

Abstract
Objective
To provide an in-depth catalog of the salivary proteome and endogenous peptidome of healthy dogs, evaluate proteins and peptides with antimicrobial properties, and compare the most common salivary proteins and peptides between different breed phylogeny groups.
Methods
36 healthy dogs without evidence of periodontal disease representing four breed phylogeny groups, based upon single nucleotide polymorphism haplotypes (ancient, herding/sighthound, and two miscellaneous groups). Saliva collected from dogs was pooled by phylogeny group and analyzed using nanoscale liquid chromatography-tandem mass spectrometry. Resulting tandem mass spectra were compared to databases for identification of endogenous peptides and inferred proteins.
Results
2,491 proteins and endogenous peptides were found in the saliva of healthy dogs with no periodontal disease. All dog phylogeny groups’ saliva was rich in proteins and peptides with antimicrobial functions. The ancient breeds group was distinct in that it contained unique proteins and was missing many proteins and peptides present in the other groups.
Conclusions and clinical relevance
Using a sophisticated nanoscale liquid chromatography-tandem mass spectrometry, we were able to identify 10-fold more salivary proteins than previously reported in dogs. Seven of the top 10 most abundant proteins or peptides serve immune functions and many more with various antimicrobial mechanisms were found. This is the most comprehensive analysis of healthy canine saliva to date, and will provide the groundwork for future studies analyzing salivary proteins and endogenous peptides in disease states.
Introduction
Saliva is composed of a complex mixture of enzymes, glycoproteins, immunoglobulins, peptides, inorganic substances, white blood cells, epithelial cells, and microflora, in addition to water. The substances in saliva originate primarily from salivary glands but also blood and nasal-bronchial secretions [1–2]. Food digestion and lubrication are well-recognized functions of saliva; however, this complex fluid also protects the oral cavity against pathogens, maintains the mouth pH and has a role in taste [3–4].
For many decades saliva has been considered the reflection of health and disease states of the oral cavity in addition to the whole body [4]. The relatively easy and non-invasive access to saliva samples compounded with the remarkable advances in the technology to investigate proteins—a major saliva component—have spiked researchers’ interest in looking at the composition of this biological fluid in healthy individuals with the ultimate goal of identifying biomarkers of diseases [4–9].
Various mass spectrometry methods are currently available and have been widely used to study the salivary proteins, and smaller, endogenous peptides. However, despite extensive investigation in humans [2,5,8,10–26], much less work has been done in other species [3,27–34]. Recently, the protein components of dog saliva were examined using SDS-PAGE-LC coupled to tandem mass spectrometry (MS/MS) [3], but only one dog sample was analyzed. Hence, a more comprehensive characterization of the salivary proteome and endogenous peptidome of healthy dogs is needed and can provide a valuable groundwork for future studies searching for specific changes in salivary protein composition associated with oral and systemic diseases.
The primary study aim was to provide an in-depth catalog of the salivary endogenous peptidome and proteome of healthy dogs. Additional aims included an evaluation of proteins and peptides with antimicrobial properties and comparison of the most common salivary proteins between different breed phylogeny groups.
Materials and methods
A summary of the study design and methodology used is shown in Fig 1.
Fig 1. Flow chart summarizing the study design.

Animals
The study was approved by the Institutional Animal Care and Use Committee of the University of Minnesota. Thirty-six clinically healthy dogs owned by faculty, staff and students at the University of Minnesota, College of Veterinary Medicine, were selected for the study. The dogs were carefully examined by a veterinarian with experience in dental disease to assure the absence of any dental abnormalities, especially periodontal disease. Sixteen dogs were males (11 intact and 5 neutered) and 20 were females (16 spayed and 4 intact). Their age ranged from 4 to 148 months (mean = 40.58 months; median = 25 months). The following breeds were represented: Hound cross (n = 6), Labrador Retriever (5), Alaskan Malamute (3), Bernese Mountain Dog (3), Siberian Husky (3), Golden Retriever (2), German Shepherd cross (2), American Staffordshire Terrier cross (1), Australian Cattle Dog (1), Belgian Tervuren (1), Boxer cross (1), French Bulldog (1), German Shorthair Pointer (1), German Wirehair Pointer (1), Irish Wolfhound (1), Jack Russell Terrier (1), Mixed breed dog (1), Newfoundland (1), and Scottish Deerhound (1) (Fig 1).
To try to investigate the potential effect of genetic lineage on the salivary proteomics and peptidomics, the dogs were initially selected to represent the four lineages that cluster based on structure analysis of microsatellite markers: asian/ancient, herding, hunting, and mastiff [35]. However, more recent analysis of single nucleotide polymorphism haplotypes divides dog breeds into 10 clusters [36]. Given this newer classification and the inclusion of some dogs with incomplete parentage information (crossbred dogs), the four groups were renamed as follows: ancient, herding-sighthound, and miscellaneous 1 and 2 (Fig 1). Saliva was pooled for dogs within each group.
Saliva collection
Whole saliva was collected without previous stimulation using saliva collection kits (SalivaBio kit; Salimetrics®, State College, PA) according to the manufacturer’s recommendation. Briefly, dog owners were asked to withhold food and water from their pets for at least 1 hour before sample collection. A cotton swab of 125mm length and 8mm diameter was placed in each dog’s cheek pouches for 45–60 seconds, with the collector gently holding the dog’s muzzle to prevent swallowing. Upon removal, the swab was placed in a special tube (Swab Storage Tube) and the saliva extracted by centrifugation at 685 × g for 15 minutes. The swab was removed from the tube and the saliva immediately stored at -80°C freezer until analysis [37].
Sample preparation (offline high pH reverse phase-liquid chromatography fractionation)
Saliva samples were thawed on ice and cleared of cells and debris by centrifugation at 3000 × g for 10 minutes and 16,100 × g for 1 minute at 4°C. Protein concentrations of the supernatants were determined by the bicinchoninic acid (BCA) assay. All samples were qualitatively analyzed by SDS-PAGE prior to being used in the study.
Pools of saliva from each group were prepared for proteomic analysis by combining equal protein amounts for each sample within a category to a total protein amount of 200 μg per pool. The pools were digested using the FASP protocol [38] using 10kDa filters (Pall Nanosep 10kDa filters; VWR, OD010C34) and the resulting peptides desalted using silica-based sorbent cartridges (Sep-Pak tC18 cartridges; Waters, WAT054925).
