

Abstract
Background
The control and prevention of dengue largely depends on vector control measures, environmental management, and personal protection. Dengue control programmes are facing great challenges due to development of insecticide resistance among vector mosquitoes. Information on susceptibility status to different insecticides is important for national programmes to formulate vector control strategies.
Methods
We have studied the larval susceptibility of Aedes albopictus to temephos and adult susceptibility to 4% DDT, 0.05% deltamethrin, and 5% malathion as per WHO protocols in the northern districts of West Bengal. Polymorphisms in the VGSC gene were studied by direct sequencing of PCR products.
Results
The Ae. albopictus larval population showed sensitive [Resistance Ratio (RR99)<3] to moderate levels of resistance (5<RR99>10) to temephos at different study sites. Adult bioassay results revealed that Ae. albopictus was highly resistant to DDT [Corrected Mortality (CM) < 90%] in all the study sites and susceptible to deltamethrin and malathion (CM > 98%), except in Dhupguri where a low level of resistance to deltamethrin (CM = 96.25%) was recorded. None of the six important kdr mutations (S953P, I975M/V, L978, V980G, F1474C, D1703Y) were found in the VGSC of studied mosquitoes, but we identified 11 synonymous and 1 non-synonymous mutation in the VGSC gene.
Conclusion
The higher susceptibility level to deltamethrin and malathion, along with the absence of important kdr mutations indicates that these two insecticides are still effective against Ae. albopictus in the study areas. The susceptibility status of temephos should be monitored closely as low to moderate levels of resistance were observed in few sites. A similar study is recommended for monitoring and early detection of insecticide resistance in other parts of the country.
Author summary
Dengue is one of the most important arboviral infections in India, and transmitted by Aedes mosquitoes. Control of dengue largely depends on effective vector control measures as no specific drugs or vaccines are available, to date. The knowledge of insecticide susceptibility status for local vector mosquitoes is essential for the formulation of effective vector control measures. Therefore, regular monitoring of insecticide susceptibility is essential for the early detection of resistance. In the present study, sensitive to moderate levels of resistance to temephos were recorded among the Aedes albopictus larval populations of different study sites. Adult Ae. albopictus were highly resistant to DDT and susceptible to deltamethrin and malathion in all the study sites, except one site where a low level of resistance to deltamethrin was observed. By sequencing the VGSC gene we observed that the studied Ae. albopictus population does not contain any of the important kdr mutations which are associated with DDT and pyrethroid resistance in other insects. We found only one non-synonymous mutation at codon S1485L, but its role in pyrethroid resistance is yet to be established.
Introduction
Dengue is a mosquito-borne flavi-viral disease and a major public health problem in more than 120 countries [1, 2]. In recent years, dengue transmission has increased predominantly in urban, semi-urban areas and has even extended to the rural areas, becoming a major public health concern globally. A recent estimate showed 390 million new dengue infections throughout the world, of which, 96 million cases manifested the severe form of the disease [2] and almost half of the world’s population are at risk of dengue infection [3]. In India, dengue is spreading into new areas and emerging as a major public health problem. In 2016, a total of 129166 dengue cases and 245 deaths were reported from India, of which 22865 cases and 45 deaths were reported from West Bengal [4]. Aedes aegypti and Aedes albopictus are the vectors of dengue along with three other important human viral diseases: yellow fever, chikungunya, and Zika. No effective vaccine against dengue is available to date. Vector control and personal protection from mosquito bites are suggested to reduce its transmission. For proper formulation and implementation of vector control strategies, thorough information about vector species distribution and their susceptibility to available insecticidal agents are necessary [5].
Four different classes of insecticides are in use as adulticides against Aedes mosquitoes: organophosphates, pyrethroids, organochlorines, and carbamates [6, 7]. Among these, pyrethroids and organophosphates are widely used throughout the world [8, 9, 10]. Pyrethroids are used as indoor residual treatment and impregnation of bed nets whereas organophosphates are used as larvicides and space treatments [6]. The National Vector Borne Disease Control Programme (NVBDCP) of India recommends different insecticides for vector management, such as temephos (50 EC) as a larvicide, DDT and synthetic pyrethroids (recently introduced) for indoor residual spray (IRS), deltamethrin (pyrethroid) for impregnation of bed nets, and malathion for ultra low volume (ULV) spray. In India, Aedes mosquito control is mainly based on anti-larval measures and the use of insecticides by space spraying of pyrethrum and fogging of malathion during a disease outbreak to kill adults. The development and spread of resistance by the vector mosquitoes against all available insecticides is a great challenge to prevent the transmission of mosquito-borne diseases. Ae. albopictus and Ae. aegypti showed resistance to DDT [11, 12, 13], but were susceptible to malathion and deltamethrin [11, 12, 14, 15] in different parts of India. Pyrethroids are synthetic analogues of naturally occurring pyrethrum from the extracts of the Chrysanthemum flower and represent the most widely used insecticide against insect vectors [16]. Unfortunately, pyrethroid efficacy is being threatened due to rapid development of resistance by the vector mosquitoes [8, 17]. The World Health Organisation (WHO) formulated standard diagnostic bioassay test kits to monitor the susceptibility of mosquitoes against different insecticides [18].
Exposure to pyrethroids and DDT results in “knockdown” (i.e., rapid paralysis) due to prolonged-activation of sodium channels. Pyrethroids and organochlorines cause overstimulation of the mosquito nervous system by repeated action potentials form the opening of the sodium channel [19, 20, 21]. Knockdown resistance (kdr) is the major mechanism of pyrethroid resistance, caused by mutations in the voltage-gated sodium channel gene (VGSC gene) [22, 23]. In insects, the voltage-gated sodium channel is an integral transmembrane protein which is composed of four homologous domains (I-IV). Each domain consists of six subunits (S1-S6) which are connected by loops. The segments S5, S6, and the P-loop between them form a central aqueous pore, and the S1-S4 segments of each domain unite to form four independent voltage-sensitive domains [24, 25]. Insects have only one functional sodium channel gene [19]. There are two receptor sites in the four-domain sodium channel for simultaneous binding of pyrethroids [26].So far, ten different mutations at eight codons comprising fifteen haplotypes have been reported in Ae. aegypti. The frequency of these mutations varies geographically [27, 28] but such reports from India are very rare.
Periodical monitoring of insecticide resistance among the prevailing vector population in a given geographical region will be helpful to formulate vector control strategies by the NVBDCP. The present work was designed to study the susceptibility status of Ae. albopictus to temephos, DDT, deltamethrin, and malathion, as well as polymorphisms in the VGSC gene in dengue endemic areas of northern West Bengal.
