

Abstract
The standard-of-care therapeutics for the treatment of ocular neovascular diseases like wet age-related macular degeneration (AMD) are biologics targeting vascular endothelial growth factor signaling. There are currently no FDA approved small molecules for treating these blinding eye diseases. Therefore, therapeutic agents with novel mechanisms are critical to complement or combine with existing approaches. Here, we identified soluble epoxide hydrolase (sEH), a key enzyme for epoxy fatty acid metabolism, as a target of an antiangiogenic homoisoflavonoid, SH-11037. SH-11037 inhibits sEH in vitro and in vivo and docks to the substrate binding cleft in the sEH hydrolase domain. sEH levels and activity are upregulated in the eyes of a choroidal neovascularization (CNV) mouse model. sEH is overexpressed in human wet AMD eyes, suggesting that sEH is relevant to neovascularization. Known sEH inhibitors delivered intraocularly suppressed CNV. Thus, by dissecting a bioactive compound’s mechanism, we identified a new chemotype for sEH inhibition and characterized sEH as a target for blocking the CNV that underlies wet AMD.
TABLE OF CONTENTS GRAPHIC

Ocular neovascularization is the underlying cause of blindness in diseases such as retinopathy of prematurity (ROP), proliferative diabetic retinopathy (PDR), and wet age-related macular degeneration (AMD), which cause blindness in infants, adults of working age and the elderly, respectively.1 Current FDA approved treatments for wet AMD focus on inhibiting the vascular endothelial growth factor (VEGF) signaling pathway using biologics such as ranibizumab and aflibercept.2 Despite the success of these therapeutic agents, their association with ocular and systemic side effects due to inhibition of such a major angiogenic pathway, and the presence of resistant and refractory patient populations complicate their use.3, 4 Thus, discovery of new therapeutic targets is crucial. Alternative angiogenic targets could lead to new therapeutics to complement and combine with the existing medications.
We previously characterized a novel antiangiogenic homoisoflavonoid derivative, SH-11037 (1; Figure 1a), in vitro5 and in vivo.6 SH-11037 had potent antiangiogenic activities in the laser-induced choroidal neovascularization (L-CNV) mouse model, a widely-used system that recapitulates some of the features of wet AMD.7 Therefore, characterization of its mechanism of action is important. We used an unbiased forward chemical genetics approach to identify SH-11037 protein targets that might be potentially druggable angiogenic mediators. We first synthesized two photoaffinity reagents 2 and 3 that retained antiangiogenic activity, and a control compound 4 (Figure 1a).8 The ester group in 2 (shared with SH-11037) was replaced by an amide in 3 for increased stability. The SH-11037-based affinity reagents 2 and 3 were immobilized and used to pull down protein binding partners from a porcine brain lysate. Affinity reagent 3 but not the negative control reagent 4 pulled down a specific protein target, which was identified as soluble epoxide hydrolase (Figure 1b; Supplementary Figure 1). Immunoblot confirmed the identity of the pulled down protein (Figure 1c).
Figure 1.

Soluble epoxide hydrolase (sEH) is a target of antiangiogenic homoisoflavonoid SH-11037. a) Structures of SH-11037 (1), ester affinity reagent (2), amide affinity reagent (3), and negative control reagent (4). BP, benzophenone moiety. b) Proteins pulled down with indicated reagents were separated by SDS-PAGE and silver stained, then identified by mass spectrometry. A unique band was present in pulldown with 3 but not 4, red box; asterisks represent non-specific bands. c) Immunoblot of pulled down protein using antibody against sEH. Silver-stained gel and immunoblot are representatives from at least two independent experiments. d) SH-11037 (yellow) docks in the substrate-binding cleft of sEH, as shown in this stereo view. Key residues are shown in green. e) 2D interaction diagram for docked SH-11037. The protein “pocket” is displayed with a line around the ligand, colored with the color of the nearest protein residue. The π-π stacking interactions are shown as green lines.
