

Abstract
Background
Asthma is a disease of chronic airway inflammation with heterogeneous features. Neutrophilic asthma is corticosteroid-insensitive asthma related to absence or suppression of TH2 process and increased TH1 and/or TH17 process. Macrolides are immunomodulatory drug that reduce airway inflammation, but their role in asthma is not fully known. The purpose of this study was to evaluate the role of macrolides in neutrophilic asthma and compare their effects with those of corticosteroids.
Methods
C57BL/6 female mice were sensitized with ovalbumin (OVA) and lipopolysaccharides (LPS). Clarithromycin (CAM) and/or dexamethasone (DXM) were administered at days 14, 15, 21, 22, and 23. At day 24, the mice were sacrificed.
Results
Airway resistance in the OVA+LPS exposed mice was elevated but was more attenuated after treatment with CAM+DXM compared with the monotherapy group (p<0.05 and p<0.01). In bronchoalveolar lavage fluid study, total cells and neutrophil counts in OVA+LPS mice were elevated but decreased after CAM+DXM treatment. In hematoxylin and eosin stain, the CAM+DXM-treated group showed less inflammation additively than the monotherapy group. There was less total protein, interleukin 17 (IL-17), interferon γ, and tumor necrosis factor α in the CAM+DXM group than in the monotherapy group (p<0.001, p<0.05, and p<0.001). More histone deacetylase 2 (HDAC2) activity was recovered in the DXM and CAM+DXM challenged groups than in the control group (p<0.05).
Conclusion
Decreased IL-17 and recovered relative HDAC2 activity correlated with airway resistance and inflammation in a neutrophilic asthma mouse model. This result suggests macrolides as a potential corticosteroid-sparing agent in neutrophilic asthma.
Keywords: Neutrophils, Asthma, Macrolides, Adrenal Cortex Hormones, Th17 Cells, Inflammation, Histone Deacetylases
Introduction
Asthma is a common chronic airway disorder characterized by variable symptoms, airway obstruction, airway inflammation, and airway remodeling1. Classically, mast cells, T helper type II cells (TH2 cells), and eosinophils play roles in asthma by cytokine-based airway inflammation2. Asthma has a complex syndrome of many different clinical features without predominance, and there are many types, such as non-TH2 cell-related, neutrophilic, and paucigranulocytic asthma3.
The burden of asthma is continuously increasing; low quality of life, disability, morbidity, and socioeconomic issues have been emerging worldwide. Refractory or persistent asthma has been the main focus in previous studies4,5,6,7,8. Asthma can be categorized according to type of underlying inflammation, and much refractory asthma is neutrophilic. Neutrophilic asthma is non-allergic asthma that often does not respond to inhaled or systemic corticosteroids9.
There is some proposed pathobiology of corticosteroid insensitivity in neutrophilic asthma. First, the TH2 process of neutrophilic asthma was absent or suppressed; corticosteroids suppressed eosinophil and T-lymphocyte inflammation but did not prevent neutrophil accumulation10. Instead, T helper 17 cells (TH17) were pivotal and linked with neutrophilic asthma11,12. Second, corticosteroid insensitivity correlated with histone deacetylase-2 (HDAC2) activity and expression, which suggests that drugs that restore HDAC activity can reduce neutrophil recruitment in asthma13.
Macrolides are antibiotics with immunomodulatory properties14,15. They may be effective in patients with severe neutrophilic asthma, but there is a lack of evidence for using macrolides for asthma16,17,18.
In this study, we set a neutrophilic asthma model, describe the possibility of macrolides as a therapeutic agent for severe and refractory neutrophilic asthma, and compare the effectiveness of macrolides, corticosteroids and concurrent treatment in the neutrophilic asthma mouse model.
Materials and Methods
1. Animal and experimental model
In a previous study, we developed eosinophilic asthma and neutrophilic asthma mouse models. Intraperitoneal ovalbumin (OVA) sensitization induced TH2-derived eosinophilic inflammation in the mouse model19,20,21. After the eosinophilic asthma mouse model was sensitized, using lipopolysaccharides (LPS) shifted the airway inflammation from eosinophil-predominant to neutrophil-predominant and highly correlated with levels of interleukin (IL)-17A and TH17 cells22. Modifying previous studies, we sensitized 6-week-old female C57BL/6 mice (Orient Inc., Seongnam, Korea) with peritoneal injection of 100 µg of OVA grade V (Sigma-Aldrich, St. Louis, MO, USA) that was dissolved in 2 mg aluminum hydroxide (Sigma-Aldrich) at days 0 and 7. Mice were exposed to intranasal 50 µg OVA at days 14, 15, 21, 22, and 23 to induce eosinophilic inflammation. We used intranasal 10 µg LPS from Escherichia coli O11:B4 (Sigma-Aldrich) to induce neutrophilic inflammation at days 21, 22, and 23.