Materials and methods
Study sites
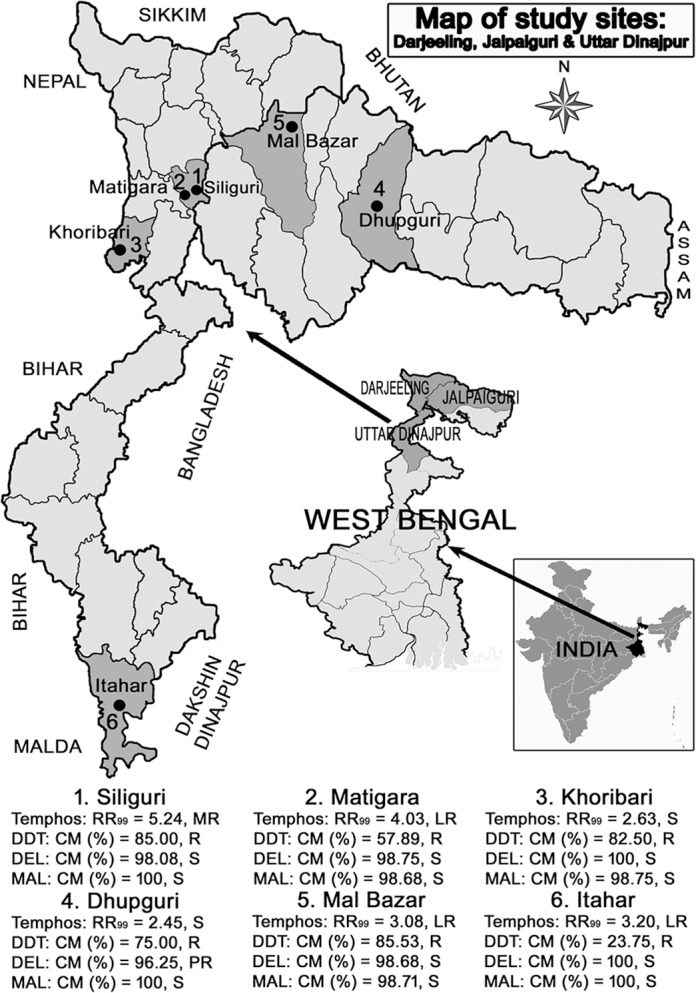
This study was carried out in one municipality and two blocks of Darjeeling, two blocks of Jalpaiguri, and one block of Uttar Dinajpur districts of West Bengal during June 2016 to September 2016. The study locations were Siliguri Municipal Corporation (SMC), Matigara, and Khoribari of the Darjeeling district; Malbazar, Dhupguri of the Jalpaiguri district, and the Itahar block of Uttar Dinajpur. Most of the study sites were sub-urban except Siliguri Municipal Corporation (urban) and Khoribari (rural) (Fig 1).
Fig 1. Map showing the study sites.
Mosquito collection, rearing and identification
The aquatic stages (larvae and pupae) of Aedes sp. were collected from the seventeen localities of three districts. For each collection site, larvae and pupae were collected from domestic, peri-domestic and natural breeding places. The collected immature stages of mosquitoes were stored in plastic containers containing water from the same breeding habitat and transferred to the laboratory. In the laboratory, the wild caught mosquito larvae and pupae were transferred into a larvae rearing tray along with water collected from the field and supplied with food for ornamental fishes available in the local market along with yeast. The mosquito larvae and pupae were reared to the adult stages in the laboratory under controlled conditions (temperature 25°C ± 2°C; relative humidity 80% ± 10%). After emergence, the adults were identified by using the standard identification keys of Barraud, 1934 [29] and Tyagi et al., 2012 [30]. The identified Ae. albopictus were allowed to breed under laboratory conditions. The larvae and adults of the F1 generation were used for larval and adult insecticide bioassays.
Larval susceptibility tests
Susceptibility of larvae to temephos (50EC; Nitapol Industries Pvt Ltd., Kolkata) was estimated using the standard WHO bioassay protocol [31]. The stock temephos solution of 1 ppm concentration and other subsequent dilutions were prepared in 95% ethanol and stored at +4°C for use in the susceptibility bioassay. Bioassays were conducted using 20–25 third instar to early fourth instar larvae (wild caught strain and laboratory strain) in disposable paper cups filled with the required concentration of insecticide solution and double distilled water at room temperature (25°C ± 2°C). Eight different concentrations (0.0005, 0.001, 0.005, 0.01, 0.05, 0.1, 0.5 and 1.0 ppm) were used as per WHO recommendation [32, 33] and each experiment was replicated at least three times. Each set of the bioassay was accompanied by two sets of controls (equal concentration of 95% ethanol). Larval mortality was recorded after 24 h of exposure. The larvae that were motionless or convulsive upon a sharp stimulation were counted as dead [31]. Larval mortality was determined by dividing the number of dead larvae by the total number tested. A test was considered as invalid if pupation rate was greater than 10%, or mortality rate in the control was greater than 20% [31]. The degree of resistance was determined by the resistance ratio (RR99), which is calculated by comparing the lethal concentration (LC99) value for a population with the LC99 value for the insecticide for a laboratory colony. The RR99 ≤3 was considered as susceptible, and 3 < RR99 ≤ 5 as low resistance, 5 < RR99 ≤ 10 as moderate resistance, and RR99> 10 as high resistance [34].
Adult susceptibility bioassay
Two to three day old laboratory emerged unfed female Ae. albopictus mosquitoes were used for the insecticide susceptibility bioassay as per WHO protocol [18]. The tested insecticides were 4% DDT, 0.05% deltamethrin, and 5% malathion. The insecticide-impregnated papers were procured from the Vector Control Research Unit (VCRU), Universiti Sains Malaysia, Malaysia. Five different holding tubes were used for each set of the experiment of which four were a test and one was a control. In each holding tube,15–20 adult female mosquitoes were kept for one hour. After one hour of holding, mosquitoes from four tubes marked as test were exposed to insecticide-impregnated papers. The control tests were performed using silicone oil, olive oil, and risella oil pre-impregnated papers for deltamethrin, malathion, and DDT, respectively. Mosquitoes were allowed in the exposure tube for one hour and cumulative knock down was recorded after 10, 15, 20, 30, 40, 50, and 60 minutes. After 60 minutes of exposure, the mosquitoes were transferred to holding tubes and fed on a 5% sucrose solution for the next 24 h. Mortality was scored after 24 h to determine the susceptibility status as per WHO recommendation [18]. Mosquitoes were considered dead if they were motionless, when they were mechanically stimulated, following the method of Gonzalez Audino [35]. The live and dead mosquitoes obtained from the adult bioassays were stored at -20°C and used for molecular biological assays.
