

Abstract
Glycogen loading (GL), a well-known type of sports conditioning, in combination with exercise and a high carbohydrate diet (HCD) for 1 week enhances individual endurance capacity through muscle glycogen supercompensation. This exercise-diet combination is necessary for successful GL. Glycogen in the brain contributes to hippocampus-related memory functions and endurance capacity. Although the effect of HCD on the brain remains unknown, brain supercompensation occurs following exhaustive exercise (EE), a component of GL. We thus employed a rat model of GL and examined whether GL increases glycogen levels in the brain as well as in muscle, and found that GL increased glycogen levels in the hippocampus and hypothalamus, as well as in muscle. We further explored the essential components of GL (exercise and/or diet conditions) to establish a minimal model of GL focusing on the brain. Exercise, rather than a HCD, was found to be crucial for GL-induced hyper-glycogen in muscle, the hippocampus and the hypothalamus. Moreover, EE was essential for hyper-glycogen only in the hippocampus even without HCD. Here we propose the EE component of GL without HCD as a condition that enhances brain glycogen stores especially in the hippocampus, implicating a physiological strategy to enhance hippocampal functions.
Introduction
Glycogen is an important energy source for muscle during exercise, and it is depleted with increased intensity and/or duration of exercise1. Such glycogen depletion leads to muscle fatigue during endurance exercise2–4. To avoid muscle fatigue, muscle glycogen-loading (GL) – a well-established sports conditioning strategy including both exercise and diet for 1 week before competition – increases muscle glycogen levels and enhances the endurance capacity in humans and animals5–8.
GL has been developed using a popular theory called “muscle glycogen supercompensation”, which is characterized by an initial depletion of muscle glycogen levels followed by a considerable replenishment of muscle glycogen 24–48 hours after acute exercise9–11. Åstrand first proposed the classic GL protocol: 3 days of exercising and a low-carbohydrate diet to induce muscle glycogen depletion followed by 3 days of a high-carbohydrate diet for hyper muscle glycogen12. However, classic GL protocol is complicated and occasionally induces restlessness with hypoglycemia due to the 3 days of low carbohydrate diet5,13. To solve these problems, Sherman et al. established a novel GL protocol using only a high-carbohydrate diet and exercises that induced hyper muscle glycogen mirroring the classic protocol7. This novel GL protocol is popular among modern endurance athletes14,15.
Interestingly, similar type of glycogen supercompensation phenomena in the brain were observed in our recent study16. We found that brain glycogen decreases with exhaustive exercise17, particularly in the hippocampus and cortex, and that supercompensation occurs, as it does in muscle, 6 hours after exhaustive exercise in rats16. These elevated glycogen levels were sustained for up to 24 hours after exhaustive exercise16. Therefore, we have hypothesized that GL increases brain glycogen storage, as observed in muscle, based on the exercise-induced glycogen-supercompensation theory as mentioned above.
Brain glycogen, which is localized in astrocytes and produces lactate as a neuronal energy source and/or neuromodulator, plays a critical role in memory function and exercise endurance18–22. Chronic exercise that enhances endurance capacity and cognitive function has also been accompanied with elevated hippocampal glycogen levels in normal rats16. In our very recent study, 4 weeks of moderate exercise is effective in improving the declining memory function (hippocampal) in type 2 diabetic rats, and this exercise-induced hippocampal-memory amelioration has been associated with hyper-glycogen levels in the hippocampus23. Indeed, recent studies have demonstrated that pharmacological or genetic inhibition of hippocampal glycogen metabolism impairs memory formation and compromises endurance capacity19–22,24. Furthermore, pharmacologically elevated brain glycogen levels in the brain protect neuronal activities under insulin-induced severe hypoglycemia25. Therefore, if GL increases brain glycogen levels as it does in muscle, GL is a possible strategy to enhance brain functions relating with memory and endurance performance.
To study the effects of GL on brain, determination of an appropriate GL condition for animals is needed. To date, Shinohara’s 1 week GL model, which is composed of an exhaustive exercise followed by a moderate exercise (10 min with a weight equal to 1–2% of body mass) and then rest with HCD, is useful as a reference because in their model hyper-glycogen storage has appeared in rat muscle after the GL8. However, their analysis is inadequate since the roles of the respective components of GL, namely EX (exhaustive exercise followed by moderate exercise and rest) and HCD, on glycogen storage is still unclear, and they did not address brain glycogen storage.
We thus performed four experiments as follows: First, we employed GL protocols, EX with HCD, in rat models and assessed whether glycogen levels increased after GL in various regions of the brains especially in the hippocampus, and in muscle (Experiment 1, Fig. 1A). Subsequently, we examined which GL component is dominant, EX or HCD, in inducing hyper-glycogen levels in the brain (HCD: Experiment 2, Fig. 1B; EX: Experiment 3, Fig. 1C, exercise conditions: Experiment 4, Fig. 1D). Through these analyses we intended to clarify whether GL or one of its two main components (EX and HCD) may have positive effects on brain glycogen storage.
Figure 1.
