

Abstract
Rising seawater temperature and ocean acidification threaten the survival of coral reefs. The relationship between coral physiology and its microbiome may reveal why some corals are more resilient to these global change conditions. Here, we conducted the first experiment to simultaneously investigate changes in the coral microbiome and coral physiology in response to the dual stress of elevated seawater temperature and ocean acidification expected by the end of this century. Two species of corals, Acropora millepora containing the thermally sensitive endosymbiont C21a and Turbinaria reniformis containing the thermally tolerant endosymbiont Symbiodinium trenchi, were exposed to control (26.5°C and pCO2 of 364 μatm) and treatment (29.0°C and pCO2 of 750 μatm) conditions for 24 days, after which we measured the microbial community composition. These microbial findings were interpreted within the context of previously published physiological measurements from the exact same corals in this study (calcification, organic carbon flux, ratio of photosynthesis to respiration, photosystem II maximal efficiency, total lipids, soluble animal protein, soluble animal carbohydrates, soluble algal protein, soluble algal carbohydrate, biomass, endosymbiotic algal density, and chlorophyll a). Overall, dually stressed A. millepora had reduced microbial diversity, experienced large changes in microbial community composition, and experienced dramatic physiological declines in calcification, photosystem II maximal efficiency, and algal carbohydrates. In contrast, the dually stressed coral T. reniformis experienced a stable and more diverse microbiome community with minimal physiological decline, coupled with very high total energy reserves and particulate organic carbon release rates. Thus, the microbiome changed and microbial diversity decreased in the physiologically sensitive coral with the thermally sensitive endosymbiotic algae but not in the physiologically tolerant coral with the thermally tolerant endosymbiont. Our results confirm recent findings that temperature-stress tolerant corals have a more stable microbiome, and demonstrate for the first time that this is also the case under the dual stresses of ocean warming and acidification. We propose that coral with a stable microbiome are also more physiologically resilient and thus more likely to persist in the future, and shape the coral species diversity of future reef ecosystems.
Introduction
Increased atmospheric carbon dioxide (CO2) is causing the oceans to warm and become more acidic, thus resulting in lower seawater pH and carbonate mineral saturation state. At the current rate of warming and ocean acidification, reefs are expected to experience significant declines in coral abundance, coral diversity, and reef growth before the end of this century [1]. Despite this, some coral species appear to be more tolerant of these predicted conditions than others [2–4]. The relationship between coral physiology and its microbiome may shed light on why some corals are more resilient to global change conditions.
Elevated seawater temperature can lead to coral bleaching: a process whereby scleractinian corals lose substantial numbers of their photosynthetic endosymbiotic dinoflagellates (Symbiodinium spp.), giving the colony a pale (hence bleached) appearance. Bleaching damages coral health, slows or arrests growth, and can lead to mortality [1,5–7]. Factors associated with the ability to tolerate and recover from bleaching include coral energy reserves (i.e., lipid, protein, carbohydrates) [8–11], heterotrophic feeding capacity and/or plasticity [12–15], and endosymbiont type shuffling [9,16]. Ocean acidification can also affect some corals. Decreases in seawater pH hamper coral calcification in some species [4,17–19], but not in others [3,4,20,21]. Additionally, ocean acidification can dramatically impact coral health in some cases [2,22], while in other cases it has little or no negative effects on coral host physiology [4,19,23] and is often coupled with a stimulating effect on the endosymbiotic algae [23–25]. However, corals are increasingly exposed to the chronic dual stress of both rising temperature and ocean acidification simultaneously, and our understanding of how corals respond to the single stress of elevated temperature or acidity do not inform us on how coral might respond to the dual stress. At the current rate of atmospheric CO2 increase, the tropical oceans are conservatively expected to warm by 1–2°C and seawater pH to decrease by ~0.3 pH units over the course of this century [26].
Coral physiological responses to elevated temperature stress tend to be more severe when simultaneously exposed to reduced pH conditions in some species [2,4,23], but not in other species [3,4,23,24]. Physiological traits associated with coral resilience to both temperature and pH stress include high energy reserves, thermally tolerant endosymbiont types, and heterotrophic feeding on zooplankton or organic matter [4,23,27]. Thus, some species are more resilient to stress conditions expected by the end of this century than others. However, we do not fully understand what drives that resilience. Documented shifts, and lack of shifts, in the microbial community composition of heat-stressed corals [28–32] suggest that the microbiome may be a critical component of coral resilience. More importantly, the relationship between coral physiology and its microbiome may shed light on why some corals are more affected by the dual stress of elevated temperature and ocean acidification than others.
Coral-associated microbial communities are highly diverse, vary seasonally, and–with some exceptions–are generally consistent within a coral species [33–38]. Elevated temperature stress has been associated with significant shifts in coral microbial community composition in some cases [28–30] that appear to return to pre-bleaching states following recovery [28]. Specific Operational Taxonomic Units (OTUs) associated with bacteria of the genus Vibrio, and other OTUs from the bacterial classes γ-Proteobacteria, δ-Proteobacteria, and Acidobacteria, coincide with bleaching in corals, suggesting that such taxa cause or facilitate coral bleaching, or that they opportunistically increase in abundance in bleached corals when the coral’s health is compromised [28,29,33,39–44]. Bourne et al. [28] hypothesized that the loss of endosymbiotic algae during coral bleaching reduces the amount of reactive oxygen species–natural bacterial inhibitors–thus allowing opportunistic bacteria to infect and/or proliferate. Conversely, Banin et al. [33] proposed that bacterial pathology increases with temperature and causes bleaching, which is consistent with studies showing that microbial pathogenesis is temperature-dependent [39]. However, more recent work indicates that the microbiome may play a role in coral resilience to heat stress. Reschef et al. [45] proposed that beneficial microbes increase in stressed corals, conferring an immune-like response coined the “Coral Probiotic Hypothesis” making corals tolerant of stressful conditions like bleaching. Santos et al. [43] suggested that increases in nitrogen-fixing bacteria in bleached corals provide an alternative mechanism for corals to acquire fixed nitrogen in the absence of abundant endosymbionts, thus helping corals tolerate climate change stresses–though Pogoreutz et al. [46] found that increases in Diazotrophs and N-fixation are a stress response that exacerbates coral-algal symbiotic breakdown and bleaching. Others have shown that thermally tolerant corals can also have stable microbial communities under temperature stress or benefit from stable microbial communities that resemble their non-bleached counterparts [31,32]. Therefore, the microbiome has the potential to play a role in coral susceptibility, resistance, and recovery from stress events that could make the difference between coral species resilience and persistence, or extinction.
Ocean acidification has also been shown to cause shifts in coral microbial community composition, though no patterns emerge from the existing literature. Decreases in seawater pH result in increases, decreases, and no change in microbial diversity and relative abundances of the dominant classes of microbes [30,47–50]. Microbially mediated nutrient cycling can also be affected by ocean acidification by causing decreases in nitrogen fixation rates [51]. Clearly, more research is needed to address the effects of ocean acidification on coral microbial communities, particularly in combination with ocean warming.
Today, both rising temperatures and ocean acidification are occurring simultaneously. Only one study to date has examined the combined effects of elevated temperature and ocean acidification on the coral microbiome [30], and none have investigated the possible connection between the coral physiology and the microbiome under these dual stress conditions. Webster et al. [30] found that the microbial community composition shifts in response to end-of-century conditions were greater for the coral Acropora millepora than for the coral Seriatopora hystrix, and that dominant changes in bacterial phyla differed between the species. While Vibrio seems to play a large role in coral responses to temperature stress [28,44], this temperature sensitivity is lost when combined with ocean acidification stress [30]. At the same time, how changes in the coral microbiome are related to changes in coral physiology (animal host and endosymbiotic algae) under both temperature and pH conditions expected by the end of this century are completely unknown. Understanding this relationship may be key to uncovering why some corals are more resilient than others to climate change (see reviews by [52–54]). Here, we assessed the effects of the dual stresses of increased temperature and ocean acidification (i.e., increased pCO2) expected later this century on coral microbial community composition. We interpreted the microbial findings within the context of previously published physiological measurements from the exact same corals in this study (i.e., animal host and endosymbiotic algae). This holobiont approach to understanding corals [37] can only increase our understanding of why some corals are more resilient than others. We hypothesize that corals with a stable microbial community composition are physiologically more resilient to combined ocean warming and acidification.
Materials and methods
Experimental design
This experiment was conducted at Reef Systems Coral Farm (New Albany, OH, USA) in summer 2011 in coarsely filtered (150 μm filters) artificial seawater and is described in detail in Schoepf et al. [4]. Additional filtering of the seawater was deemed unnecessary as corals maintain microbial community compositions that are compositionally distinct from their surrounding seawater [32,36,37,55]. Due to financial limitations, this study focused only on a subset of these corals, which were analyzed for microbial community composition. A brief description of the experimental methods that pertain only to this subset of corals is as follows.