DNA isolation and kdr mutation detection
Genomic DNA was isolated from both live and dead mosquitoes (individually) by using the DNeasy Blood & Tissue Kit (Qiagen, Germany), as per the manufacturer’s instructions. Before initiation of DNA isolation, the wings of the mosquitoes were removed and the remaining part of the mosquito was carefully homogenised by a Tissue Ruptor (Qiagen, Germany). Extracted DNA was stored at -20°C until further study.
PCR was done using three different primer pairs targeting six amino acid loci (S953P, I975M/V, L978, V980G of domain II, F1474C of domain III and D1703Y of domain IV) of the voltage-gated sodium channel gene (VGSC) of Ae. albopictus, which is responsible for knockdown resistance (kdr). The details of primers and PCR conditions are given in Table 1 as described earlier by Kasai et al., 2011 [36].
Table 1. Primer and PCR conditions used for amplification and sequencing of VGCS gene of Aedes albopictus (Kasai et al., 2011).
| Domains | Primer name | PCR Primers (5'-3') | PCR condition | Sequencing primers (5'-3') |
|---|---|---|---|---|
| II | aegSCF20 | GACAATGTGGATCGCTTCCC | Initial denaturation at 94°C for 3 min, 35 cycles each of 94°C for 15 s, 55°C for 30 s, and 72°C for 30 s, followed by a final elongation step at 72°C for 10 min | aegSCF3: GTGGAACTTCACCGACTTCA |
| aegSCR21 | GCAATCTGGCTTGTTAACTTG | aegSCR22: TTCACGAACTTGAGCGCGTTG | ||
| III | aegSCF7 | GAGAACTCGCCGATGAACTT | aegSCR8: TAGCTTTCAGCGGCTTCTTC | |
| aegSCR7 | GACGACGAAATCGAACAGGT | |||
| IV | albSCF6 | TCGAGAAGTACTTCGTGTCG | albSCF7: AGGTATCCGAACGTTGCTGT | |
| albSCR8 | AACAGCAGGATCATGCTCTG |
The quality of PCR products was ascertained by 2% agarose gel electrophoresis following ethidium bromide stain. The PCR product was gel purified using the Qiagen gel extraction kit (Qiagen, Germany) and sequencing was outsourced to Chromous Biotech, Bangalore. Four different primers i.e., aegSCF3, aegSCR22 (forward and reverse primer for domain II), aegSCR8 (reverse primer for domain III), and albSCF7 (forward primer for domain IV) were used for sequencing of the PCR products.
Analysis of sequence
In the present study, we numbered the codons of the VGSC gene according to the sequence of Ae. albopictus. The sequences were analysed using the software BioEdit Sequence Alignment Editor version 7.0.9.0. The sequences were aligned with the reference sequence for Ae. albopictus (GenBank accession no. AY663384.1), using an online multiple sequence alignment tool.
Ethics statement
Before initiation of the work, the objectives of the study were explained to the local population of each study site. Permission was taken from the owners of private houses/lands before collection of immature stages of mosquito. The study did not involve any endangered and protected species. Mosquitoes were maintained under optimal conditions such as temperature, humidity, and adequate food supply in the laboratory. The study protocol was approved by the Institutional Ethics Committee of Calcutta School of Tropical Medicine, Kolkata.
Data analysis
Larval bioassay data were analyzed using Log dose probit (Ldp) Line computer software (Ehabsoft, Cairo Egypt; available at: http://www.ehabsoft.com/ldpline) according to the Finney’s method [37]. Chi-squared (χ2) test was used to estimate the goodness of fit, while linear regression was used to evaluate the data linearity. Lethal concentrations (LC10, LC50, and LC99) along with the slope were estimated at 95% confidence intervals (CI). For adult bioassays, observed mortality was calculated by the formula: observed mortality (%) = (Total no. of dead mosquitoes / Total mosquitoes exposed) x 100. The observed mortality was corrected using Abbott’s formula when the mortality rate of control was within 5% - 20%. Corrected Mortality (CM) (%) = [(% of observed mortality—% of control mortality) / (100 - % of control mortality)] x 100. For adult bioassays, resistant/susceptibility status was defined according to WHO recommendations [18]. Mosquitoes were considered susceptible (S) if the corrected mortality (CM) rate was greater than 98% and resistant (R) if mortality rate was less than 90%. Mortality rate between 90–98% was considered as possible resistance (PR) and needs verification by alternative methods like enzyme bioassay and molecular marker studies [18]. The cumulative knock down rates (KDR) were calculated by observing the number of knocked down mosquitoes after 10, 15, 20, 30, 40, 50 and 60 minutes during the hour-long exposure period. Knockdown time (KDT10, KDT50, and KDT95) is the time required for knockdown of a particular proportion of mosquitoes following exposure to any insecticide. KDTs were determined using Log dose probit (Ldp) Line computer software (Ehabsoft, Cairo Egypt; available at: http://www.ehabsoft.com/ldpline) programme according to the Finney’s method [37].
Results
Demography of the study area
The study was conducted in one municipality and 5 different blocks of 3 districts in the northern part of West Bengal during June 2016 –September 2016. The study sites of Dhupguri and Itahar blocks were surrounded by paddy fields, whereas the presence of both paddy fields and tea gardens were characteristic of the remaining study sites except Siliguri Municipality Corporation (SMC) and Matigara. Most of the study sites were suburban in nature except the Siliguri municipality area (Urban) and Khoribari (rural) (Fig 1). Storage water tanks, discarded tyres, tree holes, construction sites, flower pots, plastic cups, coconut shells, and discarded containers were the different seasonal breeding sites found in the study area. The climatic conditions of all study sites were humid and sub-tropical in nature and the temperature varies from 8°C in winter to 40°C in summer.
Larval susceptibility status
The summary of larval bioassay results is presented in Table 2. The LC10, LC50, and LC99 values of different study sites did not follow a normal distribution for mortality to the log dose (χ2 ≥ 16.08; p ≤ 0.01). The LC50 values ranged from 0.0009 to 0.0015 mg/L and LC99 from 0.1565 to 0.3343 mg/L. The calculated RR50 and RR99 values in different study sites were ranged from 1.0 to 2.5 and 2.45 to 5.24, respectively.