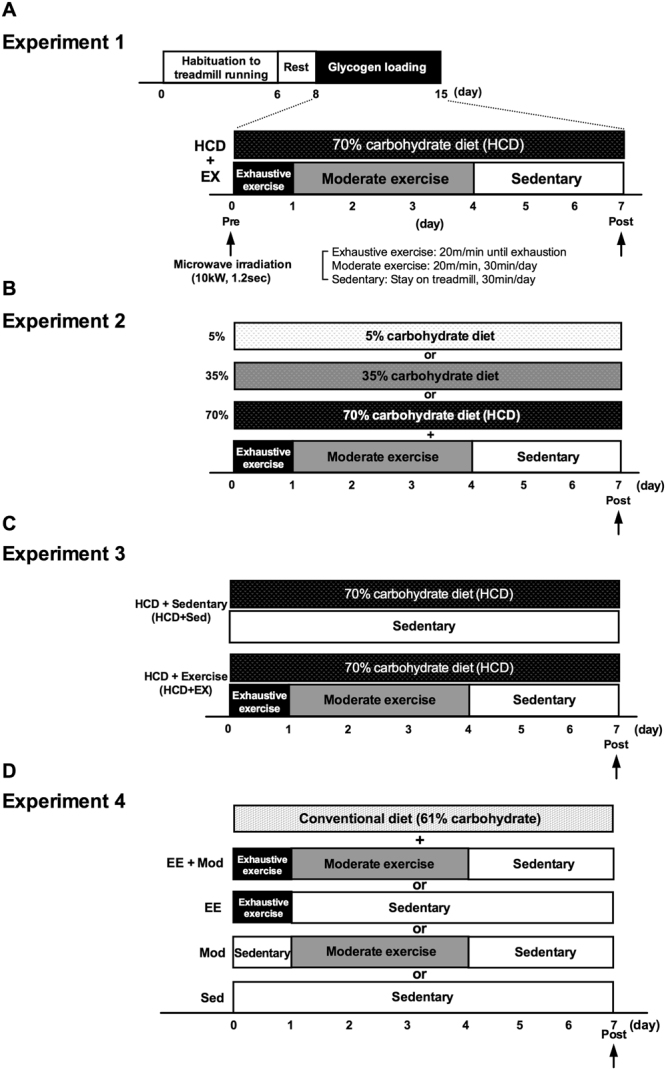
Experimental design. (A) One-week GL protocol for rat model (Experiment 1). Rats were divided into a Pre-GL and a Post-GL group, and underwent GL protocols with HCD (70% carbohydrates) and EX (day 1: exhaustive exercise (20 m/min until exhaustion), days 2–4: moderate exercise (20 m/min, 30 min/day), days 5–7: sedentary (rest on treadmill, 30 min/day)). Rats were sacrificed using high-power microwave irradiation (10 kW, 1.2 sec). (B) GL protocol to investigate the importance of the HCD component during GL (Experiment 2). Rats were divided into three diet groups each with different amount of carbohydrate (low: 5%, middle: 35%, and high: 70%) and fixed EX. Each group underwent GL protocol. (C) GL protocol to investigate the importance of the EX component during GL (Experiment 3). Rats were divided into two groups (HCD + EX and HCD + Sed), and underwent GL protocol. (D) GL with conventional-diet protocol to investigate the essential exercise condition during GL (Experiment 4). Rats were divided into four exercise groups (EE + Mod, a combination of exhaustive exercise and moderate exercise; EE, exhaustive exercise alone; Mod, moderate exercise alone; and Sed, sedentary), and underwent GL with a conventional diet (61% carbohydrates).
Results
GL increases glycogen levels in both muscle and brain
Rats underwent 1-week of GL, which consisted of several EX conditions and a HCD. Glycogen levels in muscle, liver and brain after GL were measured by using microwave irradiation (Fig. 2A). Muscle glycogen, but not liver glycogen, increased after GL (EX with HCD) (P < 0.01; Exp. 1, Fig. 2B) compared to pre-GL. Concomitantly, GL also led to increased brain glycogen levels, particularly in the hippocampus and hypothalamus (P < 0.05; Exp. 1, Fig. 2C). The rates of glycogen increase in muscle, the hippocampus and the hypothalamus were 79%, 12%, and 10%, respectively, implying that the GL model is effective for increasing brain glycogen, although the extent of the increase is much smaller than that of muscle.
Figure 2.
