

Abstract
Mitochondria are cellular powerhouses that produce ATP, lipids, and metabolites, as well as regulate calcium homeostasis and cell death. The unique cristae-rich double membrane ultrastructure of this organelle is elegantly arranged to carry out multiple functions by partitioning biomolecules. Mitochondrial ultrastructure is intimately linked with various functions; however, the fine details of these structure-function relationships are only beginning to be described. Here, we demonstrate the application of serial-section electron tomography to elucidate mitochondrial structure in Drosophila indirect flight muscle. Serial-section electron tomography may be adapted to study any cellular structure in three-dimensions.
Keywords: Molecular Biology, Issue 130, electron tomography, serial section, mitochondria, ultrastructure, Drosophila, indirect flight muscle
Introduction
Electron microscopy is a valuable tool to study the structural context of subcellular assemblies and organelles that perform cellular processes. Methods have been developed to preserve the ultrastructure of tissues or cells either by chemical fixation with aldehydes or by high pressure freezing (HPF) followed by freeze substitution (FS)1,2. The embedded specimen blocks may then be sectioned, stained, and observed with a transmission electron microscope (TEM). The HPF specimen could also be processed under cryo-condition, such as by cryo-sectioning or by focused ion beam (FIB) milling, and observed by cryo-EM3,4.
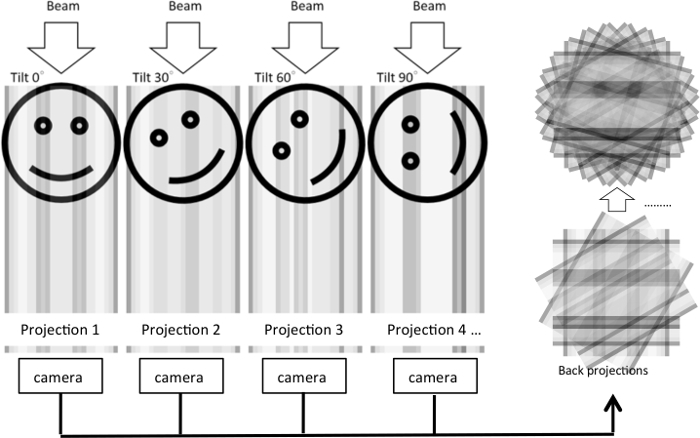
Although thin-section EM provides informative morphological insights, the resulting 2D images can only reveal the ultrastructure of a particular cross-section. How ultrastructure is organized in a 3D volume remains obscured. In order to visualize cellular ultrastructure in three dimensions, an electron tomography method was developed where series of tilt images were acquired and back projected to generate a tomographic reconstruction5 (Figure 1). A double tilt series can be collected by rotating the sample 90° and acquiring a second tilt series. This will minimize missing-wedge artifacts that result from limited sampling angles and improve resolution of the tomogram.
Here, we describe the application of serial-section electron tomography to study mitochondrial ultrastructure of Drosophila indirect flight muscle (IFM)6,7,8,9. In order to obtain 3D reconstructions covering entire mitochondria (approximately 2.5 µm-thick), serial sections were obtained from Drosophila IFM tissue blocks. Tomograms of each section were collected individually using automatic data collection software. Tomographic reconstructions were generated and serial tomograms were joined with the IMOD package to obtain a reconstructed volume of an entire mitochondrion. The joined tomograms were analyzed by 3D software. The densities of mitochondrial cristae were segmented to generate a segmentation model that revealed the organization in three-dimensions.
Protocol
1. Section Drosophila Tissues Using a Vibrating Blade Microtome
Anesthetize Drosophila on ice and immerse each single fly in 1 mL of 4% low melting agarose in phosphate buffer. Allow agarose to solidify on ice. Typically, 4 - 6 flies were processed.
Use a vibrating blade microtome to section agarose gel embedded Drosophila into slices with 100 µm thickness and immerse in fixative solution containing 2.5% glutaraldehyde in 0.1 M phosphate buffer. NOTE: Vibratome sectioning is preferred because tissue architecture remains more intact compared to other methods. Alternatively, dissecting tweezers can be used to dissect IFM into the fixative solution containing 2.5% glutaraldehyde in 0.1 M phosphate buffer.