Endogenous peptides (naturally occurring salivary peptides below 10kDa) were collected from the flow-through after the initial centrifugation step using the 10kDa filters (prior to protein alkylation). These peptides were separately reduced using 5 mM tris (2-carboxyethyl) phosphine (TCEP) and alkylated using 50 mM iodoacetamide. Endogenous peptide samples were cleaned up on mixed-mode polymeric sorbent cartridges (Oasis MCX cartridges; Waters, 186000253).
The trypsin-digested protein samples were fractionated using high-pH reversed phase with subsequent concatenation similar to the protocol of Wang et al [39]. The samples were dissolved in 200 mM ammonium formate, pH 10 containing 2% acetonitrile (ACN) and loaded onto a C18 column (Phenomenex Kinetek C18 column; 2.6 μm, 2.1 x 100 mm). Solvents A and B were 20 mM ammonium formate, pH 10, containing 2 and 90% ACN respectively. A gradient was run at 200 μL/min with the following steps: 0 min, 2% B; 5 min, 2% B; 5.5 min, 5%B; 28 min, 30% B; 31 min, 60% B; 33 min, 90% B; 40 min, 90% B; 41 min, 2% B; 45 min, 2% B. The column was heated to 55°C with a heated sleeve (Analytical Sales & Service, Inc., HSI-25L). Fractions were collected every minute, and were concatenated by combining a volume equivalent to 15 mAU from fractions 7 and 21, 8 and 22, etc., until fractions 20 and 34 to produce 14 concatenated fractions. These were dried in a speed-vac, and re-dissolved in 37.5 μL of 0.1% trifluoroacetic acid (TFA) in 2% ACN load solvent.
Nano liquid chromatography–tandem mass spectrometry (LC-MS/MS) analysis
Analysis of the concatenated fractions from the digest proteins, as well as the endogenous peptides, was performed on a mass spectrometer (Orbitrap Fusion Tribrid with Easy-nLC autosampler and LC; Thermo Scientific, Waltham, MA) equipped with an autosampler and LC system. For protein samples, 2.5 μL of each concatenated fraction was injected directly onto an in-house packed, 10 cm x 75 μm column packed with 3 μm C18 particles. Separation was achieved by a gradient from 2–30% B over 50 min, followed by a 2 min ramp to 90% B and 8 min hold at 90% B. The flow rate was 200 nL/min. The MS operated in a top speed, data-dependent mode with a cycle time of 3 s. MS1 scans were performed in the Orbitrap at 120k resolution from 400–1500 m/z with an AGC target of 4E5. Percursor isolation took place in the quadrupole with an isolation width of 1.6 m/z. CID was performed at 35% NCE and MS2 spectra were collected in the ion trap. Dynamic exclusion used a repeat count of 1 for a duration of 30 s.
Sequence database search for proteins and peptides
The data were searched against a RefSeq Canis familiaris database with common contaminant proteins, containing 47336 entries, using protein analysis software (Sequest HT node in Proteome Discoverer 2.0; Thermo Scientific, Waltham, MA). Search parameters used included trypsin enzyme specificity with ≤2 missed cleavages for analysis of intact proteins, carbamidomethyl as a fixed modification on cysteine and variable modification of methionine oxidation and protein N-terminal acetylation. Precursor and product ion mass tolerances of 35 ppm and 0.6 Da were used. For identification of endogenous peptides, all parameters were the same as above, except that no enzyme was specified.
Criteria for protein identification
MS/MS based peptide and inferred protein identifications were validated using proteomic analysis software (Scaffold; version Scaffold 4.6.1; Proteome Software Inc., Portland, OR). For the analysis of intact salivary proteins, peptide identifications were accepted if they could be established at greater than 92.0% probability by the Scaffold Local FDR algorithm. Peptide identifications were also required to exceed specific database search engine thresholds. Sequest identifications required at least deltaCn scores of greater than 0.0 and XCorr scores of greater than 1.8, 2.2, 2.5 and 3.5 for singly, doubly, triply and quadruply charged peptides. Protein identifications were accepted if they could be established at greater than 5.0% probability to achieve an estimated FDR less than or equal to 1.0% and contained at least 1 identified peptide. Protein probabilities were assigned by the Protein Prophet algorithm [40]. Proteins that contained similar peptides and could not be differentiated based on MS/MS analysis alone were grouped to satisfy the principles of parsimony. Proteins sharing significant peptide evidence were grouped into clusters. Collectively, the above criteria resulted in an estimated peptide FDR of 0.2% and an estimated protein FDR of 1.0%, both estimated using the target-decoy method. S1 Appendix contains all information on identified proteins and peptides for the analysis of intact salivary proteins.
For the identification of endogenous salivary peptides, accepted peptide identifications were stringently filtered to an estimated FDR level of 0.0% using the target-decoy method for estimation. S2 Appendix contains all information on identified endogenous peptides.
Bioinformatics analyses
The semi-quantitative protein data from the dog saliva was measured via protein spectral counts from the MS-based proteomics data. Normalized spectral counts were assigned to each identified protein using the “Quantitative Value” assignment tool within the Scaffold software used for organizing protein identifications and comparing spectral counts across samples. Quantitative values for identified proteins and endogenous peptides were compared across the four groups.
To be included in the analyses each specific protein and peptide had to meet at least one of the following criteria: (i) an assigned normalized spectral count value of 5 or higher in at least one group or (ii) a normalized spectral count value of 1 or more in at least two groups. After application of the criteria in the dataset, the gene symbols of included proteins were imported into a web-based program (Venny 2.1) for construction of Venn’s diagram [41] with the goal of comparing the protein and peptide content among groups. A two dimensional Principle Component Analysis (PCA) and Heatmap were also generated using a web tool for visualizing multivariate data (ClustVis) [42] to further evaluate similarities and differences in the salivary proteomics and peptidomics among the four groups.
Search of proteins and peptides with immune functions
Papers on salivary proteomics/peptidomics were reviewed to identify proteins and peptides with antimicrobial functions that are reported to be abundant in human saliva (specific proteins and references are provided in the results); our database was then searched for these proteins and peptides.
Results
Dog saliva proteomic and peptidomic profile
Using nanoscale LC MS/MS 2,491 proteins and endogenous peptides were identified in the dog saliva (S3 Appendix), and 1,588 of those met the defined quantitative criteria for further analysis. The top 10 most abundant proteins and their function are provided in Table 1; 7 of the 10 have immune functions. Table 2 additionally shows salivary proteins and peptides with various antimicrobial properties that have been reported to be abundant in previous studies. Most of these proteins and peptides were present in all four groups.