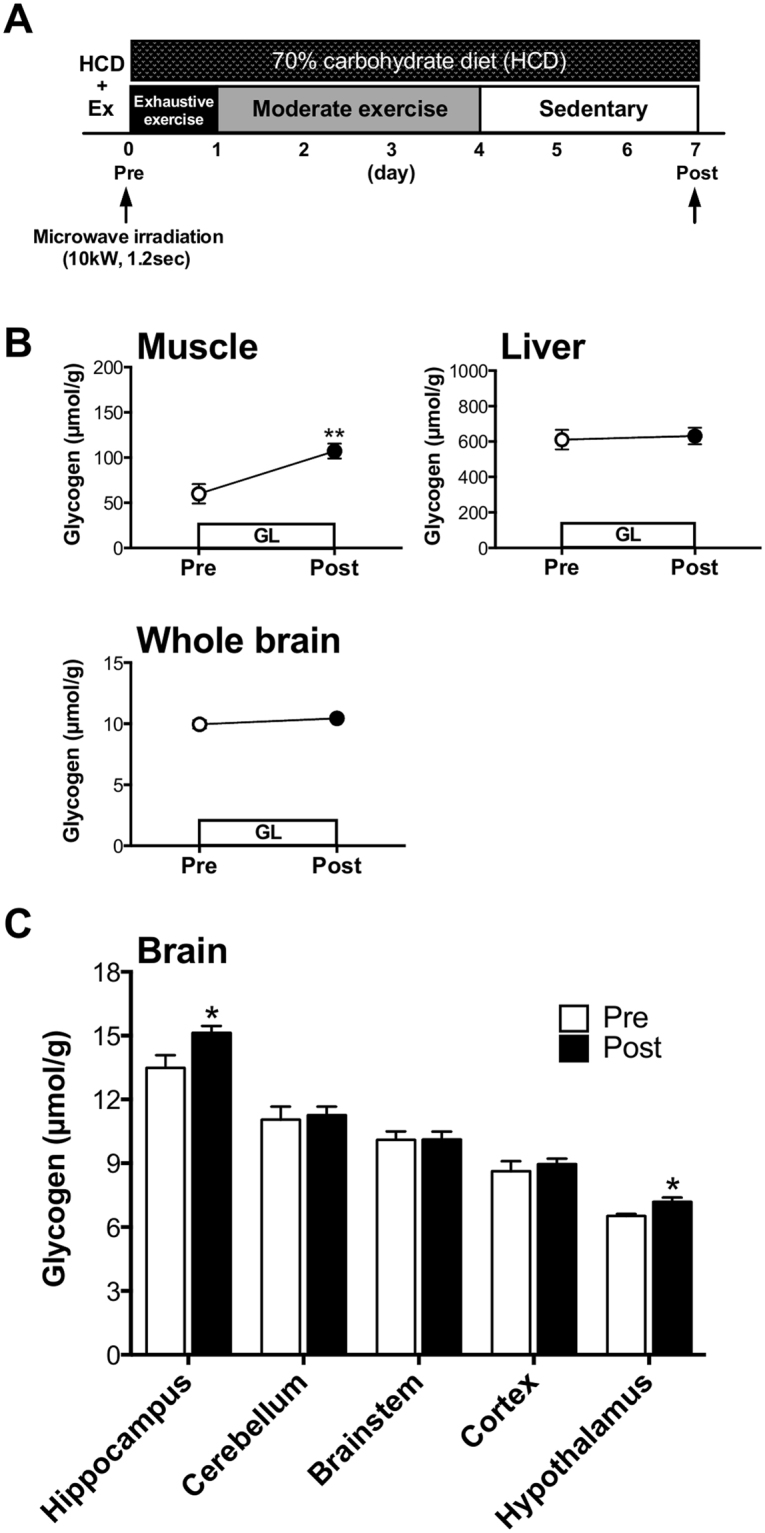
GL increased glycogen levels in muscle and the brains. (A) Experimental design. (B) Glycogen levels in the whole brain, muscle, and liver for pre- and post-GL. (C) Glycogen levels in the five brain loci (hippocampus, cerebellum, brainstem cortex and hypothalamus). Data are expressed as mean ± standard error (n = 6–8/group). *P < 0.05; **P < 0.01 versus pre-GL group (unpaired t test).
HCD during GL is not necessary for GL-induced hyper-brain glycogen levels in the brain
Rats were performed GL protocols with EX and different percentage of carbohydrate during GL (Fig. 3A). Three kinds of diets were adjusted same calorie per gram to examine whether HCD component in GL is necessary to induce hyper-hippocampal glycogen levels (Tables 1 and 2). During GL, only total carbohydrate intake increased with the increase of carbohydrate content in the diet (P < 0.001; Exp. 2, Table 3). Muscle glycogen levels did not differ significantly (Exp. 2, Fig. 3B) as the percentage of carbohydrates diet increased, but had a positive correlation with total carbohydrate intake (r = 0.43, P < 0.05; Exp. 2, Fig. 3C). In contrast, liver glycogen levels of the 70% carbohydrate diet group were 54% higher than those of the 5% carbohydrate diet group (Exp. 2, Fig. 3B). Furthermore, there was a positive correlation between carbohydrate intake and liver glycogen levels (r = 0.68, P < 0.001; Exp. 2, Fig. 3C). Regarding the relationship with fat intake and glycogen levels, there was significant negative correlation between fat intake and liver glycogen levels (r = −0.6, P < 0.01; Exp. 2, Fig. S2B), and a similar tendency was seen also in the muscle glycogen (r = −0.35, P = 0.09; Exp. 2, Fig. S2A). Of note, there was no significant change in brain glycogen levels in the carbohydrate-dose-dependent experiment without a significant correlation with carbohydrate intake (hippocampus: r = 0.37, P = 0.08, hypothalamus: r = −0.08, P = 0.71, cortex: r = 0.002, P = 0.99) (Exp. 2, Fig. 3B and C). Thus, muscle and brain glycogen levels may have similar responses to a gradual increase of carbohydrates in the diet during GL while their correlation with total carbohydrate intake was organ specific.
