2, which suppresses glioma cell growth [56]. It has been indicated that overexpression of plasminogen activator inhibitor-1 (PAI-1/SERPINE1) could enhance tumour cell proliferation as well as inhibit G(1)-phase transition complexes, cdk4/6/ cyclin D3 and promote the cell-cycle suppressors p53, p27Kip1 and p21Cip1/Waf1 [57]. IGFBP3 induced by p53 have also been verified to be related to the apoptosis of HCC cells [58].
Focal adhesion
The pathway analysis also suggests that Bcl-2, CCND1, PDGFRA, VEGFA and c-MET belong to the focal adhesion pathway. It has also been suggested that CCND1 variants may be positively correlated with the precancerous cirrhosis of hepatocarcinogenesis [59]. Increased expression of platelet-derived growth factor receptor alpha (PDGFRA) promoted by miR-146a has been verified to be associated with the microvascular invasion and poor prognosis of HCC [60]. In addition, it has been shown that the synergism of sorafenib has contributed to the therapy of HCC patients by suppressing the level of MET [61]. It has been identified that HCC patients with amplification of vascular endothelial growth factor A (VEGFA) are more likely to be sensitive to sorafenib [62]. Zhou et al. have also found that miR-503 serve as a suppressor of tumor angiogenesis by targeting VEGFA in HCC patients [63]. However, the functions of genes which were related with miR-34a expression and progression of HCC have not been identified.
In the present study, researchers have paid attention to the results of the comprehensive analyses, especially the section of the integrative –network analysis of miR-34a targets and HCC-related genes. It illustrated the most important genes and pathways involved in the functions of miR-34a in HCC, which may offer researchers more insights into understanding the relationship between miR-34a and HCC. However, the targets of miR-34a were predicted in silico, including genes verified or not verified in experiments, which could make it difficult to explore the exact mechanism of miR-34a in HCC. It is noted that the two genes CCND1 and bcl-2 seem to serve as key regulators between the expression of miR-34a and HCC. Therefore, it is imperative to find out the exact relationships among miR-34a, CCND1/bcl-2 and HCC at a further step.
Conclusions
In summary, it was worth considering that the analyses of miR-34a- targets in HCC would provide effective guidelines for the diagnosis, prognosis and therapy of patients with HCC. The results indicate that miR-34a primarily controls cell cycle, cytokine-cytokine receptor interaction, notching pathway, p53 pathway and focal adhesion pathway in regulating the tumorigenesis and process of HCC. Therefore, these pathways may offer novel insights into the functions of miR-34a in HCC and guide the researchers to find more effective methods to the prevention and treatment of HCC.
Additional files
GO enrichment analysis of HCC related genes. (XLSX 12 kb)
GO enrichment analysis of miR-34a target genes. (XLSX 14 kb)
Integrative-analysis of miR-34a target genes and the natural language processing (NLP) results. Twenty-four overlapping genes that are not only associated with the development and progression of HCC but also are the potential miR-34a target genes were obtained in this integrative–analysis (XLSX 14 kb)
Acknowledgements
Not applicable.
Funding
The study was supported partly by the Fund of National Natural Science Foundation of China (NSFC81560489, NSFC 81260222), the Fund of Guangxi Provincial Health Bureau Scientific Research Project (Z2014054), Youth Science Foundation of Guangxi Medical University (GXMUYSF201311), and Guangxi University Science and Technology Research Projects (LX2014075). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Availability of data and materials
All data generated or analyzed during this study are included in this published article (Tables 1 and 2) and its Additional files (Additional file 1: Table S1, Additional file 2: Table S2 and Additional file 3: Table S3).
Abbreviations
- 3’-UTRs
3′--untranslated regions
- 95%CI
95% confidence intervals
- AFB1
Aflatoxin-B1
- GEO
Gene expression omnibus
- GO
Gene ontology (GO)
- HBV
Hepatitis B virus
- HCC
Hepatocellular carcinoma
- HCV
Hepatitis C virus
- KEGG
Kyoto Encyclopedia of Genes and Genomes
- miRNA
microRNA
- mRNA
Messenger RNAs
- NLP
Natural language processing
- PDGFRA
Platelet-derived growth factor receptor alpha
- SMD
Std. mean difference
- TLR4
Toll-like receptor 4
- VEGFA
Vascular endothelial growth factor A
Authors’ contributions
HY, YWD, ZBF, GC and DZL designed the experiment, interpreted the data and corrected the manuscript. FHR, RQH, JNL, XGL, HWL conducted the experiment, performed the statistical analysis and prepared the manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Footnotes
Jing-ning Lu Deceased.