Table 2. Temephos susceptibility status of Aedes albopictus in three districts of West Bengal.
| Values | Study sites | |||||
|---|---|---|---|---|---|---|
| Darjeeling | Jalpaiguri | Uttar Dinajpur | ||||
| Siliguri (n = 480) |
Matigara (n = 480) |
Khoribari (n = 480) |
Dhupguri (n = 480) |
Malbazar (n = 480) |
Itahar (n = 480) |
|
| LC10 (lower limit–upper limit) [mg/L] | 0.0001 (0–0.0001) |
0.0001 (0–0.0001) |
0.0001 (0–0.0001) |
0.0001 (0–0.0001) |
0.0001 (0–0.0001) |
0.0001 (0–0.0001) |
| LC50 (lower limit–upper limit) [mg/L] | 0.0015 (0.0005–0.0028) |
0.0009 (0.0002–0.0018) |
0.001 (0.0001–0.0019) |
0.0006 (0.0001–0.0011) |
0.001 (0.0002–0.0021) |
0.0009 (0.0002–0.0019) |
| LC99 (lower limit–upper limit) [mg/L] | 0.3343 (0.1699–2.8605) |
0.2574 (0.1616–3.7848) |
0.1678 (0.2453–19.1891) |
0.1565 (0.1838–16.7816) | 0.1963 (0.1414–4.5084) |
0.2043 (0.1763–8.025) |
| X2 (p) | 16.08 (0.01) | 23.97 (0.0005) | 34.72 (<0.0001) | 27.93 (0.0001) | 23.59 (0.0006) | 26.68 (0.0002) |
| Slope | 0.99 ± 0.09 | 0.95 ±0.09 | 1.03 ± 0.11 | 0.97 ± 0.11 | 1.02 ± 0.11 | 0.99 ± 0.11 |
| R | 0.95 | 0.92 | 0.91 | 0.91 | 0.93 | 0.92 |
| G | 0.14 | 0.21 | 0.39 | 0.38 | 0.26 | 0.29 |
| RR50/RR99* | 2.5 / 5.24 | 1.5 / 4.03 | 1.67 / 2.63 | 1.0 / 2.45 | 1.67 / 3.08 | 1.5 / 3.20 |
| Status# | MR | LR | S | S | LR | LR |
n = number; LC10/LC50/LC99 = lethal concentration 10%/50%/99%, RR = resistance ratio, g = ‘g’ is a factor used for fiducial limit calculations
* The LC50 and LC99 values of laboratory strain was 0.0006mg/L and 0.0638mg/L, respectively
#Classification adapted from Mazzari and Georghiou (1995): S = Susceptible (RR < 3), LR = Low Resistance (3 < RR < 5), MR = Moderate Resistance (5 < RR < 10), HR = High Resistance (>10).
Adult susceptibility status
The results of the adult susceptibility bioassay for Ae. albopictus are given in Table 3. After 24 hours of exposure, the corrected mortality rates for 4% DDT were 23.75% to 85.53% in different study sites. The obtained mortality rates were well below the WHO recommended 90% mortality rate for resistance. So, results suggested that the Ae. albopictus population of the study areas was highly resistant to DDT. In all of the study sites, the corrected mortality rate for 0.05% deltamethrin ranged from 98.08% to 100%, except in Dhupguri where the corrected mortality was 96.25%. So, Ae. albopictus population of all the study sites was susceptible to deltamethrin except Dhupguri. The corrected mortality rate for 5% malathion was >98% in all the study sites indicating susceptibility to malathion.
Table 3. Insecticide susceptibility status of Aedes albopictus against 4% DDT, 0.05% deltamethrin, and 5% malathion in West Bengal.
| Insecticides | Districts | Blocks | Mosquito exposed | Mosquito died | Observed Mortality (%) | CM (%) | KDT10 (min.) [95% CI] |
KDT50 (min.) [95% CI] |
KDT95 (min.) [95% CI] |
χ2 (p) | Slope | Status# | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T* | C* | T* | C* | T* | C* | ||||||||||
| 4% DDT | Darjeeling | Siliguri | 160 | 40 | 136 | 0 | 85.00 | 0 | 85.00 | 12.82 [9.05–14.75] | 28.59 [23.6–34.39] | 80.10 [69.57–119.04] | 19.59 (0.002) | 3.68 (± 0.2) | R |
| Matigara | 160 | 40 | 96 | 2 | 60.00 | 5.00 | 57.89 | 11.98 [9.45–14.24] | 42.17 [37.90–47.91] | 212.11 [155.71–329.21] | 3.24 (0.7) | 2.34 (± 0.2) | R | ||
| Khoribari | 160 | 40 | 132 | 0 | 82.50 | 0 | 82.50 | 13.26 [11.47–14.89] | 30.36 [28.31–32.57] | 87.91 [76.28–105.28] | 7.53 (0.2) | 3.56 (± 0.2) | R | ||
| Jalpaiguri | Dhupguri | 160 | 40 | 122 | 2 | 76.25 | 5.00 | 75.00 | 17.39 [11.08–20.41] | 51.39 [43.11–73.09] | 206.49 [172.26–538.03] | 15.83 (0.007) | 2.72 (± 0.3) | R | |
| Malbazar | 160 | 40 | 138 | 2 | 86.25 | 5.00 | 85.53 | 7.65 [5.97–9.22] | 23.62 [21.48–25.83] | 100.42 [82.50–130.47] | 2.91 (0.7) | 2.62 (± 0.2) | R | ||
| U. Dinajpur | Itahar | 160 | 40 | 38 | 0 | 23.75 | 0 | 23.75 | 11.02 [9.04–12.82] | 31.57 [29.03–34.46] | 121.92 [99.52–159.70] | 6.06 (0.3) | 2.80 (± 0.2) | R | |
| 0.05% deltamethrin | Darjeeling | Siliguri | 160 | 40 | 157 | 1 | 98.13 | 2.50 | 98.08 | 4.48 [2.38–5.56] | 12.09 [8.84–14.35] | 43.28 [36.38–63.51] | 12.93 (0.02) | 2.97 (± 0.3) | S |
| Matigara | 160 | 40 | 158 | 0 | 98.75 | 0 | 98.75 | 6.45 [5.26–7.46] | 11.60 [10.57–12.51] | 24.63 [22.25–28.27] | 0.95 (0.9) | 5.03 (± 0.5) | S | ||
| Khoribari | 160 | 40 | 160 | 0 | 100.00 | 0 | 100 | 6.69 [5.63–7.67] | 13.07 [11.97–14.09] | 30.84 [28.20–34.41] | 0.94 (0.9) | 4.41 (± 0.3) | S | ||
| Jalpaiguri | Dhupguri | 160 | 40 | 154 | 2 | 96.25 | 5.00 | 96.25 | 7.29 [6.24–8.24] | 13.82 [12.76–14.82] | 31.43 [28.84–34.87] | 1.99 (0.8) | 4.61 (± 0.3) | PR | |
| Malbazar | 160 | 40 | 158 | 2 | 98.75 | 5.00 | 98.68 | 5.05 [3.84–6.27] | 11.86 [10.47–13.13] | 35.49 [31.37–41.68] | 6.4 (0.3) | 3.46 (± 0.2) | S | ||
| U. Dinajpur | Itahar | 160 | 40 | 160 | 0 | 100.00 | 0 | 100 | 5.38 [4.09–6.47] | 10.14 [8.94–11.14] | 22.85 [20.52–26.54] | 1.09 (0.9) | 4.67 (± 0.5) | S | |
| 5% malathion | Darjeeling | Siliguri | 160 | 40 | 160 | 0 | 100.00 | 0 | 100 | 10.85 [8.02–12.03] | 17.52 [14.65–20.53] | 32.39 [29.26–44.23] | 24.35 (0.0002) | 6.16 (± 0.4) | S |