Table 1. Most abundant canine salivary proteins and their functions.
| Protein name | Gene name | Total normalized spectral counts | Function [reference] |
|---|---|---|---|
| Fc fragment of IgG binding protein | FCGBP | 797 | Binds to IgG on mucosal surfaces [43] |
| Polymeric immunoglobulin receptor (precursor) | PIGR | 571 | Transports IgA across epithelial cells [44] |
| BPI fold-containing family A member 2 | BPIFA2 | 466 | Key components of the innate immune response against Gram-negative bacteria [45] |
| BPI fold-containing family B member 1 isoform X2 | BPIFB1 | 352 | Key components of the innate immune response against Gram-negative bacteria [45] |
| Albumin | ALB | 296 | Serum-derived protein believed to passively enter saliva. Saliva-specific functions include binding to hydroxyapatite and lubrication of oral tissues [46] |
| Ovostatin homolog 2-like | LOC611455 | 270 | GO: serine-type endopeptidase inhibitor activity |
| Mucin 19 | MUC19 | 264 | Gel-forming mucin that lubricates saliva and plays a role in reducing adherence and increasing clearance of bacteria [47] |
| Angiopoietin-related protein 5-like | LOC607055 | 261 | Not reported |
| Actin gamma 1 | ACTG1 | 244 | Cytoskeletal protein with multiple functions in the defense against intracellular pathogens [48] |
| Ig lambda chain V-I region BL2 | LOC607368 | 219 | Component of immunoglobulin light chains |
Table 2. Canine salivary proteins and peptides with antimicrobial properties.
| Protein name | Gene name | Normalized spectral counts per dog group* | Function [reference] |
|---|---|---|---|
| Antileukoproteinase or secretory leukocyte protease inhibitor (precursor) | SLPI | 4-2-2-1 | The N-terminal cationic domain has an antibacterial, antifungal and antiviral effect [49–51] |
| Beta 2 microglobulin (precursor) | B2M | 0-1-2-2 | Agglutinates bacteria (e.g. Streptococcus mutans) [50–52] |
| BPI fold-containing family A member 2 | BPIFA2 | 41-136-103-186 | Key components of the innate immune response against Gram-negative bacteria [45] |
| BPI fold-containing family B member 1 isoform X2 | BPIFB1 | 42-96-67-147 | Key components of the innate immune response against Gram-negative bacteria [45] |
| Carbonic anhydrase 6 isoform X1 | CA6 | 16-42-50-69 | Binds to Staphylococcus aureus [53] |
| Cathelicidin antimicrobial peptide (precursor) | CAMP | 1-1-1-2 | Antibacterial and antifungal effects by disruption of cell membrane. It also binds and neutralizes lipopolysaccharide from Gram-negative bacteria [50,54,55] |
| Cystatin-M | CST6 | 2-7-9-8 | Cystatins block the action of bacterial proteases [49,50,56] |
| Cystatin-A | CSTA | 2-2-0-1 | |
| Deleted in malignant brain tumors 1 protein isoform X1 | DMBT1 | 7-27-27-44 | Known as salivary agglutinin and is identical to Gp-340 expressed in lungs. Binds to a wide variety of microorganisms [49,50,53,57] |
| Elafin/skin-derived antileukoproteinase (SKALP) (precursor) | PI3 | 1-1-0-0 | Kills Gram-negative and Gram-positive bacteria [50,58] |
| Fibronectin (partial, predicted) | FN1 | 6-12-20-8 | Agglutinates bacteria and prevents its adhesion to oral surfaces [50,57] |
| Immunoglobulin J chain isoform 1 (predicted) | IGJ | 0-3-10-4 | Binds to Staphylococcus aureus [53] |
| Lactotransferrin (Precursor) | LTF | 8-43-52-80 | Bacteriostatic due to its iron-depriving effects [49,50] |
| Lactoperoxidase isoform 2 | LPO | 3-41-37-63 | Catalysis the formation of bactericidic compounds [49,50] |
| Lysozyme C, milk isozyme-like (predicted) | LYZF2 | 3-55-72-63 | Defense response to bacterium; regulation of macrophage activation; lysis bacteria cell wall polysaccharides; activates bacterial autolysins [49,50,53] |
| Lysozyme precursor (cluster) | LYZ | 28-24-19-28 | |
| Mucin-5B (predicted) | MUC5B | 14-46-69-72 | Modulates the microbial colonization of oral epithelial surfaces [49] |
| Mucin-7 (predicted) | MUC7 | 0-0-1-2 | Binds to a variety of bacteria [49,50,59–61] |
| Mucin-19 (predicted) | MUC19 | 0-65-93-106 | Gel-forming mucin that lubricates saliva and plays a role in reducing adherence and increasing clearance of bacteria [47] |
| Myeloperoxidase | MPO | 6-3-7-4 | Catalyses the hydrogen peroxide oxidation of thiocynate ions which forms the bactericidal product, hypothiocyanite [50] |
| Peptidoglycan recognition protein 1 | PGLYRP1 | 4-3-2-2 | It binds to the bacterial cell wall peptidoglycans to exert the bactericidal effect, but do not permeabilize bacterial membranes. They are bactericidal for Gram-positive bacteria and bacteriostatic for Gram-negative bacteria [50,62,63] |
| Polymeric immunoglobulin receptor (precursor) | PIGR | 2-164-204-201 | Transports IgA across epithelial cells [44] |
| Serpin B10 (predicted) | SERPINB10 | 3-8-9-14 | Positive regulation of defense response to virus by host |
| S100-A8 | S100A8 | 2-5-5-11 | Also known as calgranulin A (S100-A8) and B (S100-A9). The dimer of calgranulin A and B is called calprotectin is expressed in neutrophils, macrophages and keratinocytes cytosols. They inhibit bacterial growth by scavenging divalent cation [50,64] |
| S100-A9 isoformX3 | S100A9 | 2-4-1-8 | |
| Zymogen granule protein 16 homolog B | ZG16B | 1-5-6-15 | Binds to Staphylococcus aureus [53] |
* Groups are sequentially as follows: Ancient, Herding-Sighthound, Miscellaneous-1,Miscellaneous-2
The salivary proteomic and peptidomic content of the Ancient group stands out from other breed groups
Comparative analyses using a Venn diagram showed that the four groups shared 614 proteins and peptides representing 38.7% of the analyzed proteins/peptides (Fig 2). In contrast to the other three groups, unique proteins (SPTBN2 (normalized spectral count = 7), TMOD3 (6) and PSMC4 (5)) were identified only in the Ancient group. In addition, there were 245 (15.5%) proteins that were shared by the Herding-Sighthound and Miscellaneous 1 and 2 groups, but not the Ancient group (Fig 2).