Six colonies of the Pacific corals Acropora millepora and Turbinaria reniformis were collected from northwest Fiji and transported to Reef Systems Coral Farm (New Albany, Ohio, USA) which is a CITES permit holder. The coral colonies were maintained in a single large recirculating tank for 2.5 months prior to the experiment, and were exposed to the same recirculating seawater and environmental conditions. We assumed that the opportunity for acquiring any given bacteria was equal among all corals in the same way that it would have been had the corals been freshly collected from the reef prior to the experiment. Since corals maintain microbial community compositions that are compositionally distinct from the surrounding seawater [32,36,37,55], any differences in the microbial community composition of the corals after 2.5 months in the large tank was assumed to be due to species-specific differences in how coral establish and maintain their microbiome.
Prior to starting the manipulative experiment, 6 control tanks and their shared recirculating sump, and all 6 treatment tanks and their shared recirculating sump were filled with artificial seawater that was made in a common bath and equally partitioned among all tanks and sumps. Thus all seawater starting conditions in the experimental tanks, including the seawater microbial community composition, were the same for all corals in this study and the only differences between the treatment and control tanks during the experiment were the temperature and pCO2 levels.
Each coral colony was divided into 12 fragments. A. millepora is a branching coral that contains the thermally sensitive endosymbiont type Symbiodinium C21a, while T. reniformis is a foliose coral that contains the thermally tolerant Symbiodinium trenchi (Symbiodinium type reported in the companion paper by Hoadley et al., [23]). One fragment from each colony was assigned to each of the six control tanks (26.5°C and pCO2 of 364 μatm) and to each of the treatment tanks (29.0°C and pCO2 of 750 μatm), yielding a total sample size of 6 treatment and 6 control fragments for each species (total n = 24). This sample size is the same or larger than the sample sizes used in the vast majority of coral microbial studies (e.g., [28,30,32,44,55–57]).
Seawater pCO2 was controlled by bubbling in pure CO2, CO2-free air, or ambient air in each sump to achieve the desired pCO2. Temperature was also controlled in each sump with submerged computer-controlled titanium heaters. At the beginning of the experiment, temperature was raised gradually from 26.5°C to 31.5°C over the first 18 days to prevent heat shock, then maintained at 31.5°C for 6 additional days, for a total of 24 days with an average temperature of 29°C. pCO2 was raised gradually over the first 4 days to prevent pCO2 shock, and maintained at 750 μatm for the remaining 18 days. Corals were grown in the tanks for 24 days on a 9:15 hour light:dark cycle (275 μmol quanta m-2 s-1) and fed every three days with two-day old brine shrimp nauplii that were hatched in a separate single batch culture with new seawater. Coral were fed the brine shrimp in separate feeding containers, and returned to their experimental tanks after one hour. The feeding container water and remaining brine shrimp were discarded so as not to introduce brine shrimp into the recirculating system (see Schoepf et al., [4] for more details). The control temperature of 26.5°C represented the average summer temperature in Fiji where the coral colonies were originally sourced, while the average elevated temperature of 29.0°C represented the upper limit of current Fiji summer temperatures, but is still below the bleaching threshold at that location (www.ospo.noaa.gov/Products/ocean/index.html), and representative of the increase in baseline temperatures expected by the end of this century in tropical regions under the RCP 8.5 scenario [26]. The control and treatment pCO2 levels represented present day conditions and those expected by mid-century under the RCP 8.5 scenario and by 2100 under the RCP 6.0 scenario [26], respectively. Throughout the study, temperature, salinity, pHNBS, and total alkalinity (TA) were measured daily according to methods described in Schoepf et al. [4]. Daily pCO2, aragonite saturation state (Ωarag), and pH (reported on the pH total scale, pHT) were then calculated according to Schoepf et al. [4]. The experiment ran for 24 days from 19 July– 12 August, 2011, then coral fragments were frozen at -80°C, and transported to the lab for analyses.
16S rRNA gene sequencing and OTU table construction
Whole coral tissue and associated mucus layer was airbrushed with milliQ water to produce a slurry from the 24 frozen coral fragments and frozen at -80°C. Coral slurries were centrifuged and total genomic DNA was extracted using PowerSoil® DNA Isolation kits according to the manufacturer instructions (MoBio Laboratories, Inc., Carlsbad, CA, USA). DNA was eluted in the provided buffer and quantified on a Qubit 2.0 instrument (Invitrogen, Carlsbad, CA, USA). DNA samples were then sequenced at the Argonne National Laboratory using the Illumina MiSeq platform. The V4 region of the 16S rRNA gene was targeted with the universal primers 515F and 806R [58,59] that targeted both bacteria and archaea. The use of the 16S rRNA gene is a standard in the field for identifying microbial community composition and structure in the coral literature (e.g., [28,31,32,44,49,55,57]. Sequencing data was processed using the QIIME platform [60]. Briefly, FASTQ forward and reverse files were joined and libraries were split based on the barcodes. Sequence length control and end-trimming were performed with default parameters, and the minimum quality score was set as 19. Additional parameters included a minimum count of 10 for an OTU to be retained and presence in at least 25% of the samples. OTUs were assigned based on the release 111 of the Silva ribosomal database [61] and clustered at 97% similarity level. Chimeras were identified and removed with usearch61 [62]. After the OTU table was generated, any OTUs matching mitochondria, chloroplasts, or eukaryotes were removed from the biom table using filter_taxa_from_otu_table.py and later summarize_taxa.py. Details on PCR, sequence processing, and command line work are available in the S1 Methods of the Supporting Information.
Physiology measurements
Physiological measurements in this study were already reported in Schoepf et al. [4], Levas et al. [27], and Hoadley et al. [23] along with details of each analysis. In brief, calcification [from [4]] was measured by the buoyant weight method [63] during the study and standardized to surface area to produce calcification rates for the first and second half of the experiment. Only the latter calcification rate is used in this study as it is most relevant to the other physiological and microbial data, which were all measured either during the last few days of the experiment or at the end of the 24-day experiment. Particulate organic carbon (POC) flux [from [27]], ratio of photosynthesis to respiration (P:R) [from [23]], and photosystem II maximal efficiency (Fv/Fm) [from [23]] were measured on living corals during the last four days of the experiment. POC flux was calculated as the difference in blank-corrected POC concentration of the seawater in a sealed chamber containing a coral fragment and the initial seawater POC concentration, standardized to the incubation duration (1.5 hours) and fragment surface area. POC was defined as the concentration of organic particles captured on a GF/F (0.7 μm nominal pore size) and measured by combustion using a Costech Elemental Analyzer. When POC fluxes are positive, corals are releasing organic matter into the water, typically in the form of mucus. When POC fluxes are negative, coral are taking up POC as a source of fixed carbon [27]. Maximum net photosynthetic rate (P) and light acclimated dark respiration (R) were measured from the change in oxygen concentration of each coral fragment in a respirometry chamber and standardized to surface area. The ratio of P:R was calculated from the ratio of gross P (net P + R) to R. Dark acclimated quantum yield of photosystem II (Fv/Fm), was measured in the light by pulse amplitude modulation fluorometry (Diving PAM, Waltz, Germany). Fv/Fm is generally viewed as a proxy for coral heat sensitivity [64].
In the laboratory, the following physiological analyses were performed on all frozen coral fragments collected on the last day of the experiment and reported by Schoepf et al. [4]: total lipids, animal host soluble protein, animal host soluble carbohydrates, total biomass, endosymbiotic algal density, chlorophyll a, and total surface area. The following physiological variables were performed on the same frozen fragments and reported by Hoadley et al. [23]: soluble algal protein, and soluble algal carbohydrates.
In brief, total lipids, animal host soluble protein and animal host soluble carbohydrates were quantified from ground whole coral subsamples and standardized to ash-free dry weight. Lipids were extracted in a 2:1 chloroform:methanol solution, washed in 0.88% KCl, extracted and washed again in 100% chloroform and 0.88% KCl, respectively. For the animal host soluble protein and carbohydrate analyses, the animal fraction was separated from the endosymbiotic algae via sonication and centrifugation. Protein was extracted using the bicinchoninic method [65] with bovine serum albumin as a standard (Pierce BCA Protein Assay Kit). Carbohydrates were quantified using the phenol-sulfuric acid method with glucose as a standard [66]. Biomass was determined as the difference between the dry and burned weight of a ground coral subsample and standardized to surface area. Coral surface area was determined using the single wax dipping method [67,68] for the branching A. millepora and the aluminium foil method [69] for the plating T. reniformis.