2. Prepare EM Specimens by the High-pressure Freezing and Freeze Substitution (HPF/FS) Method
Wash tissue sections in 3 drops (~150 µL) of phosphate buffer, followed by 2 drops (~100 µL) of phosphate buffer with 20% BSA. Then place sections in gold carriers for HPF filled with buffer and 20% BSA.
Load the sample containing carriers into a high-pressure freezer according to the user manual.
After freezing, release carriers from the holder under liquid nitrogen and transfer to a freeze-substitution device cooled to -140 °C.
Perform the freeze-substitution protocol as shown in Table 1, with the FS cocktail containing 2% glutaraldehyde, 2% osmium tetroxide, and 0.1% uranyl acetate in acetone.
- Carefully remove the specimens from the carriers with a needle and embed specimens in resin at room temperature. Polymerize the resin at 65 °C for 16 h. NOTE: FS protocols should be modified to prepare other types of samples. HPF/FS is preferred to preserve the ultrastructure and minimize the loss of cellular contents.
- Alternatively, apply a chemical fixation protocol. Fix specimens with 2.5% glutaraldehyde overnight, wash with buffers and then fix with 1% osmium tetroxide for 2 h. Wash and dehydrate with ascending concentrations of ethanol and then infiltrate and embed specimens in Spurr's resin before polymerizing at 65 °C for 16 h.
3. Prepare Serial-sections of the Specimens for Electron Tomography
Trim the specimen blocks to expose the desired block face that contains tissue.
Pretreat gold particles (10 nm in diameter) with 1% BSA for 30 min. Wash and suspend gold particles in PBS buffer. Overlay gold particles on copper slot grids coated with carbon film to create fiducial markers.
Check under TEM to have sufficient fiducial markers (at least 5 - 10 markers) in the field of view of tomography acquisition.
Cut serial sections, 200 - 250 nm in thickness, using an ultramicrotome.
Collect serial sections on slot grids using a perfect loop for thin sections.
Stain the sections with Reynold's lead citrate for 10 min.
Overlay a second layer of fiducial gold particles on the top of the sections.
4. Collect Double-tilt Electron Tomography
Load the grid onto a dual-axis tomography holder and insert into the transmission electron microscope operating at 200 kV.
Align the microscope at the eucentric focus.
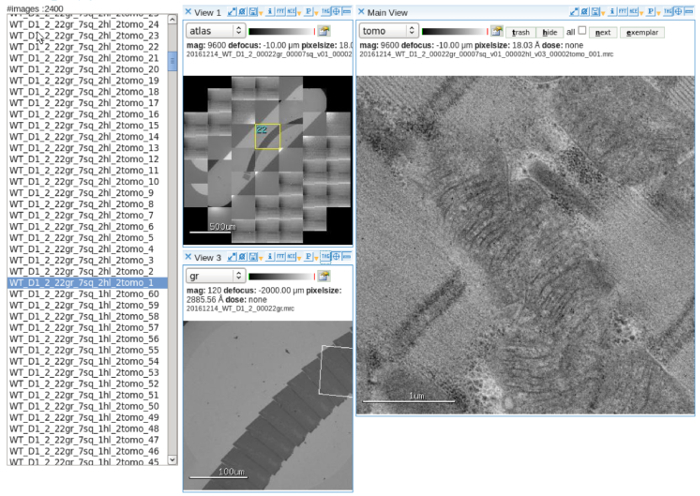
Set up the automatic data collection software. Adjust and align the electron beam at the multi-scale imaging setting. Refer to user manual for operation detail10 (Figure 2).
Acquire camera dark and bright references in an empty area without carbon film under the tomography collection setting.
Collect a grid atlas at low magnification. Select mitochondria on serial sections as targets for tomography collection.
Acquire a tilt series from -60° to +60° with 2° increments on axis-A for each target. NOTE: The tilt angles will be mechanically constrained by the design of the specimen holder. The holder will block the beam at high tilt angles.
To collect the second tilt series, rotate the specimen holder 90°. Acquire a new atlas. Select corresponding positions and acquire the tilt series on axis-B for each target. NOTE: Other software packages, such as SerialEM and Xplore3D, are available for automatic data collection.
5. Reconstruct 3D Tomograms and Segment Sub-volumes Using Software
- Reconstruct tomograms of dual-axis tilt images using IMOD software11.
- Refer to user manual for operation detail.
