

Abstract
Background
Photosynthetic systems are known to be sensitive to high temperature stress. To maintain a relatively “normal” level of photosynthetic activities, plants employ a variety of adaptive mechanisms in response to environmental temperature fluctuations. Previously, we reported that the chloroplast-targeted AtFtsH11 protease played an essential role for Arabidopsis plants to survive at high temperatures and to maintain normal photosynthetic efficiency at moderately elevated temperature. To investigate the factors contributing to the photosynthetic changes in FtsH11 mutant, we performed detailed chlorophyll fluorescence analyses of dark-adapted mutant plants and compared them to Col-0 WT plants under normal, two moderate high temperatures, and a high light conditions.
Results
We found that mutation of FtsH11 gene caused significant decreases in photosynthetic efficiency of photosystems when environmental temperature raised above optimal. Under moderately high temperatures, the FtsH11 mutant showed significant 1) decreases in electron transfer rates of photosystem II (PSII) and photosystem I (PSI), 2) decreases in photosynthetic capabilities of PSII and PSI, 3) increases in non-photochemical quenching, and a host of other chlorophyll fluorescence parameter changes. We also found that the degrees of these negative changes for utilizing the absorbed light energy for photosynthesis in FtsH11 mutant were correlated with the level and duration of the heat treatments. For plants grown under normal temperature and subjected to the high light treatment, no significant difference in chlorophyll fluorescence parameters was found between the FtsH11 mutant and Col-0 WT plants.
Conclusions
The results of this study show that AtFtsH11 is essential for normal photosynthetic function under moderately elevated temperatures. The results also suggest that the network mediated by AtFtsH11 protease plays critical roles for maintaining the thermostability and possibly structural integrity of both photosystems under elevated temperatures. Elucidating the underlying mechanisms of FtsH11 protease in photosystems may lead to improvement of photosynthetic efficiency under heat stress conditions, hence, plant productivity.
Electronic supplementary material
The online version of this article (10.1186/s12870-018-1228-2) contains supplementary material, which is available to authorized users.
Keywords: Heat stress, Thermotolerance, Photosynthesis; Thermostability, FtsH11, Chlorophyll fluorescence, High temperature
Background
Photosynthesis is the primary source of raw materials (carbohydrates, amino acids, fatty acids, etc.) for all forms of agricultural products. In nature, plants are subject to daily and seasonal temperature fluctuations in their growing environments. Therefore, photosynthetic systems are under frequent influence of temperature changes. Studies have shown that chloroplast membranes, especially thylakoid membranes and associated photosynthetic apparatus, are extremely sensitive to high temperature (heat stress) and are thought to be the primary targets of heat stress [1–4]. High temperature extremes have profound negative impacts on essentially all aspects of photosynthetic activities, from changes in enzyme activities and photochemical reactions, damages to photosystem II proteins, alternations in membrane fluidity and photosystem protein complexes, to the irreversible destruction of the entire PSII [2–7]. As the trend of global temperature increases [8, 9], the negative effects of heat stress on photosynthesis, hence on world agriculture production and food security are expected to exacerbate [10–13]. Understanding the basic mechanisms by which photosynthetic systems in plants respond and adapt to temperature upshifts in their environment is a prerequisite for any attempt to enhance or even maintain efficient photosynthesis under heat stress conditions. However, comparing to the studies on functions of FtsH proteases under light stress [14, 15], very little is known on how plants sense the temperature changes and regulate those essential responses to maintain relatively “normal” photosynthetic activities under moderate high temperatures.
Previously, we reported that AtFtsH11, a homolog of a filamentous temperature sensitive H protease (FtsH), plays an essential role in thermotolerance of Arabidopsis plants [16]. Mutations of AtFtsH11 cause reduction in photosynthetic efficiency under moderately elevated temperature of 30 °C. The FtsH proteases are a class of membrane-bound, ATP-dependent zinc-metalloprotease found in all eukaryotic organisms examined so far [17, 18]. In prokaryotic and yeast systems, FtsH proteases are essential to microbial survival at high temperature and are also involved in protection against other environmental stresses [19–21]. These proteases function as molecular chaperones and help organisms to maintain cell homeostasis under stress conditions [22–24]. Plant genomes contain multiple FtsH homolog genes, with most of them predicted to be either chloroplast or mitochondria targeted [15, 25, 26]. In Arabidopsis, major chloroplast targeted FtsH proteases (FtsH1/5, FtsH2/8, FtsH6 and FtsH7/9) have been shown to function in alleviating and repairing high light induced photodamages to photosystems (PS), especially in the turnover of photodamaged D1 protein in PSII [14, 25, 27–34]. The FtsH11 is the first plant FtsH protease reported to be involved in thermotolerance in plants. Recently, a role of FtsH6 in the regulation of HSP21 level has also be proposed [35].
Chlorophyll (Chl) fluorescence measurements have been widely used to examine the impacts of a variety of abiotic stresses on photosynthetic systems in plants [36–40]. The effects of high temperatures on photosynthetic activity and apparatus have been evaluated by both slow and fast Chl fluorescence kinetic measurements in many plant species, such as Arabidopsis, wheat and beans [41–44]. High temperature induced reduction in the efficiency of PSII photochemistry and PS1 functions have been revealed using the Chl fluorescence measurements [39, 41, 44]. Moreover, with the improved technology, Chl fluorescence has become a powerful tool for detailed analysis of functional and structural changes of PS under a variety environmental stresses and in characterization of heat stress tolerance of plant varieties [37, 38, 40, 42]. In Arabidopsis FtsH11 mutants, the decrease in thermotolerance appears to be associated with decreased photosynthetic efficiency (quantum yield) above 30 °C [16]. However, which specific photosynthetic factors contribute to such decreases, and exactly how mutation of FtsH11 altered their normal functions are still unknown and need to be investigated. In this study, we applied the Chl florescence approach to examine the effects of FtsH11 mutation on a range of photosynthetic parameters under normal and moderately high temperature conditions in Col-0 and Ftsh11 mutant plants. We identified the major Chl fluorescence parameters that were affected by Ftsh11 mutations and how their changes contributed to the detected decreases in PSII efficiency in FtsH11 mutant under moderate high temperatures. The study also revealed that mutations in FtsH11 gene had much broader negative impact on photosynthetic systems in plants.
Methods
Plant growth conditions
Arabidopsis (Arabidopsis thaliana) Ftsh11 knockout mutant salk033047 and the corresponding Col-0 wild type (WT) used in this study for all Chl fluorescence measurements were obtained from the Arabidopsis Biological Resource Center (https://abrc.osu.edu). Arabidopsis seeds were suspended in 0.1% agar solution and cold-treated at 4 °C for 2 days. The vernalized seeds were sowed on well-moistened Sunshine 3 potting soil (Sun Gro Horticulture, Bellevue, WA) in Arabaskets placed in an Araflat (Lehle Seeds, Round Rock, TX). Each Araflat contained half Col-0 and half salk033047 plants. The Araflats were covered with clear plastic covers for 5 day after planting to retain soil moisture and allow seed to germinate. Arabidopsis seedlings were thinned to 1 plant per Arabasket at 2 true-leaf stage and grown in a Reach-In Conviron growth chamber (E-15 model, Conviron, Winnipeg, Canada) for 3 weeks under optimal conditions (21 °C, 12 h/12 h d/n at 70 μmol quanta m−2 s−1). The plants were fertilized weekly with diluted Miracle-Gro nutrient (Scotts Miracle-Gro Products, Port Washington, NY). A total of six Araflats were planted for each experiments and the flats were rotated weekly.
Treatments
For Chl fluorescence measurements, the Arabaskets with 21d–old, uniformly grown, WT and salk033047 plants were selected and randomly rearranged into 4 treatment groups. 1) 21 °C control with normal light intensity (70 μmol quanta m−2 s−1), 2) 30 °C moderately elevated temperature with normal light intensity, 3) 35 °C elevated temperature with normal light intensity, and 4) 21 °C with high light (HL) intensity (400 μmol quanta m−2 s−1). The chambers used for the treatments were the same as the E15 Conviron model for growing the plants. Light intensity in each chamber was adjusted to the aforementioned levels using a light meter, and temperature settings in all chambers were checked with a thermocouple. The plants in groups 2 and 3 were placed into the heat treatment chambers with temperatures set at 30 °C or 35 °C, respectively, and light intensity was the same as that of optimal control conditions. The plants in HL treatment group 4 were placed to a 21 °C chambers with light intensity set at 400 μmol quanta m−2 s−1 whereas plants in group 1 remained in the original chamber. The photoperiod for all groups was the same 12 h/12 h light/dark cycle.