| Matigara | 160 | 36 | 158 | 2 | 98.75 | 5.56 | 98.68 | 10.85 [8.15–12.58] | 23.79 [20.27–27.49] | 65.17 [56.16–87.18] | 13.64 (0.01) | 3.76 (± 0.2) | S | ||
| Khoribari | 160 | 40 | 158 | 0 | 98.75 | 0 | 98.75 | 9.31 [7.76–10.76] | 23.49 [21.67–25.36] | 77.04 [66.47–92.98] | 9.73 (0.08) | 3.19 (± 0.2) | S | ||
| Jalpaiguri | Dhupguri | 160 | 40 | 160 | 2 | 100.00 | 5.00 | 100 | 13.70 [10.09–15.51] | 25.31 [20.97–29.99] | 55.59 [49.19–76.13] | 24.83 (0.0002) | 4.81 (± 0.3) | S | |
| Malbazar | 160 | 35 | 158 | 1 | 98.75 | 2.86 | 98.71 | 9.75 [8.40–10.98] | 20.90 [19.44–22.37] | 55.64 [49.69–63.99] | 6.31 (0.3) | 3.87 (± 0.2) | S | ||
| U. Dinajpur | Itahar | 160 | 40 | 156 | 0 | 97.50 | 0 | 97.50 | 12.71 [6.77–13.47] | 21.68 [15.34–28.79] | 43.08 [40.94–81.72] | 65.38 (<0.0001) | 5.52 (± 0.3) | S | |
*T = Test, C = Control, CM = Corrected Mortality
#S = Susceptible (CM ≥98%); R = Confirmed Resistance (CM <90%); PR = Possible Resistance (CM = 90–97%)
The knock down time (KDT10, KDT50, KDT95) for DDT, deltamethrin, and malathion showed a linear probit for knock-down rates with time in most of the study sites (Table 3). The observed KDT50 values were 23.62 to 51.39 mins for DDT, 10.14 to 13.82 mins for deltamethrin, and 17.52 to 25.31 mins for malathion. The KDT95 values for DDT were 80.10 to 212.11 mins, for deltamethrin 22.85 to 43.28 mins and for malathion 32.39 to 77.04 mins. The survival rate of Ae. albopictus against DDT, deltamethrin, and malathion over an exposure time of 1 hour is given in Fig 2A–2C. During 1 hour of exposure, the knock down rate (KDR) varies from 68.75% - 93.75% for DDT, 100% for deltamethrin, and 95.00% - 100% for malathion.
Fig 2.

Survival rate of Aedes albopictus against 4% DDT (A), 0.05% deltamethrin (B), 5% malathion (C) in West Bengal.
Detection of kdr mutations in Ae. albopictus
DNA was isolated from 30 dead and 10 alive, deltamethrin-exposed Ae. albopictus mosquitoes and used for PCR amplification. For detection of kdr mutations three DNA fragments of 480 bp, 740 bp, and 280 bp for domain II, III, and IV of VGSC gene were amplified, respectively. None of the six important kdr mutations (i.e., S953P, I975M/V, L978, V980G, F1474C, D1703Y) were found among the studied mosquitoes. We detected 3 synonymous mutations in domain II, 1 non-synonymous and 3 synonymous mutations in domain III, and 5 synonymous mutations in domain IV. The frequencies of observed mutations are presented in Table 4. The DNA sequences have been submitted to GenBank under accession nos.MF776970 and MF774494.
Table 4. Prevalence of SNPs in VGSC gene of Aedes albopictus in West Bengal.
| Domains | SNPs | Occurrence of mutations | |||
|---|---|---|---|---|---|
| Amino acids | Codon change | N | % | 95% CI | |
| II | V942V | GTG>GTA | 6 | 15.0% | 7.06–29.07 |
| L946L | CTG>TTG | 5 | 12.5% | 5.46–26.11 | |
| C947C | TGT>TGC | 40 | 100% | 91.24–100.00 | |
| III | D1445D | GAT>GAC | 6 | 15.0% | 7.06–29.07 |
| G1453G | GGC>GGA | 5 | 12.5% | 5.46–26.11 | |
| F1468F | TTC>TTT | 7 | 17.5% | 8.75–31.95 | |
| S1485L | TCG>TTG | 3 | 7.5% | 2.58–19.86 | |
| IV | A1691A | GCT>GCC | 4 | 10.0% | 3.96–23.05 |
| G1694G | GGG>GGC | 3 | 7.5% | 2.58–19.86 | |
| D1709D | GAC>GAT | 5 | 12.5% | 5.46–26.11 | |
| N1712N | AAT>AAC | 6 | 15.0% | 7.06–29.07 | |
| F1713F | TTT>TTC | 8 | 20.0% | 10.5–34.76 | |
Discussion
Emergence and spread of insecticide resistance is the biggest challenge to control vector-borne disease transmission [38]. In Aedes mosquitoes there are two major mechanisms for pyrethroid resistance: increased detoxification and mutation in the VGSC gene. To date more than 50 different VGSC mutations have been identified in different insect species [19]. The six non-synonymous amino acid substitutions: S989P, I1011M, L1014F, V1016G in domain II, F1534C in domain III and D1763Y in domain IV of house fly are found to be associated with pyrethroid resistance. These codons are orthologous to the codons 953, 975, 978, 980, 1474 and 1703, respectively of Ae. albopictus. The involvement of other mutations in pyrethroid resistance remains to be investigated. The L1014F, at S6 subunit of domain II was the first pyrethroid-resistance-associated mutation identified in the house fly and German cockroach [39, 40, 41]. I1011M was identified in domestic house fly from Brazil, Guyana, whereas V1016G was identified from Indonesia and Thailand [42]. Later, different substitutions, I1011V and V1016I, were found in Ae. aegypti populations from Latin America [43]. The most significant F1534C, located in S6 subunit of domain III was discovered in DDT/permethrin-resistant Ae. aegypti in Thailand and Vietnam [44, 45]. The adult insecticide susceptibility bioassay is applied to determine the lethal dose of different insecticides by direct exposure. Additional tests, such as polymorphisms in marker genes and biochemical assays of different enzymes are used as supplementary evidence to clarify the results of bioassays and potential mechanisms.