2. Drug challenges
Figure 1 describes the drug challenges in the neutrophilic asthma mouse model. We randomly allocated the mice to the following groups: (1) control, (2) OVA+LPS+0.5% carboxymethyl cellulose (CMC), (3) OVA+LPS+clarithromycin (CAM) (10 mg/kg), (4) OVA+LPS+dexamethasone (DXM; 1 mg/kg)+CMC, and (5) OVA+LPS+CAM (10 mg/kg)+DXM (1 mg/kg).
Figure 1. Schematic outline of the neutrophilic asthma mouse model. Mice were sensitized via OVA on days 0 and 7 by i.p. injection and challenged through inhalation of OVA on days 14, 15, 21, 22, and 23. For neutrophilic asthma, mice were additionally challenged with LPS on days 21, 22, and 23. DXM by i.p. injection and CAM via oral gavage were challenged every 24 hours on days 14, 15, 16, 17, 18, 21, and 23. OVA: ovalbumin; i.p.: intraperitoneally; LPS: lipopolysaccharide; DXM: dexamethasone; CAM: clarithromycin; i.n: intranasally.

The control group comprised five mice, and the drug-challenge groups comprised seven. We used 0.5% CMC (Sigma-Aldrich) as a vehicle to compare with other groups. The mice were exposed to peritoneal injection of 1 mg/kg DXM (Sigma-Aldrich) and 10 mg/kg CAM (Sigma-Aldrich) via oral gavage every 24 hours on days 14, 15, 16, 17, 18, 21, 22, and 23. The mice were sacrificed at day 24. All animal protocols were approved by the Animal Subjects Committee of the Catholic University of Korea.
3. Airway responsiveness
We calculated airway hyperresponsiveness by respiratory system resistance (Rrs) with methacholine (Mch; Sigma-Aldrich) in the FlexiVent system (SCIREQ, Montreal, QC, Canada)23,24. Mice were exposed to nebulized phosphate-buffered saline (PBS) for 3 minutes for baseline Rrs value, and we assessed Rrs after challenging them with increasing concentrations of Mch (6.25, 12.5, 25, and 50 mg/mL; Sigma-Aldrich) in PBS using an Aerosonic ultrasonic nebulizer (DeVilbiss, Somerset, PA, USA). We averaged and expressed the Rrs measured during each 3-minute sequence for each Mch concentration.
4. Bronchoalveolar lavage
The trachea was exposed and cannulated with silicone tubing attached to a 22-gauge needle on a 1-mL tuberculin syringe. We withdrew bronchoalveolar lavage (BAL) fluid after instilling 0.8 mL cold sterile PBS through the trachea into the lung. We cytospun the recovered BAL fluid for 10 minutes at 1,000 ×g at 4℃ and stored supernatant at –80℃ until required for further analysis. We counted total cells in the BAL fluid us-ing a hemocytometer. After we adjusted the number of cells to 1×106/mL, we centrifuged 50 µL aliquots of BAL fluid onto microscope slides for 5 minutes at 43 g and stained the slides with Diff-Quick (Sysmax, Kobe, Japan). We obtained the percentages of macrophages, eosinophils, lymphocytes, and neutrophils in the BAL fluid by counting 500 leucocytes in randomly selected fields of the slides under a light microscope.
5. Lung tissue preparation for histopathology
After the mice were killed, their lungs were inflated, fixed in 4% paraformaldehyde for 24 hours, and embedded in paraffin wax. We cut sections at a thickness of 4 µm using a microtome and subjected the deparaffinized tissue sections to hematoxylin and eosin (H&E) staining for microscopic examination (magnification of ×200).
6. Measurement of cytokines
We measured the concentrations of IL-5 and IL-17 in the BAL fluids and IL-13, interferon γ(IFN-γ), and tumor necrosis factor α(TNF-α) in the lung homogenates using an enzyme-linked immunosorbent assay kit (R&D Systems, Minneapolis, MN, USA). The assay sensitivities were 7, 5, 2, and 7.21 pg/mL, respectively. We conducted the protocol according to manufacturer's instructions.
7. Western blot analysis
We disrupted separated lung tissues that had been immediately frozen in liquid nitrogen using a Polytron homogenizer (Pellet pestles cordless motor; Sigma) and centrifuged them. We purified the proteins from the supernatant and assessed the concentrations using the Bradford method. We separated the protein samples by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred them to a polyvinylidene difluoride membrane (Amersham Pharmacia Biotech, Little Chalfont, UK). After blocking with 5% skimmed milk (Difco, BD, San Jose, CA, USA), we incubated the membrane overnight with an anti-HDAC2 antibody (1:1,000, rabbit monoclonal; Abcam, Cambridge, UK). We washed the membrane three times with TBST and incubated it with a secondary antibody for 2 hours. Horseradish peroxidase-conjugated goat anti rabbit IgG (1:2,000, Santa Cruz Biotech, Dallas, TX, USA) was used as the secondary antibody. We detected the target proteins using the ECL Western Blotting Analysis System (Thermo Fisher Scientific, Rockford, IL, USA). We either exposed the membrane to X-ray film or analyzed it with an LAS 3000 (Fujifilm, Tokyo, Japan) image analyzer using Multi Gauge v. 3.0 software. We also quantified the HDAC2 and ß-actin band intensities with Multi Gauge.