Figure 3.

GL-induced hyper-glycogen in the muscle, but not in brain, is associated with carbohydrate intake. (A) Experimental design. (B) Glycogen levels in muscle, liver, and brain (hippocampus, hypothalamus, and cortex). Data are expressed as mean ± standard error (n = 7–8/group) **P < 0.01 versus 5% group (Dunnett’s post hoc test). (C) Correlation between carbohydrate intake and glycogen levels in muscle, liver, hippocampus, hypothalamus and cortex. Data are expressed as mean ± standard error (n = 7–8/group). Correlations are shown between the carbohydrate intake and glycogen levels. Lines in the scatter plots show significant correlation (by Pearson’s product-moment correlations test).
Table 1.
Compositions of diets with different amounts of carbohydrates.
| Ingredients/Diet | 5% | 35% | 70% |
|---|---|---|---|
| Casein | 23.1 | 23.1 | 23.1 |
| DL-methionine | 0.3 | 0.3 | 0.3 |
| Corn starch | 0.0 | 27.8 | 60.1 |
| Sucrose | 4.6 | 4.6 | 4.6 |
| Corn oil | 9.6 | 5.5 | 0.7 |
| Lard | 19.2 | 10.9 | 1.4 |
| Cellulose powder | 38.5 | 23.1 | 5.1 |
| AIN-76 mineral mix | 3.5 | 3.5 | 3.5 |
| AIN-76 vitamin mix | 1.0 | 1.0 | 1.0 |
| Choline bitartrate | 0.2 | 0.2 | 0.2 |
| Total (%) | 100.0 | 100.0 | 100.0 |
Table 2.
Carbohydrate, fat, and protein ratios of three diets.
| Diet | Nutrients | Content (g) | Calorie ratio (%) | Energy (kcal/100 g) | Total calorie (kcal/100 g) |
|---|---|---|---|---|---|
| 5% | Carbohydrate | 4.6 | 5.0 | 18.5 | 370.0 |
| Fat | 28.8 | 70.0 | 259.0 | ||
| Protein | 23.1 | 25.0 | 92.5 | ||
| 35% | Carbohydrate | 32.4 | 35.0 | 129.5 | 370.0 |
| Fat | 16.4 | 40.0 | 148.0 | ||
| Protein | 23.1 | 25.0 | 92.5 | ||
| 70% | Carbohydrate | 64.8 | 70.0 | 259.0 | 370.0 |
| Fat | 2.1 | 5.0 | 18.5 | ||
| Protein | 23.1 | 25.0 | 92.5 |
Table 3.
Baseline physiological variables after GL.
| Parameter/Carbohydrate ratio in diet | 5% | 35% | 70% |
|---|---|---|---|
| Body weight (g) | 296.0 ± 3.2 | 298.0 ± 3.1 | 295.0 ± 4.4 |
| Blood glucose (mM) | 4.8 ± 0.1 | 4.5 ± 0.1 | 4.8 ± 0.1 |
| Total food intake (g/6 days) | 92.6 ± 5.9 | 90.7 ± 4.3 | 94.5 ± 6.5 |
| Total calorie intake (kcal/100 g/6 days) | 273.0 ± 20.0 | 268.5 ± 14.7 | 285.0 ± 24.0 |
| Total carbohydrate intake (g/6 days) | 3.6 ± 0.3 | 25.4 ± 1.4*** | 54.1 ± 4.5*** |
Data are expressed as mean ± standard error (n = 7–8/group). ***P < 0.001 versus 5%.
Exercise during GL is required for GL-induced hyper-glycogen levels in muscle and brain
Since GL is composed of HCD and EX, next, we examined whether EX component in GL is necessary inducing hyper-hippocampal glycogen levels (Fig. 4A). Muscle glycogen levels increased with exercise (P < 0.05; Exp. 3, Fig. 4B). Brain glycogen levels in the hippocampus and hypothalamus, but not in the cortex, increased with EX (P < 0.05; Exp. 3, Fig. 4D). The rates of glycogen increase in muscle, the hippocampus and the hypothalamus were 34%, 13%, and 29%, respectively, implying that the GL, particularly the EX component, is crucial for increasing muscle and brain glycogen levels and, in particular, EX-induced brain glycogen increases are region specific.
Figure 4.