In the present study, we determined the susceptibility status of Ae. albopictus against DDT, deltamethrin, and malathion. The results showed that Ae. albopictus is significantly resistant to DDT with a higher KDT and KDR and a lower mortality rate. Similar observations have also been reported from other parts of the country [11, 12, 14]. Though DDT is not in use against Aedes vector mosquitoes, this compound is still in use for control of malaria vectors. The present study areas have been highly endemic for malaria for a long time, with the exception of Itahar. Thus, Aedes mosquitoes have been exposed to DDT for many generations which might be the cause of the high level of resistance that has developed to DDT. Ae. albopictus from the present study areas were susceptible to deltamethrin and malathion. Pyrethroid resistance in adult Aedes sp. is a problem worldwide. The level of resistance varies from region to region. A lower level of resistance is found in Asian, African, and Northern American countries, [46, 47, 48, 49] whereas higher levels of resistance are found in South American countries [50, 51]. In the present study, lower values of knock down time and knock down rate were observed in Ae. albopictus against deltamethrin and malathion. The KDT values recorded in the present study did not follow a normal distribution pattern which indicates that the prevailing Ae. albopictus population is susceptible to these insecticides.
In India, temephos is used as larvicidal agent. In contrast to adult susceptibility, higher levels of larval resistance have been found in Asian, African, and North American countries [49, 52, 53, 54, 55]. In the present study we found that the Ae. albopictus larvae were sensitive to temephos in Khoribari and Dhupguri (RR99<3); showed a low level of resistance in Matigara, Malbazar, and Itahar (3<RR99>5), and moderate resistance in Siliguri (5<RR99>10) [33].A similar type of observation was also reported from the north eastern part of India [11, 12]. The Siliguri Municipal Corporation is the only urban site in the present study, where temephos has been in use for a long time. A longer duration of exposure to temephos might be the cause of the observed moderate level of resistance against it in Ae. albopictus from Siliguri. In contrast, a recent report from the northern part of West Bengal showed susceptibility of Ae. albopictus larvae to temephos assessed by larval susceptibility and bioassay of detoxifying enzymes [56].
The KDR is a mechanism of DDT and pyrethroid resistance. Mutations at codons 953, 975, 978, 980, 1474, 1703 of the VGSC gene of Ae. albopictus have been found to be associated with reduced susceptibility to both DDT and pyrethroids [22, 23]. As per our present study, the only previous report from India did not reveal any mutation in the VGSC gene of Ae. albopictus [8] but two other reports reveal mutations at codon F1534C [57] and at codon T1520I + F1534C of the Ae. aegypti VGSC gene [13]. In the present study, we detected only one non-synonymous mutation at S1485L in three samples. Interestingly, all three mosquitoes were susceptible to deltamethrin. So, the role of this mutation in pyrethroid resistance cannot be explained. We also detected 11 synonymous mutations among both dead as well as live deltamethrin-exposed mosquitoes.
We did not assess the detoxifying enzyme levels associated with DDT and deltamethrin resistance. The higher susceptibility level in deltamethrin with absence of important kdr mutations and higher susceptibility to malathion indicate that these two insecticides are still effective in the study areas. The susceptibility status of temephos as a larvicide should be monitored closely as moderate and lower levels of resistance were observed in mosquitoes from a few study sites. A similar study is highly recommended for monitoring and early detection of pyrethroid and malathion resistance in other parts of the country.
Acknowledgments
We thank all the population of the study villages for their help during sample collection. We are also thankful to the Chief Medical Officer of Health and Deputy Chief Medical Officer of Health–II of Darjeeling district for their kind cooperation during the mosquito collection.
Data Availability
All relevant data are available within the paper.
Funding Statement
The authors did not received any specific funding for the present work, but received partial financial assistance from Department of Science and Technology, Govt. of West Bengal. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Ng LC. Challenges in dengue surveillance and control. Western Pac Surveill Response. 2011; J 2:1–3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bhatt S, Gething PW, Brady OJ, Messina JP, Farlow AW, Moyes CL, et al. The global distribution and burden of dengue. Nature. 2013; 496: 504–507. doi: 10.1038/nature12060 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Brady OJ, Gething PW, Bhatt S, Messina JP, Brownstein JS, Hoen AG, et al. Refining the global spatial limits of dengue virus transmission by evidence-based consensus. PLoS Negl Trop Dis. 2012; 6:e1760 doi: 10.1371/journal.pntd.0001760 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.NVBDCP, 2017. Dengue/DHF situation in India. New Delhi, NVBDCP. (available at: http://nvbdcp.gov.in/den-cd.html).
- 5.Kerah-Hinzoumbé C, Péka M, Nwane P, Donan-Gouni I, Etang J, Samè-Ekobo A. et al. Insecticide resistance in Anopheles gambiae from south-western Chad, Central Africa. Malar J.2008; 7: 192 doi: 10.1186/1475-2875-7-192 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Berg HVD, Zaim M, Yadav R, Soares A, Ameneshewa B, Mnzava A, et al. Global trends in the use of insecticides to control vector-borne diseases. Environ. Health Persp. 2012; 120: 577–582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.David JP, Ismail H, Chandor-Proust A, Paine M. Role of cytochrome P450s in insecticide resistance: impact on the control of mosquito-borne diseases and use of insecticides on Earth, Phil. Trans. Royal Soc. London B Biol. Sci., 2013; 368 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.WHO, 2009. Dengue guidelines for diagnosis, treatment, prevention and control, World Health Organization (WHO) and the Special Programme for Research and Tropical Diseases (TDR).