Docking of SH-11037 to sEH (Figure 1d) shows a binding mode in which the compound occupies nearly the entire active site of the enzyme. The homoisoflavonoid group of SH-11037 occupies the site where the catalytic Asp335 and Tyr466 residues of sEH are located.9 Both aromatic rings of the homoisoflavonoid group are involved in π-π interactions with sEH residues that include His524 and Trp336 (Figure 1e). The benzyl substituent of the peptidic moiety of SH-11037 is ensconced into a hydrophobic cavity created by Trp473, Met503, Ile363, and Phe362. The substituent is located near an opening through which the linker of 2 or 3 is attached to SH-11037. Extensive molecular dynamics simulations reveal that this moiety is flexible as illustrated by root-mean-square deviations for SH-11037 that range from 1.2 to 2.8 Å (Supplementary Figure 2). Animation of the molecular dynamics simulations (Supplementary movie) reveals that the benzene ring periodically adopts a conformation whereby the para position of the ring is exposed to solvent. This is likely the conformation that is adopted by the benzyl group in compounds 2 and 3.
Soluble epoxide hydrolase (sEH, encoded by EPHX2) is a 62 kDa bifunctional enzyme that has N-terminal lipid phosphatase (EC 3.1.3.76) and C-terminal epoxide hydrolase (EC 3.3.2.10) activities.10 While the physiological role of the lipid phosphatase activity of sEH is not fully understood, its epoxide hydrolase activity has been extensively studied due to its role in the metabolism of arachidonic acids’ epoxide derivatives, epoxyeicosatrienoic acids (EETs).11 EETs have proangiogenic effects resulting in accelerating tumor growth,12 and play a role in hypertension, pain and inflammation. Therefore, sEH inhibitors have been in clinical trials for treating related conditions.13, 14 But additionally, sEH is involved in the metabolism of the epoxides of ω-3 fatty acids, docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA), which are more abundant in the eye than EETs.15
Following the identification of sEH as a binding target of SH-11037, we tested whether the compound interferes with the epoxide hydrolase activity of sEH in vitro, compared to known sEH inhibitors. Trans-4-(4-[3-adamantan-1-yl-ureido]-cyclohexyloxy)-benzoic acid (t-AUCB (5); Figure 2a) is a specific inhibitor of the epoxide hydrolase activity of sEH, widely used in preclinical studies.16 Meanwhile, 7-(trifluoromethyl)-N-(4-(trifluoromethyl)phenyl) benzo[d]isoxazol-3-amine (7), is a structurally distinct benzisoxazole inhibitor with excellent potency and pharmacokinetic properties.17 Interestingly, SH-11037 inhibited sEH enzymatic activity in vitro in a concentration-dependent manner (Figure 2a), although not as potently as t-AUCB or 7. To test whether these effects are specific to SH-11037, we used as a negative control SH-11098 (6), which is a homoisoflavonoid that was found to be inactive in angiogenesis assays in vitro.5 This related compound had minimal inhibitory activity, suggesting that structural features of SH-11037 specifically interact with sEH.
Figure 2.

SH-11037 is an sEH inhibitor. a) SH-11037 (1) but not its inactive analog SH-11098 (6) significantly suppressed sEH enzymatic activity in vitro, IC50 = 0.15 μM (SH-11098 IC50 > 10 μM). The specific sEH inhibitors t-AUCB (5) and compound 7 were used as positive controls, IC50 = 9.5 nM for each. Mean ± SEM from triplicate wells shown. b) Michaelis-Menten kinetic response plot for sEH-mediated hydrolysis of fluorogenic substrate, PHOME, for varying SH-11037 concentrations. Mean ± SEM from triplicate wells shown. c) Lineweaver-Burk plot of these data suggests mixed-type inhibition. d) Dixon plot further supports mixed-type inhibition. e) Lipid profile of retina/choroid for DHA-related metabolites from L-CNV (three days post-laser) or control mice treated with vehicle, 10 μM t-AUCB, or 10 μM SH-11037. EDP, epoxydocosapentaenoic acids; DHDP, dihydroxydocosapentaenoic acids. f) The ratio of 19,20 EDP/DHDP between different treatment conditions and vehicle only (no laser) control mice indicates increased sEH levels/activity three days following laser induction compared to no laser control, **P<0.01, and a significant sEH inhibition by SH-11037, *P<0.05 vs. vehicle. One-way ANOVA, Dunnett’s post hoc tests. Mean ± SEM, n = 5 mice/treatment. Activity assay and kinetic analyses are representatives from at least two independent experiments.