8. HDAC2 activity
We measured HDAC2 activity using the colorimetric EpiQuik HDAC2 Activity Assay Kit (Epigentek, Brooklyn, NY, USA). We incubated the nuclear extracts with specific substrate for 90 minutes at 37℃, followed by capture antibody for 60 minutes, and detection antibody for 30 minutes at room temperature. We measured absorbance at 450 nm in a microelisa reader.
9. Statistical analysis
All data are expressed as the means±SEM, and we used GraphPad Prism 5.01 (GraphPad Software, San Diego, CA, USA) for the data analysis. We performed the statistical analyses using one-way ANOVA and t tests and considered p<0.05 to indicate statistical significance.
Results
1. Combination therapy of macrolide and corticosteroid attenuates airway resistance in the neutrophilic asthma mouse model
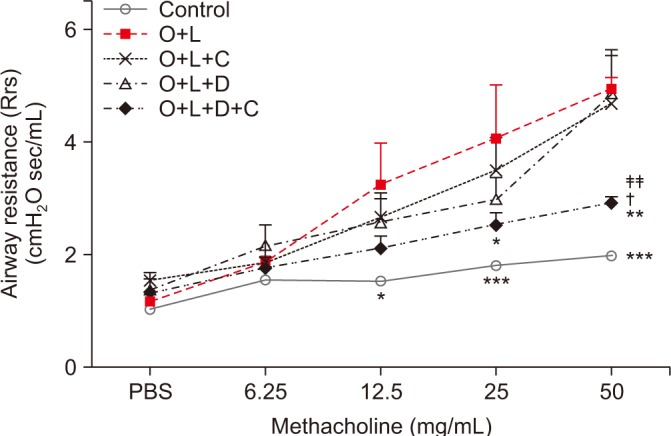
The mice in the OVA+LPS group showed greater airway resistance after they received 12.5 mg/mL of Mch than did the control group (p<0.05) (Figure 2). CAM or DXM monotherapy group did not decrease Rrs after 12.5, 25, or 50 mg/mL of Mch, but the OVA+LPS-challenged mice treated with CAM+DXM showed decreased Rrs at 25 and 50 mg/mL of Mch challenge (p<0.05 and p<0.01). The change was significant compared with findings in the CAM and DXM monotherapy groups, especially after 50 mg/mL of Mch (p<0.05 and p<0.01).
Figure 2. Airway responsiveness to methacholine by forced oscillation technique. Airway responsiveness was expressed in terms of respiratory systemic Rrs. Baseline Rrs value was assessed after 3 minutes of nebulized PBS. Rrs was measured in response to increasing challenges of methacholine dose (6.25 mg/mL, 12.5 mg/mL, 25 mg/mL, and 50 mg/mL) in control, O+L, O+L+C, O+L+D, and O+L+D+C group. Rrs was increased after OVA+LPS challenge. Rrs of combination of CAM+DXM group was significantly changed compared to the CAM or DXM group especially at 50 mg/mL of methacholine challenge. O+L vs. all (*p<0.05, **p<0.01, ***p<0.001), O+L+C vs. O+L+D+C (†p<0.05), O+L+D vs. O+L+D+C (‡‡p<0.01). Rrs: resistance; PBS: phosphate buffered saline; O: ovalbumin; L: lipopolysaccharide; C: clarithromycin; D: dexamethasone.
2. Combination therapy of macrolide and corticosteroid reduces lung inflammation in the neutrophilic asthma mouse model
In BAL fluid, total cell counts were higher in OVA+LPS-challenged mice (p<0.001) (Figure 3A). These mice that were treated with CAM+DXM revealed lower total cell counts (p<0.001) compared with those in the monotherapy groups. We conducted differential cell counts to evaluate reduced inflammation in the pathways based on microscopic morphological criteria. In OVA+LPS-challenged mice, alveolar neutrophil counts percentage was 20.7% and eosinophil counts percentage was 18.6%, which is higher compared to control group (0.2% and 0%). The numbers of alveolar neutrophils, eosinophils, and lymphocytes did not decrease with CAM or DXM monotherapy, but with the two together, the numbers of neutrophils decreased (16.3%, p<0.05) but not eosinophils (16.3%). On H&E staining, inflammatory cell accumulation improved in the DXM monotherapy group. In the CAM+DXM group, accumulation of inflammation cells improved more than it did in the monotherapy groups (Figure 3B).