Fig 2. Venn diagram displaying the overlapped and unique proteins among the four groups.
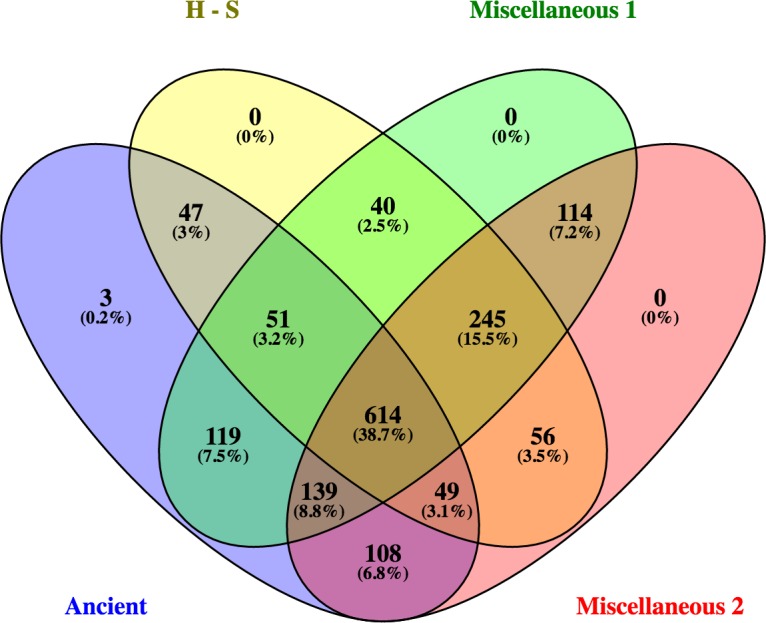
H-S = Herding-Sighthound.
The unique protein profile in the Ancient group was also reflected in the PCA analysis (Fig 3). The Ancient group nested distant from the other three groups at the far end of PC1 (X-axis), which explained 47.6% of the total variance. When taking into consideration PC2 (Y-axis), which explained 26.8% of total variance, Herding-Sighthound and Miscellaneous group 2 clustered more closely than Miscellaneous group 1 and Ancient group.
Fig 3. Two dimensional principal coordinate analysis showing the salivary protein and peptide profile relationship among the groups.

The intergroup relationship in the PCA analysis is also confirmed by the dendrogram in Fig 4, which shows the relationship of the four groups. The predominance of blue color in the Ancient group indicates relatively less abundance of specific proteins and peptides in this group compared to the other groups.
Fig 4. Heatmap showing the relative abundance (color) and relationship (dendogram) of salivary proteins and peptides among the groups.

Discussion
Using nanoscale LC–MS/MS we identified 2,491 proteins and peptides in the saliva of healthy dogs with no periodontal diseases. In contrast, a recent study described only 244 proteins in dog saliva [3]. The substantially higher number of proteins identified in this study could be partially explained by differences in sample collection but a main contributor is most likely the mass spectrometer instrumentation used. For our study, we used a sophisticated mass spectrometer (Orbitrap Fusion Tribrid with Easy-nLC autosampler and LC (Thermo Scientific, Waltham, MA) for LC-MS/MS analysis which provides some of the highest sensitivity currently available for analysis of complex protein mixtures [65]. The prior study by de Sousa-Pereira and colleagues utilized an older generation MALDI-TOF/TOF instrument [3], which most likely explains the order of magnitude difference in proteins identified. It also highlights the depth of our study in terms of proteins and endogenous peptides identified, which provides a much more comprehensive view of the salivary proteome and peptidome in healthy dogs.
One of the most important functions of saliva is to protect the oral cavity and indirectly other organs against infections. In this study, 7 of the top 10 most abundant proteins have immune functions. Additionally, we identified 26 peptides and proteins (as well as some isoforms) that have been reported to have antimicrobial functions in human saliva; 4 of these were also in the top 10 most abundant in canine saliva. Six of the 26 proteins and peptides were not present in all four breed groups indicating the variability among individual dogs or dog breeds. There are likely many additional proteins and peptides with antimicrobial functions in the 2,491 identified in the study.
Antimicrobial peptides (AMP) are small molecular weight, typically cationic peptides that have a broad spectrum of action against bacteria, fungi, parasites and some viruses [66]. They are an important part of the innate immune response of almost all living organisms including plants, invertebrates and vertebrates and generally function by forming holes in the microorganisms’ cell membrane. Various AMP such as, alfa and beta defensins, cathelicidin, adrenomedullin, histatins, elafin, secretory leukocyte protease inhibitor (SLPI) and lysozyme have been found in human saliva [49,50,67]. Of these, we identified precursors of elafin, SLPI, cathelicidin and lysozyme and, de Sousa-Pereira and collaborators listed lysozyme in the saliva of the dog included in their study [3]. β-defensins are expressed in many epithelial tissues and have been identified in the skin of healthy dogs [68–70]. The absence of this category of AMP in the dog saliva was, to some extent, unexpected but, defensins were not reported in the saliva of dog, cattle, sheep, horse, rabbit and rat in a recent study [3]. Moreover, the lack of identification could be explained by their presence below the limit of detection of the method used. Differences in the expression of proteins and peptides in the saliva of humans and dogs could be partially explained by phylogenetic and dietary variations between these species. However, additional studies including a large number of dogs will be needed to corroborate our findings.
The dogs in this study were selected to represent diverse ancestral lineages both to provide a comprehensive dataset of the canine salivary proteome and peptidome and to determine if there were clear differences between breed groups. While only two of the four groups ultimately represented distinct genetic clusters based on the most recent canine genomics data, differences were evident, with only 38.7% of the analyzed proteins and peptides shared by all groups. The Ancient group, which included Siberian Huskies and Alaskan Malamutes, was the most distinct, and the only one with unique proteins. This parallels genetic differences in the breed groups; the breeds within the ancient group have a high level of divergence from other breeds. It cannot be determined from this data whether the unique proteins identified in the Ancient group are specifically characteristic of the northern group (the clade comprising the Siberian Husky and Alaskan Malamute) or if they are also a feature of saliva from other ancient breeds [36]. Future studies including a larger breed representation in the various phylogenic groups and a larger number of dogs per breed will help answer this question. Interestingly, a recent study showed unique proteins in the saliva of Korean people when compared to a comprehensive database of human salivary proteins indicating ethnic differences in the human saliva proteome [71].