Endosymbiotic algal density, chlorophyll a, soluble protein, and soluble carbohydrates were determined on airbrushed coral slurry where the algae were then separated from the coral host via centrifugation, and standardized to surface area. The number of algal cells were counted on six replicate subsamples using a hemocytometer and a Nikon microphot-FXA epifluorescent microscope. Chlorophyll a was extracted from another subsample in methanol and quantified spectrophotometrically according to Porra et al. [70]. Soluble algal protein and carbohydrates were determined using the same methods as above for the animal host.
Statistical analyses
The data were analyzed using non-parametric multivariate techniques to determine if the microbial community composition and structure varied among coral species and treatments. As rare OTUs can sometimes play an interesting role in microbial ecology [65], we included all sequences in the analyses. The OTU table was used as the input for microbial community analyses in PRIMER E v. 1.0.6 (Quest Research Limited, Auckland, New Zealand). The sequence data was evaluated at three levels: Phylum, Class, and OTU. These levels of analyses were done to facilitate comparisons with other publications. At the phylum level (Level 2 table from Qiime), the sum of the abundances of all OTUs within each phylum was square root-transformed prior to construction of a Bray-Curtis resemblance matrix. The same was performed at the Class (Level 4) level. Analysis of Similarity (ANOSIM) was used to test for the effect of species and treatment on the microbial community composition at the phylum and class levels. Similarity Percentage (SIMPER) analyses were conducted to determine the degree of dissimilarity in microbial communities between coral species, between control and treatment corals within a species, and to determine which phyla or classes were responsible for the largest portion of those dissimilarities. The same analyses were then conducted using all of the individual OTU sequence abundances. In addition, the Shannon Diversity Index at the OTU level was computed for each species and treatment. A univariate two-way analysis of variance (ANOVA) was used to test the effects of species and treatment on the Shannon Diversity Index, where the data was first tested for normality using the Shapiro-Wilk’s test and homogeneity of variance was assessed with plots of expected vs. residual values. A posteriori Slice tests (i.e., tests of simple effects, Winer [71]) were used to determine if treatment corals differed from controls within each species.
The physiological measurements were analyzed using non-parametric techniques to determine if corals differed physiologically between species and treatments. Since these data are a subset of those from Schoepf et al. [4], Hoadley et al. [23], and Levas et al. [27], and presented together for the first time, new statistical analyses were necessary using only the data from the samples in this study. Univariate Kruskal-Wallis tests were conducted on each individual variable to determine if they differed between control and treatment corals of each species. A Euclidean distance-based resemblance matrix was then constructed using normalized data. ANOSIM was used to test for the effect of species and treatment on the overall coral physiology. SIMPER analyses were conducted to determine which physiological variable(s) were responsible for the largest portion of the differences detected in the ANOSIM. These multi-variate analyses of the physiological data are unique to the current study.
Non-parametric multidimensional scaling (NMDS) analyses were performed to graphically represent relationships between the microbial abundance variability and the coral physiology variability between species and treatments in multidimensional space. The two sets of data (microbial and coral physiology) were then compared to determine if microbial community composition and structure co-varied with coral physiology using two strategies: 1) vectors were added to NMDS plots (Pearson correlations > 0.1) to show the direction and magnitude of the influence of each physiological variable to the distribution of the microbial-based data points in 2D NMDS space, and 2) Spearman correlations were performed to compare the physiology-based NMDS distribution pattern to the microbial-based NMDS distribution pattern (BEST test).
The Kruskal-Wallis tests were performed using SAS software version 9.2. All other analyses were generated using the software package Primer V6 [72,73]. Where appropriate, p < 0.05 was considered significant.
Since the coral fragments in the control and treatment tanks were all from the same parent colonies (i.e., one fragment from each colony and species in each tank), changes in the microbiome were due to treatment effects and not due to genetic differences between colonies. Overall, the experimental design allowed us to detect changes in the coral microbial community composition due to treatment effects, independent of starting microbial community composition and inter-colony variability. Thus, any observed differences in the coral microbial community composition in the experiment were due to innate differences between species and treatment effects alone.
Results
All corals survived the experiment, though their physical appearance varied by species and treatment. Half of the treatment A. millepora were pale at the end of the study and all other fragments retained a healthy brown (for control A. millepora) or mustard yellow (for both control and treatment T. reniformis) color (Fig 1). The average seawater temperature, pHT, pCO2, total alkalinity, and Ωarag for the control and treatment tanks are given in Table 1.
Fig 1. Photographs of representative coral fragments at the end of the experiment.

A. millepora and T. reniformis after 24 days in the experiment under control (26.5°C and pCO2 of 364 μatm) (left side) and treatment (29.0°C and pCO2 of 750 μatm) (right side) conditions. The treatment A. millepora fragment typifies the paling that was observed in half of the fragments in this group. Photos by V Schoepf.
Table 1. Average conditions (± 1standard error) for the control and treatment tanks.
| Control | Treatment | |
|---|---|---|
| Temperature (°C) | 26.45 ± 0.01 | 28.93 ± 0.02 |
| pHT | 8.07 ± 0.01 | 7.81 ± 0.01 |
| pCO2 (μatm) | 364 ± 10 | 750 ± 26 |
| TA (μmol kg-1) | 2269 ± 11 | 2305 ± 9 |
| Ωarag | 3.69 ± 0.07 | 2.52 ± 0.06 |
pHT = pH reported on the total scale, TA = total alkalinity, Ωarag = aragonite saturation state.
Modified from Schoepf et al. [4].
Microbial community composition
Overall, there were 831 OTUs across all coral fragments spanning 48 Phyla: 565 for A. millepora and 650 for T. reniformis (S1 Table). In addition, 388 OTUs were shared by both coral species, 175 were unique to A. millepora, 263 were unique to T. reniformis. We observed that sequences affiliated with members of the Proteobacteria (primarily Alphaproteobacteria and Gammaproteobacteria classes) and Firmicutes Phyla were the most abundant in treatment and controls of both coral species, with sequences matching Actinobacteria, Bacteroidetes, and Acidobacteria Phyla being the next most abundant and typically present in both species and treatments (Fig 2A). However, ANOSIM revealed that at the Phylum level, bacterial communities on average did not significantly differ between species, or between treatment and controls of either species (ANOSIMs Global R -0.036, p = 0.66) (Fig 2B).
Fig 2. Microbial relative abundances by phylum.

(A) Acropora millepora (Am) and Turbinaria reniformis (Tr) under control (26.5°C and pCO2 of 364 μatm) and treatment (29.0°C and pCO2 of 750 μatm) conditions for each sample (1–6). (B) Average microbial relative abundances by Phylum.
Significant differences in the coral microbial communities were found at the Class and OTU levels between the species, and between treatment and controls of A. millepora, but not between treatment and control T. reniformis according to ANOSIM analyses (Table 2, S1 Fig). At the OTU level, the microbial community composition of treatment and control A. millepora and T. reniformis were 77% and 59% dissimilar, respectively (S2 Table). In A. millepora, this dramatic dissimilarity was due to increases in OTUs associated with Sphingomonas, Pseudomonas, and Halanaerobium, the virtual appearance of Rhodococcus fascians, and decreases in 16S rRNA gene sequence associated with the Rhodobacteraceae in treatment corals compared to the controls (Fig 3, S2 Table). The virtual disappearance of Pseudovibrio was due to a single coral fragment (Fig 3, S2 Table). At the same time, the Shannon Diversity Index decreased significantly from an average of 3.21 in control to 1.98 in treatment A. millepora (S3 Table). Though variability among treatment and control A. millepora fragments was high (Fig 3A), several overarching patterns emerged in this coral species. Treatment conditions resulted in relative abundance increases in sequences affiliated with Sphingomonas (observed in 5 of the 6 fragments), Pseudomonas (observed in 4 of the 6 fragments), Halenaerobium (observed in 5 of 6 fragments) and R. fascians (observed in all 6 fragments) coupled with decreases in a 16S rRNA gene sequence associated with the Rhodobacteraceae (observed in 4 of 6 fragments) (Fig 3A, S1 Table). On average, this resulted in -9 to +100-fold changes in these OTUs (Fig 3B). Microbial OTU diversity did not significantly differ between control and treatment T. reniformis (average Shannon Diversity Index of 3.4 vs 2.9, respectively) nor between species (S3 Table).