- 9.Baldacchino F, Caputo B, Chandre F, Drago A, Torre A, Montarsi F, et al. Control methods against invasive Aedes mosquitoes in Europe: a review. Pest Man. Sci. 2015; 71: 1471–1485. [DOI] [PubMed] [Google Scholar]
- 10.Carter SW. A review of the use of synthetic pyrethroids in public health and vector pest control. Pestic. Sci. 1989: 27. [Google Scholar]
- 11.Dhiman S, Rabha B, Yadav K, Baruah I, Veer V. Insecticide susceptibility and dengue vector status of wild Stegomyia albopicta in a strategically important area of Assam, India. Parasites & Vectors. 2014; 7: 295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Yadav K, Rabha B, Dhiman S, Veer V. Multi-insecticide susceptibility evaluation of dengue vectors Stegomyia albopicta and St. aegypti in Assam, India. Parasites & Vectors. 2015; 8: 143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kushwah RBS, Dykes CL, Kapoor N, Adak T, Singh OP. Pyrethroid-resistance and presence of two knockdown resistance (kdr) mutations, F1534C and a Novel Mutation T1520I, in Indian Aedes aegypti. PLoS Negl Trop Dis. 2015; 9(1): e3332 doi: 10.1371/journal.pntd.0003332 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kushwah RBS, Mallick PK, Ravikumar H, Dev V, Kapoor N, Adak T, et al. Status of DDT and pyrethroid resistance in Indian Aedes albopictus and absence of knockdown resistance (kdr) mutation. J Vector Borne Dis. 2015; 52: 95–98. [PubMed] [Google Scholar]
- 15.Das M, Dutta P. Status of insecticide resistance and detoxifying enzyme activity of Aedes albopictus population in Sonitpur district of Assam, India. International Journal of Mosquito Research. 2014; 1(4): 35–41. [Google Scholar]
- 16.Elliott M. Synthetic Pyrethroids American Chemical Society; Washington, DC, USA: 1977; 1–28. [Google Scholar]
- 17.Ranson H, Lissenden N. Insecticide resistance in african Anopheles mosquitoes: A worsening situation that needs urgent action to maintain malaria control. Trends Parasitol. 2016; 32: 187–196. doi: 10.1016/j.pt.2015.11.010 [DOI] [PubMed] [Google Scholar]
- 18.WHO, 2016. Test procedures for insecticide resistance monitoring in malaria vector mosquitoes– 2nd ed. World Health Organization, 20 Avenue Appia, Geneva, Switzerland. http://www.who.int/malaria/areas/vector_control/insecticide_resistance/en/
- 19.Dong K, Du Y, Rinkevich F, Nomura Y, Xu P, Wang L, et al. Molecular biology of insect sodium channels and pyrethroid resistance. Insect Biochem. Mol. Biol. 2014; 50: 1–17. doi: 10.1016/j.ibmb.2014.03.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Narahashi T, Ginsburg KS, Nagata K, Song JH, Tatebayashi H. Ion channels as targets for insecticides. Neurotoxicology. 1998; 19: 581–590. [PubMed] [Google Scholar]
- 21.Silver KS, Du Y, Nomura Y, Oliveira EE, Salgado VL, Zhorov BS, et al. Voltage-Gated Sodium Channels as Insecticide Target Sites. Elsevier; Burlington, MA, USA: 2014; 46: 389–433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Soderlund DM. Sodium channels In: Gilbert L.I., Iatrou K., Gill S.S., editors. Comprehensive Molecular Insect Science. Volume 5 Elsevier; Amsterdam, The Netherlands: 2005. pp. 1–24. [Google Scholar]
- 23.Soderlund DM, Bloomquist JR. Neurotoxic actions of pyrethroid insecticides. Ann. Rev. Entomol. 1989; 34: 77–96. [DOI] [PubMed] [Google Scholar]
- 24.Catterall WA, Chandy KG, Clapham DE, Gutman GA, Hofmann F, Harmar AJ, et al. International Union of Pharmacology: Approaches to the nomenclature of voltage-gated ion channels. Pharmac. Rev. 2003; 55(4):573–574. [Google Scholar]
- 25.Davies TGE, Field LM, Usherwood PN, Williamson MS. A comparative study of voltage-gated sodium channels in the Insecta: implications for pyrethroid resistance in Anopheline and other Neopteran species. Insect. Mol. Biol. 2007; 16(3):361–375. doi: 10.1111/j.1365-2583.2007.00733.x [DOI] [PubMed] [Google Scholar]
- 26.Du Y, Nomura Y, Satar G, Hu Z, Nauen R, He SY, et al. Molecular evidence for dual pyrethroid-receptor sites on a mosquito sodium channel. Proc. Natl. Acad. Sci. USA. 2013; 110: 11785–11790 doi: 10.1073/pnas.1305118110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Moyes CL, Vontas J, Martins AJ, Ng LC, Koou SY, Dusfour I, et al. Contemporary status of insecticide resistance in the major Aedes vectors of arboviruses infecting humans. PLoS Negl Trop Dis. 2017; 11(7): e0005625 doi: 10.1371/journal.pntd.0005625 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Smith LB, Kasai S and Scott JG. Pyrethroid resistance in Aedes aegypti and Aedes albopictus: Important mosquito vectors of human diseases. Pesticide Biochemistry and Physiology. 2016; 133:1–12. doi: 10.1016/j.pestbp.2016.03.005 [DOI] [PubMed] [Google Scholar]
- 29.Barraud PJ. The fauna of British India including Burma and Ceylon (Diptera: Culicidae), Tribes Megarhinini and Culicini, Vol 5, London: Taylor and Francis, 1934; 1–452. [Google Scholar]
- 30.Tyagi BK, Munirathinam A, Krishnamoorthi R, Venkatesh A. A field based hand book of identification keys to mosquitoes of public health importance in India. Centre for research in Medical Entomology, Madurai, India. 2012; 25–27. [Google Scholar]
- 31.World Health Organization (WHO). 2005. Guidelines for laboratory and field testing of mosquito larvicides. WHO/CDS/WHOPES/GCDPP.13, Geneva, Switzerland: World Health Organization; [Google Scholar]
- 32.World Health Organization (WHO). 1981(a).Criteria and meaning of tests for determining the susceptibility or resistance of insects to insecticides. WHO/VBC/81.806, Geneva.
- 33.World Health Organization (WHO). 1981(b). Instructions for determining the susceptibility or resistance of adult mosquitoes to organochlorine, organophosphate and carbamate insecticides. Establishment of the baseline. WHO/VBC/81.805, Geneva,.
- 34.Mazzarri MB, Georghiou GP. Characterization of resistance to organophosphate, carbamate, and pyrethroid insecticides in field populations of Aedes aegypti from Venezuela. J. Am. Mosq. Control. Assoc. 1995; 11:315–322 [PubMed] [Google Scholar]
- 35.Gonzalez Audino P, Vassena C, Barrios S, Zerba E, Picollo MI. Role of enhanced detoxication in a deltamethrin-resistant population of Triatoma infestans (Hemiptera, Reduviidae) from Argentina. Mem Inst Oswaldo Cruz. 2004; 99(3): 335–339. [DOI] [PubMed] [Google Scholar]
- 36.Kasai S, Ng LC, Lam-Phua SG, Tang CS, Itokawa K, Komagata O, et al. First detection of a putative knockdown resistance gene in major mosquito vector, Aedes albopictus. Jpn J Infect Dis. 2011; 64: 217–221. [PubMed] [Google Scholar]
- 37.Finney J. 1972. Probit analysis. 3rd En London, Cambridge University Press; 68–78. [Google Scholar]
- 38.World Health Organization (WHO). 1976. WHO, 22nd Report by the Expert Committee on Insecticides, WHO Tech. Rpt. Ser., pp. 77.