Chlorophyll fluorescence measurements
The most recent fully-expanded intact rosette leaf from each Arabidopsis plant was used for Chl fluorescence measurements after 2 h dark-adaption. Dark treatment was applied by placing the enclosed leaf clips on to the selected rosette leaves. The fluorescence slow induction curve (IC) and the light response curve (LC) of the intact leaves were measured [45] at the onset of the treatment (0 day) and then daily for 5 days (1, 2, 3, 4, and 5 days) using a pulse amplitude modulation fluorometer equipped with a dual wavelength emitter detector unit (DualPAM 100, Walz, Effeltrich, FRG). At the set treatment times, the 2 h dark-adapted leaf together with the enclosed leaf clip on was carefully removed from base of the petiole and then immediately mounted on the measuring head of DualPam100 system under dim light. At the beginning of each measurement, actinic light (20 Hz) was provided to measure the minimal (Fo) and maximal (Fm) Chl fluorescence of the dark-adapted leaf tissues and to determine the maximum quantum efficiency [Fv/Fm = (Fm-Fo)/Fm] of PSII photochemistry. The Fv/Fm was automatically calculated where Fv represents the maximum variable fluorescence (Fv = Fm-Fo). Forty seconds after the onset of actinic light, a saturating pulse (SP) was given in 20 s intervals over a 5-min period for IC measurements. The SP had an intensity of 6000μE m−2.s−1 at an emission peak of 660 nm and pulsed for 1 s. For the LC measurement, the red actinic light was automatically applied in a stepwise increasing in photosynthetically active radiation (PAR) from 0 to 8, 15, 24, 55, 97, 128, 218, 341, 533 and 827 in 30 s intervals over a 5-min period using a DualPam100 system. Fluorescence parameter changes (response curves) of the leaf tissues in response to the SP (IC) and light intensity increases (LC) were recorded over each measurement [46]. The changes of the P700 redox state of PSI were recorded during the IC at an absorbance wavelength of 830 nm. For each treatment, leaves from 6 to 8 plants were used for daily Chl fluorescence measurements. The experiment was repeated 4 times. Information about the Chl fluorescence parameters reported here and equations for their calculation are provided as the supplementary material (Additional file 1: Table S1).
Results
The maximum quantum efficiency of PSII (Fv/Fm) decreased significantly in FtsH11 under elevated temperatures
Under normal 21 °C growing temperature, no significant difference was detected in the measured Chl fluorescence parameters between the dark-adapted Ftsh11 mutant salk033047 plants and WT Col-0 plants (Figs. 1 and 2, 0d). At 21 °C, Fv/Fm of salk033047 and Col-0 plants were 0.769 and 0.752, respectively (Fig. 1a, 0 d). Highlight (400 μmol quanta m−2 s−1) at 21 °C had no significant effect on Fv/Fm in both Ftsh11 mutant and WT plants (Fig. 2a). However, after subjecting to elevated temperatures of 30 °C and 35 °C, the Fv/Fm of mutant plants decreased rapidly over the course of high temperature treatments (Figs. 1 and 2a). At 30 °C, the Fv/Fm of salk033047 decreased from 0.752 at 0d (21 °C) to 0.639, 0.524, 0.426, 0.297 and 0.214 after 1d, 2d, 3d, 4d, and 5d of heat exposure, respectively (Fig. 1a). The average reduction of Fv/Fm was −0.109 per day with a R2 of 0.998. At 35 °C, Fv/Fm of the mutant plants decreased from 0.749 (0d) to 0.492 after only 1d of heat exposure and it plummeted to 0.145 at the end of 2d treatment (Fig. 2a). Therefore, the 35 °C treatment was terminated after 2 days. The average reduction of Fv/Fm was −0.302 per day in 35 °C-treated salk033047 plants with a R2 of 0.993, much faster than that occurred at 30 °C.
Fig. 1.
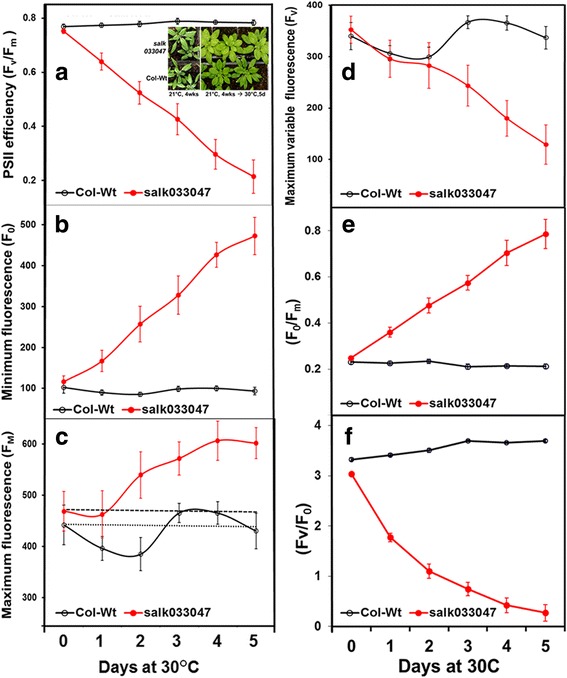
Changes in chlorophyll fluorescence in dark-adapted leaves under moderately elevated temperature at 30 °C. a the maximal PSII efficiency (Fv/Fm), (b) maximal Chl fluorescence (Fm), (c) minimal Chl fluorescence (Fo), (d) maximal variable Chl fluorescence (Fv), (e) the correlation of Fm and Fv with Fo and (f) the daily Fv/Fo ratio changes in Col WT and (salk033047) FtsH11 mutant over a 5 day 30 °C-treatment. The first data point in (a) and the data point at the 0d represent the data collected from 21 °C-grown plants prior to the onset of heat treatment. Three-week old Col WT and FtsH11 mutant (salk033047) plants grown at 21 °C 70 μmol with 12 h/12 h d/n cycle were transferred to a 30 °C growth chamber. Changes in dark-adapted chlorophyll fluorescence were determined daily for five days. For each sampling point, the most recently matured leaves from 6 to 8 plants were used. The experiments were repeated 4 times. Each data point represents mean ± SD
Fig. 2.
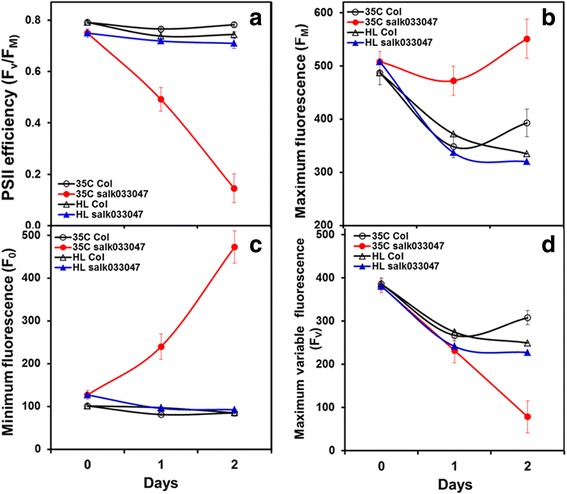
Changes in chlorophyll fluorescence of dark adapted leaves of Col WT and FtsH11 mutant (salk033047) under moderately high temperature at 35 °C or a 2d high light treatment (HL, 400 μmol quanta m−2 s−1) at normal temperature (21 °C). a the Fv/Fm, (b) Fm, (c) Fo, and (d) Fv values in rosette leaves were measured daily after 2 h of dark adaption
Changes in individual Chl fluorescence parameters in FtsH11 under elevated temperature conditions
As indicated by the way the Fv/Fm was calculated [(Fm – Fo)/Fm], the decreases in Fv/Fm in heat-treated FtsH11 mutant could be a result from either an increase in Fo or a decrease in Fm or a combination of both (decrease in maximal variable fluorescence Fv). To determine the cause for Fv/Fm decrease in salk033047 and its association with Ftsh11 mutation, we examined the kinetic responses of each individual Chl fluorescence parameter to the applied heat treatments in mutant plants and compared with those in WT under the same conditions.
Upon exposure to moderately heat treatments, Fo of salk033047 plants displayed a dramatic increase while it showed no significant change in Col-0 plants under both elevated temperatures. After only 1 day of exposure to the 30 °C treatment, the Fo of salk033047 plants increased substantially (Fig. 1b). The Fo of mutant plants increased continually with heat exposure time, whereas Fo of WT plants showed little change under the same treatment (Fig. 1b). The rate of Fo increase in 30 °C-treated mutant plants averaged 75.2 per day with a R2 of 0.992. The average Fo in 5d, 30 °C-treated mutant plants increased more than 4-fold from 116 to 472 (Fig. 1b). The increase in Fo accelerated further after exposing mutant plants to a 35 °C heat treatment, at a rate (172.8/day) that was twice of 30 °C-treated mutant plants (Fig. 2b). The Fo of 2d, 35 °C-treated salk033047 plants exceeded more than 4-folds of 21 °C-controls (0d). The differences of Fo levels were highly significant not only between mutant and WT plants that underwent the same treatment and between control and heat-treated mutant plants, but also between the 30 °C and 35 °C treated mutant plants (Fig. 1 and 2b).