Table 2. One-way ANOSIMs of microbial community composition with pairwise tests of each coral species and treatment combination at the a) Class and b) OTU level.
| Pairwise tests of groups | R statistic | P-value |
|---|---|---|
| a) Class level | ||
| Am Control vs. Am Treatment | 0.296 | 0.028 |
| Am Control vs. Tr Control | 0.254 | 0.043 |
| Am Control vs. Tr Treatment | 0.433 | 0.002 |
| Am Treatment vs. Tr Control | 0.356 | 0.013 |
| Am Treatment vs. Tr Treatment | 0.230 | 0.002 |
| Tr Control vs. Tr Treatment | 0.039 | 0.284 |
| b) OTU level | ||
| Am Control vs. Am Treatment | 0.220 | 0.048 |
| Am Control vs. Tr Control | 0.263 | 0.037 |
| Am Control vs. Tr Treatment | 0.394 | 0.009 |
| Am Treatment vs. Tr Control | 0.426 | 0.011 |
| Am Treatment vs. Tr Treatment | 0.302 | 0.004 |
| Tr Control vs. Tr Treatment | 0.059 | 0.234 |
The overall model at the Class and OTU levels were significant (Global R = 0.256, p<0.01, 999 permutations and R = 0.257, p<0.02, 999 permutations, respectively). Bolded p-values are significant. Am = Acropora millepora, Tr = Turbinaria reniformis, Control = 26.5°C and pCO2 of 364 μatm, Treatment = 29.0°C and pCO2 of 750 μatm.
Fig 3. Relative abundance of the nine OTUs contributing the most to the dissimilarity between treatment and control corals as determined by SIMPER analysis.

(A) Results for Acropora millepora (Am) and Turbinaria reniformis (Tr) under control (26.5°C and pCO2 of 364 μatm) and treatment (29.0°C and pCO2 of 750 μatm) conditions for (A) each sample and (B) on average per treatment and species. Average relative abundance details in S2 Table. Dissimilarity analyses details by SIMPER are in S2 Table. ANOSIM results in Table 2 correspond to panel (B). Note that the proportionate contributions of the OTUs here do not sum to 1 because only the nine OTUs contributing the most to the dissimilarities are shown. Sphingomonas, Ruegeria, Pseudovibrio, Methylobacterium, and Rhodobacteraceae belong to the class Alphaproteobacteria. Pseudomonas, Acinetobacter, and Coxiella belong to the class Gammaproteobacteria. Rhodococcus fascians belongs to the phylum Actinobacteria, Halanaerobium and Staphylococcus to Firmicutes, and Percinobacter to Bacteroidetes.
Coral physiology
Calcification, Fv/Fm, and algal carbohydrate concentration significantly declined by 136%, 23%, and 32%, respectively, while animal host protein concentration increased by 51% in treatment compared to control A. millepora (Fig 4A, S4 Table). In T. reniformis, the endosymbiotic algal protein was four-fold higher and algal carbohydrate concentrations were 34% lower in the treatment compared to control corals (Fig 4B, S4 Table). No other significant differences were detected between treatment and control corals of either species (S4 Table). It is worth noting that host protein concentrations as well as the total energy reserves (i.e., lipid, protein, and carbohydrates) in treatment T. reniformis were 100% and 13% greater than in treatment A. millepora, respectively (Fig 4). When all of the physiology data were evaluated as a whole, significant overall differences between coral species, and between treatments within species, were found (Table 3). Animal host protein and endosymbiotic algal carbohydrates contributed the most, and algal cell density and P:R the least, to the overall physiological differences between the two coral species (S5 Table).
Fig 4. Average (± 1SE) physiological variables.
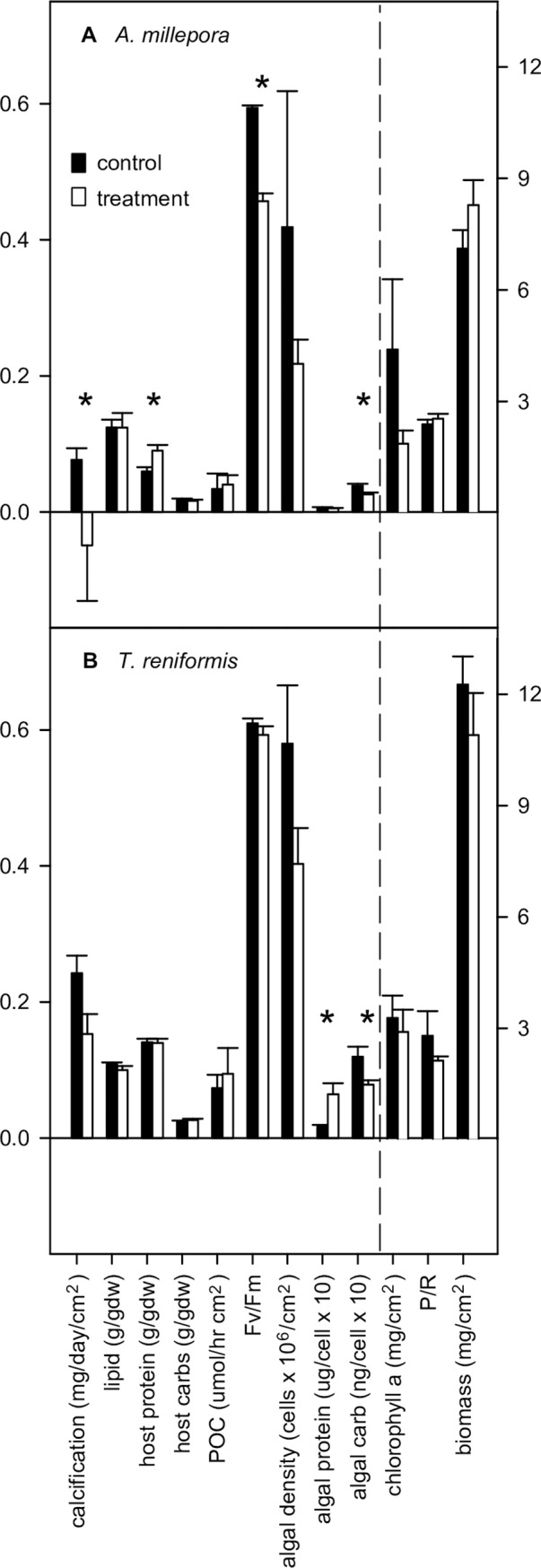
Results given for (A) Acropora millepora and (B) Turbinaria reniformis. Measurement units for each variable in brackets along the x-axis. Left-hand y-axis scale corresponds to the first 9 variables and the right-hand y-axis scale corresponds to the last three variables (separated by a dashed line). Black bars = control coral (26.5°C and pCO2 of 364 μatm), white bars = treatment coral (29.0°C and pCO2 of 750 μatm), carbs = carbohydrates, POC = particulate organic carbon flux, * = significant difference between control and treatment averages for a given variable by Kruskal-Wallis test (details in S3 Table). Data from Schoepf et al. [4], Hoadley et al. [23], and Levas et al. [27].
Table 3. One-way ANOSIM of coral physiology with pairwise tests of each species and treatment combination.
| Pairwise tests of groups | R statistic | P-value |
|---|---|---|
| Am Control vs. Am Treatment | 0.265 | 0.015 |
| Am Control vs. Tr Control | 0.515 | 0.002 |
| Am Control vs. Tr Treatment | 0.537 | 0.002 |
| Am Treatment vs. Tr Control | 0.807 | 0.002 |
| Am Treatment vs. Tr Treatment | 0.754 | 0.002 |
| Tr Control vs. Tr Treatment | 0.178 | 0.024 |
The overall model was significant (Global R = 0.509, p<0.001, with 999 permutations). Bolded p-values are significant. Am = Acropora millepora, Tr = Turbinaria reniformis, Control = 26.5°C and pCO2 of 364 μatm, Treatment = 29.0°C and pCO2 of 750 μatm.
Coral microbiome and physiology
NMDS analyses of the microbial OTU relative abundance data revealed clear separation of the coral microbial communities between the coral species, and between treatment and controls of A. millepora, but not between treatment and control T. reniformis (Fig 5, Table 2). Of the physiological variables, host protein and Fv/Fm best described the overall microbiology NMDS pattern of the corals (BEST, spearman correlation R = 0.344, p = 0.26), though the results were not statistically significant. Nevertheless, the host protein vector appears to define an axis that best describes differences in the microbial communities between the species whereas the Fv/Fm vector appear to define an axis separating the microbial communities of treatment and control corals (Fig 5). BEST analysis was also conducted on A. millepora alone as it expressed significantly different microbial communities between treatment and control fragments. However, the model was not significant and no physiological variables described the overall microbiology NMDS pattern within A. millepora (Global R = 0.21, p = 0.95).
Fig 5. NMDS ordination using microbial OTU community composition from all 24 coral fragments.