Figure 3. Combination therapy of macrolide and corticosteroid inhibited lung inflammation in murine model of neutrophilic asthma. (A) BALF analysis was performed to evaluate lung inflammation. The number of total cell counts, eosinophil, and neutrophil was significantly increased after OVA+LPS challenge. The number of total cell counts was significantly more decreased in CAM+DXM group compared to decrease in CAM or DXM group. Neutrophils in combination of CAM+DXM treatment group was decreased and it was not seen in CAM or DXM group. The number of eosinophils was not decreased in CAM, DXM, and CAM+DXM group. (B) Accumulation of inflammatory cells improved significantly more in CAM+DXM group compared to DXM group in H&E staining. O+L vs. all (*p<0.05, **p<0.01, ***p<0.001), O+L+C vs. O+L+D+C (†p<0.05, ††p<0.01, †††p<0.001), O+L+D vs. O+L+D+C (‡‡p<0.01, ‡‡‡p<0.001). BALF: bronchoalveolar lavage fluid; O: ovalbumin (OVA); L: lipopolysaccharide (LPS); C: clarithromycin (CAM); D: dexamethasone (DXM).

3. Macrolides have more potent effects than corticosteroids on attenuating TH17 inflammation in the neutrophilic asthma mouse model
OVA+LPS challenge increased the total protein amounts in the BAL fluid (p<0.01). After treatment with DXM monotherapy and a combination of CAM+DXM, total protein amounts in the BAL fluid decreased (p<0.001). Protein amounts were lower in the combination group than in the DXM monotherapy group, but difference was not statistically significant (Figure 4A).
Figure 4. TH17 inflammation was attenuated by combination of macrolide and corticosteroid. Total protein, TH2 inflammation, TH17 inflammation, and proinflammatory cytokine in BALF analysis was increased after OVA+LPS challenge. (A) The number of total protein in BALF was decreased in DXM and CAM+DXM group. (B) IL-5 in BALF, marker of TH2 inflammation, was not significantly decreased in DXM or CAM+DXM group. (C) IL-17, marker of TH17 inflammation, was significantly decreased in DXM and CAM+DXM group. (D) IL-13, marker of TH2 inflammation, was evaluated in lung tissue. There was no significant decrease in DXM or CAM+DXM group. (E, F) Proinflammatory cytokine was decreased in combination of CAM+DXM and it was not decreased in monotherapy of CAM or DXM. O+L vs. all (*p<0.05, **p<0.01, ***p<0.001), O+L+C vs. O+L+D+C (†p<0.05, †††p<0.001). BALF: bronchoalveolar lavage fluid; O: ovalbumin (OVA); L: lipopolysaccharide (LPS); C: clarithromycin (CAM); D: dexamethasone (DXM); IL: interleukin; IFN-γ: interferon γ; TNF-α: tumor necrosis factor α.

TH2 inflammation marker (IL-5 and IL-13) and TH17 inflammation marker (IL-17A) were induced after OVA+LPS challenge in the mouse model (p<0.05; Figure 4B, C and p<0.001; Figure 4D); IFN-γ, and TNF-αincreased also (p<0.01) (Figure 4E, F). After treatment with CAM only, IL-5 and IL-13 decreased (p<0.05 for both). After treatment with DXM, total protein (p<0.001) (Figure 4A) and IL-17A decreased (p<0.01) (Figure 4B, C). We also noted decreases in the CAM+DXM group. IFN-γand TNF-αdecreased with CAM+DXM, but there was no attenuation in either the CAM or the DXM monotherapy groups (p<0.05) (Figure 4E, F).
4. Macrolides act on HDAC2 activity and have potent effects with corticosteroids
HDAC activity is correlated to corticosteroid activity and decreases in many cases of corticosteroid insensitivity25. HDAC2 activity in OVA+LPS challenged mice was lower than the activity in the control group (p<0.01) (Figure 5A) and it was not recovered after CAM treatment. HDAC2 activity was more recovered in the DXM-challenged group than in the control group (p<0.01) (Figure 5A). The CAM+DXM-challenged group recovered more HDAC2 activity than either the control or the CAM-challenged groups (p<0.05 for both) (Figure 5A).
Figure 5. Macrolide has potent effects of recovering HDAC2 activity with corticosteroid. (A) HDAC2 activity was decreased after OVA+LPS challenge. It was not recovered after CAM treatment, but it was recovered after DXM or CAM+DXM treatment. (B) DXM or DXM+CAM group showed higher density than OVA+LPS or CAM group in western blot analysis. (C) Relative density of HDAC2/β-actin was well correlated these tendency and CAM+DXM group was further increased in density of HDAC2/β-actin than DXM group. O+L vs. all (*p<0.05, **p<0.01), O+L+C vs. O+L+D+C (†p<0.05, †††p<0.001), O+L+D vs. O+L+D+C (‡‡p<0.01). HDAC2: histone deacetylase-2; O: ovalbumin (OVA); L: lipopolysaccharide (LPS); C: clarithromycin (CAM); D: dexamethasone (DXM).