Enzyme kinetics analysis showed that increasing concentrations of SH-11037 decreased Vmax and increased KM, revealing that SH-11037 is a mixed-type inhibitor of sEH (Figure 2b, c, d), with Ki = 1.73 ± 0.45 μM. Compound 7 is also a mixed-type inhibitor (Supplementary Figure 3). Furthermore, secondary plots of KMapp/Vmaxapp and 1/Vmaxapp vs. [SH-11037] and [7] fit the curves expected for mixed-type inhibition (Supplementary Figure 4). The catalytic mechanism of sEH proceeds as a nucleophilic attack onto the epoxide substrate by an Asp residue, which results in a tetrahedral intermediate, requiring activated water to release the diol and regenerate free enzyme.9 Given that sEH has two substrates (in our assay, the fluorogenic substrate 3-phenyl-cyano(6-methoxy-2-naphthalenyl)methyl ester-2-oxiraneacetic acid [PHOME] and water), and involves a covalent intermediate, it is possible that SH-11037 may bind and stabilize an enzyme species late in the catalytic cycle that is still in conformational equilibrium with the free enzyme. SH-11037 binding in the active site of such an enzyme species may not compete with the substrates. Taken together, these findings indicate that SH-11037 represents a novel, distinct chemotype from known sEH inhibitors,18 although interestingly, some other flavonoid natural products have previously been identified as mixed-type sEH inhibitors.19
After establishing SH-11037’s in vitro inhibition of sEH activity, it was crucial to assess whether the previously documented antiangiogenic effects of SH-110376 were mediated through the inhibition of sEH in the L-CNV model. Therefore, we analyzed the lipid profiles of the retina/choroid layers from mice, at an acute timepoint three days after CNV induction and intravitreal injections of 10 μM SH-11037 or t-AUCB. Since DHA is the most abundant bioactive lipid in the eye,15 DHA epoxy and dihydroxy metabolite levels were evaluated to investigate sEH activity in vivo. Interestingly, 19,20-epoxydocosapentaenoic acid (EDP) and its dihydroxy metabolite, 19,20-dihydroxydocosapentaenoic acid (DHDP) were the DHA metabolites most affected by sEH inhibition (Figure 2e). The ratio of 19,20-EDP to 19,20-DHDP decreased after induction of neovascularization, indicative of enhanced sEH activity under these conditions. However, this ratio was partially normalized after SH-11037 or t-AUCB treatment compared to the vehicle treated controls, indicating sEH inhibition in vivo (Figure 2f). Despite being less potent than t-AUCB in vitro (Figure 2a), SH-11037 performed comparably in vivo (Figure 2f), perhaps indicative of better ocular bioavailability than the existing inhibitor. To our knowledge, this is the first evidence that local sEH chemical inhibition can alter the lipid balance in the eye.
Given the significantly suppressed ratio of 19,20 EDP/DHDP after induction of CNV compared to the untreated control, suggestive of increased sEH activity, we investigated whether there are differences in sEH expression during neovascularization. Intriguingly, L-CNV treated mice demonstrated substantial upregulation of sEH in photoreceptor layers, both within and surrounding the neovascular lesion, compared to untreated eyes (Figure 3a). This upregulation of sEH in L-CNV was further confirmed in immunoblots of retina and choroid layers of laser-treated mouse eyes relative to untreated controls (Figure 3b). Co-immunostaining revealed co-localization of upregulated sEH levels with rod photoreceptors in the eyes of L-CNV mice compared to controls (Figure 3c, Supplementary Figure 5), but no overlap with markers of other retinal cell types, including retinal ganglion cells, horizontal cells, Müller glia, and cone photoreceptors (Supplementary Figures 5, 6, 7, 8). This increase in immunostaining corresponded to an increase in sEH activity in L-CNV eye lysates, which could be normalized by SH-11037 or compound 7 treatment (Figure 3d). Surprisingly, postmortem human wet AMD patients’ eyes also revealed changed sEH expression in the central retina compared to age-matched controls: an increase in the staining pattern of sEH in the inner retina seen in age-matched control retina, and aberrant expression in some photoreceptors (Figure 3e; Supplementary Figure 9). Together, these data strongly suggest a role for retinal sEH in the CNV process both in mice and humans.
Figure 3.