Adjustments for Fm were observed in both WT and mutant plants within the first day exposure to either moderately elevated temperatures or HL treatment (Figs. 1 and 2c). Fm displayed an initial decrease followed by an increase during the course of heat treatments. Although the kinetic change of Fm in mutant plants resembled those exhibited by WT in response to the heat treatments, the degrees of initial decrease in heat-treated salk033047 was considerable less than those seen in WT plants (Fig. 1 and 2c). The Fm levels of heat-treated mutant plants were also much higher than those of WT plants. The Fm of 2d and 4d 30 °C-treated mutant plants were 539 and 606 respectively comparing to 385 and 465 in heat-treated WT (Fig. 1b). The differences in Fm between mutant and WT leaves became significant after 2 days of 30 °C treatment (Fig. 1c). The difference in Fm was widened further between mutant and WT plants under 35 °C condition (Fig. 2c).
Fv is the difference between Fm and Fo and represent the maximum variable fluorescence for photochemical quenching of PSII. After the onset of the heat treatments, the Fv of salk033047 plants showed a steady decrease (Figs. 1 and 2d), despite of the detected Fm increases (Fig. 1 and 2c). The Fv of 30 °C-treated salk033047 plants showed a clear negative correlation with Fo (Fig. 1e). The similar correlation relationship was also observed in 35 °C-treated mutant plants (data not shown). The moderate increases in Fmdetected in heat-treated mutant plants (Fig. 1b) was simply unable to compensate for the effect of heat induced Fo increase for Fv (Fig. 1e). Therefore, the decreases in Fv of salk033047 plants resulted solely from the drastic increases in Fo occurred under heat treatment conditions (Fig. 1f). Consequently, the heat induced increases in Fo primarily resulted in the significant decrease in Fv/Fm in heat-treated FtsH11 mutant plants (Figs. 1 and 2).
The FtsH11 mutant displayed similar Chl fluorescence as the WT in response to high light stress at 21 °C
To determine if the Ftsh11 mutation also affects the response of Arabidopsis to HL stress, we measured the Chl fluorescence yields of salk033047 and Col-0 under a light intensity about five folds higher than optimal level at normal 21 °C. We found that, in response to a light intensity increase from 70 μmol quanta m−2 s−1 to 400 μmol quanta m−2 s−1, the Fm of salk033047 and Col-0 mutant plants decreased within the first day of HL exposure but exhibited no changes in Fo (Fig. 2). The Fv changes in HL-stressed mutant and WT basically followed the same pattern of Fm changes (Fig. 2c and d). As a result, no HL induced net decreases in Fv/Fm were detected in either plant line. Overall, the HL-stressed salk033047 and Col-0 plants showed no significant differences in Fo Fm Fm, and Fv/Fm over the period of 2d treatment (Fig. 2), indicating that mutation of Ftsh11 did not affect the adaption processes of salk033047 leaves to high light stress in Arabidopsis under normal temperature.
The electron transfer rates (ETR) of PSII and PSI decreased significantly in Ftsh11 mutant at 30 °C
The IC and LC measurements were used to further dissect the effects of FtsH11 mutations on photosynthetic systems under moderate heat stress. Significant differences in a range of measured parameters were detected between the heat-treated salk033047 and Col-0 plants and between heat-treated and 21 °C-control salk033047 plants. No differences in the measured parameters were detected between control salk033047 and Col-0 and between heat-treated and control WT plants. The LC of ERT(II), qN and Y(II) and the Fm, maximal levels of ERT(II) and qN of PSII obtained in the IC in control (0d) and 2d, and 5d 30 °C-treated salk033047 and control and 5d 30 °C Col-0 plants are presented in Fig. 3. The ETR(I) and maximal P700 of PSI (Pm) are presented in Fig. 4. The kinetic responses of major Chl fluorescence parameters for PSII and PSI in dark-adapted plants obtained from IC are presented in Additional file 2: Figure S1. A table summarizing the responses of all parameters of IC and LC is provided as supplemental materials (Additional file 1: Table S1).
Fig. 3.
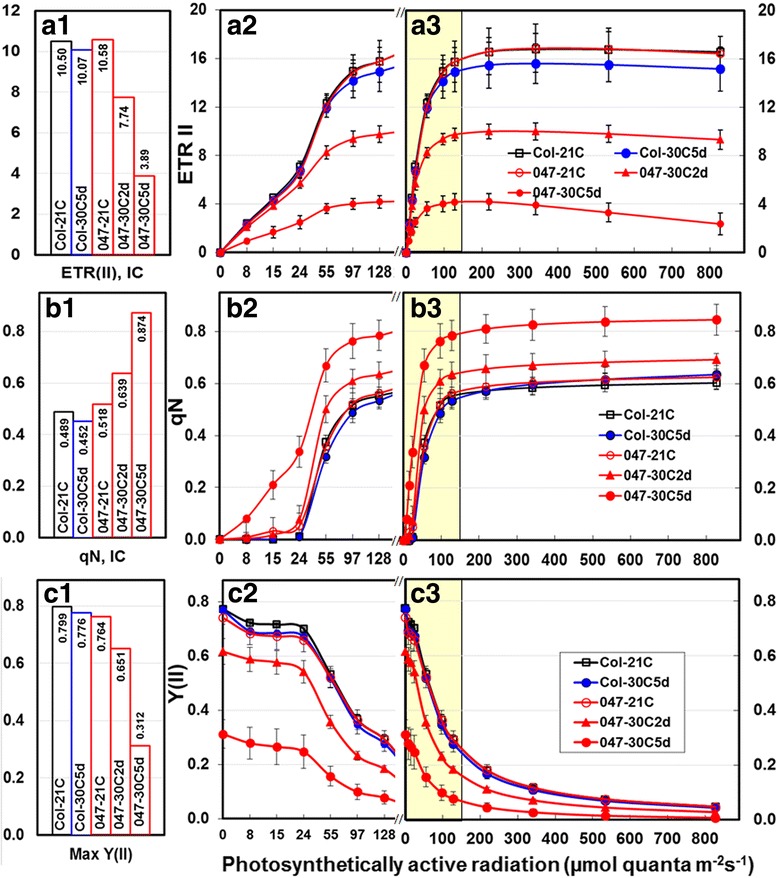
Light response curve measured in light adapted leaves in 21 °C, and 30 °C5d Col WT and in 21 °C, 30 °C2d and 30 °C5d treated FtsH11 mutant (047) plants. a Electron transfer rate of PSII, ETR(II); (b) Coefficient of nonphotochemical quenching of PSII, qN; (c) Effective quantum yield of PSII, Y(II). The maximal levels of the corresponding parameters (number inside the graph) obtained in IC measurement are presented here as fig. a1, b1 and c1. The X axis for fig. a2, b2 and c2 was plotted with even interval for the first six low light intensities measurement to provide readers a better view for the data points at lower light intensity range. The corresponding data points were shaded in fig. a3, b3, and c3. For each sampling point, 6–8 plants were used. The experiments were repeated 4 times. Each data point represents mean ± SD. Here, the mutant salk033047 was written as 047 due to space restraint
Fig. 4.
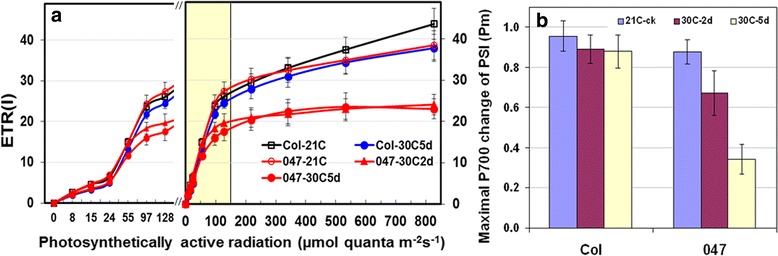
Light response curve of electron transfer rate of PSI, ETR(I) (a) and maximal P700 of PSI, Pm (b) measured in light adapted Col WT and FtsH11 mutant (047) plants at normal 21 °C, 2d and 5d of 30 °C treatment
In 30 °C-treated salk033047, significant decreases in ETR (II) were detected at all light intensity applied in 5d–treated plants and at 55 PAR and above light intensity in 2d–treated plants (Fig. 3a). At the onset of light induction 8 PAR, the ETR(II)s were 2.08 and 0.94 in 2d and 5d–treated mutant comparing to the 2.24 and 2.41 measured in 21 °C-contrl mutant and 5d–treated WT. Such ETR(II) difference expanded as the applied light intensity increased over the course of LC measurement. The levels of heat induced ETR(II) decreases in salk033047 plants also showed an apparent correlation with the heat exposure times. The ETR(II)s in 5d–treated salk033047 were not only significantly lower than those of heat-treated Col-0 plants, but also were significantly lower than those of 2d–treated mutant plants (Fig. 3a). At 97 PAR, the average ETR(II)s in 2d and 5d treated salk033047 plants were 9.38, 4.01 respectively, comparing the 14.17 in 5d–treated Col-0. At 827 PAR, ETR(II)s changed to 9.33, 2.34 and 15.17 in these corresponding plants. The IC measurements detected similar ETR(II) changes in heat-stressed FtsH11 mutant (Additional file 2: Figure S1). At the end of IC illuminations, ETR(II)s of 2d and 5d treated salk033047 were 7.74, 3.89, respectively, comparing 10.58 in control mutant and 10.07 in 5d treated WT (Fig. 3a).