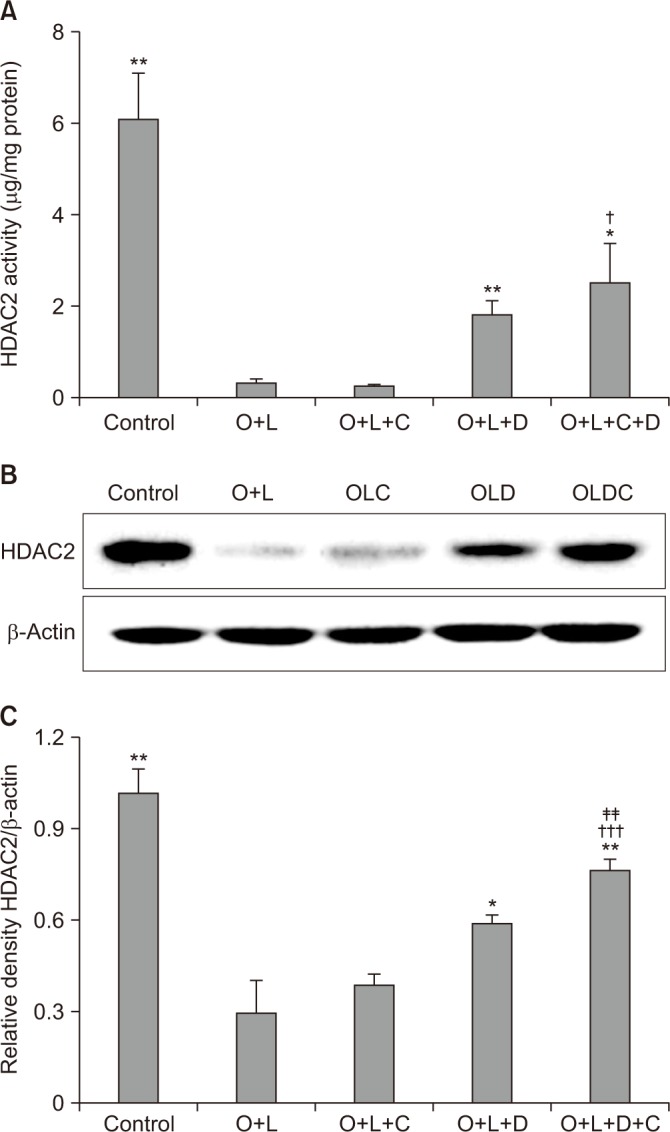
Relative density analysis of HDAC2/β-actin showed significant differences. In the OVA+LPS-challenged mice, the relative density of HDAC2/β-actin was lower than in the control group (p<0.01) (Figure 5C); it was not significantly higher in the CAM-challenged group than in the control group. In the DXM-challenged group, relative density of HDAC2/β-actin was greater than that in the control group (p<0.05) (Figure 5C), and it was greater in the CAM+DXM-challenged group than in the DXM group (p<0.01) (Figure 5C).
Discussion
We developed a neutrophilic asthma mouse model by challenging OVA+LPS to C57BL/6 female mice, and we found neutrophilia and eosinophilia in the OVA+LPS murine model. We demonstrated that macrolide and corticosteroid treatment reduced airway inflammation through attenuating airway resistance in the neutrophilic asthma mouse model. A combination of macrolide and corticosteroid reduced the neutrophil counts in BAL fluid but not eosinophil counts. TH17 inflammation marker was attenuated by corticosteroid, and the macrolide potentiated the effect of the corticosteroid. Pro-inflammatory cytokines did not decrease with macrolide alone or corticosteroid alone but with the combination of the two. The combination of macrolide and corticosteroid recovered relative HDAC2 activity that accounted for improvement in corticosteroid insensitivity.
Neutrophilic asthma is characterized by sputum neutro-philia, low lung function, more trapped air, and thicker airway walls than non-neutrophilic asthma; little is understood about its related molecular elements and pathophysiology12,26,27. It has remained controversial whether neutrophilia is an independent driving component and synergistic factor with eosinophilia or a consequence of corticosteroid therapy.
Airway resistance and airway inflammation on H&E stain were not attenuated by macrolide alone or corticosteroid alone but with the combination of the two, the neutrophil count decreased. This demonstrates that macrolide and corticosteroid together decreased airway resistance and inflammation by attenuating neutrophilic inflammation.