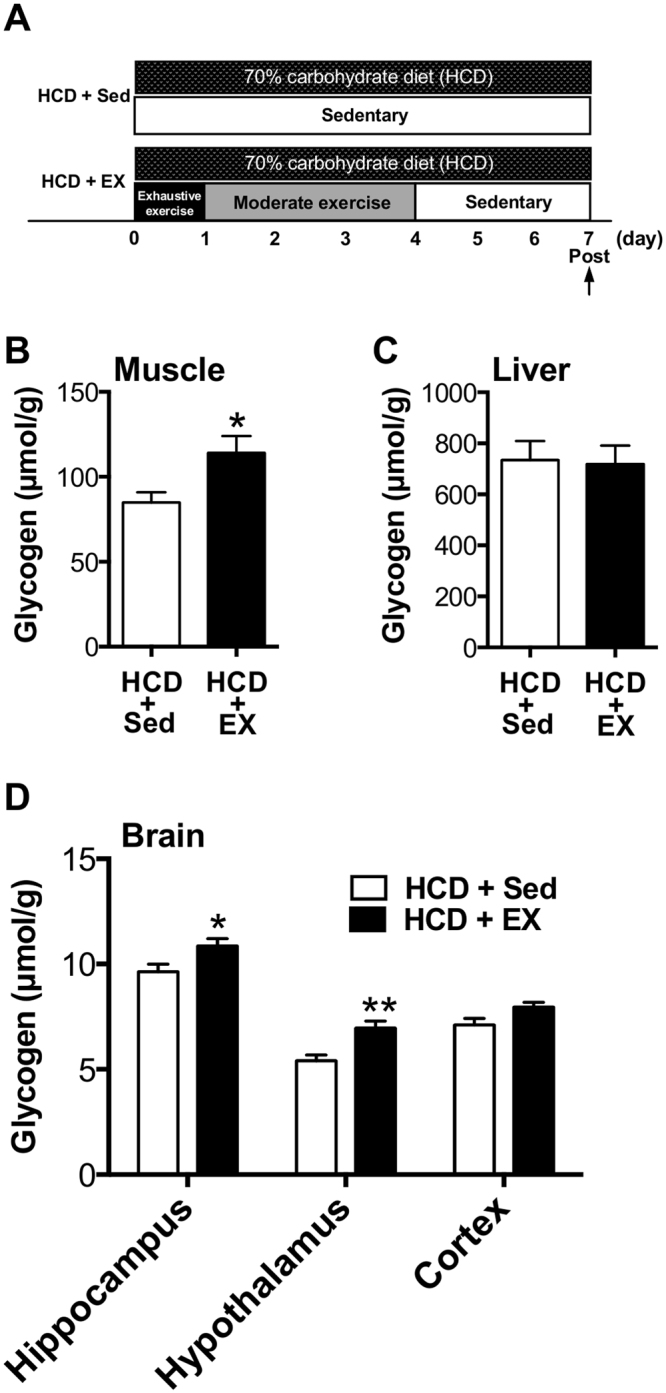
EX during GL is required for GL-induced hyper-glycogen in the muscle and the brain. (A) Experimental design. (B) Muscle glycogen. (C) Liver glycogen. (D) Brain glycogen levels in hippocampus, hypothalamus, and cortex. Data are expressed as mean ± standard error (n = 4–5/group) *P < 0.05; **P < 0.01 versus HCD ± Sed group (unpaired t test).
Exhaustive exercise is an essential for inducing hyper-glycogen levels in the hippocampus without a HCD
Finally, we tried to determine which conditions of exercise (EE + Mod: a combination of exhaustive exercise and moderate exercise, EE: exhaustive exercise alone, Mod: moderate exercise alone, Sed: and sedentary) during GL protocol is essential for inducing hyper-hippocampal glycogen levels, thus, modified GL model, which consisted of several conditions of exercise and conventional diet (61% carbohydrate) was used in this experiment (Fig. 5A). Muscle and liver glycogen levels were unchanged in all exercise regimen groups irrespective of exercise conditions (Exp. 4, Fig. 5B,C). However, glycogen levels in the hippocampus, but not in the hypothalamus or cortex, increased with exhaustive exercise alone (8%) or with a combination of exhaustive and moderate exercise (10%) (P < 0.05; Exp. 4, Fig. 5D), implying that exhaustive exercise alone is sufficient to induce hyper-glycogen levels in the hippocampus with the possibility that this is a hippocampal-specific novel innovation of GL for the brain.
Figure 5.

Exhaustive, but not moderate, exercise in GL icreasesd hippocampal glycogen with conventional diet. (A) Experimental design. (B) Muscle glycogen. (C) Liver glycogen. (D) Hippocampal, hypothalamic and cortical glycogen. Data are expressed as mean ± standard error (n = 5–10/group). *P < 0.05; **P < 0.01 versus sedentary group (Dunnett’s post hoc test).
Discussion
In the present study, we aimed to clarify whether GL or one or both of its two main components, EX (exhaustive exercise followed by moderate exercise and rest) and HCD (70% high carbohydrates diet), have positive effects on brain glycogen storage. We first tested the hypothesis that 1-week GL model of previous trials, EX with HCD6,8, woud have more impact on glycogen storage in some brain regions in a similar manner as in muscle, and, if so, we sought to better characterize the new GL effect of inducing hyper-glycogen storage in the brain in terms of the essential components of the GL using the GL protocol (see experimental protocol in Fig. 1). As a result, we have demonstrated that GL increases brain glycogen, at least in the hippocampus and hypothalamus with concomitant muscle glycogen accumulation. Further, the hyper-glycogen levels induced in the hippocampus by the GL are not dependent on carbohydrate intake during the GL, but are dependent on exhaustive exercise, a one of the EX components, implying the physiological significance of EX in enhancing brain glycogen synthesis.