The response of ETR(I) of PSI in FtsH11 mutant to the 30 °C heat treatment were similar to those of ETR(II), except that the exposure to 30 °C beyond the 2nd day did not induce further decrease in ETR(I) of the mutant plants (Fig. 4a). The results indicate that mutations of FtsH11 in Arabidopsis reduce the electron transfer efficiency at elevated temperatures.
Nonphotochemical quenching (NPQ) increased significantly in Ftsh11 mutant at 30 °C
The coefficient of NPQ, qN, was used to evaluate the effects of FtsH11 mutation on the efficiency of PSII in the energy dissipation in Arabidopsis chloroplast. The higher the qN, the higher proportion of the absorbed light energy being dissipated (quenched) into heat, indicates the less efficiency of PSII in the use of excitation energy for photochemical reaction. In both IC and LC measurements, we detected increases in qNs in the FtsH11 mutant after subjecting the plants to the 30 °C treatment (Fig. 3b, Additional file 2: Figure S1). Opposite to what occurred in the 5d–treated WT (a slight delayed response) the qNs in 5d–treated mutant plants raised much fast after the onset of induction and were substantially higher than those of WT plant as well as those of 21 °C-grown mutant plants (Fig. 3b). The qNs of 5d–treated mutant plants were also significantly higher than those of 2d, treated mutant plants, indicating a positive correlation between the qN increases of mutant plants and the heat treatment times. At 827 PAR, the measured qNs in 2d and 5d treated salk033047 plants were 0.692 and 0.845, respectively comparing 0.625 in control mutant and 0.635 of 5d–treated Col-0 plants (Fig. 3b). The highest qN obtained in IC were from either 5th or 6th SP, with relative values of 0.639, 0.874, 0.518 and 0.452 for the corresponding plants (Fig. 3b1).
The effective quantum yield of PSII (YII) and maximal P700 of PSI (Pm) decreased significantly in Ftsh11 mutant under moderately elevated temperatures
Both IC and LC measurements detected reductions of YII in 30 °C-treated FtsH11 mutant. Consistent with the Fv/Fm change presented above, the YIIs of salk033047 were significantly lower than those of heat treated Col-0 WT and 21 °C-control plants (Fig. 3c). Again, here we observed a positive correlation between the heat exposure times with the degree of the YII reduction in salk033047 plants (Fig. 3c). We also observed high light induced decreases in YII in WT and mutant plants in LC measurements (Fig. 3c), a phenomenon called photoinhibition. However, the responses of salk033047 plants to the photoinhibition were remarkably similar to those of WT plants and showed no association with the heat treatment (Fig. 3).
The IC and LC measurements also revealed significant decreases in the maximal P700 of PSI (Pm) in 30 °C-treated salk033047 plants (Fig. 4b). The decreases in Pm of mutant plants also showed an apparent correlation with the duration of the heat exposure (Fig. 4b). The longer the heat treatment, the larger decreases in Pm of mutant plants. Furthermore, moving the 21 °C-grown plants to 25 °C also induced a moderate reduction in Pm of mutant plants even though no changes in PSII efficiency of mutant plant were detected at this temperature (Additional file 3: Figure S2). Also, the Pm of control salk033047 plants (0.867) was slightly lower than Pm of control Col-0 (0.949), much similar to that (0.889) of 5d–treated WT (Fig. 4b).
Discussion
FtsH11 protease play an essential role in maintaining photosynthetic efficiency of photosystems at elevated temperatures in Arabidopsis
The FtsH11 is a chloroplast targeted protease [25] located exclusively in the chloroplast envelope [47]. It is the first plant FtsH that has been shown to play an essential role in thermotolerance in Arabidopsis [16], while most other well-studied chloroplast FtsH proteases functions in protecting the photosystems from high light stress [14, 15, 48]. Mutations in FtsH11 cause significant decreases in photosynthetic capability and a bleaching chlorotic leaf phenotype [16]. Similar chlorotic leaf phenotype has been observed in ftsh11 when grown under natural sunlight in the field in northern Sweden [28] and under continuous light at relatively high light intensities [49], conditions that are known to induce photoinhibition and cause damages to PSII [50–52]. Hence, a role of FtsH11 in protecting PSII from light stress has also been proposed [49]. Because high temperatures, even moderately high, can induce biochemical and structural changes and cause damages to PSII very much like those of high light stress [4, 53, 54], we used experimental conditions that can distinguish the changes in PS performance induced by heat from those induced by HL in FtsH11 mutant in this study. The light condition used for growing the plants and for the heat treatments was at the low end of optimal light condition (70 to 100 μmol quanta m−2 s−1) for Arabidopsis plant (12 h/12 h d/n) and generally do not impose light stress to photosystems of normal Arabidopsis plant. Likewise, the HL treatment performed at about 6 folds higher light intensity than of control but normal 21 °C temperature eliminated the likelihood of heat stress imposing on photosystems in Arabidopsis plant. Therefore, the differences detected between heat-stressed (30 °C and 35 °C) and control (21 °C) plants under normal light are likely to be caused by the moderately elevated temperatures while those detected between HL-stressed and normal light grown plants at 21 °C are likely to be induced by light stress. Under all the treatment conditions, we detected significant decreases in both Fv/Fm of PSII and Pm of PSI only in moderate heat-stressed ftsh11 plants while no differences were shown under either the control or HL-stress condition between the WT and FtsH11 mutant (Figs. 1, 2, 3 and 4). In plants, Fv/Fm and Pm are photosynthetic parameters indicating an overall photochemical efficiency of PSII and PSI, respectively. Decreases in Fv/Fm and Pm under stress conditions are indicators of stress induced damages to PS [55], mostly occurred at the reaction centers (RC) and the light harvesting complexes (LHC) [5, 40]. The specific association of Fv/Fm and Pm decreases with FtsH11 mutation under moderate temperatures suggests a primary function of this protease in maintaining photosynthetic activities of both PSII and PSI as the environmental temperature elevated above optimal. The correlation relationship between the degrees of Fv/Fm and Pm decreases in FtsH11 mutant and the durations and levels of the heat stress applied revealed in this study (Figs. 1a, 2a and 3c) further imply the active involvement of the FtsH11 in the adaptation and/or protection of the photosynthetic systems to/from heat stress in Arabidopsis. Furthermore, the study also revealed a negative correlation between the increases in Fo and the decreases in Fv during the course of heat treatment in mutant plants (Fig. 1e), therefore, uncovering the main contributing factor (Fo) to the detected decreases in Fv/Fm in ftsh11 (Figs. 1 and 2).
The role of FtsH11 in regulating the adaptation of photosystems to temperature increases in Arabidopsis
The present study has shown that the mutation of FtsH11 caused alterations in the array of Chl fluorescence parameters under moderately elevated temperatures of 30 °C and 35 °C (Figs. 1, 2, 3 and 4). The results reveal several lines of evidence for a critical role of FtsH11 in maintaining the overall structural and/or biochemical stability of photosynthetic systems at moderately elevated temperatures. First, the gradual but significant increase in Fo occurred only in heat stressed FtsH11 mutants, indicating damages to PSII RCs in FtsH11 plants under moderately high temperatures (Figs. 1 and 2c). The Fo in the dark adapted plant represents the dark fluorescence yield where all RCs of PSII are open. It has been shown that structural and biochemical alternations to the PSII RCs, such as dissociation of LHC from RC and monomerization of LHC can increase Fo [56, 57]. The increases in Fo have also been associated with PSII damages caused by both high temperature and high light stresses in plants [58]. The level of Fo is known to be closely associated with the thermostability of PSII and has been used to examine the damages to PSII caused by high temperature [42, 43]. High temperature induced severe structural and function damages to PSII is indicated by the significant increase Fo in Chl fluorescence measurements [42, 43]. Normally, the rapid increase of Fo occurs at temperatures about 40 °C or above, ranging from 40 to 50 °C depending on plant species, indicating severe damage to PSII [41, 42, 45]. Here we detected significant increases in Fo in FtsH11 mutant at 30 °C, a temperature that does not usually cause noticeable changes in PSII photochemistry, even for a cool season plants of Arabidopsis (Figs. 1, 2, 3 and 4). The results indicate severe damages to PSII LHC and RCs caused by either structural or biochemical alternations or a combination of both in PSII of FtsH11 plants under moderately elevated temperatures of 30 °C. It is known that chloroplasts respond to temperature increases by adjusting membrane properties [59, 60], modification of the configuration and/or spatial organization of protein complexes on thylakoids, regulation of the turnover of specific proteins such as D1 protein of PSII [34], and repairing damaged proteins to maintain structural intactness and function normality [5, 54, 57]. The proteolytic activities mediated by various enzymes, including FtsH proteases, are essential for efficient photosynthesis and critical for maintaining stability of various complexes in chloroplasts in response to daily temperature and light fluctuation and under high temperature and radiation stress [26, 31, 32, 51, 61]. The significant Fo increases in FtsH11 under moderately elevated temperatures indicates that FtsH11 protease plays essential roles in maintaining thermostability of photosystems in response to temperature increases. Moreover, no significant changes in Fo at HL condition and no significant differences in other Chl fluorescence parameters between WT and FtsH11mutant plants under control and HL conditions suggest that FtsH11 function specifically in maintaining thermostability of PSII under high temperature conditions. Therefore, maintaining a normal function of FtsH11 in chloroplast could be vital for plant’s adaptability to the daily and seasonal temperature fluctuation.