Neutrophilic asthma often responds less to inhaled corticosteroids or to high-dose of systemic corticosteroids. There is no proven mechanism of corticosteroid insensitivity in neutrophilic asthma. Characteristics of neutrophilic asthma, such as more fixed airway obstruction and thicker airway walls, may be related to unresponsiveness to corticosteroids. Previous studies revealed that the corticosteroid insensitivity in neutrophilic asthma is because of the absence of or decreased TH2 inflammation mechanism and increased TH1 and/or TH17 inflammation. Murine models have similar helper T cell phenotypes to those in human28. TH2 and TH17 cytokines and pro-inflammatory cytokines were elevated in the OVA+LPS murine model. TH2 inflammation did not decrease with DXM or CAM+DXM. TH17 inflammation did decreased with DXM and CAM to potentiate its effect. Pro-inflammatory cytokines decreased only with CAM+DXM, which reveals that the macrolide potentiated the corticosteroid to attenuate TH17 inflammation, which is evidence that CAM has an adjunctive rather than direct effect on DXM in attenuating the inflammation cascade.
Macrolides have a wide spectrum of functions in the airway, such as reducing airway secretion, chronic inflammation, bacterial loads, and virulence29. Macrolides have an action of immunomodulation, host-pathogen interaction, regulating signaling pathways and cytokine response, reducing oxidative stress, and controlling innate immunity30. Macrolides are well-known in chronic obstructive pulmonary disease (COPD), and the inflammation in neutrophilic asthma has similar aspects to COPD31. Evidence of the benefits of macrolides in asthma is scant, but they have been widely used for various conditions of asthma, especially severe refractory asthma. Simpson et al.17 documented that clarithromycin treatment in severe uncontrolled asthma modulated IL-8 and the accumulation and activation of neutrophils. Brusselle et al.18 demonstrated that azithromycin was effective in reducing acute and severe exacerbation in severe non-eosinophilic asthma patients. There have been some proposed mechanisms of macrolide. Shinkai et al.15 suggested that one possible mechanism is that it suppresses IL-8 and activates extracellular signal-regulated kinase reversely related to neutrophil migration.
There is another action of macrolides in neutrophilic asthma. Corticosteroid insensitivity correlated with airway hyperresponsiveness with decreased HDAC2 activity and expression13. HDAC2 can inhibit inflammatory protein coding genes such as granulocyte macrophage colony stimulating factor or cyclooxygenase 2, promoted by IL-1β, TNF-α, or nuclear factor κB kinase32. Macrolides reversed corticosteroid insensitivity by restoring HDAC activity and activating phosphoinositol 3 kinase33. This suggests that macrolides may reduce neutrophil recruitment in asthma by restoring corticosteroid insensitivity13.
HDAC is a key protein in corticosteroid insensitivity. We decided to check HDAC2 activity and relative density of HDAC2/β-actin, and the macrolide did not restore HDAC2 activity; the activity was restored after treatment with DXM or CAM+DXM, which showed that the macrolide was not directly related to HDAC2 activity. Relative density of HDAC2/β-actin was significantly higher in the OVA+LPS+CAM+DXM group than in the OVA+LPS+DXM group, which suggested that the corticosteroid restored HDAC2 activity and the macrolide acted indirectly with the corticosteroid to potentiate its effects on HDAC2 activity. This finding suggests that macrolides have corticosteroid-sparing effects. Finally, the macrolide in this study potentiated corticosteroid`s effects on attenuating airway hyperresponsiveness and inflammation by restoring HDAC2 activity.
Our study had limitations. We investigated a novel neutro-philic asthma model, but novel model needed to be validated for compatibility. Although we validated the neutrophilic mouse model's compatibility at a precedent-setting study, additional studies with novel models are needed to demonstrate the reproducibility of these study results. Another limitation was that we demonstrated a possible relationship between airway inflammation and CAM via HDAC2 activity recovery, but we did not ultimately validate the mechanism in neutro-philic asthma. Additional investigations are needed to identify the precise pathways of the mechanism. Finally, even if the asthma inflammation phenotype in the mouse model was not different from that in humans, these findings still need to be validated in human studies. Future research is will needed that analyzes the long-term outcomes of macrolides in neutrophilic asthma.
In conclusion, for this study, we set a neutrophilic asthma mouse model and demonstrated how a macrolide acted with a corticosteroid to adjunctively reduce airway hyperresponsiveness and inflammation in the mouse model. The activity may be related to attenuation of TH17 inflammation and recovery of HDAC2 activity, and the findings suggest macrolides as potential corticosteroid-sparing agents in neutrophilic asthma.
Footnotes
Conflicts of Interest: No potential conflict of interest relevant to this article was reported.