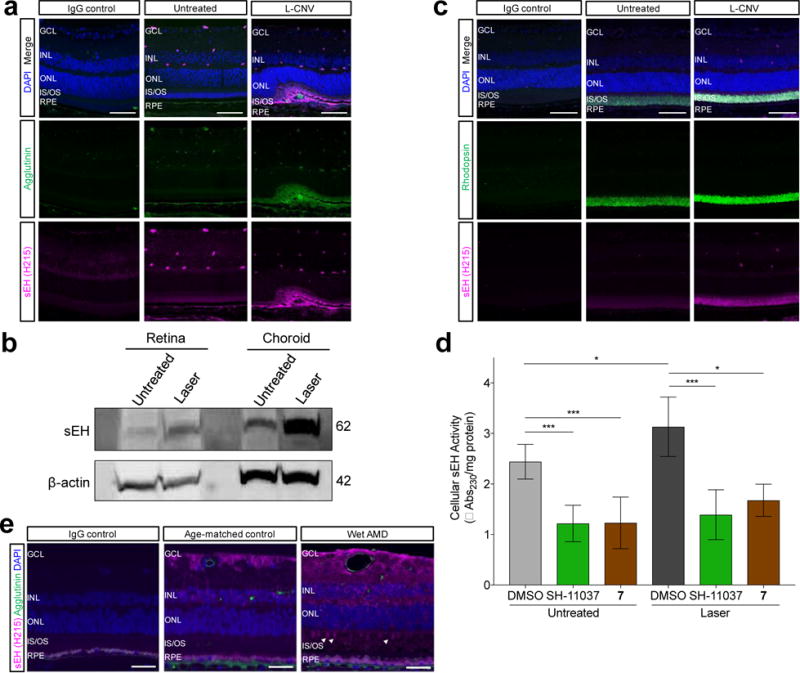
sEH is upregulated in the eyes of mice and humans undergoing neovascularization. a) Representative images of retinal sections from L-CNV and control eyes stained with DAPI (blue), agglutinin for vasculature (green), and sEH (magenta), showing upregulation of sEH in the outer retina in L-CNV sections three days post-laser. b) Immunoblot of sEH protein levels in mouse retina and choroid sections of laser treated mouse eyes three days post-laser compared to untreated controls; β-actin is a loading control. Pooled eyes from two independently-treated animals per condition. c) Representative images of retinal sections from L-CNV (three days post-laser) and control eyes stained with DAPI (blue), sEH (magenta), and rod marker rhodopsin (green), showing co-localization of upregulated sEH with rod photoreceptors. d) sEH activity is upregulated in L-CNV eye tissue (*P<0.05) and normalized by 20 μM SH-11037 or 7 treatment (***P<0.001), as indicated in a trans-stilbene oxide enzymatic activity assay performed three days post-laser. Mean ± SEM, ANOVA with Tukey’s post hoc tests. Pooled data from three experiments, n = 2–3 animals per condition per experiment. e) Representative images of central retinal sections from eyes of human wet AMD patients (78 years old) and age-matched controls (68 years old). sEH is magenta, vasculature (FITC-agglutinin) is green, and nuclei (DAPI) are blue. In wet AMD, sEH is increased in the inner retina, and aberrantly expressed in some photoreceptors (arrowheads). Scale bars = 50 μm. IgG is a negative control with preimmune primary antibodies. GCL, ganglion cell layer; INL, inner nuclear layer; ONL, outer nuclear layer; IS/OS, photoreceptor inner segments/outer segments; RPE, retinal pigment epithelium.
sEH is widely expressed in various tissues, with the highest levels of expression seen in liver, kidney, and brain in neuronal cell bodies and astrocytes.20, 21 In mouse eye development it is expressed in Müller glial cells,22 and in oxygen-induced retinopathy in neonatal mice, expression is seen in retinal ganglion cells, neovessels, and inner nuclear layer neurons.23 Our data suggest a differential expression pattern in the adult mouse eye, especially under CNV stress. Aged human eyes, in turn, have another pattern of expression, but this could also be a species difference: the rod-specific sEH expression seen in response to CNV in the rod-dominant, nocturnal murine eye might be expected to be shared by other cell types, as we observed, in the cone-dominant human macula.
Several studies have demonstrated potent antiangiogenic effects of DHA and its epoxides in the eye after genetic manipulation and/or dietary or i.p. administration,22, 24–28 although a few studies showed proangiogenic effects.23, 29, 30 To date however, inhibition of sEH using small molecules has only been tested in L-CNV using systemic administration through i.p. injections28, 29, and showed antiangiogenic effects only when diet was supplemented with ω-3 fatty acids or their epoxides.28 Given that the arachidonic acid pathway leading to EET formation is dominant systemically,31 unlike in the eye, the effects of EDP buildup after systemic sEH inhibition might be partially offset by the concomitant proangiogenic EET buildup. Thus, local sEH inhibition is appealing.