Second, the significant increases in qN of heat-treated FtsH11 plants (Fig. 3b) implied that the mutation in FtsH11 had a negative effect on the PSII photochemical efficiency in using the absorbed energy when growth temperature increased above optimal. The qN is a Chl fluorescence parameter commonly used to describe the efficiency of PSII in energy dissipation. In plants, the increase in NPQ often occurs when the light intensity of the growth environments exceeds the capacity for photosynthesis [5, 62]. It is a protective mechanism of plants to dissipate excess absorbed light energy as heat in order to prevent the generation of reactive oxygen species (ROS) and photodamages to photosystems (especially PSII) under high light condition [5, 62]. NPQ is a sensitive parameter for monitoring damage to photosystems caused by various environmental stresses [37, 40, 55]. Conformational changes of the thylakoid membrane, unstacking of grana and conformation change, monomerization of LHCII, and damages to PSII-LHCII induced by environmental factors such as high temperature and/or high light, are thought to be major causes for NPQ increases [57, 61, 63–65]. Although photosystems of FtsH11 plants appeared to function similarly as those of WT plants under normal temperature, the significant qN increase in FtsH11 after exposing to 30 °C (Fig. 3 and Additional file 2: Figure S1) indicates that the impairment of photosystems in utilizing the absorbed light energy for photochemistry in heat stressed mutant plants. The correlation of qN increases with the heat exposure times further point to the heat induced damages to photosystems in mutant plants. The results revealed an important role of FtsH11 protease in regulating functional stability of photosystems in response to environmental temperature increases in Arabidopsis.
Third, the detected decreases in ETR provide direct evidence on the effect of FtsH11 mutation on electron transport in both PSII and PSI (Fig. 3). High temperature stress induced changes in ETR have been reported in sweet sorghum and other species [5, 39, 44, 53]. It is known that increases in temperatures not only increase thylakoid membrane fluidity but also cause rearrangement and even disassociation of protein complexes and photosystems [4, 5]. Extreme heat induced structural changes in protein complexes, damage to photosystems, and the loss of oxygen evolving activity all reduce the ability of photosystems in electron transport [4, 53]. Here we showed significant ETR(II) and ETR(I) decreases in FtsH11 under a moderately elevated temperature which caused no ETR change in WT plants (Figs. 3 and 4). The results demonstrate the involvement of FtsH11 protease in maintaining effective electron transport of photosystems as environmental temperature increases in Arabidopsis.
In most studies, heat stress induced drastic functional and structural changes of photosystems often occur at extreme temperatures [39, 42, 44]. For most plants, it happens at mid to upper 40s°C [36, 43, 44, 66]. For cool-season species, such as Arabidopsis, wheat and barley, it occurs at temperature of about 40 °C to lower 40s [41, 42, 45]. The results of WT plants presented in this study show normal photosynthetic activities of Arabidopsis plants at treatment temperatures of 30 °C and 35 °C (Figs. 1, 2, 3 and 4, Additional file 2: Figure S1). The results of this study (Figs. 1, 2, 3 and 4, Additional file 2: Figure S1), however, reveal drastic changes for a range of Chl fluorescence parameters in FtsH11 mutant at 30 °C, a temperature much lower than typically reported temperature that alter the efficiency of photosynthetic systems. The findings all point to severe damages to photosynthetic systems in FtsH11 plants under this moderately elevated temperature at which WT plants functioned normally. The fact that photosynthetic systems of FtsH11 mutant functioned similarly as those of WT plants at optimal temperature under either normal light or high light conditions (Figs. 1, 2, 3 and 4) suggests specific role of FtsH11 in the maintenance of functional and structural stability of photosynthetic systems as the growth temperature elevated above optimal. The Fo increases in FtsH11 at 30 °C and 35 °C are likely the result of heat-induced dissociation of light harvesting complex (LHC) from the reaction center complexes and/or monomerization of LHC, reported phenomenon observed under extreme high temperature conditions [64]. The significant increases in qN and decreases in ETR(II) and YII of PSII and ETR(I) of PSI in 30 °C-treated FtsH11 plants are likely to be the results of heat induced damages to photosystems and possibly heat-induced configuration changes of protein complexes on thylakoid membranes.
The FtsH11 mediated thermotolerance of photosynthetic systems appears to be independent from oxidative stress
It is well known that heat stress induces the production of ROS such as hydrogen peroxide and superoxide [5]. Bursts of ROS (oxidative stress) can inhibit the PSII repair processes and the turnover of damaged protein subunits of PSII [52]. Using spinach thylakoids, Yamashita and colleagues have showed that the heat inactivation of PSII is caused by ROS produced during moderate heat stress processes [67]. Some of the Chl fluorescence parameter changes observed in the 30 °C-treated FtsH11 plants resemble those reported in oxidative stressed plants (Figs 1 and 2). Therefore, the possible association of the detected changes in Chl fluorescence in 30 °C-treated FtsH11 with overproduction of ROS was examined. The accumulations of hydrogen peroxide and superoxide in the most recent expanded rosette leaves of mutant and WT plants were compared by nitroblue tetrazolium and 3,3′-diaminobenzidine staining, respectively. Surprisingly, we found no noticeable difference in the level of these ROS accumulation (Additional file 4: Figure S3). The result also showed very minimal ROS induction under the moderate heat condition used in this study, thus rule out the possibility of ROS accumulation as a cause for the phenotypic and Chl fluorescence changes observed in heat-treated FtsH11 plants. Therefore, the thermotolerance mechanism of photosynthetic systems mediated by FtsH11 protease in chloroplasts is likely to be independent of oxidative stress.
The increase in Fo in FtsH11 mutants was not affected by HCF genes
The high-chlorophyll-fluorescence mutants (hcf) have been known to have high Fo value [68, 69], a phenotype similar to that observed in ftsh11 plants under moderately elevated temperatures (Figs. 1 and 2c). To determine the possible association of FtsH11 with any of the HCF genes as well as their association with moderate heat stress, we measured the Fo and Fv/Fm levels in all Arabidopsis HCF mutants available in ABRC (http://www.arabidopsis.org/) over a 5 d, 30 °C treatment. Although the Fo levels of all HCF mutants were higher than those of Col-0 WT and ftsh11 plants under normal 21 °C temperature, no further increase in Fo was observed in any of HCF mutants under the 30 °C treatment (Additional file 5: Table S2). Also, unlike FtsH11 mutants, we detected no significant decreases in Fv/Fm in 30 °C-treated HCF mutants. All leaves of HCF mutants stayed green in color and chlorophyll content remained at the same level throughout the course of 30 °C treatment. These results suggest that FtsH11 protease mediated mechanisms in maintaining normal photosynthetic activities at moderately elevated temperatures are likely to be independent from the functions of those HCF genes examined in this study.
Conclusions
This study shows that mutations in the FtsH11 protease gene altered an array of Chl fluorescence parameters, resulting in significant reduction in photosynthetic efficiency of PSII and PSI within a few days of a 30 °C challenge. It also reveals that the drastic Fo increases contributed the most to the detected decreases in maximum quantum efficiency of PSII photochemistry of FtsH11 mutant plants at elevated temperatures. The findings of this study show that the FtsH11 protease mediated network is essential in maintaining normal photosynthetic activities of photosystems possibly via its regulatory functions in the thermostability of photosynthetic machineries as environmental temperature rises above optimal. Such roles of FtsH11 protease appear to be specific to the adaptive mechanism of photosystems to temperature increases in chloroplasts. Disruption of the FtsH11 renders FtsH11 plants unable to perform these required regulatory functions in chloroplast. To understand the network regulated by FtsH11, it is necessary to identify the substrate of FtsH11 and the downstream components using genetic, biochemical and molecular approaches. To translate the discovery from Arabidopsis, the FtsH11 homologs should be evaluated in crop species for their functions in alleviating negative impacts of heat stress to photosystems. These experiments will offer new insights into the proteolytic processes of FtsH11 in regards to maintaining efficient photosynthesis under moderate heat stress and a foundation for improving photosynthetic efficiency of crop plants, hence sustain agricultural productivity under unfavorable environments.