References
- 1.Reddel HK, Hurd SS, FitzGerald JM. World Asthma Day. GINA 2014: a global asthma strategy for a global problem. Int J Tuberc Lung Dis. 2014;18:505–506. doi: 10.5588/ijtld.14.0246. [DOI] [PubMed] [Google Scholar]
- 2.Pelaia G, Vatrella A, Maselli R. The potential of biologics for the treatment of asthma. Nat Rev Drug Discov. 2012;11:958–972. doi: 10.1038/nrd3792. [DOI] [PubMed] [Google Scholar]
- 3.Wenzel SE. Complex phenotypes in asthma: current definitions. Pulm Pharmacol Ther. 2013;26:710–715. doi: 10.1016/j.pupt.2013.07.003. [DOI] [PubMed] [Google Scholar]
- 4.Barry LE, Sweeney J, O'Neill C, Price D, Heaney LG. The cost of systemic corticosteroid-induced morbidity in severe asthma: a health economic analysis. Respir Res. 2017;18:129. doi: 10.1186/s12931-017-0614-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Accordini S, Corsico AG, Braggion M, Gerbase MW, Gislason D, Gulsvik A, et al. The cost of persistent asthma in Europe: an international population-based study in adults. Int Arch Allergy Immunol. 2013;160:93–101. doi: 10.1159/000338998. [DOI] [PubMed] [Google Scholar]
- 6.Murray CJ, Vos T, Lozano R, Naghavi M, Flaxman AD, Michaud C, et al. Disability-adjusted life years (DALYs) for 291 diseases and injuries in 21 regions, 1990-2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet. 2012;380:2197–2223. doi: 10.1016/S0140-6736(12)61689-4. [DOI] [PubMed] [Google Scholar]
- 7.Lai CK, Kim YY, Kuo SH, Spencer M, Williams AE. Cost of asthma in the Asia-Pacific region. Eur Respir Rev. 2006;15:10–16. [Google Scholar]
- 8.Bahadori K, Doyle-Waters MM, Marra C, Lynd L, Alasaly K, Swiston J, et al. Economic burden of asthma: a systematic review. BMC Pulm Med. 2009;9:24. doi: 10.1186/1471-2466-9-24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Alam R, Good J, Rollins D, Verma M, Chu H, Pham TH, et al. Airway and serum biochemical correlates of refractory neutrophilic asthma. J Allergy Clin Immunol. 2017;140:1004–1014. doi: 10.1016/j.jaci.2016.12.963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Green RH, Brightling CE, Woltmann G, Parker D, Wardlaw AJ, Pavord ID. Analysis of induced sputum in adults with asthma: identification of subgroup with isolated sputum neutrophilia and poor response to inhaled corticosteroids. Thorax. 2002;57:875–879. doi: 10.1136/thorax.57.10.875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lambrecht BN, Hammad H. The immunology of asthma. Nat Immunol. 2015;16:45–56. doi: 10.1038/ni.3049. [DOI] [PubMed] [Google Scholar]
- 12.Wenzel SE. Asthma phenotypes: the evolution from clinical to molecular approaches. Nat Med. 2012;18:716–725. doi: 10.1038/nm.2678. [DOI] [PubMed] [Google Scholar]
- 13.Ito K, Herbert C, Siegle JS, Vuppusetty C, Hansbro N, Thomas PS, et al. Steroid-resistant neutrophilic inflammation in a mouse model of an acute exacerbation of asthma. Am J Respir Cell Mol Biol. 2008;39:543–550. doi: 10.1165/rcmb.2008-0028OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kanoh S, Rubin BK. Mechanisms of action and clinical application of macrolides as immunomodulatory medications. Clin Microbiol Rev. 2010;23:590–615. doi: 10.1128/CMR.00078-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Shinkai M, Henke MO, Rubin BK. Macrolide antibiotics as immunomodulatory medications: proposed mechanisms of action. Pharmacol Ther. 2008;117:393–405. doi: 10.1016/j.pharmthera.2007.11.001. [DOI] [PubMed] [Google Scholar]
- 16.Wong EH, Porter JD, Edwards MR, Johnston SL. The role of macrolides in asthma: current evidence and future directions. Lancet Respir Med. 2014;2:657–670. doi: 10.1016/S2213-2600(14)70107-9. [DOI] [PubMed] [Google Scholar]
- 17.Simpson JL, Powell H, Boyle MJ, Scott RJ, Gibson PG. Clarithromycin targets neutrophilic airway inflammation in refractory asthma. Am J Respir Crit Care Med. 2008;177:148–155. doi: 10.1164/rccm.200707-1134OC. [DOI] [PubMed] [Google Scholar]
- 18.Brusselle GG, Vanderstichele C, Jordens P, Deman R, Slabbynck H, Ringoet V, et al. Azithromycin for prevention of exacerbations in severe asthma (AZISAST): a multicentre randomised double-blind placebo-controlled trial. Thorax. 2013;68:322–329. doi: 10.1136/thoraxjnl-2012-202698. [DOI] [PubMed] [Google Scholar]