To address this, we sought to test sEH inhibitors locally, using intravitreal injections, to minimize any systemic side effects and focus on understanding the effects of sEH in the eye specifically. We previously showed that SH-11037 was effective at doses ≥ 1 μM in this context.6 Here, to validate sEH as a key target, we assessed the antiangiogenic effect of two chemically distinct small molecule inhibitors of sEH, t-AUCB (5) and compound 7, in L-CNV (Figure 4). A single injection of either t-AUCB or compound 7 dose-dependently suppressed CNV lesion vascular volume compared to vehicle (Figure 4a–c), and comparable to the standard of care-equivalent anti-VEGF164 antibody, suggesting indeed that sEH inhibition directly in the eye does not require ω-3 supplementation for antiangiogenic efficacy. Additionally, in order to confirm our observations in a different model system, we tested t-AUCB and compound 7 in the choroidal sprouting assay, as an ex vivo model of CNV.6 Interestingly, both t-AUCB and compound 7 suppressed the ability of choroidal tissues to form sprouts (Figure 4d–e).
Figure 4.

Local application of sEH inhibitors dose-dependently suppresses neovascularization. a) Representative images from confocal microscopy of agglutinin stained CNV lesions 14 days post-laser, scale bar = 50 μm. b) & c) Dose-dependent inhibition of the volume of CNV lesions by b) t-AUCB (5) and c) sEH inhibitor 7 compared to vehicle control. Mean ± SEM, n = 6–15 animals/treatment (one eye per animal). *P<0.05, **P<0.01, ***P<0.001 compared to vehicle, one-way ANOVA, Dunnett’s post hoc tests. d) & e) Inhibition of mouse choroidal sprouting ex vivo by d) t-AUCB (5) and e) sEH inhibitor 7 compared to vehicle control. Mean ± SEM, n = 4 eyes/treatment, representative data from at least two independent experiments. Axes for measurement of sprouting distance shown in yellow. Scale bars = 1 mm. *P<0.05, **P<0.01 compared to vehicle, one-way ANOVA, Dunnett’s post hoc tests. f) Summary of SH-11037’s mechanism. By inhibiting sEH, SH-11037 decreases the formation of 19,20-DHDP (dihydroxydocosapentaenoic acid), and increases levels of docosahexaenoic acid (DHA)-derived 19,20-EDP (epoxydocosapentaenoic acid), with antiangiogenic effects.
The effect of ω-3 polyunsaturated fatty acids (PUFA) levels on choroidal neovascularization has been previously reported in vivo in the L-CNV model24, 25 and in wet AMD patients.32 Interestingly, in the retina of sEH knockout mice, pronounced changes in the levels of 19,20-DHDP, but not the epoxy metabolites of DHA, compared to wild-type control mice were observed.22 Moreover, 19,20-DHDP could rescue the suppressed developmental retinal angiogenesis phenotype observed with these mice,22 and as noted i.p. injections of 19,20-EDP suppressed CNV lesion volume in the L-CNV mouse model.25, 28 While the explanation for the absence of changes in other EDPs after sEH inhibition or knockout is not clear, it is possible that the DHA epoxy (19,20-EDP) and diol (19,20-DHDP) metabolites and the activity of sEH are the major factors underlying the observed changes in retinal and choroidal angiogenesis rather than DHA levels themselves. Despite the role of DHA in the eye, the variability in the circulating levels of DHA among individuals suggests that dietary intake of ω-3 PUFA would not be sufficient alone for the prevention or treatment of wet AMD.32 Therefore, local small molecule inhibition of sEH is an appealing therapeutic approach of significant interest for wet AMD patients to augment DHA epoxy metabolite levels with or without dietary supplementation of ω-3 PUFA (Figure 4f).
In conclusion, our findings reveal not only the target of an antiangiogenic molecule and a novel chemotype for sEH inhibition, but also a central role for local sEH in ocular neovascularization. Developing novel, ocular specific sEH-targeted therapies is a possible approach to complement or combine with the existing anti-VEGF medications to overcome their limitations and tackle multiple angiogenesis signaling pathways for improved treatment of wet AMD; we already showed that SH-11037 can synergize with an anti-VEGF antibody in L-CNV.6 Additionally, sEH and its inhibition using small molecules such as SH-11037 could be further investigated in other ocular neovascular diseases including PDR and ROP.
METHODS
Details of experimental procedures are provided in the Supporting Information.