Gray vector overlay shows the proportional influence of each physiology variable to the NMDS plot distribution. Am = Acropora millepora (circles), Tr = Turbinaria reniformis (open squares), Control (blue) = 26.5°C and pCO2 of 364 μatm, Treatment (black) = 29.0°C and pCO2 of 750 μatm. Calc = calcification rate during the last two weeks of the study, a_cells = endosymbiotic algal cell density, Chla = chlorophyll a concentration, carbs = carbohydrate concentration, lipid = total lipid concentration, protein = soluble protein concentration, POC = particulate organic carbon flux, h = animal host, a = endosymbiotic algae.
Discussion
When corals are exposed to stress such as increased temperature or decreases in seawater pH, the coral microbial community often undergoes compositional changes [28,29,40,44,48,49,55,74,75]. The current study is the first to simultaneously examine the coral microbial community composition, coral host physiology, and endosymbiotic algal physiology associated with the dual stress of increased seawater temperature and lower pH. When comparing the two coral species here, we find that T. reniformis with its thermally tolerant endosymbiont Symbiodinium trenchi has a stable microbial community composition and is only slightly affected physiologically when exposed to conditions expected by the end of this century for 24 days, whereas A. millepora with its more thermally sensitive endosymbiont type C21a suffers both a decline in microbial diversity and a shift in its microbial community composition combined with a more dramatic physiological decline under the same stressful conditions.
The coral microbiomes of A. millepora and T. reniformis were dominated by members of the phylum Proteobacteria (Fig 2), which is a pattern found in most coral microbial communities (e.g., [30,48,55,76]). However, when exposed to the dual stress of increased temperature and lower pH, we found no significant change in the phylum-level microbial community composition of A. millepora (Fig 2B) whereas Webster et al. [30] did. This could be due to the different experimental conditions of both studies. Namely, Webster et al. [30] corals were collected two weeks before the experiment, maintained at 28°C or 31°C during the experiment, and were kept in 1μm filtered seawater and starved of any heterotrophic food source. In the current study, the corals were collected three months prior to the experiment, maintained at 26.5°C or 29°C in 150 μm filtered seawater, and fed Artemia nauplii twice a week.
At the OTU level, the microbial community composition changed and diversity decreased for A. millepora, but not T. reniformis, when exposed to the dual stress treatment (Table 2, S3 Table). In the field of ecology, the insurance hypothesis states that microbial diversity stabilizes microbial community function [77]. Therefore, the loss of diversity observed in dually stressed A. millepora may indicate a destabilized microbial community function. However, increases in coral microbial diversity in response to sewage and sedimentation stress have also been interpreted to mean microbial community destabilization [78]. Here, we find that the large shifts in OTUs most responsible for the differences between treatment and control A. millepora corals do suggest an association between the microbiome and decreased coral health, which suggests destabilized microbial function (Fig 3). Specifically, the large increases in OTU relative abundance of Sphingomonas and Pseudomonas (Fig 3B) may be an indication of declining coral health as Sphingomonas-like bacteria and P. aeruginosa are associated with coral disease [44,79]. The large declines in Rhodobacteraceae could indicate a decrease in nitrogen-fixing ability [43,51,80]. Lastly, the known plant pathogen Rhodococcus fascians dramatically increased in all six A. millepora fragments (Fig 3A). Though we have no direct evidence that any of the bacterial shifts in A. millepora led to specific diseases or health decline, none of these OTU shifts suggest an acclimation response that could be conferring a probiotic [45] or protective effect in response to the dual stress of elevated temperature and ocean acidification. Alternatively, these large OTU shifts may indicate a restructuring of the microbial community in order to facilitate adaptation to the dual-stress conditions [31]. The maintenance of diversity and the microbial community stability observed in T. reniformis might have been because this species was already pre-adapted to the dual stress conditions, as has been demonstrated for some populations of Acropora hyacinthus from American Samoa [31]. However, it is unlikely that the microbial community of T. reniformis was already pre-adapted to the dual stress of elevated temperature and pCO2 while A. millepora was not, since both sets of coral colonies were collected from the same site with the same temperature and pH history. Additional studies in other coral species from other regions are needed to further evaluate these findings.
More importantly, the combined microbial and physiological changes in A. millepora indicate that this species was broadly compromised under treatment conditions. Major components of the holobiont–the animal host, endosymbiotic alga, and the microbiome–all deteriorated under treatment conditions. In addition to the decline in microbial diversity and shifts in microbial community composition (Fig 3), decreases in calcification, Fv/Fm, and algal carbohydrate energy reserves in A. millepora indicate that both the host and algal physiological functions were compromised under treatment conditions (Fig 4A). In contrast, the T. reniformis holobiont was less affected with no negative impacts on the coral host physiology, a decline only in the algal carbohydrates, and no significant changes in the microbiome community or diversity (Fig 4B, Table 2, S3 Table). Though overall physiology was significantly different between treatment and controls for both species, the degree of separation between the two groups was greater for A. millepora than for T. reniformis (Table 3). In addition, treatment T. reniformis had double the host protein concentrations and slightly greater total energy reserves (i.e., lipid, protein, and carbohydrates) compared to treatment A. millepora (Fig 4). High energy reserves have been shown to be a key component to coral resilience in the face of temperature stress [8,9]. Interestingly, host animal protein was one of two variables that explained the highest amount of variation in microbial community composition (Fig 5) and overall physiological differences between the two species (S5 Table). In addition, T. reniformis releases almost twice as much organic matter (probably as mucus) as A. millepora under control and treatment conditions (Fig 4) [27], which could provide a stable medium for cultivating its microbiome since bacterial growth is dramatically enhanced on coral mucus [81,82]. This could be advantageous as microbial community composition stability has been linked to coral health in some cases [31,32,81]. It is also possible that T. reniformis has greater heterotrophic capacity and/or plasticity under dual stress conditions than does A. millepora. Previous work has shown that corals that are either heterotrophically plastic or have high heterotrophic capacity recover more quickly from temperature stress [12,14]. Heterotrophic plasticity or high capacity under the dual stress of elevated temperature and pCO2 could potentially underlie some of the holobiont resilience observed here in T. reniformis, though further research is needed to test this hypothesis. Finally, T. reniformis hosted a thermally tolerant endosymbiont type Symbiodinium trenchi and maintained Fv/Fm under dual stress, whereas A. millepora hosted the more sensitive endosymbiont C21a and Fv/Fm rates dropped (Fig 4). While endosymbiont type does play a role in thermal sensitivity, T. reniformis also has a much thicker tissue layer than A. millepora, which would provide greater photoprotection to its S. trenchi endosymbionts when under stress [23]. At this time, it is not clear what the link between endosymbiont type and microbial community composition shifts (or stability) under a dual temperature and acidity stress is. What does emerge from this study is that the physiological traits of T. reniformis, combined with a thermally tolerant Symbiodinium type and stable microbiome together appear to make it more tolerant to warmer and more acidic seawater conditions than A. millepora.
Summary
Overall, we show that the bacterial microbiome changed in the physiologically sensitive coral A. millepora but not in the physiologically more tolerant coral T. reniformis in response to conditions of elevated temperature and OA expected later this century. These findings are consistent with previous physiological work on the coral host and endosymbiotic algae of these species demonstrating the more resilient nature of the T. reniformis holobiont compared to the A. millepora holobiont [4,23,27]. These findings support the hypothesis that coral with a stable and diverse microbiome are also physiologically more resilient to the dual stress of elevated temperature and OA. Furthermore, our findings suggest that the animal host may play a role in determining the microbial community composition and diversity, and that multiple traits across the holobiont (i.e., host energy reserves, mucus production, Symbiodinium type, microbiome stability) appear to be involved in coral sensitivity or resilience to changes in seawater conditions expected on reefs later this century. We hypothesize that coral holobiont deterioration in the face of climate change may be in part triggered by a combination of the animal host’s protein and mucus influence on the microbiome and the Symbiodinium type. Further research is needed to determine if shifts in the coral holobiont physiology and/or Symbiodinium type cause shifts in the microbiome or vice versa, or if the two are independent of each other. Since some coral diseases and bleaching are known to be caused by bacteria [39,44,83,84], it is unlikely that the microbiome community changes in response to environmental stress are independent of the coral physiology. We further suggest that large organic matter release rates (typically in the form of mucus) may be important for supporting stable and presumably healthier microbial symbionts for corals. Our results are consistent with recent findings that some temperature-stress tolerant corals have a stable microbiome or benefit from stable microbial communities that resemble their non-bleached counterparts [31,32], and demonstrate for the first time that this also appears to be the case under the dual stresses of ocean warming and acidification. Additional studies with different coral species are needed to fully test this hypothesis. Model projections of coral persistence over the next century might need to consider not just coral host and endosymbiotic algae physiological responses to stress, but the combined responses of the coral host, endosymbiotic algae, and microbiome.