Electronic supplementary material
The online version of this article (10.1186/s12885-017-3941-x) contains supplementary material, which is available to authorized users.
Contributor Information
Fang-Hui Ren, Email: renfanghui3825318@163.com.
Hong Yang, Email: yanghonggx@163.com.
Rong-quan He, Email: 254199854@qq.com.
Jing-ning Lu, Email: smmdct@qq.com.
Xing-gu Lin, Email: linxinggu@hotmail.com.
Hai-Wei Liang, Email: 1071949934@qq.com.
Yi-Wu Dang, Email: dangyiwu@126.com.
Zhen-Bo Feng, Email: fengzhenbo_gxmu@163.com.
Gang Chen, Email: chen_gang_triones@163.com.
Dian-Zhong Luo, Email: 13878802796@163.com.
References
- 1.Liu Y, Buil A, Collins BC, Gillet LC, Blum LC, Cheng LY, et al. Quantitative variability of 342 plasma proteins in a human twin population. Mol Syst Biol. 2015;11(1):786. doi: 10.15252/msb.20145728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.de Martel C, Ferlay J, Franceschi S, Vignat J, Bray F, Forman D, et al. Global burden of cancers attributable to infections in 2008: a review and synthetic analysis. Lancet Oncol. 2012;13(6):607–615. doi: 10.1016/S1470-2045(12)70137-7. [DOI] [PubMed] [Google Scholar]
- 3.Yang X, Xie X, Xiao YF, Xie R, CJ H, Tang B, et al. The emergence of long non-coding RNAs in the tumorigenesis of hepatocellular carcinoma. Cancer Lett. 2015;360(2):119–124. doi: 10.1016/j.canlet.2015.02.035. [DOI] [PubMed] [Google Scholar]
- 4.Pan L, Ren F, Rong M, Dang Y, Luo Y, Luo D, et al. Correlation between down-expression of miR-431 and clinicopathological significance in HCC tissues. Clin Transl Oncol. 2015;17(7):557–63. doi: 10.1007/s12094-015-1278-y. [DOI] [PubMed] [Google Scholar]
- 5.Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–297. doi: 10.1016/S0092-8674(04)00045-5. [DOI] [PubMed] [Google Scholar]
- 6.Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136(2):215–233. doi: 10.1016/j.cell.2009.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Krol J, Loedige I, Filipowicz W. The widespread regulation of microRNA biogenesis, function and decay. Nat Rev Genet. 2010;11(9):597–610. doi: 10.1038/nrg2843. [DOI] [PubMed] [Google Scholar]
- 8.De Cecco L, Dugo M, Canevari S, Daidone MG, Callari M. Measuring microRNA expression levels in oncology: from samples to data analysis. Crit Rev Oncog. 2013;18(4):273–287. doi: 10.1615/CritRevOncog.2013007207. [DOI] [PubMed] [Google Scholar]
- 9.Nie J, Ge X, Geng Y, Cao H, Zhu W, Jiao Y, et al. miR-34a inhibits the migration and invasion of esophageal squamous cell carcinoma by targeting yin Yang-1. Oncol Rep. 2015;34(1):311–317. doi: 10.3892/or.2015.3962. [DOI] [PubMed] [Google Scholar]
- 10.Zhang H, Li S, Yang J, Liu S, Gong X, Yu X. The prognostic value of miR-34a expression in completely resected gastric cancer: tumor recurrence and overall survival. Int J Clin Exp Med. 2015;8(2):2635–2641. [PMC free article] [PubMed] [Google Scholar]
- 11.Lou G, Liu Y, Wu S, Xue J, Yang F, Fu H, et al. The p53/miR-34a/SIRT1 positive feedback loop in Quercetin-induced apoptosis. Cell Physiol Biochem. 2015;35(6):2192–2202. doi: 10.1159/000374024. [DOI] [PubMed] [Google Scholar]
- 12.Li R, Shi X, Ling F, Wang C, Liu J, Wang W, et al. MiR-34a suppresses ovarian cancer proliferation and motility by targeting AXL. Tumour Biol. 2015; [DOI] [PubMed]