One week after the GL, we found that brain glycogen levels had increased significantly in the hippocampus and hypothalamus as well as in muscle (pre-post muscle glycogen increase rate in ours vs. previous, 79% vs. 24%) (Exp. 1, Fig. 2B and C), which is consistent with previous studies in humans and rats5–8. These results showed that not only the validity of the current GL model for muscle glycogen storage, but also newly observed positive effects on glycogen storage in the brain, particularly in the hippocampus and hypothalamus (Exp. 1, Fig. 2C). Previous studies have shown that brain glycogen decreases with severe physiological conditions but it does not reduce after exposure to milder stimuli (e.g. fasting for 24 hours, sleep deprivation for over 12 hours, exhaustive exercise for 2 hours, etc.)17,26,27. In particular, Matsui et al.17 showed that 30 minutes of moderate exercise, which is the same condition with moderate exercise (day 2 to day 4) of GL, does not decrease brain glycogen levels. Thus, the combination of HCD and EX might contribute to the GL-induced hyper-glycogen levels in the hippocampus and hypothalamus as well as in the muscle. Furthermore, we examined the dynamics of muscle glycogen levels during GL (Fig. S1A) and confirmed increased glycogen at day 5–7 compared to pre-GL (P < 0.01) (Fig. S1). The GL-induced peak and timing of glycogen increase in the muscle are also consistent with previous studies5–8, indicating the physiological validity of our rat model mimicking human GL. These findings suggest the potential application of GL-induced hyper-hippocampal glycogen to enhance not only the endurance capacity but also the cognitive function of athletes and normal subjects.
Next, we examined which GL component is more dominant in inducing hyper-glycogen stores, the HCD or EX component, in Exp. 2 and 3. First, we examined the role of the HCD component in GL-induced hyper-brain glycogen storage in the hippocampus. In Exp. 2 (Fig. 3), food intake, total calorie intake during the GL were unchanged in the three groups with different amounts of carbohydrates in their diets (5%, 35%, and 70%). Furthermore, there was no significant difference between body weight and blood glucose among the three diet groups. Therefore, it is unlikely that insulin resistance occurred through the 5%-carbohydrate (high fat) diet. In these groups only the total carbohydrate intake increased, dependent on the percentage of carbohydrates in their respective diet (P < 0.001) (Table 3), and the results showed that the hyper-glycogen stores in all tissues of the three groups were not impacted (Fig. 3B). Furthermore, glycogen levels in peripheral tissues (liver and muscle) depended upon and had a positive correlation with the amount of carbohydrate intake, whereas brain glycogen levels did not, suggesting that hyper-glycogen levels in the brain (hippocampus and hypothalamus) occurred independently of carbohydrate intake (Fig. 3C). Regarding the liver, there was a negative correlation between fat intake and liver glycogen. However, the recovery of liver glycogen levels after exercise, which is the basis of GL, is strongly affected by carbohydrate intake28, furthermore, there is a strong positive correlation between carbohydrate intake and liver glycogen levels in this experiment 2 (Fig. 3C), suggesting the importance of carbohydrate intake increasing glycogen levels in liver.
As for a EX, we examined the effects of EX component in GL-induced hyper-hippocampal glycogen storage and found that EX with a HCD showed a significant hyper-hippocampal glycogen storage rather than Sed with HCD in both muscle and brain tissues (hippocampus and hypothalamus) (Fig. 4B and D) as well as pre-post changes in Exp. 1 (Fig. 2B). The present study provided a new hypothesis that EX is more dominant than HCD in terms of hyper- hippocampal glycogen stores. We further examined the effects of EX combined with a conventional diet (61% carbohydrates) and found that exhaustive exercise (EE) with and without moderate exercise (Mod) (EE + Mod group: exhaustive exercise followed by moderate exercise and rest, EE group: exhaustive exercise followed by rest) resulted in similar hyper-glycogen stores only in the hippocampus (Exp. 4, Fig. 5D). The Mod (moderate exercise followed by sedentary) and Sed (sedentary) conditions produced no effects on hyper-glycogen storage in the hippocampus (Exp. 4, Fig. 5D). Thus, the implementation of exhaustive exercise itself is a crucial factor in long-term (one week) hyper-glycogen storage in the hippocampus. In our previous study, supercompensation occurs rapidly in brain glycogen16: within 6 hours after exhaustive exercise; the rates of supercompensation peak were 29–60% in the brain (whole brain: 46%, cortex: 60%, hippocampus: 33%, hypothalamus: 29%, etc.), and 46% in the plantaris muscle at 24 h after exhaustive exercise. Additionally, significant increases remained at 24 h after exercise in the hippocampus and cortex16. While the mechanism remains unknown, the hyper- glycogen stores in the hippocampus over one week might be in part due to the first exhaustive exercise-induced supercompensation. Finally, EX with HCD showed a significant hyper-glycogen storage rather than Sed with HCD in both muscle and brain tissues (hippocampus and hypothalamus) (Fig. 4B and D) as well as pre-post changes in Exp.1 (Fig. 2B). On the other hand, a conventional diet did not lead to increased muscle glycogen (Exp. 4, Fig. 5B), but a HCD did (Exp. 3, Fig. 4B). These results further suggest the importance of a HCD, not EX alone, in inducing hyper-glycogen levels in muscle, supporting the results that GL induces increased hyper-glycogen levels in muscle in a manner dependent on carbohydrate intake.