In addition to genetic differences, other factors not investigated in this study could have also played a role in variations noted between the dog groups. Age, diurnal variation, health status and individual variation have all been shown to influence the composition of proteins in the saliva in humans [72–78]. These variables, and possibly others, most likely also impact the salivary protein and peptide profiles of dogs and need to be carefully and urgently investigated before we can obtain accurate information on changes in saliva protein and peptide components in disease states.
This study provides a comprehensive catalog of the proteins and endogenous peptides present in canine saliva. We included 36 dogs and divided them in groups based on breed phylogeny which revealed differences that parallel genetic clusters. Samples were pooled within each group, and this could have masked any inter-individual or gender variances in the salivary protein composition of the dogs. An important next step is to evaluate any possible influence of age, gender, breed and individual in the composition of proteins and peptides of the dog saliva.
Supporting information
(XLSX)
(XLSX)
(XLSX)
Acknowledgments
The authors thank Dr. Kevin Stepaniuk for helping with sample collection. The authors also thank the Center for Mass Spectrometry and Proteomics at the University of Minnesota for assistance with data generation and analysis, and maintenance of required instrumentation.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
The work was self-funded by the corresponding author. Partial funding for E.F. was provided by the Office of the Director, National Institute of Health (NIH) under award number K01OD019912. The funder had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Castagnola M, Cabras T, Vitali A, Sanna MT, Messana I. Biotechnological implications of the salivary proteome. Trends Biotechnol 2011; 29(8): 409–418. doi: 10.1016/j.tibtech.2011.04.002 [DOI] [PubMed] [Google Scholar]
- 2.Guo T, Rudnick PA, Wang W, Lee CH, DeVoe DL, Balgley BM. Characterization of the human salivary proteome by capillary isoelectric focusing/nanoreversed-phase liquid chromatography coupled with ESI-tandem MS. J Proteome Res 2006; 5(6):1469–1478. doi: 10.1021/pr060065m [DOI] [PubMed] [Google Scholar]
- 3.de Sousa-Pereira P, Cova M, Abrantes J, Ferreira R, Trindade F, Barros A et al. Cross-species comparison of mammalian saliva using an LC-MALDI based proteomic approach. Proteomics 2015; 15(9):1598–1607. doi: 10.1002/pmic.201400083 [DOI] [PubMed] [Google Scholar]
- 4.Lima DP, Diniz DG, Moimaz SAS, Sumida DH, Okamoto AC. Saliva: reflection of the body. Int J Infect Dis 2010;14(3): e184–188. doi: 10.1016/j.ijid.2009.04.022 [DOI] [PubMed] [Google Scholar]
- 5.Al-Tarawneh SK, Border MB, Dibble CF, Bencharit S. Defining salivary biomarkers using mass spectrometry-based proteomics: a systematic review. OMICS 2011; 15:353–361. doi: 10.1089/omi.2010.0134 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Katsiougiannis S, Wong DTW. The Proteomics of Saliva in Sjogren's Syndrome. Rheumatic Disease Clinics 2016; 42(3): 449–456. doi: 10.1016/j.rdc.2016.03.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Podzimek S, Vondrackova L, Duskova J, Janatova T, Broukal Z. Salivary markers for periodontal and general diseases. Dis Markers 2016; (2016): 9179632 doi: 10.1155/2016/9179632 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Xie H, Rhodus NL, Griffin RJ, Carlis JV, Griffin TJ. A catalogue of human saliva proteins identified by free flow electrophoresis-based peptide separation and tandem mass spectrometry. Mol Cell Proteomics 2005; 4(11): 1826–1830. doi: 10.1074/mcp.D500008-MCP200 [DOI] [PubMed] [Google Scholar]
- 9.Zhang Y, Sun J, Lin CC, Abemayor E, Wang MB, Wong DT. The emerging landscape of salivary diagnostics. Periodontol 2000 2016; 70(1): 38–52. doi: 10.1111/prd.12099 [DOI] [PubMed] [Google Scholar]
- 10.Camisasca DR, Gonçalves LDR, Soares MR, Sandimb V, Nogueira FCS, Garcia CHS, et al. A proteomic approach to compare saliva from individuals with and without oral leukoplakia. J Proteomics. 2017; 151: 43±52. https://doi.org/10.1016/j.jprot.2016.07.029 [DOI] [PubMed] [Google Scholar]
- 11.Cargile BJ, Talley DL, Stephenson JL Jr. Immobilized pH gradients as a first dimension in shotgun proteomics and analysis of the accuracy of pI predictability of peptides. Electrophoresis 2004; 25(6): 936–945. doi: 10.1002/elps.200305722 [DOI] [PubMed] [Google Scholar]
- 12.Csősz É, Kalló G, Márkus B, Deák E, Csutak A, Tőzsér J. Quantitative body fluid proteomics in medicine—A focus on minimal invasiveness. J Proteomics. 2017; 153: 30±43. https://doi.org/10.1016/j.jprot.2016.08.009 [DOI] [PubMed] [Google Scholar]
- 13.Ghafouri B, Tagesson C, Lindahl M. Mapping of proteins in human saliva using two-dimensional gel electrophoresis and peptide mass fingerprinting. Proteomics 2003; 3(6): 1003–1015. doi: 10.1002/pmic.200300426 [DOI] [PubMed] [Google Scholar]
- 14.Hardt M, Thomas LR, Dixon SE, Newport G, Agabian N, Prakobphol A et al. Toward defining the human parotid gland salivary proteome and peptidome: identification and characterization using 2D SDS-PAGE, ultrafiltration, HPLC, and mass spectrometry. Biochemistry 2005; 44(8): 2885–2899. doi: 10.1021/bi048176r [DOI] [PubMed] [Google Scholar]