- 13.Kang L, Mao J, Tao Y, Song B, Ma W, Lu Y, et al. MicroRNA-34a suppresses the breast cancer stem cell-like characteristics by downregulating Notch1 pathway. Cancer Sci. 2015;106(6):700–708. doi: 10.1111/cas.12656. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 14.Lai M, Du G, Shi R, Yao J, Yang G, Wei Y, et al. MiR-34a inhibits migration and invasion by regulating the SIRT1/p53 pathway in human SW480 cells. Mol Med Rep. 2015;11(5):3301–3307. doi: 10.3892/mmr.2015.3182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Chen WY, Liu SY, Chang YS, Yin JJ, Yeh HL, Mouhieddine TH, et al. MicroRNA-34a regulates WNT/TCF7 signaling and inhibits bone metastasis in Ras-activated prostate cancer. Oncotarget. 2015;6(1):441–457. doi: 10.18632/oncotarget.2690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Zhou JY, Chen X, Zhao J, Bao Z, Chen X, Zhang P, et al. MicroRNA-34a overcomes HGF-mediated gefitinib resistance in EGFR mutant lung cancer cells partly by targeting MET. Cancer Lett. 2014;351(2):265–271. doi: 10.1016/j.canlet.2014.06.010. [DOI] [PubMed] [Google Scholar]
- 17.Cao C, Sun J, Zhang D, Guo X, Xie L, Li X, et al. The long intergenic noncoding RNA UFC1, a target of MicroRNA 34a, interacts with the mRNA stabilizing protein HuR to increase levels of beta-catenin in HCC cells. Gastroenterology. 2015;148(2):415–426. doi: 10.1053/j.gastro.2014.10.012. [DOI] [PubMed] [Google Scholar]
- 18.Dang Y, Luo D, Rong M, Chen G. Underexpression of miR-34a in hepatocellular carcinoma and its contribution towards enhancement of proliferating inhibitory effects of agents targeting c-MET. PLoS One. 2013;8(4):e61054. doi: 10.1371/journal.pone.0061054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Settles B. ABNER an open source tool for automatically tagging genes, proteins and other entity names in text. Bioinformatics. 2005;21(14):3191–3192. doi: 10.1093/bioinformatics/bti475. [DOI] [PubMed] [Google Scholar]
- 20.Zhang X, Tang W, Chen G, Ren F, Liang H, Dang Y, et al. An encapsulation of gene signatures for Hepatocellular carcinoma, MicroRNA-132 predicted target genes and the corresponding overlaps. PLoS One. 2016;11(7):e0159498. doi: 10.1371/journal.pone.0159498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Mewes HW, Albermann K, Heumann K, Liebl S, Pfeiffer FMIPS. A database for protein sequences, homology data and yeast genome information. Nucleic Acids Res. 1997;25(1):28–30. doi: 10.1093/nar/25.1.28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Liu K, Liu S, Zhang W, Jia B, Tan L, Jin Z, et al. miR-494 promotes cell proliferation, migration and invasion, and increased sorafenib resistance in hepatocellular carcinoma by targeting PTEN. Oncol Rep. 2015;34(2):1003–10. doi: 10.3892/or.2015.4030. [DOI] [PubMed] [Google Scholar]
- 23.Liu K, Liu S, Zhang W, Ji B, Wang Y, Liu Y. miR222 regulates sorafenib resistance and enhance tumorigenicity in hepatocellular carcinoma. Int J Oncol. 2014;45(4):1537–1546. doi: 10.3892/ijo.2014.2577. [DOI] [PubMed] [Google Scholar]
- 24.Lin Y, Lin L, Jin Y, Wang D, Tan Y, Zheng C. Combination of Matrine and Sorafenib decreases the aggressive phenotypes of Hepatocellular carcinoma cells. Chemotherapy. 2014;60(2):112–118. doi: 10.1159/000371736. [DOI] [PubMed] [Google Scholar]
- 25.Vaira V, Roncalli M, Carnaghi C, Faversani A, Maggioni M, Augello C, et al. MicroRNA-425-3p predicts response to sorafenib therapy in patients with hepatocellular carcinoma. Liver Int. 2015;35(3):1077–1086. doi: 10.1111/liv.12636. [DOI] [PubMed] [Google Scholar]