- Align the individual tilt series by the positions of gold fiducial markers. Reconstruct tomograms either by the Back Projection method or by the Simultaneous Iterative Reconstruction Technique (SIRT) method for both axis-A and axis-B, respectively (Figure 3).
- Combine tomograms of axis-A and axis-B to generate a double-tilt tomogram with reduced missing wedge artifact.
- Join together double-tilt tomograms of serial sections to obtain reconstructions covering entire mitochondrion volume. Model the gaps between serial sections by the IMOD program.
- Trim the joined tomograms and bin to a desired size for volume segmentation.
- Analyze joined serial tomograms using 3D software.
- Refer to user manual for operation detail.
- Filter the tomograms with a Gaussian filter (or other desired method) to improve the contrast of features and reduce background densities.
- Segment the ultrastructure of mitochondria either manually or automatically.
- Display segmentation models to allow inspection in 3D.
- Generate movies using tools available in 3D.

Representative Results
We applied serial-section electron tomography to analyze structural features of mitochondrial cristae that reflects its energetic state and aging. We showed that mitochondria of Drosophila IFM form an integrated cristae and matrix network in 3D (Figure 4)7. In addition, mutant flies with mitochondrial DNA replication defect and an accelerated aging phenotype accumulated mitochondria that contained subsections of onion-like swirling core (Figure 5)7.
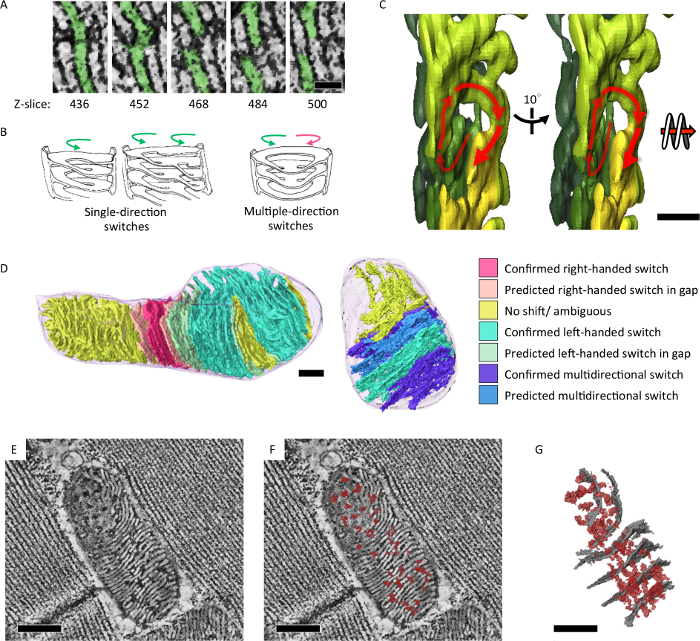
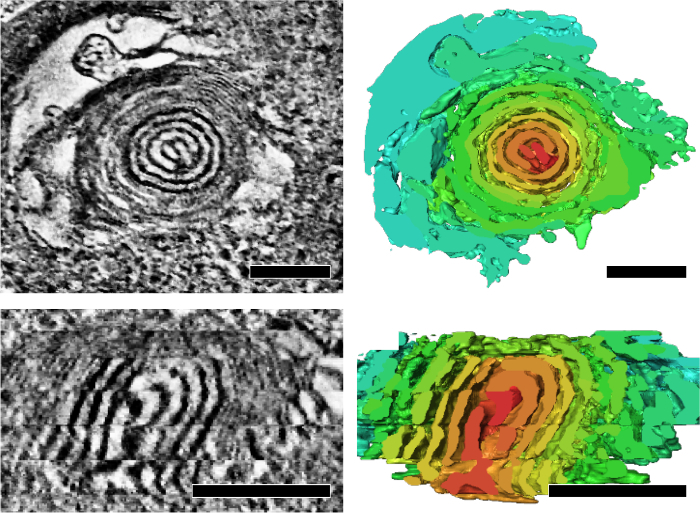
Figure 1: Illustration of electron tomography reconstruction from a tilt series. In the experiment, an electron beam is passed through a 3D object as the object is tilted to various degrees. For each tilt condition, a 2D projection is generated and captured with a camera. The 2D projections are then back projected to reconstruct the 3D object based on the central slice theorem. In the illustration, the same process is shown for an original 2D image that is projected into a single dimension under various tilt angles. The 1D projections are then used to reconstruct the 2D image. Please click here to view a larger version of this figure.