- 19.Bergquist M, Jonasson S, Hjoberg J, Hedenstierna G, Hanrieder J. Comprehensive multiplexed protein quantitation delineates eosinophilic and neutrophilic experimental asthma. BMC Pulm Med. 2014;14:110. doi: 10.1186/1471-2466-14-110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zhang F, Huang G, Hu B, Qian GS, Song Y. Recombinant HMGB1 A box protein inhibits Th17 responses in mice with neutrophilic asthma by suppressing dendritic cell-mediated Th17 polarization. Int Immunopharmacol. 2015;24:110–118. doi: 10.1016/j.intimp.2014.11.005. [DOI] [PubMed] [Google Scholar]
- 21.Zhao Y, Huang Y, He J, Li C, Deng W, Ran X, et al. Rosiglitazone, a peroxisome proliferator-activated receptor-gamma agonist, attenuates airway inflammation by inhibiting the proliferation of effector T cells in a murine model of neutrophilic asthma. Immunol Lett. 2014;157:9–15. doi: 10.1016/j.imlet.2013.11.004. [DOI] [PubMed] [Google Scholar]
- 22.Zhao S, Jiang Y, Yang X, Guo D, Wang Y, Wang J, et al. Lipopolysaccharides promote a shift from Th2-derived airway eosinophilic inflammation to Th17-derived neutrophilic inflammation in an ovalbumin-sensitized murine asthma model. J Asthma. 2017;54:447–455. doi: 10.1080/02770903.2016.1223687. [DOI] [PubMed] [Google Scholar]
- 23.McGovern TK, Robichaud A, Fereydoonzad L, Schuessler TF, Martin JG. Evaluation of respiratory system mechanics in mice using the forced oscillation technique. J Vis Exp. 2013;(75):e50172. doi: 10.3791/50172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Shalaby KH, Gold LG, Schuessler TF, Martin JG, Robichaud A. Combined forced oscillation and forced expiration measurements in mice for the assessment of airway hyperresponsiveness. Respir Res. 2010;11:82. doi: 10.1186/1465-9921-11-82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Marwick JA, Ito K, Adcock IM, Kirkham PA. Oxidative stress and steroid resistance in asthma and COPD: pharmacological manipulation of HDAC-2 as a therapeutic strategy. Expert Opin Ther Targets. 2007;11:745–755. doi: 10.1517/14728222.11.6.745. [DOI] [PubMed] [Google Scholar]
- 26.Wenzel SE, Szefler SJ, Leung DY, Sloan SI, Rex MD, Martin RJ. Bronchoscopic evaluation of severe asthma: persistent inflammation associated with high dose glucocorticoids. Am J Respir Crit Care Med. 1997;156(3 Pt 1):737–743. doi: 10.1164/ajrccm.156.3.9610046. [DOI] [PubMed] [Google Scholar]
- 27.Jatakanon A, Uasuf C, Maziak W, Lim S, Chung KF, Barnes PJ. Neutrophilic inflammation in severe persistent asthma. Am J Respir Crit Care Med. 1999;160(5 Pt 1):1532–1539. doi: 10.1164/ajrccm.160.5.9806170. [DOI] [PubMed] [Google Scholar]
- 28.Annunziato F, Romagnani S. Mouse T helper 17 phenotype: not so different than in man after all. Cytokine. 2011;56:112–115. doi: 10.1016/j.cyto.2011.06.009. [DOI] [PubMed] [Google Scholar]
- 29.Spagnolo P, Fabbri LM, Bush A. Long-term macrolide treatment for chronic respiratory disease. Eur Respir J. 2013;42:239–251. doi: 10.1183/09031936.00136712. [DOI] [PubMed] [Google Scholar]
- 30.Friedlander AL, Albert RK. Chronic macrolide therapy in inflammatory airways diseases. Chest. 2010;138:1202–1212. doi: 10.1378/chest.10-0196. [DOI] [PubMed] [Google Scholar]
- 31.Kim RY, Pinkerton JW, Gibson PG, Cooper MA, Horvat JC, Hansbro PM. Inflammasomes in COPD and neutrophilic asthma. Thorax. 2015;70:1199–1201. doi: 10.1136/thoraxjnl-2014-206736. [DOI] [PubMed] [Google Scholar]
- 32.Barnes PJ. Corticosteroid resistance in patients with asthma and chronic obstructive pulmonary disease. J Allergy Clin Immunol. 2013;131:636–645. doi: 10.1016/j.jaci.2012.12.1564. [DOI] [PubMed] [Google Scholar]
- 33.Kobayashi Y, Wada H, Rossios C, Takagi D, Charron C, Barnes PJ, et al. A novel macrolide/fluoroketolide, solithromycin (CEM-101), reverses corticosteroid insensitivity via phosphoinositide 3-kinase pathway inhibition. Br J Pharmacol. 2013;169:1024–1034. doi: 10.1111/bph.12187. [DOI] [PMC free article] [PubMed] [Google Scholar]