- 26.Sun H, Cui C, Xiao F, Wang H, Xu J, Shi X, et al. miR-486 regulates metastasis and chemosensitivity in hepatocellular carcinoma by targeting CLDN10 and CITRON. Hepatol Res. 2015; [DOI] [PubMed]
- 27.Gougelet A, Sartor C, Bachelot L, Godard C, Marchiol C, Renault G, et al. Antitumour activity of an inhibitor of miR-34a in liver cancer with beta-catenin-mutations. Gut. 2015; [DOI] [PubMed]
- 28.Zhu L, Gao J, Huang K, Luo Y, Zhang B, Xu W. miR-34a screened by miRNA profiling negatively regulates Wnt/beta-catenin signaling pathway in Aflatoxin B1 induced hepatotoxicity. Sci Rep. 2015;5:16732. doi: 10.1038/srep16732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Li N, Fu H, Tie Y, Hu Z, Kong W, Wu Y, et al. miR-34a inhibits migration and invasion by down-regulation of c-met expression in human hepatocellular carcinoma cells. Cancer Lett. 2009;275(1):44–53. doi: 10.1016/j.canlet.2008.09.035. [DOI] [PubMed] [Google Scholar]
- 30.Cheng J, Zhou L, Xie QF, Xie HY, Wei XY, Gao F, et al. The impact of miR-34a on protein output in hepatocellular carcinoma HepG2 cells. Proteomics. 2010;10(8):1557–1572. doi: 10.1002/pmic.200900646. [DOI] [PubMed] [Google Scholar]
- 31.Jiang ZC, Tang XM, Zhao YR, Zheng L. A functional variant at miR-34a binding site in toll-like receptor 4 gene alters susceptibility to hepatocellular carcinoma in a Chinese Han population. Tumour Biol. 2014;35(12):12345–12352. doi: 10.1007/s13277-014-2547-z. [DOI] [PubMed] [Google Scholar]
- 32.Shi L, Lin H, Li G, Sun Y, Shen J, Xu J, et al. Cisplatin enhances NK cell immunotherapy efficacy to suppress HCC progression via altering the androgen receptor (AR)-ULBP2 signals. Cancer Lett. 2016; [DOI] [PMC free article] [PubMed]
- 33.Han K, Chen X, Bian N, Ma B, Yang T, Cai C, et al. MicroRNA profiling identifies MiR-195 suppresses osteosarcoma cell metastasis by targeting CCND1. Oncotarget. 2015;6(11):8875–8889. doi: 10.18632/oncotarget.3560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Karpel-Massler G, Shu C, Chau L, Banu M, Halatsch ME, Westhoff MA, et al. Combined inhibition of Bcl-2/Bcl-xL and Usp9X/Bag3 overcomes apoptotic resistance in glioblastoma in vitro and in vivo. Oncotarget. 2015;6(16):14507–21. doi: 10.18632/oncotarget.3993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Attwooll C, Lazzerini Denchi E, Helin K. The E2F family: specific functions and overlapping interests. EMBO J. 2004;23(24):4709–4716. doi: 10.1038/sj.emboj.7600481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Xu T, Zhu Y, Xiong Y, Ge YY, Yun JP, Zhuang SM. MicroRNA-195 suppresses tumorigenicity and regulates G1/S transition of human hepatocellular carcinoma cells. Hepatology. 2009;50(1):113–121. doi: 10.1002/hep.22919. [DOI] [PubMed] [Google Scholar]
- 37.Xiao F, Zhang W, Chen L, Chen F, Xie H, Xing C, et al. MicroRNA-503 inhibits the G1/S transition by downregulating cyclin D3 and E2F3 in hepatocellular carcinoma. J Transl Med. 2013;11:195. doi: 10.1186/1479-5876-11-195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Zou C, Li Y, Cao Y, Zhang J, Jiang J, Sheng Y, et al. Up-regulated MicroRNA-181a induces carcinogenesis in hepatitis B virus-related hepatocellular carcinoma by targeting E2F5. BMC Cancer. 2014;14:97. doi: 10.1186/1471-2407-14-97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Xue J, Niu YF, Huang J, Peng G, Wang LX, Yang YH, et al. miR-141 suppresses the growth and metastasis of HCC cells by targeting E2F3. Tumour Biol. 2014;35(12):12103–12107. doi: 10.1007/s13277-014-2513-9. [DOI] [PubMed] [Google Scholar]