Supplementary Material
Acknowledgments
We thank M. Sturek for the porcine brain, and K. Maddipati at the Wayne State University Lipidomics Core, supported in part by NIH/NCRR S10RR027926. T.W.C. acknowledges support from the International Retinal Research Foundation, the Retina Research Foundation, the BrightFocus Foundation, NIH/NEI R01EY025641, and an unrestricted grant from Research to Prevent Blindness, Inc. S.-Y.S. was supported by the Bio & Medical Technology Development Program of the National Research Foundation funded by the Korean government, MSIP (NRF-2017M3A9C8027781) and the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI-HI14C1135). This publication was also made possible in part by R01EY018358 (M.E.B.), NIH/NCATS UL1TR001108 and KL2TR001106.
Footnotes
Supporting Information Available: This material is available free of charge via the Internet. Experimental procedures, Supplementary Figures 1−9, and Supplementary Movie.
AUTHOR INFORMATION
R.S.S., B.L., S.Y.S., X.F., and T.W.C. are named inventors on patent applications related to the current work.
No other authors declare competing financial interests.
References
- 1.Das A, McGuire PG. Retinal and choroidal angiogenesis: pathophysiology and strategies for inhibition. Prog Retinal Eye Res. 2003;22:721–748. doi: 10.1016/j.preteyeres.2003.08.001. [DOI] [PubMed] [Google Scholar]
- 2.Prasad PS, Schwartz SD, Hubschman JP. Age-related macular degeneration: current and novel therapies. Maturitas. 2010;66:46–50. doi: 10.1016/j.maturitas.2010.02.006. [DOI] [PubMed] [Google Scholar]
- 3.Falavarjani KG, Nguyen QD. Adverse events and complications associated with intravitreal injection of anti-VEGF agents: a review of literature. Eye. 2013;27:787–794. doi: 10.1038/eye.2013.107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Lux A, Llacer H, Heussen FM, Joussen AM. Non-responders to bevacizumab (Avastin) therapy of choroidal neovascular lesions. Br J Ophthalmol. 2007;91:1318–1322. doi: 10.1136/bjo.2006.113902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Basavarajappa HD, Lee B, Lee H, Sulaiman RS, An H, Magana C, Shadmand M, Vayl A, Rajashekhar G, Kim EY, Suh YG, Lee K, Seo SY, Corson TW. Synthesis and biological evaluation of novel homoisoflavonoids for retinal neovascularization. J Med Chem. 2015;58:5015–5027. doi: 10.1021/acs.jmedchem.5b00449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Sulaiman RS, Merrigan S, Quigley J, Qi X, Lee B, Boulton ME, Kennedy B, Seo SY, Corson TW. A novel small molecule ameliorates ocular neovascularisation and synergises with anti-VEGF therapy. Sci Rep. 2016;6:25509. doi: 10.1038/srep25509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Grossniklaus HE, Kang SJ, Berglin L. Animal models of choroidal and retinal neovascularization. Prog Retinal Eye Res. 2010;29:500–519. doi: 10.1016/j.preteyeres.2010.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Lee B, Sun W, Lee H, Basavarajappa H, Sulaiman RS, Sishtla K, Fei X, Corson TW, Seo SY. Design, synthesis and biological evaluation of photoaffinity probes of antiangiogenic homoisoflavonoids. Bioorg Med Chem Lett. 2016;26:4277–4281. doi: 10.1016/j.bmcl.2016.07.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Borhan B, Jones AD, Pinot F, Grant DF, Kurth MJ, Hammock BD. Mechanism of soluble epoxide hydrolase. Formation of an alpha-hydroxy ester-enzyme intermediate through Asp-333. J Biol Chem. 1995;270:26923–26930. doi: 10.1074/jbc.270.45.26923. [DOI] [PubMed] [Google Scholar]