Supporting information
Additional details.
(DOCX)
(A) Acropora millepora (Am) and Turbinaria reniformis (Tr) under control (26.5°C and pCO2 of 364 μatm) and treatment (29.0°C and pCO2 of 750 μatm) conditions for each sample (1–6). (B) Average microbial relative abundances by Class.
(TIFF)
Results for Acropora millepora and Turbinaria reniformis under control (26.5°C and pCO2 of 364μatm) and treatment (29.0°C and pCO2 of 750μatm) conditions. Samples are from colonies numbered 1–6.
(XLSX)
Average percent dissimilarity between species and between treatments within species in microbial OTU abundance. Control = 26.5°C and pCO2 of 364μatm, Treatment = 29.0°C and pCO2 of 750μatm. Am = Acropora millepora, Tr = Turbinaria reniformis, Av.Abund = average abundance, Av.Diss = average dissimilarity, Diss/SD = Dissimilarity /standard deviation, Contrib% = contribution percent, Cum.% = Cumulative percent contribution.
(DOCX)
Effects of species and treatment were fixed and fully crossed. A posteriori slice tests were used to test for differences between treatment and controls within species. df = degrees of freedom, SS = type III sum of squares of the main effects, F = F-statistic. Significant effects are bolded.
(DOCX)
Significant differences (p ≤ 0.05) are bolded. calc = calcification, chla = chlorophyll a, cells = endosymbiotic algal cell density, lipid = total lipids, protein = soluble animal protein concentration, carbs = carbohydrate concentration, biomass = coral ash free dry weight per area, h = host, a = endosymbiotic algae.
(DOCX)
Summary of average squared distance (Av.Sq.Dist = 29.54) in coral physiology between Acropora millepora and Turbinaria reniformis across all control and treatment coral fragments. SD = standard deviation, Av.Value = average, Contrib % = percent contribution, Cum. % = cumulative percent.
(DOCX)
Acknowledgments
We thank the staff at Reef Systems Coral Farm, E. Zebrowski, M. Berzelis, M. Ringwald, S. Blackhurst, K. Dobson, C. Saup, and Y. Matsui for their assistance with the experiment. This work was supported by funding from the U.S. National Science Foundation (NSF-EF-1041124, 1040940, 1041070 to AGG, MEW, and WJC, respectively) and from The Ohio State University (start-up funds to MJW). Sequences have been deposited in NCBI’s Sequence Read Archive under accession number PRJNA344759. The physiological data are archived at www.bco-dmo.org/project/528004.
Data Availability
Sequences have been deposited in NCBI’s Sequence Read Archive under accession number PRJNA344759. The physiological data are archived at www.bco-dmo.org/project/528004.
Funding Statement
Funding by the U.S. National Science Foundation (NSF-EF-1041124, 1040940, 1041070 to AGG, MEW, and WJC, respectively) and from The Ohio State University (start-up funds to MJW). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. Reef Systems Coral Farm funder provided space for the experiment and contributed to the physical construction and maintenance of the experimental setup, but did not have any additional role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript. The specific roles of these authors are articulated in the ‘author contributions’ section.
References
- 1.Veron JEN, Hoeg-Guldberg O, Lenton TM, Lough JM, Obura DO, Pearce-Kelly P, et al. (2009) The coral reef crisis: The critical importance of <350 ppm CO2. Marine Pollution Bulletin 58: 1428–1436. doi: 10.1016/j.marpolbul.2009.09.009 [DOI] [PubMed] [Google Scholar]
- 2.Anthony KRN, Kline DI, Diaz-Pulido G, Dove S, Hoegh-Guldberg O (2008) Ocean acidification causes bleaching and productivity loss in coral reef builders. Proceedings of the National Academy of Sciences of the United States of America 105: 17442–17446. doi: 10.1073/pnas.0804478105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Edmunds PJ, Brown D, Moriarty V (2012) Interactive effects of ocean acidification and temperature on two scleractinian corals from Moorea, French Polynesia. Global Change Biology 18: 2173–2183. [Google Scholar]
- 4.Schoepf V, Grottoli A, Warner M, Cai W, Melman T, Hoadley K, et al. (2013) Coral energy reserves and calcification in a high-CO2 world at two temperatures. PLoS ONE 8: e75049 doi: 10.1371/journal.pone.0075049 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hoegh-Guldberg O (1999) Climate change, coral bleaching and the future of the world's coral reefs. Marine Freshwater Research 50: 839–866. [Google Scholar]
- 6.Brown BE (1997) Coral bleaching: causes and consequences. Coral Reefs 16 suppl: s129–s138. [Google Scholar]
- 7.Buddemeier RW, Fautin DG (1993) Coral bleaching as an adaptive mechanism. BioScience 43: 320–326. [Google Scholar]
- 8.Anthony KRN, Hoogenboom MO, Maynard JA, Grottoli AG, Middlebrook R (2009) Energetics approach to predicting mortality risk from environmental stress: a case study of coral bleaching. Functional Ecology 23: 539–550. [Google Scholar]
- 9.Grottoli A, Warner M, Levas S, Aschaffenburg M, Schoepf V, McGiinley M, et al. (2014) The cumulative impact of annual coral bleaching can turn some coral species winners into losers. Global Change Biology: doi: 10.1111/gcb.12658 [DOI] [PubMed] [Google Scholar]
- 10.Rodrigues LJ, Grottoli AG (2007) Energy reserves and metabolism as indicators of coral recovery from bleaching. Limnology and Oceanography 52: 1874–1882. [Google Scholar]
- 11.Schoepf V, Grottoli AG, Levas SJ, Aschaffenburg M, Baumann J, Matsui Y, et al. (2015) Annual coral bleaching and the long-term recovery capacity of coral. Proceedings of the Royal Society B 282: 20151887 doi: 10.1098/rspb.2015.1887 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Grottoli AG, Rodrigues LJ, Palardy JE (2006) Heterotrophic plasticity and resilience in bleached corals. Nature 440: 1186–1189. doi: 10.1038/nature04565 [DOI] [PubMed] [Google Scholar]
- 13.Houlbreque F, Ferrier-Pages C (2009) Heterotrophy in Tropical Scleractinian Corals. Biological Reviews 84: 1–17. doi: 10.1111/j.1469-185X.2008.00058.x [DOI] [PubMed] [Google Scholar]
- 14.Levas S, Grottoli AG, Hughes AD, Osburn C, Matsui Y (2013) Physiological and biogeochemical traits of bleaching and recovery in the mounding species of coral Porites lobata: implications for resilience in mounding corals. PLoS ONE 8: e63267 doi: 10.1371/journal.pone.0063267 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Tremblay P, Gori A, Maguer J, Hoogenboom M, Ferrier-Pages C (2016) Heterotrophy promotes the re-establishment of photosynthate translocation in a symbiotic coral after heat stress. Scientific Reports 6: 38112 doi: 10.1038/srep38112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Berkelmans R, van Oppen MJH (2006) The role of zooxanthellae in the thermal tolerance of corals: a 'nugget of hope' for coral reefs in an era of climate change. Proceedings of the Royal Society B-Biological Sciences 273: 2305–2312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Comeau S, Edmunds P, Spindel N, Carpenter RC (2013) The responses of eight coral reef calcifiers to increasing partial pressure of CO2 do not exhibit a tipping point. Limnology and Oceanography 58: 388–398. [Google Scholar]
- 18.Jokiel PL, Rodgers KS, Kuffner IB, Andersson AJ, Cox EF, Mackenzie FT (2008) Ocean acidification and calcifying reef organisms: a mesocosm investigation. Coral Reefs 27: 473–483. [Google Scholar]
- 19.Krief S, Hendy E, Fine M, ruth Y, Anders M, Foster G, et al. (2010) Physiological and isotopic responses of scleractinian corals to ocean acidification. Geochimica et Cosmochimica Acta 74: 4988–5001. [Google Scholar]
- 20.Reynaud S, Leclercq N, Romaine-Lioud S, Ferrier-Pagés C, Jaubert J, Gattuso J-P (2003) Interacting effects of CO2 partial pressure and temperature on photosynthesis and calcification in a scleractinian coral. Global Change Biology 9: 1660–1668. [Google Scholar]
- 21.Comeau S, Carpenter R, Edmunds P (2013) Effects of feeding and light intensity on the response of the coral Porites rus to ocean acidification. Marine Biology 160: 1127–1134. [Google Scholar]