- 40.Cao T, Li H, Hu Y, Ma D, Cai X. miR-144 suppresses the proliferation and metastasis of hepatocellular carcinoma by targeting E2F3. Tumour Biol. 2014;35(11):10759–10764. doi: 10.1007/s13277-014-2017-7. [DOI] [PubMed] [Google Scholar]
- 41.Su J, Wang Q, Liu Y, Zhong M. miR-217 inhibits invasion of hepatocellular carcinoma cells through direct suppression of E2F3. Mol Cell Biochem. 2014;392(1–2):289–296. doi: 10.1007/s11010-014-2039-x. [DOI] [PubMed] [Google Scholar]
- 42.Lee JM, Heo MJ, Lee CG, Yang YM, Kim SG. Increase of miR-199a-5p by protoporphyrin IX, a photocatalyzer, directly inhibits E2F3, sensitizing mesenchymal tumor cells to anti-cancer agents. Oncotarget. 2015;6(6):3918–3931. doi: 10.18632/oncotarget.2928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Lennartsson J, Ronnstrand L. Stem cell factor receptor/c-kit: from basic science to clinical implications. Physiol Rev. 2012;92(4):1619–1649. doi: 10.1152/physrev.00046.2011. [DOI] [PubMed] [Google Scholar]
- 44.Mansuroglu T, Baumhoer D, Dudas J, Haller F, Cameron S, Lorf T, et al. Expression of stem cell factor receptor c-kit in human nontumoral and tumoral hepatic cells. Eur J Gastroenterol Hepatol. 2009;21(10):1206–1211. doi: 10.1097/MEG.0b013e328317f4ef. [DOI] [PubMed] [Google Scholar]
- 45.Yang P, Li QJ, Feng Y, Zhang Y, Markowitz GJ, Ning S, et al. TGF-beta-miR-34a-CCL22 signaling-induced Treg cell recruitment promotes venous metastases of HBV-positive hepatocellular carcinoma. Cancer Cell. 2012;22(3):291–303. doi: 10.1016/j.ccr.2012.07.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Xiao F, Zhang W, Zhou L, Xie H, Xing C, Ding S, et al. microRNA-200a is an independent prognostic factor of hepatocellular carcinoma and induces cell cycle arrest by targeting CDK6. Oncol Rep. 2013;30(5):2203–2210. doi: 10.3892/or.2013.2715. [DOI] [PubMed] [Google Scholar]
- 47.Morell CM, Strazzabosco M. Notch signaling and new therapeutic options in liver disease. J Hepatol. 2014;60(4):885–890. doi: 10.1016/j.jhep.2013.11.028. [DOI] [PubMed] [Google Scholar]
- 48.Wu G, Wilson G, George J, Qiao L. Modulation of notch signaling as a therapeutic approach for liver cancer. Curr Gene Ther. 2015;15(2):171–181. doi: 10.2174/1566523214666141224100319. [DOI] [PubMed] [Google Scholar]
- 49.Xue TC, Zou JH, Chen RX, Cui JF, Tang ZY, Ye SL. Spatial localization of the JAG1/Notch1/osteopontin cascade modulates extrahepatic metastasis in hepatocellular carcinoma. Int J Oncol. 2014;45(5):1883–1890. doi: 10.3892/ijo.2014.2630. [DOI] [PubMed] [Google Scholar]
- 50.Wang M, Xue L, Cao Q, Lin Y, Ding Y, Yang P, et al. Expression of Notch1, Jagged1 and beta-catenin and their clinicopathological significance in hepatocellular carcinoma. Neoplasma. 2009;56(6):533–541. doi: 10.4149/neo_2009_06_533. [DOI] [PubMed] [Google Scholar]
- 51.Sun Q, Wang R, Luo J, Wang P, Xiong S, Liu M, et al. Notch1 promotes hepatitis B virus X protein-induced hepatocarcinogenesis via Wnt/beta-catenin pathway. Int J Oncol. 2014;45(4):1638–1648. doi: 10.3892/ijo.2014.2537. [DOI] [PubMed] [Google Scholar]
- 52.Sun Q, Wang R, Wang Y, Luo J, Wang P, Cheng B. Notch1 is a potential therapeutic target for the treatment of human hepatitis B virus X protein-associated hepatocellular carcinoma. Oncol Rep. 2014;31(2):933–939. doi: 10.3892/or.2013.2917. [DOI] [PubMed] [Google Scholar]