- 10.Harris TR, Hammock BD. Soluble epoxide hydrolase: gene structure, expression and deletion. Gene. 2013;526:61–74. doi: 10.1016/j.gene.2013.05.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zhang G, Kodani S, Hammock BD. Stabilized epoxygenated fatty acids regulate inflammation, pain, angiogenesis and cancer. Prog Lipid Res. 2014;53:108–123. doi: 10.1016/j.plipres.2013.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Panigrahy D, Edin ML, Lee CR, Huang S, Bielenberg DR, Butterfield CE, Barnes CM, Mammoto A, Mammoto T, Luria A, Benny O, Chaponis DM, Dudley AC, Greene ER, Vergilio JA, Pietramaggiori G, Scherer-Pietramaggiori SS, Short SM, Seth M, Lih FB, Tomer KB, Yang J, Schwendener RA, Hammock BD, Falck JR, Manthati VL, Ingber DE, Kaipainen A, D’Amore PA, Kieran MW, Zeldin DC. Epoxyeicosanoids stimulate multiorgan metastasis and tumor dormancy escape in mice. J Clin Invest. 2012;122:178–191. doi: 10.1172/JCI58128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Morisseau C, Hammock BD. Impact of soluble epoxide hydrolase and epoxyeicosanoids on human health. Annu Rev Pharmacol Toxicol. 2013;53:37–58. doi: 10.1146/annurev-pharmtox-011112-140244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Lazaar AL, Yang L, Boardley RL, Goyal NS, Robertson J, Baldwin SJ, Newby DE, Wilkinson IB, Tal-Singer R, Mayer RJ, Cheriyan J. Pharmacokinetics, pharmacodynamics and adverse event profile of GSK2256294, a novel soluble epoxide hydrolase inhibitor. Br J Clin Pharmacol. 2016;81:971–979. doi: 10.1111/bcp.12855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Arterburn LM, Hall EB, Oken H. Distribution, interconversion, and dose response of n-3 fatty acids in humans. Am J Clin Nutr. 2006;83:1467S–1476S. doi: 10.1093/ajcn/83.6.1467S. [DOI] [PubMed] [Google Scholar]
- 16.Fromel T, Jungblut B, Hu J, Trouvain C, Barbosa-Sicard E, Popp R, Liebner S, Dimmeler S, Hammock BD, Fleming I. Soluble epoxide hydrolase regulates hematopoietic progenitor cell function via generation of fatty acid diols. Proc Natl Acad Sci U S A. 2012;109:9995–10000. doi: 10.1073/pnas.1206493109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Shen HC, Ding FX, Deng Q, Xu S, Tong X, Zhang X, Chen Y, Zhou G, Pai LY, Alonso-Galicia M, Roy S, Zhang B, Tata JR, Berger JP, Colletti SL. A strategy of employing aminoheterocycles as amide mimics to identify novel, potent and bioavailable soluble epoxide hydrolase inhibitors. Bioorg Med Chem Lett. 2009;19:5716–5721. doi: 10.1016/j.bmcl.2009.08.006. [DOI] [PubMed] [Google Scholar]
- 18.Shen HC, Hammock BD. Discovery of inhibitors of soluble epoxide hydrolase: a target with multiple potential therapeutic indications. J Med Chem. 2012;55:1789–1808. doi: 10.1021/jm201468j. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Thao NP, Luyen BT, Kim JH, Jo AR, Dat NT, Kiem PV, Minh CV, Kim YH. Identification, characterization, kinetics, and molecular docking of flavonoid constituents from Archidendron clypearia (Jack) Nielsen leaves and twigs. Bioorg Med Chem. 2016;24:3125–3132. doi: 10.1016/j.bmc.2016.05.034. [DOI] [PubMed] [Google Scholar]
- 20.Norwood S, Liao J, Hammock BD, Yang GY. Epoxyeicosatrienoic acids and soluble epoxide hydrolase: potential therapeutic targets for inflammation and its induced carcinogenesis. Am J Transl Res. 2010;2:447–457. [PMC free article] [PubMed] [Google Scholar]
- 21.Sura P, Sura R, Enayetallah AE, Grant DF. Distribution and expression of soluble epoxide hydrolase in human brain. J Histochem Cytochem. 2008;56:551–559. doi: 10.1369/jhc.2008.950659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hu J, Popp R, Fromel T, Ehling M, Awwad K, Adams RH, Hammes HP, Fleming I. Müller glia cells regulate Notch signaling and retinal angiogenesis via the generation of 19,20-dihydroxydocosapentaenoic acid. J Exp Med. 2014;211:281–295. doi: 10.1084/jem.20131494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Shao Z, Fu Z, Stahl A, Joyal JS, Hatton C, Juan A, Hurst C, Evans L, Cui Z, Pei D, Gong Y, Xu D, Tian K, Bogardus H, Edin ML, Lih F, Sapieha P, Chen J, Panigrahy D, Hellstrom A, Zeldin DC, Smith LE. Cytochrome P450 2C8 ω3-long-chain polyunsaturated fatty acid metabolites increase mouse retinal pathologic neovascularization–brief report. Arterioscler Thromb, Vasc Biol. 2014;34:581–586. doi: 10.1161/ATVBAHA.113.302927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Moghaddam-Taaheri S, Agarwal M, Amaral J, Fedorova I, Agron E, Salem N, Jr, Chew E, Becerra SP. Effects of docosahexaenoic acid in preventing experimental choroidal neovascularization in rodents. J Clin Exp Ophthalmol. 