- 15.Hirtz C, Chevalier F, Centeno D, Egea JC, Rossignol M, Sommerer N et al. Complexity of the human whole saliva proteome. J Physiol Biochem 2005; 61(3): 469–480. doi: 10.1007/BF03168453 [DOI] [PubMed] [Google Scholar]
- 16.Hu S, Xie Y, Ramachandran P, Ogorzalek Loo RR, Li Y, Loo JA et al. Large-scale identification of proteins in human salivary proteome by liquid chromatography/mass spectrometry and two-dimensional gel electrophoresis-mass spectrometry. Proteomics 2005; 5(6):1714–1728. doi: 10.1002/pmic.200401037 [DOI] [PubMed] [Google Scholar]
- 17.Jacobs R, Maasdorp E, Malherbe S, Loxton AG, Kim S, der Spuy GV, et al. Diagnostic potential of novel salivary host biomarkers as candidates for the immunological diagnosis of tuberculosis disease and monitoring of tuberculosis treatment response. PLOS ONE. 2016; 11: e0160546 https://doi.org/10.1371/journal.pone.0160546 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Loseva OI, Gavryushkin AV, Osipov VV, Vanyakin EN. Application of free-flow electrophoresis for isolation and purification of proteins and peptides. Electrophoresis 1998; 19(7): 1127–1134. doi: 10.1002/elps.1150190712 [DOI] [PubMed] [Google Scholar]
- 19.Mireya GB, Lu B, Liao L, Xu T, Bedi G, Melvin JE, et al. Characterization of the human submandibular/sublingual saliva glycoproteome using lectin affinity chromatography coupled to multidimensional protein identification technology. J Proteome Res. 2011; 10: 5031±5046. https://doi.org/10.1021/pr200505t [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Moritz RL, Ji H, Schutz F, Connolly LM, Kapp EA, Speed TP et al. A proteome strategy for fractionating proteins and peptides using continuous free-flow electrophoresis coupled off-line to reversed-phase high-performance liquid chromatography. Anal Chem 2004; 76(16): 4811–4824. doi: 10.1021/ac049717l [DOI] [PubMed] [Google Scholar]
- 21.Pfaffe T, Justin CW, Beyerlein P, Konstner K, Punyadeera C. Diagnostic potential of saliva: current state and future applications. Clin Chem. 2011; 57: 675±687. https://doi.org/10.1373/clinchem.2010.153767 [DOI] [PubMed] [Google Scholar]
- 22.Sivadasan P, Gupta MK, Sathe GJ, Balakrishnan L, Palit P, Gowda H, et al. Human salivary proteome—a resource of potential biomarkers for oral cancer. J Proteomics. 2015; 127: 89±95. https://doi.org/10.1016/j.jprot.2015.05.039 [DOI] [PubMed] [Google Scholar]
- 23.Vitorino R, Lobo MJ, Ferrer-Correira AJ, Dubin JR, Tomer KB, Domingues PM et al. Identification of human whole saliva protein components using proteomics. Proteomics 2004; 4(4): 1109–1115. doi: 10.1002/pmic.200300638 [DOI] [PubMed] [Google Scholar]
- 24.Wang Q, Yu Q, Lin Q, Duan Y. Emerging salivary biomarkers by mass spectrometry. Clin Chim Acta. 2015; 438: 214±221. https://doi.org/10.1016/j.cca.2014.08.037 [DOI] [PubMed] [Google Scholar]
- 25.Wilmarth PA, Riviere MA, Rustvold DL, Lauten JD, Madden TE, David LL. Two-dimensional liquid chromatography study of the human whole saliva proteome. J Proteome Res 2004; 3(5): 1017–1023. doi: 10.1021/pr049911o [DOI] [PubMed] [Google Scholar]
- 26.Yao Y, Berg EA, Costello CE, Troxier RF, Oppenheim FG. Identification of protein components in human acquired enamel pellicle and whole saliva using novel proteomics approaches. J Biol Chem 2003; 278(7): 5300–5308. doi: 10.1074/jbc.M206333200 [DOI] [PubMed] [Google Scholar]
- 27.Alterman A, Mathison R, Coronel CE, Stroppa MM, Finkelberg AB, Gallara RV. Functional and proteomic analysis of submandibular saliva in rats exposed to chronic stress by immobilization or constant light. Arch Oral Biol 2012; 57(6): 663–669. doi: 10.1016/j.archoralbio.2011.12.008 [DOI] [PubMed] [Google Scholar]
- 28.Ang CS, Binos S, Knight MI, Moate PJ, Cocks BG, McDonagh MB. Global survey of the bovine salivary proteome: integrating multidimensional prefractionation, targeted, and glycocapture strategies. J Proteome Res 2011; 10(11): 5059–5069. doi: 10.1021/pr200516d [DOI] [PubMed] [Google Scholar]
- 29.de Sousa-Pereira P, Abrantes J, Pinheiro A, Colaco B, Vitorino R, Esteves PJ. Evolution of C, D and S-type cystatins in mammals: an extensive gene duplication in primates. PLoS One 2014; 9(10): e109050 doi: 10.1371/journal.pone.0109050 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Gutierrez AM, Miller I, Hummel K, Nobauer K, Martinez-Subiela S, Razzazi-Fazeli E et al. Proteomic analysis of porcine saliva. Vet J 2011; 187(3): 356–362. doi: 10.1016/j.tvjl.2009.12.020 [DOI] [PubMed] [Google Scholar]
- 31.Hornshoj H, Bendixen E, Conley LN, Andersen PK, Hedegaard J, Panitz F. et al. Transcriptomic and proteomic profiling of two porcine tissues using high-throughput technologies. BMC Genomics 2009; 10:30 doi: 10.1186/1471-2164-10-30 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lamy E, da Costa G, Santos R, Capela e Silva F, Potes J, Pereira A et al. Effect of condensed tannin ingestion in sheep and goat parotid saliva proteome. J Anim Physiol Anim Nutr (Berl) 2011;95:304–312. doi: 10.1111/j.1439-0396.2010.01055.x [DOI] [PubMed] [Google Scholar]
- 33.Lamy E, Costa G, Santos R, Capela e Silva F, Potes J, Pereira A et al. Sheep and goat saliva proteome analysis: a useful tool for ingestive behavior research? Physiol Behav 2009; 98(4): 393–401. doi: 10.1016/j.physbeh.2009.07.002 [DOI] [PubMed] [Google Scholar]
- 34.Lamy E, Graca G, Costa G, Franco C, Silva FC, Baptista ES et al. Changes in mouse whole saliva soluble proteome induced by tannin-enriched diet. Proteome Sci 2010; 8:65 doi: 10.1186/1477-5956-8-65 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ostrander EA, Wayne RK. The canine genome. Genome Res 2005; 15(12): 1706–1716. doi: 10.1101/gr.3736605 [DOI] [PubMed] [Google Scholar]