Figure 2: Automatic data collection. An image viewer display of software showing the atlas of a slot grid containing serial sections, multi-scale targeting of mitochondria and acquired tilt images. Scale bar = 500 µm (left top panel), 100 µm (left bottom panel), 1 µm (right panel). Please click here to view a larger version of this figure.
Figure 3: Serial-section electron tomography of a single mitochondrion in Drosophila indirect flight muscle. (A) 2D micrographs and (B) 3D tomograms from serial sections covering the entire volume of a mitochondrion. (C) The joined serial-section tomograms are projected to create a longitudinal section, with the z-axis shown vertically. Notably, tissue sectioning led to loss of material, leaving gaps between joined tomograms. Scale bar = 500 nm. Please click here to view a larger version of this figure.
Figure 4: Integrated intra-mitochondria cristae and matrix network revealed in 3D. (A) Slices of mitochondrial electron tomographic reconstructions showing switches between lamellar membranes through the z-axis. (B) Illustrations of the observed cristae switching patterns of right handed and/or left-handed spirals. (C) Tomographic segmentation illustrating a left-handed spiral in 3D (D) Cristae switching patterns were analyzed and color-rendered on the segmentation model (E, F). Tomographic slice showing lateral matrix confluency (dark densities, marked red) across cristae membranes (white densities). (G) Segmentation model of the tomogram in (E) showing lateral matrix confluency (in red) and representative cristae (in gray). Scale bar = 50 nm (A), 200 nm (C), and 300 nm (D, E, F, G). The figure was the reprint from Jiang et al.7Please click here to view a larger version of this figure.
Figure 5: Mitochondria with onion-like swirling cores accumulated in flies with mitochondrial DNA replication defects during aging. Representative tomographic slices and the corresponding segmentation (right panels) showing a cross-sectional view (top) and a longitudinal-section view (bottom) of a swirling core. Volume segmentation is indicated by arbitrary color rendering to highlight the swirling core. Scale bar: 200 nm. The figure was the reprint from Jiang et al.7Please click here to view a larger version of this figure.
| Step | Temp | Time | Solution |
| 1 | -140 °C to -9 0°C | 30 min | Liquid nitrogen |
| 2 | -90 °C | 96 hr | FS cocktail |
| 3 | -90 °C to -60 °C | 6 hr (5 °C/hr) | FS cocktail |
| 4 | -60 °C | 12 hr | FS cocktail |
| 5 | -60 °C to -25 °C | 7 hr (5 °C/hr) | FS cocktail |
| 6 | -25 °C | 12 hr | FS cocktail |
| 7 | -25 °C to 0 °C | 5 hr (5 °C/hr) | FS cocktail |
| 8 | 0 °C | 1 hr x 3 times | Acetone |
| 9 | Room temp | Resin infiltration |
Table 1: Freeze-substitution protocol.
Discussion
In this protocol, we describe an optimized workflow for applying serial-section electron tomography to study 3D mitochondrial ultrastructure of Drosophila indirect flight muscle. Preservation of ultrastructure in the sample is the primary technical challenge for this type of analysis. In order to best preserve ultrastructure two methodological steps were included. First, tissue was sampled by sectioning with vibrating blade microtome in order to maintain the tissue architecture as much as possible. Second, a HPF/FS protocol was optimized to preserve organelle ultrastructure during the preparation of embedded specimen blocks. Specimens were frozen under high pressure, which lowers the freezing point of water and reduces the formation of ice crystals that damage ultrastructure1. Specimens as thick as 0.1 mm can be vitrified instantaneously, and then subjected to freeze substitution to generate specimen blocks for EM analysis. The improved preservation of ultrastructure by HPF/FS was noted, when compared to chemical fixation methods. Using this series of steps for sample preparation, preservation of mitochondrial double membranes and cristae membranes was dramatically improved.