2011;2:187. doi: 10.4172/2155-9570.1000187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Yanai R, Mulki L, Hasegawa E, Takeuchi K, Sweigard H, Suzuki J, Gaissert P, Vavvas DG, Sonoda KH, Rothe M, Schunck WH, Miller JW, Connor KM. Cytochrome P450-generated metabolites derived from omega-3 fatty acids attenuate neovascularization. Proc Natl Acad Sci U S A. 2014;111:9603–9608. doi: 10.1073/pnas.1401191111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Stahl A, Sapieha P, Connor KM, Sangiovanni JP, Chen J, Aderman CM, Willett KL, Krah NM, Dennison RJ, Seaward MR, Guerin KI, Hua J, Smith LE. Short communication: PPAR gamma mediates a direct antiangiogenic effect of omega 3-PUFAs in proliferative retinopathy. Circ Res. 2010;107:495–500. doi: 10.1161/CIRCRESAHA.110.221317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Connor KM, SanGiovanni JP, Lofqvist C, Aderman CM, Chen J, Higuchi A, Hong S, Pravda EA, Majchrzak S, Carper D, Hellstrom A, Kang JX, Chew EY, Salem N, Jr, Serhan CN, Smith LE. Increased dietary intake of omega-3-polyunsaturated fatty acids reduces pathological retinal angiogenesis. Nat Med. 2007;13:868–873. doi: 10.1038/nm1591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Hasegawa E, Inafuku S, Mulki L, Okunuki Y, Yanai R, Smith KE, Kim CB, Klokman G, Bielenberg DR, Puli N, Falck JR, Husain D, Miller JW, Edin ML, Zeldin DC, Lee KSS, Hammock BD, Schunck WH, Connor KM. Cytochrome P450 monooxygenase lipid metabolites are significant second messengers in the resolution of choroidal neovascularization. Proc Natl Acad Sci U S A. 2017;114:E7545–E7553. doi: 10.1073/pnas.1620898114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Gong Y, Fu Z, Edin ML, Liu CH, Wang Z, Shao Z, Fredrick TW, Saba NJ, Morss PC, Burnim SB, Meng SS, Lih FB, Lee KS, Moran EP, SanGiovanni JP, Hellstrom A, Hammock BD, Zeldin DC, Smith LE. Cytochrome P450 oxidase 2C inhibition adds to omega-3 long-chain polyunsaturated fatty acids protection against retinal and choroidal neovascularization. Arterioscler Thromb, Vasc Biol. 2016;36:1919–1927. doi: 10.1161/ATVBAHA.116.307558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Gong Y, Shao Z, Fu Z, Edin ML, Sun Y, Liegl RG, Wang Z, Liu CH, Burnim SB, Meng SS, Lih FB, SanGiovanni JP, Zeldin DC, Hellstrom A, Smith LE. Fenofibrate inhibits cytochrome P450 epoxygenase 2c activity to suppress pathological ocular angiogenesis. EBioMedicine. 2016;13:201–211. doi: 10.1016/j.ebiom.2016.09.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Wang W, Zhu J, Lyu F, Panigrahy D, Ferrara KW, Hammock B, Zhang G. omega-3 polyunsaturated fatty acids-derived lipid metabolites on angiogenesis, inflammation and cancer. Prostaglandins Other Lipid Mediators. 2014;113–115:13–20. doi: 10.1016/j.prostaglandins.2014.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Merle BM, Benlian P, Puche N, Bassols A, Delcourt C, Souied EH, Nutritional A. M. D. Treatment Study Group Circulating omega-3 fatty acids and neovascular age-related macular degeneration. Invest Ophthalmol Visual Sci. 2014;55:2010–2019. doi: 10.1167/iovs.14-13916. [DOI] [PubMed] [Google Scholar]
- 32.Souied EH, Delcourt C, Querques G, Bassols A, Merle B, Zourdani A, Smith T, Benlian P, Nutritional A.M.D. Treatment Study Group Oral docosahexaenoic acid in the prevention of exudative age-related macular degeneration: the Nutritional AMD Treatment 2 study. Ophthalmology. 2013;120:1619–1631. doi: 10.1016/j.ophtha.2013.01.005. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.