- 22.Dove SG, Kline DI, Pantos O, Angly FE, Tyson GW, Hoegh-Guldberg O (2013) Future reef decalcification under a business-as-usual CO2 emission scenario. Proceedings of the National Academy of Sciences of the United States of America 110: 15342–15347. doi: 10.1073/pnas.1302701110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hoadley K, Pettay D, Grottoli A, Cai W, Melman T, Schoepf V, et al. (2015) Physiological response to elevated temperature and pCO2 varies across four Pacific coral species: Understanding the unique host+symbiont response. Scientific Reports 5: 18371 doi: 10.1038/srep18371 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Noonan SHC, Fabricius KE (2016) Ocean acidification affects productivity but not the severity of thermal bleaching in some tropical corals. ICES Journal of Marine Science 73: 715–726. [Google Scholar]
- 25.Langdon C, Atkinson MJ (2005) Effect of elevated pCO2 on photosynthesis and calcification on corals and interactions with seasonal change in temperature/irradiance and nutrient enrichment. Journal of Geophysical Research 110: doi: 10.1029/2004JC002576 [Google Scholar]
- 26.IPCC (2013) Summary for Policymakers. Cambridge, UK and New York, NY, USA: Cambridge University Press. [Google Scholar]
- 27.Levas S, Grottoli A, Warner M, Cai W, Bauer J, Schoepf V, et al. (2015) Organic carbon fluxes mediated by corals at elevated pCO2 and temperature. Marine Ecology Progress Series 519: 153–164. [Google Scholar]
- 28.Bourne D, Iida Y, Uthicke S, Smith-Keune C (2008) Changes in coral-associated microbial communities during a bleaching event. Isme Journal 2: 350–363. doi: 10.1038/ismej.2007.112 [DOI] [PubMed] [Google Scholar]
- 29.Littman RA, Bourne DG, Willis BL (2010) Responses of coral-associated bacterial communities to heat stress differ with Symbiodinium type on the same coral host. Molecular Ecology 19: 1978–1990. doi: 10.1111/j.1365-294X.2010.04620.x [DOI] [PubMed] [Google Scholar]
- 30.Webster NS, Negri AP, Botte ES, Laffy PW, Flores F, Noonan S, et al. (2016) Host-associated coral reef microbes respond to the cumulative pressures of ocean warming and ocean acidification. Scientific Reports 6: 19324 doi: 10.1038/srep19324 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ziegler M, Seneca F, Yum L, Palumbi S, Voolstra C (2017) Bacterial community dynamics are linked to patterns of coral heat tolerance Nature Communications 8: 14213 doi: 10.1038/ncomms14213 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Hadaidi G, Rothig T, Yum L, Ziegler M, Arif C, Roder C, et al. (2017) Stable mucus-associated bacterial communities in bleached and healthy corals of Porites lobata from the Arabian Seas. Scientific Reports 7: 45362 doi: 10.1038/srep45362 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Banin F, Ben-Haim Y, Israely T, Loya Y, Rosenberg E (2000) Effect of the environment on the bacterial bleaching of corals. Water Air and Soil Pollution 123: 337–352. [Google Scholar]
- 34.Chen CP, Tseng CH, Chen CA, Tang SL (2011) The dynamics of microbial partnerships in the coral Isopora palifera. Isme Journal 5: 728–740. doi: 10.1038/ismej.2010.151 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Koren O, Rosenberg E (2006) Bacteria associated with mucus and tissues of the coral Oculina patagonica in summer and winter. Applied and Environmental Microbiology 72: 5254–5259. doi: 10.1128/AEM.00554-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Rohwer F, Breitbart M, Jara J, Azam F, Knowlton N (2001) Diversity of bacteria associated with the Caribbean coral Montastraea franksi. Coral Reefs 20: 85–91. [Google Scholar]
- 37.Rohwer F, Seguritan V, Azam F, Knowlton N (2002) Diversity and distribution of coral-associated bacteria. Marine Ecology Progress Series 243: 1–10. [Google Scholar]
- 38.Sunagawa S, Woodley CM, Medina M (2010) Threatened Corals Provide Underexplored Microbial Habitats. Plos One 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ben-Haim Y, Thompson FL, Thompson CC, Cnockaert MC, HOste B, Swings J, et al. (2003) Vibrio coralliilyticus sp. nov., a temperature-dependent pathogen of the coral Pocillopora damicornis. International Journal of Systematic and Evolutionary Microbiology 309–315. doi: 10.1099/ijs.0.02402-0 [DOI] [PubMed] [Google Scholar]
- 40.Mouchka ME, Hewson I, Harvell CD (2010) Coral-associated bacterial assemblages: current knowledge and the potential for climate-driven impacts. Integrative and Comparative Biology 50: 662–674. doi: 10.1093/icb/icq061 [DOI] [PubMed] [Google Scholar]
- 41.Ritchie KB (2006) Regulation of microbial populations by coral surface mucus and mucus-associated bacteria. Marine Ecology Progress Series 322: 1–14. [Google Scholar]
- 42.Ritchie KB, Smith GW (1995) Preferential carbon utilization by surface bacterial communities from water mass, normal, and white-band diseased Acropora cervicornis. Molecular Marine Biology and Biotechnology 4: 345–352. [Google Scholar]
- 43.Santos HF, Carmo FL, Duarte G, Dini-Andreote F, Castro CB, Rosado AS, et al. (2014) Climate change affects key nitrogen-fixing bacterial populations on coral reefs. Isme Journal 8: 2272–2279. doi: 10.1038/ismej.2014.70 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Thurber RV, Willner-Hall D, Rodrigues-Mueller B, Desnues C, Edwards RA, Angly FE, et al. (2009) Metagenomic analysis of stressed coral holobionts. Environmental Microbiology 11: 2148–2163. doi: 10.1111/j.1462-2920.2009.01935.x [DOI] [PubMed] [Google Scholar]
- 45.Reshef L, Koren O, Loya Y, Zilber-Rosenberg I, Rosenberg E (2006) The coral probiotic hypothesis. Environmental Microbiology 8: 2068–2073. doi: 10.1111/j.1462-2920.2006.01148.x [DOI] [PubMed] [Google Scholar]
- 46.Pogoreutz C, Radecker N, Cardenas A, Gardes A, Voolstra C, Wild C (2017) Sugar enrichment provides evidence for a role of nitrogen fixation in coral bleaching. Global Change Biology 23: 3838–3848. doi: 10.1111/gcb.13695 [DOI] [PubMed] [Google Scholar]
- 47.Meron D, Buia MC, Fine M, Banin E (2013) Changes in Microbial Communities Associated with the Sea Anemone Anemonia viridis in a Natural pH Gradient. Microbial Ecology 65: 269–276. doi: 10.1007/s00248-012-0127-6 [DOI] [PubMed] [Google Scholar]
- 48.Morrow KM, Bourne DG, Humphrey C, Botte ES, Laffy P, Zaneveld J, et al. (2015) Natural volcanic CO2 seeps reveal future trajectories for host-microbial associations in corals and sponges. ISME Journal 9: 894–908. doi: 10.1038/ismej.2014.188 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Webster NS, Negri AP, Flores F, Humphrey C, Soo R, Botte ES, et al. (2013) Near-future ocean acidification causes differences in microbial associations within diverse coral reef taxa. Environmental Microbiology Reports 5: 243–251. doi: 10.1111/1758-2229.12006 [DOI] [PubMed] [Google Scholar]
- 50.Zhou G, Yuan T, CAi L, Zhang W, Tian R, Tong H, et al. (2016) Changes in microbial communities, photosynthesis and calcification of the coral Acropora gemmifera in response to ocean acidification. Scientific Reports 6: 35971 doi: 10.1038/srep35971 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Radecker N, Meyer F, Bednarz V, Cardini U, Wild C (2014) Ocean acidification rapidly reduces dinitrogen fixation associated with the hermatypic coral Seriatopora hystrix. Marine Ecology Progress Series 511: 297–302. [Google Scholar]
- 52.Ainsworth TD, Thurber RV, Gates RD (2010) The future of coral reefs: a microbial perspective. Trends in Ecology & Evolution 25: 233–240. [DOI] [PubMed] [Google Scholar]
- 53.Hernandez-Agreda A, GAtes R, Ainsworth T (in press) Defining theCoreMicrobiome in Corals’ Microbial Soup. Trends in Microbiology: 16. [DOI] [PubMed] [Google Scholar]
- 54.Sharp KH, Ritchie KB (2012) Multi-Partner Interactions in Corals in the Face of Climate Change. Biological Bulletin 223: 66–77. doi: 10.1086/BBLv223n1p66 [DOI] [PubMed] [Google Scholar]