Additional files
Summary of the IC and LC chlorophyll fluorescence parameters in Col-0 WT and FtsH11 mutant salk033047 plants in response to the 30 °C-treatment. Definition for the Chl fluorescence parameters and their calculation equations are provided in the second sheet. (XLSX 19 kb)
Induction response curves of chlorophyll fluorescence parameters of PSII and PSI in dark adapted Col-0 WT and salk033047 mutant (047) plants (plots with the “Time” as X-axis units). The light response curves of the Y(II), Y(NPQ), and Y(NO) of light adapted plants was also included here (plots with the light intensity “PAR” as the X-axis unit). (TIFF 9702 kb)
Measured P700 of PSI (Pm) and photosynthetic efficiency of PSII (YII) changes in 2 and 5 day, 25 °C treated Col WT and FtsH11 mutant salk033047 (047) leaves. The 25 °C-treatet mutant plants showed marginal decreases only for Pm in dark adapted plants and PSI and PSII activities were determined immediately on DualPAM100. (TIFF 2259 kb)
Accumulation of ROS in rosette leaves of 21 °C-control and 30 °C-treated Col-0 WT and FtsH11 mutant salk033047 plants. The leave tissues were harvested and stained in nitroblue tetrazolium (NB) and 3,3′-diaminobenzidine (DAB) solution to examine the production of hydrogen peroxide and superoxide respectively. (TIFF 1831 kb)
A) Comparisons of minimum chlorophyll fluorescence yield (Fo) of high chlorophyll fluorescence (hcf) mutant with salk0333047 plants at 21 °C and after exposure to 30 °C. b) Comparisons of photosystem II activity (Fv/Fm) of high chlorophyll fluorescence (hcf) mutant with salk0333047 plants at 21 °C and after exposure to 30 °C. (DOCX 40 kb)
Acknowledgements
The authors would like to thank Ms. Jing Wang for technical supports.
Funding
This work was supported partially by a grant from the US-Israel Binational Agricultural Research and Development Fund (US-422-09) and by USDA-ARS CRIS project.
Availability of data and materials
The datasets acquired during the current study are available from the corresponding author on reasonable request.
Disclaimer
Mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture.
Abbreviations
- Chl
Chlorophyll
- ETR(I)
PSI electron transport rate
- ETR(II)
PSII electron transport rate
- F0
Minimum Chl fluorescence yield in the dark-adapted state
- Fm
Maximum Chl fluorescence yield in the dark-adapted state (Dark fluorescence yield)
- Fm′
Maximal fluorescence yield in the illuminated sample
- F0/FM
A parameter related to changes in heat dissipation in the photosystem II antenna
- FtsH
Filamentous temperature sensitive H
- Fv
Maximum Variable fluorescence defined as FM-F0
- Fv/Fm
Maximum quantum yield of PSII photochemistry
- HCF
High-chlorophyll-fluorescence
- HL
High light
- IC
Fluorescence induction curve
- LC
Light response curve
- LHC
Light harvesting complex
- NPQ
Non-photochemical quenching
- OEX
Oxygen-evolving complex
- PAR
Photosynthetically active radiation
- PSI
Photosystem I
- PSII
Photosystem II
- qN
Coefficient of nonphotochemical quenching
- RC
Reaction center
- ROS
Reactive oxygen species
- SP
Saturating pulse (SP)
- WT
Wild type
- YII
Effective PSII quantum yield
Authors’ contributions
JC conceived the project, designed and performed the experiments, and wrote the manuscript, JC, JB and ZX analyzed data and wrote the manuscript. All the authors read and approved the final manuscript.
Ethics approval and consent to participate
The salk033047 and hcf mutants were purchased from the Arabidopsis Biological Resource Center (https://abrc.osu.edu). No permission is required for using these materials in this study.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Footnotes
Electronic supplementary material
The online version of this article (10.1186/s12870-018-1228-2) contains supplementary material, which is available to authorized users.
Contributor Information
Junping Chen, Email: junping.chen@ars.usda.gov.
John J. Burke, Email: john.burke@ars.usda.gov
Zhanguo Xin, Email: zhanguo.xin@ars.usda.gov.
References
- 1.Kim K, Portis AR., Jr Temperature dependence of photosynthesis in Arabidopsis plants with modifications in rubisco activase and membrane fluidity. Plant Cell Physiol. 2005;46(3):522–530. doi: 10.1093/pcp/pci052. [DOI] [PubMed] [Google Scholar]
- 2.Weis E, Berry JA. Plants and high temperature stress. Symp Soc Exp Biol. 1988;42:329–346. [PubMed] [Google Scholar]
- 3.Crafts-Brandner SJ, Salvucci ME. Sensitivity of photosynthesis in a C4 plant, maize, to heat stress. Plant Physiol. 2002;129(4):1773–1780. doi: 10.1104/pp.002170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Mathur S, Agrawal D, Jajoo A. Photosynthesis: response to high temperature stress. J Photochem Photobiol B. 2014;137:116–126. doi: 10.1016/j.jphotobiol.2014.01.010. [DOI] [PubMed] [Google Scholar]
- 5.Yamamoto Y. Quality control of photosystem II: the mechanisms for avoidance and tolerance of light and heat stresses are closely linked to membrane fluidity of the thylakoids. Front Plant Sci. 2016;7:1136. doi: 10.3389/fpls.2016.01136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Komayama K, Khatoon M, Takenaka D, Horie J, Yamashita A, Yoshioka M, Nakayama Y, Yoshida M, Ohira S, Morita N, et al. Quality control of photosystem II: cleavage and aggregation of heat-damaged D1 protein in spinach thylakoids. Biochim Biophys Acta. 2007;1767(6):838–846. doi: 10.1016/j.bbabio.2007.05.001. [DOI] [PubMed] [Google Scholar]
- 7.Essemine J, Govindachary S, Ammar S, Bouzid S, Carpentier R. Abolition of photosystem I cyclic electron flow in Arabidopsis Thaliana following thermal-stress. Plant Physiol Biochem. 2011;49(3):235–243. doi: 10.1016/j.plaphy.2010.11.002. [DOI] [PubMed] [Google Scholar]
- 8.Tebaldi C, Hayhoe K, Arblaster JM, Meehl GA. Going to the extremes. Clim Chang. 2006;79(3):185–211. [Google Scholar]
- 9.Solomon S, Rosenlof KH, Portmann RW, Daniel JS, Davis SM, Sanford TJ, Plattner GK. Contributions of stratospheric water vapor to decadal changes in the rate of global warming. Science. 2010;327(5970):1219–1223. doi: 10.1126/science.1182488. [DOI] [PubMed] [Google Scholar]
- 10.Teixeira EI, Fischer G, van Velthuizen H, Walter C, Ewert F. Global hot-spots of heat stress on agricultural crops due to climate change. Agric For Meteorol. 2013;170(0):206–215. [Google Scholar]
- 11.Rosenzweig C, Elliott J, Deryng D, Ruane AC, Muller C, Arneth A, Boote KJ, Folberth C, Glotter M, Khabarov N, et al. Assessing agricultural risks of climate change in the 21st century in a global gridded crop model intercomparison. Proc Natl Acad Sci U S A. 2014;111(9):3268–3273. doi: 10.1073/pnas.1222463110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Boyer JS. Plant productivity and environment ( crop genetic improvement) Science. 1982;218(4571):443–448. doi: 10.1126/science.218.4571.443. [DOI] [PubMed] [Google Scholar]
- 13.Lobell DB, Banziger M, Magorokosho C, Vivek B. Nonlinear heat effects on African maize as evidenced by historical yield trials. Nature Clim Chang. 2011;1(1):42–45. [Google Scholar]
- 14.Yoshioka-Nishimura M, Yamamoto Y. Quality control of photosystem II: the molecular basis for the action of FtsH protease and the dynamics of the thylakoid membranes. J Photochem Photobiol B. 2014;137:100–106. doi: 10.1016/j.jphotobiol.2014.02.012. [DOI] [PubMed] [Google Scholar]
- 15.Wagner R, Aigner H, Funk C. FtsH proteases located in the plant chloroplast. Physiol Plant. 2012;145(1):203–214. doi: 10.1111/j.1399-3054.2011.01548.x. [DOI] [PubMed] [Google Scholar]
- 16.Chen J, Burke JJ, Velten J, Xin Z. FtsH11 protease plays a critical role in Arabidopsis thermotolerance. Plant J. 2006;48(1):73–84. doi: 10.1111/j.1365-313X.2006.02855.x. [DOI] [PubMed] [Google Scholar]
- 17.Sakamoto W, Tamura T, Hanba-Tomita Y, Murata M. The VAR1 locus of Arabidopsis encodes a chloroplastic FtsH and is responsible for leaf variegation in the mutant alleles. Genes Cells. 2002;7(8):769–780. doi: 10.1046/j.1365-2443.2002.00558.x. [DOI] [PubMed] [Google Scholar]