- 39.Miyazaki M, Ohyama K, Dunlap DY, Matsumura F. Cloning and sequencing of the para-type sodium channel gene from susceptible and kdr-resistant german cockroaches (Blattella germanica) and house fly (Musca domestica) Mol. Gen. Genet. 1996; 252: 61–68. [PubMed] [Google Scholar]
- 40.Williamson MS, Martinez-Torres D, Hick CA, Devonshire AL. Identification of mutations in the housefly para-type sodium channel gene associated with knockdown resistance (kdr) to pyrethroid insecticides. Mol. Gen. Genet. 1996; 252: 51–60. [DOI] [PubMed] [Google Scholar]
- 41.Dong K. A single amino acid change in the para sodium channel protein is associated with knockdown-resistance (kdr) to pyrethroid insecticides in german cockroach. Insect Biochem. Mol. Biol. 1997; 27: 93–100. [DOI] [PubMed] [Google Scholar]
- 42.Brengues C, Hawkes NJ, Chandre F, McCarroll L, Duchon S, Guillet P, et al. Pyrethroid and DDT cross-resistance in Aedes aegypti is correlated with novel mutations in the voltage-gated sodium channel gene. Med. Vet. Entomol. 2003; 17: 87–94. [DOI] [PubMed] [Google Scholar]
- 43.Saavedra-Rodriguez K, Urdaneta-Marquez L, Rajatileka S, Moulton M, Flores AE, Fernandez-Salas I, et al. A mutation in the voltage-gated sodium channel gene associated with pyrethroid resistance in Latin American Aedes aegypti. Insect Mol. Biol. 2007; 16: 785–798. doi: 10.1111/j.1365-2583.2007.00774.x [DOI] [PubMed] [Google Scholar]
- 44.Kawada H, Higa Y, Komagata O, Kasai S, Tomita T, Thi Yen N, et al. Widespread distribution of a newly found point mutation in voltage-gated sodium channel in pyrethroid-resistant Aedes aegypti populations in Vietnam. PLoS Negl. Trop. Dis. 2009; 3: e527 doi: 10.1371/journal.pntd.0000527 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Yanola J, Somboon P, Walton C, Nachaiwieng W, Somwang P, Prapanthadara LA. High-throughput assays for detection of the F1534C mutation in the voltage-gated sodium channel gene in permethrin-resistant Aedes aegypti and the distribution of this mutation throughout Thailand. Trop. Med. Int. Health. 2011; 16: 501–509. doi: 10.1111/j.1365-3156.2011.02725.x [DOI] [PubMed] [Google Scholar]
- 46.World Health Organization (WHO). 2006. WHO, Pesticides and Their Application for the Control of Vectors and Pests of Public Health Importance, in: DCNTD (Ed.).
- 47.Sivan A, Shriram AN, Sunish IP, Vulhya PT. Studies on insecticide susceptibility of Aedes aegypti (Linn) and Aedes albopictus (Skuse) vectors of dengue and chikungunya in Andaman and Nicobar Islands, India, Parasit. Res. 2015; 114: 4693–4702. [DOI] [PubMed] [Google Scholar]
- 48.Ishak IH, Jaal Z, Ranson H, Wondji C. Contrasting patterns of insecticide resistance and knockdown resistance (kdr) in the dengue vectors Aedes aegypti and Aedes albopictus from Malaysia. Parasit Vectors. 2015; 8: 181 doi: 10.1186/s13071-015-0797-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Wan-Norafikah O, Nazni WA, Lee HL, Zainol-Ariffin P, Sofian-Azirun M. Permethrin resistance in Aedes aegypti (Linnaeus) collected from Kuala Lumpur, Malaysia, J. Asia-Pacific Entomol. 2010; 13: 175–182. [Google Scholar]
- 50.Marcombe S, Darriet F, Tolosa M, Agnew P, Duchon S, Etienne M, et al. Pyrethroid resistance reduces the efficacy of space sprays for dengue control on the island of Martinique (Caribbean), PLoS Negl Trop Dis. 2011; 5: e1202 doi: 10.1371/journal.pntd.0001202 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Dusfour I, Thalmensy V, Gaborit V, Issaly J, Carinci R, Girod R. Multiple insecticide resistance in Aedes aegypti (Diptera: Culicidae) populations compromises the effectiveness of dengue vector control in French Guiana, Memorias do Instituto Oswaldo Cruz. 2011; 106: 346–352. [DOI] [PubMed] [Google Scholar]
- 52.Li CX, Kaufman PE, Xue RD, Zhao MH, Wang G, Yan T, et al. Relationship between insecticide resistance and kdr mutations in the dengue vector Aedes aegypti in Southern China, Parasite Vector. 2015; 8: 325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Shetty V, Sanil D, Shetty NJ. Insecticide susceptibility status in three medically important species of mosquitoes, Anopheles stephensi, Aedes aegypti and Culex quinquefasciatus, from Bruhat Bengaluru Mahanagara Palike, Karnataka, India, Pest Manag Sci. 2013; 69: 257–267. doi: 10.1002/ps.3383 [DOI] [PubMed] [Google Scholar]
- 54.Ponlawat A, Scott JG, Harrington LC. Insecticide susceptibility of Aedes aegypti and Aedes albopictus across Thailand, J Med Entomol. 2005; 42: 821–825. doi: 10.1603/0022-2585(2005)042[0821:ISOAAA]2.0.CO;2 [DOI] [PubMed] [Google Scholar]
- 55.Luna JED, Martins MF, Anjos AF, Kuwabara EF, Navarro-Silva MA. Susceptibility of Aedes aegypti to temephos and cypermethrin insecticides, Brazil. Rev. Saúde Pública. 2004; 38: 842–843. [DOI] [PubMed] [Google Scholar]
- 56.Bharati M, Saha D. Insecticide susceptibility status and major detoxifying enzymes activities in Aedes albopictus (Skuse), vector of dengue and chikungunya in Northern part of West Bengal, India. Accepted manuscript. http://dx.doi.org/doi:10.1016/j.actatropica.2017.02.029 [DOI] [PubMed]
- 57.Muthusamy R, Shivakumar MS. Involvement of metabolic resistance and F1534C kdr mutation in the pyrethroid resistance mechanisms of Aedes aegypti in India. Acta Tropica. 2015; 148: 137–141. doi: 10.1016/j.actatropica.2015.04.026 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are available within the paper.