- 55.Meron D, Atias E, Kruh LI, Elifantz H, Minz D, Fine M, et al. (2011) The impact of reduced pH on the microbial community of the coral Acropora eurystoma. ISME Journal 5: 51–60. doi: 10.1038/ismej.2010.102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Ceh J, Raina JB, Soo RM, van Keulen M, Bourne DG (2012) Coral-Bacterial Communities before and after a Coral Mass Spawning Event on Ningaloo Reef. Plos One 7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Littman RA, Willis BL, Pfeffer C, Bourne DG (2009) Diversities of coral-associated bacteria differ with location, but not species, for three acroporid corals on the Great Barrier Reef. Fems Microbiology Ecology 68: 152–163. doi: 10.1111/j.1574-6941.2009.00666.x [DOI] [PubMed] [Google Scholar]
- 58.Lane DJ (1991) 16S/23S rRNA sequencing In: Stackebrandt E, Goodfellow M, editors. Nucleic Acid Techniques in Bacterial Systematics. New York, USA: Wiley; pp. 115–148. [Google Scholar]
- 59.Caporaso J, Lauber C, Walters W, Berg-Lyons D, Huntley J, Fierer N, et al. (2012) Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. The International Society for Microbial Ecology Journal 6: 1621–1624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, et al. (2010) QIIME allows analysis of high-throughput community sequencing data. Nature Methods 7: 335–336. doi: 10.1038/nmeth.f.303 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T, Yarza P, et al. (2013) The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Research 41: D590–D596. doi: 10.1093/nar/gks1219 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Edgar RC, Haas BJ, Clemente JC, Quince C, Knight R (2011) UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27 2194–2200. doi: 10.1093/bioinformatics/btr381 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Jokiel PL, Maragos JE, Franzisket L (1978) Coral growth: buoyant weight technique In: Stoddart DR, Johannes RE, editors. Coral Reefs: Research Methods. Paris, France: UNESCO; pp. 529–541. [Google Scholar]
- 64.Warner ME, Lesser MP, Ralph PJ (2010) Chlorophyll fluorescence in reef building corals In: Suggett DJ, Prasil O., and Borowitzka M., editor. Chlorophyll Fluorescence in Aquatic Sciences: Methods and Applications: Springer, Berlin. [Google Scholar]
- 65.Smith PK, Krohn RI, Hermanson GT, Mallia AK, Gartner FH, Provenzano MD, et al. (1985) Measurement of Protein using Bicinchonic Acid. Analytical Biochemistry 150: 76–85. [DOI] [PubMed] [Google Scholar]
- 66.Dubois M, Giles KA, Hamilton JK, Rebers PA, Smith F (1956) Colorimetric method for determination of sugars and related substances. Analytical Chemistry 28: 350–356. [Google Scholar]
- 67.Stimson J, Kinzie RA III (1991) The temporal pattern and rate of release of zooxanthellae from the reef coral Pocillopora damicornis (Linnaeus) under nitrogen-enrichment and control conditions. Journal of Experimental Marine Biology and Ecology 153: 63–74. [Google Scholar]
- 68.Veal CJ, Carmi M, Fine M, Hoegh-Guldberg O (2010) Increasing the accuracy of surface area estimation using single wax dipping of coral fragments. Coral Reefs 29: 893–897. [Google Scholar]
- 69.Marsh JA (1970) Primary productivity of reef-building calcareous red algae. Ecology 51: 255–263. [Google Scholar]
- 70.Porra R, Tompson W, Kreidemann P (1989) Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochimica et Biophysica Acta 975: 384–394. [Google Scholar]
- 71.Winer B (1971) Statistical principles in experimental design. New York: McGraw-Hill. [Google Scholar]
- 72.Primer6 (2013) PRIMER-E Ltd. 3 Meadow View, Lutton, Ivybridge, PL21 9RH, UK.
- 73.Clarke KR, Gorley RN (2006) PRIMER v6: user manual/tutorial. Plymouth: PRIMER-E. [Google Scholar]
- 74.Littman RA, Willis BL, Bourne DG (2011) Metagenomic analysis of the coral holobiont during a natural bleaching event on the Great Barrier Reef. Environmental Microbiology Reports 3: 651–660. doi: 10.1111/j.1758-2229.2010.00234.x [DOI] [PubMed] [Google Scholar]
- 75.Meron D, Rodolfo-Metalpa R, Cunning R, Baker AC, Fine M, Banin E (2012) Changes in coral microbial communities in response to a natural pH gradient. Isme Journal 6: 1775–1785. doi: 10.1038/ismej.2012.19 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Barott KL, Rodriguez-Brito B, Janouskovec J, Marhaver KL, Smith JE, Keeling P, et al. (2011) Microbial diversity associated with four functional groups of benthic reef algae and the reef-building coral Montastraea annularis. Environmental Microbiology 13: 1192–1204. doi: 10.1111/j.1462-2920.2010.02419.x [DOI] [PubMed] [Google Scholar]
- 77.Yachi S, Loreau M (1999) Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis. Proceedings of the National Academy of Sciences of the United States of America 96: 1463–1468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Ziegler M, Roik A, Porter A, Zubier K, Mudarris M, Ormond R, et al. (2016) Coral microbial community dynamics in response to anthropogenic impacts near a major city in the central Red Sea. Marine Pollution Bulletin 105: 629–640. doi: 10.1016/j.marpolbul.2015.12.045 [DOI] [PubMed] [Google Scholar]
- 79.Miller J, Rogers C, Waara R (2003) Monitoring the coral disease, plague type II, on coral reefs in St. John, U.S. Virgin Islands. Revista Biologica Tropical 51 suppl 4: 47–55. [PubMed] [Google Scholar]
- 80.Sasikala K, Ramana C, Raghuveer Rao P, Subrahmanyam M (1990) Photoproduction of hydrogen, nitrogenase and hydrogenase activities of free and immobilized whole cells of Rhodobacter sphaeroides O.U. 001. FEMS Microbiology Letters 72: 23–28. [Google Scholar]
- 81.Kvennefors EC, Sampayo E, Kerr C, Vieira G, Roff G, Barnes AC (2012) Regulation of bacterial communities through antimicrobial activity by the coral holobiont. Microbial Ecology 63: 605–618. doi: 10.1007/s00248-011-9946-0 [DOI] [PubMed] [Google Scholar]
- 82.Thompson JR, Rivera HE, Closek CJ, Medina M (2015) Microbes in the coral holobiont: partners through evolution, development, and ecological interactions. Frontiers in Cellular and Infection Microbiology 4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Kline DI, Kuntz NM, Breitbart M, Knowlton N, Rohwer F (2006) Role of elevated organic carbon levels and microbial activity in coral mortality. Marine Ecology Progress Series 314: 119–125. [Google Scholar]
- 84.Rosenberg E (2004) The bacterial disease hypothesis of coral bleaching; Rosenberg E, Loya Y, editors. 445–461 p. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional details.
(DOCX)
(A) Acropora millepora (Am) and Turbinaria reniformis (Tr) under control (26.5°C and pCO2 of 364 μatm) and treatment (29.0°C and pCO2 of 750 μatm) conditions for each sample (1–6). (B) Average microbial relative abundances by Class.
(TIFF)
Results for Acropora millepora and Turbinaria reniformis under control (26.5°C and pCO2 of 364μatm) and treatment (29.0°C and pCO2 of 750μatm) conditions. Samples are from colonies numbered 1–6.
(XLSX)
Average percent dissimilarity between species and between treatments within species in microbial OTU abundance. Control = 26.5°C and pCO2 of 364μatm, Treatment = 29.0°C and pCO2 of 750μatm. Am = Acropora millepora, Tr = Turbinaria reniformis, Av.Abund = average abundance, Av.Diss = average dissimilarity, Diss/SD = Dissimilarity /standard deviation, Contrib% = contribution percent, Cum.% = Cumulative percent contribution.
(DOCX)
Effects of species and treatment were fixed and fully crossed. A posteriori slice tests were used to test for differences between treatment and controls within species. df = degrees of freedom, SS = type III sum of squares of the main effects, F = F-statistic. Significant effects are bolded.
(DOCX)
Significant differences (p ≤ 0.05) are bolded. calc = calcification, chla = chlorophyll a, cells = endosymbiotic algal cell density, lipid = total lipids, protein = soluble animal protein concentration, carbs = carbohydrate concentration, biomass = coral ash free dry weight per area, h = host, a = endosymbiotic algae.
(DOCX)
Summary of average squared distance (Av.Sq.Dist = 29.54) in coral physiology between Acropora millepora and Turbinaria reniformis across all control and treatment coral fragments. SD = standard deviation, Av.Value = average, Contrib % = percent contribution, Cum. % = cumulative percent.
(DOCX)
Data Availability Statement
Sequences have been deposited in NCBI’s Sequence Read Archive under accession number PRJNA344759. The physiological data are archived at www.bco-dmo.org/project/528004.