- 18.Lindahl M, Spetea C, Hundal T, Oppenheim AB, Adam Z, Andersson B. The thylakoid FtsH protease plays a role in the light-induced turnover of the photosystem II D1 protein. Plant Cell. 2000;12(3):419–431. doi: 10.1105/tpc.12.3.419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Fischer B, Rummel G, Aldridge P, Jenal U. The FtsH protease is involved in development, stress response and heat shock control in Caulobacter crescentus. Mol Microbiol. 2002;44(2):461–478. doi: 10.1046/j.1365-2958.2002.02887.x. [DOI] [PubMed] [Google Scholar]
- 20.Tomoyasu T, Gamer J, Bukau B, Kanemori M, Mori H, Rutman AJ, Oppenheim AB, Yura T, Yamanaka K, Niki H, et al. Escherichia Coli FtsH is a membrane-bound, ATP-dependent protease which degrades the heat-shock transcription factor sigma 32. EMBO J. 1995;14(11):2551–2560. doi: 10.1002/j.1460-2075.1995.tb07253.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bourdineaud JP, Nehme B, Tesse S, Lonvaud-Funel A. The ftsH gene of the wine bacterium Oenococcus oeni is involved in protection against environmental stress. Appl Environ Microbiol. 2003;69(5):2512–2520. doi: 10.1128/AEM.69.5.2512-2520.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Schumann W. FtsH--a single-chain charonin? FEMS Microbiol Rev. 1999;23(1):1–11. doi: 10.1111/j.1574-6976.1999.tb00389.x. [DOI] [PubMed] [Google Scholar]
- 23.Bittner LM, Arends J, Narberhaus F. When, how and why? Regulated proteolysis by the essential FtsH protease in Escherichia Coli. Biol Chem. 2017;398(5–6):625–635. doi: 10.1515/hsz-2016-0302. [DOI] [PubMed] [Google Scholar]
- 24.Fiocco D, Collins M, Muscariello L, Hols P, Kleerebezem M, Msadek T, Spano G. The lactobacillus plantarum ftsH gene is a novel member of the CtsR stress response regulon. J Bacteriol. 2009;191(5):1688–1694. doi: 10.1128/JB.01551-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Sakamoto W, Zaltsman A, Adam Z, Takahashi Y. Coordinated regulation and complex formation of yellow variegated1 and yellow variegated2, chloroplastic FtsH metalloproteases involved in the repair cycle of photosystem II in Arabidopsis thylakoid membranes. Plant Cell. 2003;15(12):2843–2855. doi: 10.1105/tpc.017319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Adam Z, Rudella A, van Wijk KJ. Recent advances in the study of Clp, FtsH and other proteases located in chloroplasts. Curr Opin Plant Biol. 2006;9(3):234–240. doi: 10.1016/j.pbi.2006.03.010. [DOI] [PubMed] [Google Scholar]
- 27.Nixon PJ, Barker M, Boehm M, de Vries R, Komenda J. FtsH-mediated repair of the photosystem II complex in response to light stress. J Exp Bot. 2005;56(411):357–363. doi: 10.1093/jxb/eri021. [DOI] [PubMed] [Google Scholar]
- 28.Wagner R, Aigner H, Pruzinska A, Jankanpaa HJ, Jansson S, Funk C. Fitness analyses of Arabidopsis Thaliana mutants depleted of FtsH metalloproteases and characterization of three FtsH6 deletion mutants exposed to high light stress, senescence and chilling. New Phytol. 2011;191(2):449–458. doi: 10.1111/j.1469-8137.2011.03684.x. [DOI] [PubMed] [Google Scholar]
- 29.Zaltsman A, Ori N, Adam Z. Two types of FtsH protease subunits are required for chloroplast biogenesis and photosystem II repair in Arabidopsis. Plant Cell. 2005;17(10):2782–2790. doi: 10.1105/tpc.105.035071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Yu F, Park S, Rodermel SR. The Arabidopsis FtsH metalloprotease gene family: interchangeability of subunits in chloroplast oligomeric complexes. Plant J. 2004;37(6):864–876. doi: 10.1111/j.1365-313x.2003.02014.x. [DOI] [PubMed] [Google Scholar]
- 31.Kato Y, Ozawa S, Takahashi Y, Sakamoto W. D1 fragmentation in photosystem II repair caused by photo-damage of a two-step model. Photosynth Res. 2015;126(2–3):409–416. doi: 10.1007/s11120-015-0144-7. [DOI] [PubMed] [Google Scholar]
- 32.Yoshioka M, Nakayama Y, Yoshida M, Ohashi K, Morita N, Kobayashi H, Yamamoto Y. Quality control of photosystem II: FtsH hexamers are localized near photosystem II at grana for the swift repair of damage. J Biol Chem. 2010;285(53):41972–41981. doi: 10.1074/jbc.M110.117432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Chen M, Choi Y, Voytas DF, Rodermel S. Mutations in the Arabidopsis VAR2 locus cause leaf variegation due to the loss of a chloroplast FtsH protease. Plant J. 2000;22(4):303–313. doi: 10.1046/j.1365-313x.2000.00738.x. [DOI] [PubMed] [Google Scholar]
- 34.Kato Y, Sun X, Zhang L, Sakamoto W. Cooperative D1 degradation in the photosystem II repair mediated by chloroplastic proteases in Arabidopsis. Plant Physiol. 2012;159(4):1428–1439. doi: 10.1104/pp.112.199042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Sedaghatmehr M, Mueller-Roeber B, Balazadeh S. The plastid metalloprotease FtsH6 and small heat shock protein HSP21 jointly regulate thermomemory in Arabidopsis. Nat Commun. 2016;7:12439. doi: 10.1038/ncomms12439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Chen K, Sun X, Amombo E, Zhu Q, Zhao Z, Chen L, Xu Q, Fu J. High correlation between thermotolerance and photosystem II activity in tall fescue. Photosynth Res. 2014;122(3):305–314. doi: 10.1007/s11120-014-0035-3. [DOI] [PubMed] [Google Scholar]
- 37.Dabrowski P, Baczewska AH, Pawluskiewicz B, Paunov M, Alexantrov V, Goltsev V, Kalaji MH. Prompt chlorophyll a fluorescence as a rapid tool for diagnostic changes in PSII structure inhibited by salt stress in perennial ryegrass. J Photochem Photobiol B. 2016;157:22–31. doi: 10.1016/j.jphotobiol.2016.02.001. [DOI] [PubMed] [Google Scholar]
- 38.Guo Y, Tan J. Recent advances in the application of chlorophyll a fluorescence from photosystem II. Photochem Photobiol. 2015;91(1):1–14. doi: 10.1111/php.12362. [DOI] [PubMed] [Google Scholar]
- 39.Mathur S, Jajoo A. Alterations in photochemical efficiency of photosystem II in wheat plant on hot summer day. Physiology and molecular biology of plants : an international journal of functional plant biology. 2014;20(4):527–531. doi: 10.1007/s12298-014-0249-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kalaji HM, Bosa K, Koscielniak J, Hossain Z. Chlorophyll a fluorescence--a useful tool for the early detection of temperature stress in spring barley (Hordeum Vulgare L) Omics : a journal of integrative biology. 2011;15(12):925–934. doi: 10.1089/omi.2011.0070. [DOI] [PubMed] [Google Scholar]
- 41.Brestic M, Zivcak M, Kunderlikova K, Allakhverdiev SI. High temperature specifically affects the photoprotective responses of chlorophyll b-deficient wheat mutant lines. Photosynth Res. 2016;130(1–3):251–266. doi: 10.1007/s11120-016-0249-7. [DOI] [PubMed] [Google Scholar]
- 42.Brestic M, Zivcak M, Kalaji HM, Carpentier R, Allakhverdiev SI, Photosystem II. Thermostability in situ: environmentally induced acclimation and genotype-specific reactions in Triticum Aestivum L. Plant Physiol Biochem. 2012;57:93–105. doi: 10.1016/j.plaphy.2012.05.012. [DOI] [PubMed] [Google Scholar]
- 43.Pastenes C, Horton P. Resistance of photosynthesis to high temperature in two bean varieties (Phaseolus Vulgaris L) Photosynth Res. 1999;62(2):197–203. [Google Scholar]
- 44.Yan K, Chen P, Shao H, Shao C, Zhao S, Brestic M. Dissection of photosynthetic electron transport process in sweet sorghum under heat stress. PLoS One. 2013;8(5):e62100. doi: 10.1371/journal.pone.0062100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Havaux M. Stress tolerance of photosystem II in vivo: antagonistic effects of water, heat, and Photoinhibition stresses. Plant Physiol. 1992;100(1):424–432. doi: 10.1104/pp.100.1.424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.van Kooten O, Snel JF. The use of chlorophyll fluorescence nomenclature in plant stress physiology. Photosynth Res. 1990;25(3):147–150. doi: 10.1007/BF00033156. [DOI] [PubMed] [Google Scholar]