- 36.Vonholdt BM, Pollinger JP, Lohmueller KE, Han E, Parker HG, Quignon P et al. Genome-wide SNP and haplotype analyses reveal a rich history underlying dog domestication. Nature 2010; 464(7290): 898–902. doi: 10.1038/nature08837 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Hekman JP, Karas AZ, Dreschel NA. Salivary cortisol concentrations and behavior in a population of healthy dogs hospitalized for elective procedures. Appl Anim Behav Sci 2012; 141(3–4): 149–157. doi: 10.1016/j.applanim.2012.08.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Wisniewski JR, Zougman A, Nagaraj N, Mann N. Universal sample preparation method for proteome analysis. Nat Methods 2009; 6(5): 359–362. doi: 10.1038/nmeth.1322 [DOI] [PubMed] [Google Scholar]
- 39.Wang Y, Yang F, Gritsenko MA, Wang Y, Clauss T, Liu T. Reversed-phase chromatography with multiple fraction concatenation strategy for proteome profiling of human MCF10A cells. Proteomics 2011; 11(10): 2019–2026. doi: 10.1002/pmic.201000722 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Nesvizhskii AI, Keller A, Kolker E, Aebersold R. A statistical model for identifying proteins by tandem mass spectrometry. Anal Chem 2003; 75(17): 4646–4658. [DOI] [PubMed] [Google Scholar]
- 41.Oliveros J. VENNY. An interactive tool for comparing lists with Venn Diagrams, 2007. Available from: http://bioinfogp.cnb.csic.es/tools/venny/index.html
- 42.Metsalu T, Vilo J. ClustVis: a web tool for visualizing clustering of multivariate data using principal component analysis and heatmap. Nucleic Acids Res 2015; 43(W1): W566–570. doi: 10.1093/nar/gkv468 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kobayashi K, Ogata H, Morikawa M, Ilijima S, Harada N, Yoshida T, et al. Distribution and partial characterisation of IgG Fc binding protein in various mucin producing cells and body fluids. Gut 2002; 51(2): 169–176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Asano M, Komiyama K. Polymeric immunoglobulin receptor. J Oral Sci 2011; 53(2): 147–156. doi: 10.2334/josnusd.53.147 [DOI] [PubMed] [Google Scholar]
- 45.Bingle L, Barnes FA, Lunn H, Musa M, Webster S, Ian Douglas CW, et al. Characterisation and expression of SPLUNC2, the human orthologue of rodent parotid secretory protein. Histochem Cell Biol 2009; 132(3): 339–349. doi: 10.1007/s00418-009-0610-4 [DOI] [PubMed] [Google Scholar]
- 46.Schenkels LC, Veerman EC, Nieuw Amerongen AV. Biochemical composition of human saliva in relation to other mucosal fluids. Crit Rev Oral Biol Med 1995; 6(2): 161–175. [DOI] [PubMed] [Google Scholar]
- 47.Culp DJ, Robinson B, Cash MN, Bhattacharyva I, Stewart C, Cuadra-Saenz G. Salivary mucin 19 glycoproteins: innate immune functions in Streptococcus mutans-induced caries in mice and evidence for expression in human saliva. J Biol Chem 2015; 290(5): 2993–3008. doi: 10.1074/jbc.M114.597906 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Mostowy S, Shenoy AR. The cytoskeleton in cell-autonomous immunity: structural determinants of host defence. Nat Rev Immunol 2015; 15(9): 559–573. doi: 10.1038/nri3877 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Amerongen AV, Veerman EC. Saliva—the defender of the oral cavity. Oral Dis 2002; 8(1): 12–22. doi: 10.1034/j.1601-0825.2002.1o816.x [DOI] [PubMed] [Google Scholar]
- 50.Gorr SU. Antimicrobial peptides of the oral cavity. Periodontol 2000 2009; 51(:152–180. doi: 10.1111/j.1600-0757.2009.00310.x [DOI] [PubMed] [Google Scholar]
- 51.McNeely TB, Dealy M, Dripps DJ, Orenstein JM, Eisenberg SP, Wahl SM. Secretory leukocyte protease inhibitor: a human saliva protein exhibiting anti-human immunodeficiency virus 1 activity in vitro. J Clin Invest 1995; 96(1): 456–464. doi: 10.1172/JCI118056 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Ericson D. Agglutination of Streptococcus mutans by low-molecular-weight salivary components: effect of beta 2-microglobulin. Infect Immun 1984; 46(2): 526–530. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Heo SM, Choi KS, Kazim LA, Reddy MS, Haase EM, Scannapieco FA, et al. Host defense proteins derived from human saliva bind to Staphylococcus aureus. Infect Immun 2013; 81(4): 1364–1373. doi: 10.1128/IAI.00825-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.den Hertog AL, van Marle J, van Veen HA, Van’t Hof W, Boscher JG, Veerman EC, et al. Candidacidal effects of two antimicrobial peptides: histatin 5 causes small membrane defects, but LL-37 causes massive disruption of the cell membrane. Biochem J 2005; 388(Pt 2): 689–695. doi: 10.1042/BJ20042099 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Ji S, Hyun J, Park E, Lee BL, Kim KK, Choi Y. Susceptibility of various oral bacteria to antimicrobial peptides and to phagocytosis by neutrophils. J Periodontal Res 2007; 42(5): 410–419. doi: 10.1111/j.1600-0765.2006.00962.x [DOI] [PubMed] [Google Scholar]
- 56.de Sousa-Pereira P, Amado F, Abrantes J, Ferreira R, Esteves PJ, Vitorino R An evolutionary perspective of mammal salivary peptide families: cystatins, histatins, statherin and PRPs. Arch Oral Biol 2013; 58(5): 451–458. doi: 10.1016/j.archoralbio.2012.12.011 [DOI] [PubMed] [Google Scholar]
- 57.Scannapieco FA. Saliva-bacterium interactions in oral microbial ecology. Crit Rev Oral Biol Med 1994; 5(3–4): 203–248. [DOI] [PubMed] [Google Scholar]