We found that 1 week of GL with HCD resulted in hyper-glycogen stores only in the hippocampus (Fig. S1), which were kept at the same levels during last three days of GL, probably due to an altered set-point of brain glycogen synthesis and usage. Although the role and underlying mechanisms of hyper-hippocampal glycogen stores remains unclear, noradrenaline might be involved. Noradrenergic neuron are activated during intense/prolonged acute exercise17,29–32, and its noradrenergic metabolism is associated with an exercise-induced brain glycogen decrease17. Meanwhile, in cultured astrocytes, noradrenaline injection not only stimulates glycogenolysis within 30–60 minutes, but also stimulates its supercompensation via the expression of protein targeting to glycogen (PTG), an activator for glycogen synthase, 4–24 hours after injection33,34. Such the intriguing noradrenergic metabolic dynamics of astrocytic glycogen is consistent with an acute exercise effect17, suggesting a possible role of noradrenaline in the post-exercise hippocampal glycogen supercompensation contributing to the long-term hyper-hippocampal glycogen stores in this study.
What is the role of the hippocampal hyper-glycogen stores? Current studies show that hippocampal glycogen contributes to memory formation via lactate production and transportation21,22, and this is also the case in exercise endurance19. Exploring, with the use of behavioral study21,22, immunohistochemistry for measuring brain glycogen35, and LTP assessment22, whether or not such hyper-glycogen stores enhance memory should be the next step in this assessment.
Our findings provide the first evidence that GL, a combination with HCD and EX, increases brain glycogen, at least in the hippocampus and hypothalamus, with concomitant muscle glycogen deposition. Further, the increase in GL-induced hyper-glycogen levels in the hippocampus does not depend on carbohydrate intake, but does depend on exhaustive exercise, one component of EX (exhaustive exercise followed by moderate exercise and rest), implying the physiological significance of exercise in enhancing brain glycogen synthesis. This new perspective on exercise and brain glycogen will ultimately lead to novel sports/nutrition conditioning for memory function and exercise endurance.
Methods
Materials
All chemicals, including amyloglucosidase, hexokinase, NADP+-dependent glucose-6-phosphate dehydrogenase, NADP+, ATP, EDTA, MgSO4, glucose, glucose-6-phosphate, KOH, imidazole, perchloric acid, and Tris-HCl are from Sigma (St Louis, MO, USA) and Nacalai tesque (Kyoto, Japan).
Animals
Adult male Wistar rats (250–270 g; SLC Inc., Shizuoka, Japan) were housed individually, cared for in an animal facility and fed a Conventional diet (Oriental Yeast Co., Ltd, Ibaraki, Japan) with free access to water from the first week of acclimatization to habituation to treadmill running. The composition of the Conventional diet was 26% protein, 13% fat, and 61% carbohydrates. The room temperature was maintained between 22 and 24 °C under a 12:12 hours light/dark cycle (light on 7:00–19:00). All experimental protocols were approved by the Institutional Animal Care and Use Committee of the University of Tsukuba, and all procedures and methods were performed in accordance with the relevant guidelines laid down by animal ethics committee (Animal ethical approval number; 15–055). Every effort was made to minimize the number of animals used as well as any pain and discomfort.
Habituation to treadmill running
Rats were habituated to running on a treadmill (SN-460, Shinano, Tokyo, Japan) for a total of 5 sessions over 6 days after a 1-week acclimatization period. The running duration was 30 min/day, and the running speed was gradually increased from 5 to 25 m/min16,17,19,36.
Experimental procedures
Experiment 1
The experimental design of Experiment 1 is shown in Fig. 1A and Fig. 2A. Two days after the habituation period, rats underwent 1-week of GL, which consisted of several EX conditions and a HCD. The GL protocol in the present study was a modified version of 1-week muscle GL protocol that has been described previously8. Rats were fed a 70% carbohydrate (HCD) powder diet (Oriental Yeast Co., Ltd, Ibaraki, Japan) during the GL period. The composition of the HCD was 25% protein, 5% fat, and 70% carbohydrates. First, rats were divided randomly into a pre-GL and a post-GL group. As for EX protocol, on day 1 of the GL period, rats initially performed exhaustive exercise (EE): treadmill running at moderate intensity (20 m/min) until exhaustion, which has been determined as 50–70% VO2max for rats. Exhaustion was considered to have occurred when the rat was unable to keep pace with the treadmill, lay flat, and stayed on the grid positioned at the back of the treadmill for a period of 30 seconds despite being gently pushed with sticks or breathed on16,17,19,37. From day 2 to 4, rats performed additional moderate-intensity exercise (Mod) (20 m/min, 30 min/day), and then rats were allowed to rest (Sed) on the treadmill (0 m/min, 30 min) from day 5 to 7. Rats were sacrificed using microwave irradiation on day 0 (Pre) and day 7 (Post) in GL. Following microwave irradiation, five brain loci (the cortex, hippocampus, hypothalamus, cerebellum and brainstem) were collected using a method modified from Hirano et al.38. Skeletal muscle and liver were also collected.