- 53.Zhu P, Wang Y, Du Y, He L, Huang G, Zhang G, et al. C8orf4 negatively regulates self-renewal of liver cancer stem cells via suppression of NOTCH2 signalling. Nat Commun. 2015;6:7122. doi: 10.1038/ncomms8122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Huntzicker EG, Hotzel K, Choy L, Che L, Ross J, Pau G, et al. Differential effects of targeting notch receptors in a mouse model of liver cancer. Hepatology. 2015;61(3):942–952. doi: 10.1002/hep.27566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Khoo KH, Verma CS, Lane DP. Drugging the p53 pathway: understanding the route to clinical efficacy. Nat Rev Drug Discov. 2014;13(3):217–236. doi: 10.1038/nrd4236. [DOI] [PubMed] [Google Scholar]
- 56.Chen L, Zhang R, Li P, Liu Y, Qin K, Fa ZQ, et al. P53-induced microRNA-107 inhibits proliferation of glioma cells and down-regulates the expression of CDK6 and Notch-2. Neurosci Lett. 2013;534:327–332. doi: 10.1016/j.neulet.2012.11.047. [DOI] [PubMed] [Google Scholar]
- 57.Giacoia EG, Miyake M, Lawton A, Goodison S, Rosser CJ. PAI-1 leads to G1-phase cell-cycle progression through cyclin D3/cdk4/6 upregulation. Mol Cancer Res. 2014;12(3):322–334. doi: 10.1158/1541-7786.MCR-13-0543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Han JJ, Xue DW, Han QR, Liang XH, Xie L, Li S, et al. Induction of apoptosis by IGFBP3 overexpression in hepatocellular carcinoma cells. Asian Pac J Cancer Prev. 2014;15(23):10085–10089. doi: 10.7314/APJCP.2014.15.23.10085. [DOI] [PubMed] [Google Scholar]
- 59.Zeng Z, Tu J, Cheng J, Yao M, Wu Y, Huang X, et al. Influence of CCND1 G870A polymorphism on the risk of HBV-related HCC and cyclin D1 splicing variant expression in Chinese population. Tumour Biol. 2015;36(9):6891–900. doi: 10.1007/s13277-015-3401-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Zhu K, Pan Q, Zhang X, Kong LQ, Fan J, Dai Z, et al. MiR-146a enhances angiogenic activity of endothelial cells in hepatocellular carcinoma by promoting PDGFRA expression. Carcinogenesis. 2013;34(9):2071–2079. doi: 10.1093/carcin/bgt160. [DOI] [PubMed] [Google Scholar]
- 61.Xiang Q, Chen W, Ren M, Wang J, Zhang H, Deng DY, et al. Cabozantinib suppresses tumor growth and metastasis in hepatocellular carcinoma by a dual blockade of VEGFR2 and MET. Clin Cancer Res. 2014;20(11):2959–2970. doi: 10.1158/1078-0432.CCR-13-2620. [DOI] [PubMed] [Google Scholar]
- 62.Horwitz E, Stein I, Andreozzi M, Nemeth J, Shoham A, Pappo O, et al. Human and mouse VEGFA-amplified hepatocellular carcinomas are highly sensitive to sorafenib treatment. Cancer Discov. 2014;4(6):730–743. doi: 10.1158/2159-8290.CD-13-0782. [DOI] [PubMed] [Google Scholar]
- 63.Zhou B, Ma R, Si W, Li S, Xu Y, Tu X, et al. MicroRNA-503 targets FGF2 and VEGFA and inhibits tumor angiogenesis and growth. Cancer Lett. 2013;333(2):159–169. doi: 10.1016/j.canlet.2013.01.028. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
GO enrichment analysis of HCC related genes. (XLSX 12 kb)
GO enrichment analysis of miR-34a target genes. (XLSX 14 kb)
Integrative-analysis of miR-34a target genes and the natural language processing (NLP) results. Twenty-four overlapping genes that are not only associated with the development and progression of HCC but also are the potential miR-34a target genes were obtained in this integrative–analysis (XLSX 14 kb)
Data Availability Statement
All data generated or analyzed during this study are included in this published article (Tables 1 and 2) and its Additional files (Additional file 1: Table S1, Additional file 2: Table S2 and Additional file 3: Table S3).