- 47.Ferro M, Brugière S, Salvi D, Seigneurin-Berny D, Court M, Moyet L, Ramus C, Miras S, Mellal M, Le Gall S, et al. AT_CHLORO, a comprehensive chloroplast proteome database with Subplastidial localization and curated information on envelope proteins. Mol Cell Proteomics. 2010;9(6):1063–1084. doi: 10.1074/mcp.M900325-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Nishimura K, Kato Y, Sakamoto W. Chloroplast proteases: updates on proteolysis within and across Suborganellar compartments. Plant Physiol. 2016;171(4):2280–2293. doi: 10.1104/pp.16.00330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Wagner R, von Sydow L, Aigner H, Netotea S, Brugiere S, Sjogren L, Ferro M, Clarke A, Funk C. Deletion of FtsH11 protease has impact on chloroplast structure and function in Arabidopsis Thaliana when grown under continuous light. Plant Cell Environ. 2016;39(11):2530–2544. doi: 10.1111/pce.12808. [DOI] [PubMed] [Google Scholar]
- 50.Yamamoto Y, Kai S, Ohnishi A, Tsumura N, Ishikawa T, Hori H, Morita N, Ishikawa Y. Quality control of PSII: behavior of PSII in the highly crowded grana thylakoids under excessive light. Plant Cell Physiol. 2014;55(7):1206–1215. doi: 10.1093/pcp/pcu043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Nixon PJ, Michoux F, Yu J, Boehm M, Komenda J. Recent advances in understanding the assembly and repair of photosystem II. Ann Bot. 2010;106(1):1–16. doi: 10.1093/aob/mcq059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Murata N, Takahashi S, Nishiyama Y, Allakhverdiev SI. Photoinhibition of photosystem II under environmental stress. Biochim Biophys Acta. 2007;1767(6):414–421. doi: 10.1016/j.bbabio.2006.11.019. [DOI] [PubMed] [Google Scholar]
- 53.Marutani Y, Yamauchi Y, Kimura Y, Mizutani M, Sugimoto Y. Damage to photosystem II due to heat stress without light-driven electron flow: involvement of enhanced introduction of reducing power into thylakoid membranes. Planta. 2012;236(2):753–761. doi: 10.1007/s00425-012-1647-5. [DOI] [PubMed] [Google Scholar]
- 54.Zhang R, Wise RR, Struck KR, Sharkey TD. Moderate heat stress of Arabidopsis Thaliana leaves causes chloroplast swelling and plastoglobule formation. Photosynth Res. 2010;105(2):123–134. doi: 10.1007/s11120-010-9572-6. [DOI] [PubMed] [Google Scholar]
- 55.Ritchie GA. Riley, LE; Dumroese, RK; Landis, TD, tech coords 2006 National Proceedings: Forest and conservation nursery associations - 2005 proc RMRS-P-43 Fort Collins, CO: US Department of Agriculture, Forest Service, Rocky Mountain Research Station. 2006. Chlorophyll fluorescence: what is it and what do the numbers mean? pp. 34–42. [Google Scholar]
- 56.Haferkamp S, Kirchhoff H. Significance of molecular crowding in grana membranes of higher plants for light harvesting by photosystem II. Photosynth Res. 2008;95(2–3):129–134. doi: 10.1007/s11120-007-9253-2. [DOI] [PubMed] [Google Scholar]
- 57.Minagawa J. Dynamic reorganization of photosynthetic supercomplexes during environmental acclimation of photosynthesis. Front Plant Sci. 2013;4:513. doi: 10.3389/fpls.2013.00513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Saakov VS. High-temperature stress-related changes in the harmonics F0, Fm, and Fv of pulse-amplitude modulated fluorescence signals: locating thermal damage in reaction centers of photosystem II. Dokl Biochem Biophys. 2002;382:4–9. doi: 10.1023/a:1014438703467. [DOI] [PubMed] [Google Scholar]
- 59.Essemine J, Govindachary S, Ammar S, Bouzid S, Carpentier R. Functional aspects of the photosynthetic light reactions in heat stressed Arabidopsis deficient in digalactosyl-diacylglycerol. J Plant Physiol. 2011;168(13):1526–1533. doi: 10.1016/j.jplph.2011.01.015. [DOI] [PubMed] [Google Scholar]
- 60.Chen J, Burke JJ, Xin Z, Xu C, Velten J. Characterization of the Arabidopsis thermosensitive mutant atts02 reveals an important role for galactolipids in thermotolerance. Plant Cell Environ. 2006;29(7):1437–1448. doi: 10.1111/j.1365-3040.2006.01527.x. [DOI] [PubMed] [Google Scholar]
- 61.Yoshioka-Nishimura M, Nanba D, Takaki T, Ohba C, Tsumura N, Morita N, Sakamoto H, Murata K, Yamamoto Y. Quality control of photosystem II: direct imaging of the changes in the thylakoid structure and distribution of FtsH proteases in spinach chloroplasts under light stress. Plant Cell Physiol. 2014;55(7):1255–1265. doi: 10.1093/pcp/pcu079. [DOI] [PubMed] [Google Scholar]
- 62.Ebbert V, Demmig-Adams B, Adams WW, 3rd, Mueh KE, Staehelin LA. Correlation between persistent forms of zeaxanthin-dependent energy dissipation and thylakoid protein phosphorylation. Photosynth Res. 2001;67(1–2):63–78. doi: 10.1023/A:1010640612843. [DOI] [PubMed] [Google Scholar]
- 63.Essemine J, Govindachary S, Joly D, Ammar S, Bouzid S, Carpentier R. Effect of moderate and high light on photosystem II function in Arabidopsis Thaliana depleted in digalactosyl-diacylglycerol. Biochimica et Biophysica Acta (BBA) - Bioenergetics. 2012;1817(8):1367–1373. doi: 10.1016/j.bbabio.2012.02.004. [DOI] [PubMed] [Google Scholar]
- 64.Santabarbara S, Horton P, Ruban AV. Comparison of the thermodynamic landscapes of unfolding and formation of the energy dissipative state in the isolated light harvesting complex II. Biophys J. 2009;97(4):1188–1197. doi: 10.1016/j.bpj.2009.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Demmig-Adams B, Muller O, Stewart JJ, Cohu CM, Adams WW., 3rd Chloroplast thylakoid structure in evergreen leaves employing strong thermal energy dissipation. J Photochem Photobiol B. 2015;152(Pt B):357–366. doi: 10.1016/j.jphotobiol.2015.03.014. [DOI] [PubMed] [Google Scholar]
- 66.Dreyer E, Le Roux X, Montpied P, Daudet FA, Masson F. Temperature response of leaf photosynthetic capacity in seedlings from seven temperate tree species. Tree Physiol. 2001;21(4):223–232. doi: 10.1093/treephys/21.4.223. [DOI] [PubMed] [Google Scholar]
- 67.Yamashita A, Nijo N, Pospisil P, Morita N, Takenaka D, Aminaka R, Yamamoto Y. Quality control of photosystem II: reactive oxygen species are responsible for the damage to photosystem II under moderate heat stress. J Biol Chem. 2008;283(42):28380–28391. doi: 10.1074/jbc.M710465200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Meurer J, Meierhoff K, Westhoff P. Isolation of high-chlorophyll-fluorescence mutants of Arabidopsis Thaliana and their characterisation by spectroscopy, immunoblotting and northern hybridisation. Planta. 1996;198(3):385–396. doi: 10.1007/BF00620055. [DOI] [PubMed] [Google Scholar]
- 69.Wei L, Guo J, Ouyang M, Sun X, Ma J, Chi W, Lu C, Zhang L. LPA19, a Psb27 homolog in Arabidopsis Thaliana, facilitates D1 protein precursor processing during PSII biogenesis. J Biol Chem. 2010;285(28):21391–21398. doi: 10.1074/jbc.M110.105064. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Summary of the IC and LC chlorophyll fluorescence parameters in Col-0 WT and FtsH11 mutant salk033047 plants in response to the 30 °C-treatment. Definition for the Chl fluorescence parameters and their calculation equations are provided in the second sheet. (XLSX 19 kb)
Induction response curves of chlorophyll fluorescence parameters of PSII and PSI in dark adapted Col-0 WT and salk033047 mutant (047) plants (plots with the “Time” as X-axis units). The light response curves of the Y(II), Y(NPQ), and Y(NO) of light adapted plants was also included here (plots with the light intensity “PAR” as the X-axis unit). (TIFF 9702 kb)
Measured P700 of PSI (Pm) and photosynthetic efficiency of PSII (YII) changes in 2 and 5 day, 25 °C treated Col WT and FtsH11 mutant salk033047 (047) leaves. The 25 °C-treatet mutant plants showed marginal decreases only for Pm in dark adapted plants and PSI and PSII activities were determined immediately on DualPAM100. (TIFF 2259 kb)
Accumulation of ROS in rosette leaves of 21 °C-control and 30 °C-treated Col-0 WT and FtsH11 mutant salk033047 plants. The leave tissues were harvested and stained in nitroblue tetrazolium (NB) and 3,3′-diaminobenzidine (DAB) solution to examine the production of hydrogen peroxide and superoxide respectively. (TIFF 1831 kb)
A) Comparisons of minimum chlorophyll fluorescence yield (Fo) of high chlorophyll fluorescence (hcf) mutant with salk0333047 plants at 21 °C and after exposure to 30 °C. b) Comparisons of photosystem II activity (Fv/Fm) of high chlorophyll fluorescence (hcf) mutant with salk0333047 plants at 21 °C and after exposure to 30 °C. (DOCX 40 kb)
Data Availability Statement
The datasets acquired during the current study are available from the corresponding author on reasonable request.