Obtaining serial sections of the specimen is the most challenging step of the method. As the electron beam has limited penetration power, the thickness of the section is restricted to either 250 nm or 500 nm using a TEM operating at 200 kV or 300 kV, respectively. Because the thickness of a mitochondrion may be more than 2 µm, serial sections are required to obtain full volume reconstructions. However, recovering a sufficient number of serial sections to span an entire organelle is a technical challenge. Trimming the block face to be as flat as possible between the bases of the trapezoid can allow a slot grid to accommodate more sections and thus cover larger volumes. Additionally, using a perfect loop for thin sections increases the success rate of transferring the serial sections to the grid.
Serial-section electron tomography can be accomplished with standard EM core equipment. However, the method has some inevitable limitations that arise from technical constraints. One is that material is inevitably lost between serial sections, leaving gaps in the joined reconstruction. Second is the missing wedge artifact, which arises due to the limited tilt angles that are achievable. This restriction occurs because the specimen holder cannot be turned in a full rotation without blocking the electron beam. Despite these limitations, serial-section electron tomography provides sufficient resolution to reveal cellular and organelle ultrastructure in 3D.
For smaller scale imaging, cryo-electron tomography is an emerging technology that can be used to obtain the structure of macromolecular complexes and assemblies in situ at nm or sub-angstrom resolution in combination with sub-tomographic reconstruction3. In this application, cells are thinned by sectioning or by focused ion beam milling under liquid nitrogen. The tomograms are collected under cryo-conditions where molecular structures are preserved close to the native state without chemical fixation, dehydration, or embedding. At the other end of scale, to analyze large tissue volumes at the expense of resolution, serial block-face scanning electron microscopy is an appealing modality even though it requires a specific instrument4.
Disclosures
The authors declare that they have no competing financial interests.
Acknowledgments
The studies were performed in the EM core at the Institute of Cellular and Organismic Biology and the cryo-EM core of Academia Sinica, Taipei, Taiwan. The work was supported by Academia Sinica and MOST.
References
- Dahl R, Staehelin LA. High-pressure freezing for the preservation of biological structure: theory and practice. J Electron Microsc Tech. 1989;13(3):165–174. doi: 10.1002/jemt.1060130305. [DOI] [PubMed] [Google Scholar]
- Sabatini DD, Bensch K, Barrnett RJ. Cytochemistry and electron microscopy. The preservation of cellular ultrastructure and enzymatic activity by aldehyde fixation. J Cell Biol. 1963;17:19–58. doi: 10.1083/jcb.17.1.19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rigort A, et al. Micromachining tools and correlative approaches for cellular cryo-electron tomography. J Struct Biol. 2010;172(2):169–179. doi: 10.1016/j.jsb.2010.02.011. [DOI] [PubMed] [Google Scholar]
- Denk W, Horstmann H. Serial block-face scanning electron microscopy to reconstruct three-dimensional tissue nanostructure. PLoS Biol. 2004;2(11):e329. doi: 10.1371/journal.pbio.0020329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lucic V, Forster F, Baumeister W. Structural studies by electron tomography: from cells to molecules. Annu Rev Biochem. 2005;74:833–865. doi: 10.1146/annurev.biochem.73.011303.074112. [DOI] [PubMed] [Google Scholar]
- Soto GE, et al. Serial section electron tomography: a method for three-dimensional reconstruction of large structures. Neuroimage. 1994;1(3):230–243. doi: 10.1006/nimg.1994.1008. [DOI] [PubMed] [Google Scholar]
- Jiang YF, et al. Electron tomographic analysis reveals ultrastructural features of mitochondrial cristae architecture which reflect energetic state and aging. SciRep. 2017;7:45474. doi: 10.1038/srep45474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cogliati S, Enriquez JA, Scorrano L. Mitochondrial Cristae: Where Beauty Meets Functionality. TrendsBiochemSci. 2016;41(3):261–273. doi: 10.1016/j.tibs.2016.01.001. [DOI] [PubMed] [Google Scholar]
- Friedman JR, Nunnari J. Mitochondrial form and function. Nature. 2014;505(7483):335–343. doi: 10.1038/nature12985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suloway C, et al. Fully automated, sequential tilt-series acquisition with Leginon. J Struct Biol. 2009;167(1):11–18. doi: 10.1016/j.jsb.2009.03.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mastronarde DN, Held SR. Automated tilt series alignment and tomographic reconstruction in IMOD. J Struct Biol. 2017;197(2):102–113. doi: 10.1016/j.jsb.2016.07.011. [DOI] [PMC free article] [PubMed] [Google Scholar]