- 58.Simpson AJ, Maxwell AI, Govan JR, Haslett C, Sallenave JM. Elafin (elastase-specific inhibitor) has anti-microbial activity against gram-positive and gram-negative respiratory pathogens. FEBS Lett 1999; 452(3): 309–313. [DOI] [PubMed] [Google Scholar]
- 59.Groenink J, Ligtenberg AJ, Veerman EC, Bolscher JG, Nieuw Amerongen AV. Interaction of the salivary low-molecular-weight mucin (MG2) with Actinobacillus actinomycetemcomitans. Antonie Van Leeuwenhoek 1996; 70(1): 79–87. [DOI] [PubMed] [Google Scholar]
- 60.Moshier A, Reddy MS, Scannapieco FA. Role of type 1 fimbriae in the adhesion of Escherichia coli to salivary mucin and secretory immunoglobulin A. Curr Microbiol 1996; 33(3): 200–208. [DOI] [PubMed] [Google Scholar]
- 61.Reddy MS, Levine MJ, Paranchych W. Low-molecular-mass human salivary mucin, MG2: structure and binding of Pseudomonas aeruginosa. Crit Rev Oral Biol Med 1993; 4(3–4): 315–323. [DOI] [PubMed] [Google Scholar]
- 62.Lu X, Wang M, Qi J, Wang H, Li X, Gupta D, et al. Peptidoglycan recognition proteins are a new class of human bactericidal proteins. J Biol Chem 2006; 281(9): 5895–5907. doi: 10.1074/jbc.M511631200 [DOI] [PubMed] [Google Scholar]
- 63.Royet J, Dziarski R. Peptidoglycan recognition proteins: pleiotropic sensors and effectors of antimicrobial defences. Nat Rev Microbiol 2007;5(4): 264–277. doi: 10.1038/nrmicro1620 [DOI] [PubMed] [Google Scholar]
- 64.Donato R, Cannon BR, Sorci G, Riuzzi F, Hsu K, Weber DJ, et al. Functions of S100 proteins. Curr Mol Med 2013; 13(1): 24–57. doi: 10.2174/156652413804486214 [PMC free article] [PubMed] [Google Scholar]
- 65.Hebert AS, Richards AL, Bailey DJ, Ulbrich A, Coughlin EE, Westphall MS, et al. The one hour yeast proteome. Mol Cell Proteomics 2014; 13(1): 339–347. doi: 10.1074/mcp.M113.034769 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.da Costa JP, Cova M, Ferreira R, Vitorino R. Antimicrobial peptides: an alternative for innovative medicines? Appl Microbiol Biotechnol 2015; 99(5): 2023–2040. doi: 10.1007/s00253-015-6375-x [DOI] [PubMed] [Google Scholar]
- 67.Khurshid Z, Naseem M, Sheikh Z, Najeeb S, Shahab S, Zafar MS. Oral antimicrobial peptides: Types and role in the oral cavity. Saudi Pharm J 2016; 24(5): 515–524. doi: 10.1016/j.jsps.2015.02.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Leonard BC, Marks SL, Outerbridge CA, Affolter VK, Kananurak A, Young A, et al. Activity, expression and genetic variation of canine beta-defensin 103: a multifunctional antimicrobial peptide in the skin of domestic dogs. J Innate Immun 2012; 4(3): 248–259. doi: 10.1159/000334566 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Santoro D, Bunick D, Graves TK, Campbell KL. Expression and distribution of antimicrobial peptides in the skin of healthy beagles. Vet Dermatol 2011; 22(1): 61–67. doi: 10.1111/j.1365-3164.2010.00911.x [DOI] [PubMed] [Google Scholar]
- 70.Wingate KV, Torres SM, Silverstein KA, Hendrickson JA, Rutherford MS. Expression of endogenous antimicrobial peptides in normal canine skin. Vet Dermatol 2009; 20(1): 19–26. doi: 10.1111/j.1365-3164.2008.00707.x [DOI] [PubMed] [Google Scholar]
- 71.Cho HR, Kim HS, Park JS, Park SC, Kim KP, Wood TD, et al. Construction and characterization of the Korean whole saliva proteome to determine ethnic differences in human saliva proteome. PLoS ONE 2017; 12: e0181765 https://doi.org/10.1371/journal.pone.0181765 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Cabras T, Pisano E, Boi R, Olianas A, Manconi B, Inzitari R, et al. Age-dependent modifications of the human salivary secretory protein complex. J Proteome Res 2009; 8(8): 4126–4134. doi: 10.1021/pr900212u [DOI] [PubMed] [Google Scholar]
- 73.Hardt M, Witkowska HE, Webb S, Thomas LR, Dixon SE, Hall SC, et al. Assessing the effects of diurnal variation on the composition of human parotid saliva: quantitative analysis of native peptides using iTRAQ reagents. Anal Chem 2005; 77(15): 4947–4954. doi: 10.1021/ac050161r [DOI] [PubMed] [Google Scholar]
- 74.Jehmlich N, Dinh KH, Gesell-Salazar M, Hammer E, Stell L, Dhople VM, et al. Quantitative analysis of the intra- and inter-subject variability of the whole salivary proteome. J Periodontal Res 2013; 48(3): 392–403. doi: 10.1111/jre.12025 [DOI] [PubMed] [Google Scholar]
- 75.Manconi B, Cabras T, Pisano E, et al. Modifications of the acidic soluble salivary proteome in human children from birth to the age of 48months investigated by a top-down HPLC-ESI-MS platform. J Proteomics 2013;91:536–543. doi: 10.1016/j.jprot.2013.08.009 [DOI] [PubMed] [Google Scholar]
- 76.Messana I, Cabras T, Iavarone F, Manconi B, Huang L, Martelli C, et al. Chrono-proteomics of human saliva: variations of the salivary proteome during human development. J Proteome Res 2015;14(4): 1666–1677. doi: 10.1021/pr501270x [DOI] [PubMed] [Google Scholar]
- 77.Messana I, Inzitari R, Fanali C, Cabras T, Castagnola M. Facts and artifacts in proteomics of body fluids. What proteomics of saliva is telling us? J Sep Sci 2008; 31(11): 1948–1963. doi: 10.1002/jssc.200800100 [DOI] [PubMed] [Google Scholar]
- 78.Quintana M, Palicki O, Lucchi G, Ducoroy P, Chambon C, Salles C, et al. Inter-individual variability of protein patterns in saliva of healthy adults. J Proteomics 2009; 72(5): 822–830. doi: 10.1016/j.jprot.2009.05.004 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(XLSX)
(XLSX)
(XLSX)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.