Experiment 2
The experimental design of Experiment 2 is shown in Figs 1B and 3A. Another series of rats divided into two groups (5%, 35%, and 70%) and underwent GL protocols with various percentages of carbohydrates (5%, 35%, and 70% (HCD)) in their diets and EX protocol as for the GL protocol mentioned above. An experimental approach using calorie-controlled diets by adjusting the amount of cellulose powder is standard in the field of nutrition11,39,40, thus, the calories of the three diet groups were unified (Tables 1 and 2). Finally, rats were sacrificed using microwave irradiation on day 7 in GL. Following microwave irradiation the hippocampus and hypothalamus were collected along with the cortex as a negative control for brain regions relating to the cognitive function. These same brain regions were also collected in subsequent experiments as well. Skeletal muscle and liver were also collected.
Experiment 3
The experimental design of Experiment 3 is shown in Figs 1C and 4A. Another series of rats divided into two groups (HCD + Sed, HCD + EX) and underwent GL protocols with HCD and Sedentary or EX protocol as for the modified GL protocol mentioned above. Finally, rats were sacrificed using microwave irradiation on day 7 in GL.
Experiment 4
The experimental design of Experiment 2 is shown in Figs 1D and 5A. Another series of rats were divided into four groups (EE + Mod: a combination of exhaustive exercise and moderate exercise, EE: exhaustive exercise alone, Mod: moderate exercise alone, Sed: sedentary alone) and underwent GL with various exercise conditions and a conventional diet (61% of carbohydrates) for 1 week. Finally, rats were sacrificed using microwave irradiation on day 7 in GL.
Tissue preparation
Rats were anesthetized with isoflurane (a mixture of 30% vol/vol isoflurane in propylene glycol; Dainippon Sumitomo Pharma Co., Ltd., Osaka, Japan) in a bell jar and sacrificed using high-power microwave irradiation (NJE-2603, New Japan Radio Co., Ltd., Tokyo, Japan; 10-kW, 1.2 sec). Following the microwave irradiation, 5 brain loci (hippocampus, cortex, hypothalamus, cerebellum and brainstem) were collected. Muscle (plantaris and/or soleus), liver, and blood samples were also collected. All tissue samples were stored at −80 °C for subsequent biochemical analysis.
Blood glucose, lactate assays
Whole-blood glucose and lactate levels in the whole blood were measured using an automated glucose-lactate analyzer (2300 Stat Plus, Yellow Springs Instruments, USA).
Glycogen assay
The methods of glycogen and glucose measurement were consistent with the method of our previous studies16,17,19. Tissues were homogenized (Polytron, Kinematica, Kriens-Luzern, Switzerland; 4,100 g, 30 seconds, 3 times) in ice-cold 6% perchloric acid (PCA) containing 1 mM EDTA. For tissue glycogen content measurements, glycogen in 100 µl of homogenate was hydrolyzed to glucose by incubating for 3 hours at 37 °C with 1 ml of 0.2 M sodium acetate, 20 µl of 1.0 M KHCO3, and 20 U/ml of amyloglucosidase. To stop the subsequent enzymatic reaction, 0.5 ml of PCA was added. After centrifugation (14,000 g, 10 minutes, 4 °C), and supernatants were neutralized with a KOH solution, consisting of 3 M KOH, 0.3 M imidazole and 0.4 M KCl. The supernatants were then centrifuged (16,000 g, 10 minutes, 4 °C) and measured for glucose content. To measure endogenous glucose levels, non-hydrolyzed samples were obtained by centrifuging homogenates (14,000 g, 10 minutes, 4 °C) and the pH of the supernatants was controlled to a final pH of 6–8 with KOH solution. Neutralized samples were mixed thoroughly, centrifuged (16,000 g, 10 minutes 4 °C), and measured for endogenous glucose levels. The glucose content measurement was performed in 96-well plates using a coupled enzyme assay method. A total of 200 µl of a reaction solution including 50 mM Tris-HCl (pH 8.1), 0.5 mM ATP, 0.5 mM NADP+, 5 mM MgSO4, and 0.1 U/ml glucose-6-phosphate dehydrogenase was added to each well. Then, the plate was placed in the fluorescence plate reader (Arvo, Perkin Elmer, Groningen, Netherlands) and shaken, and measurements of the resultant NADPH were taken at 350 nm excitation and 450 nm emission. The plates were shaken after the addition of hexokinase (0.3U) to each well, and measurements were taken after a 30-min incubation period. Tissue glycogen levels, indicted as glucose units, were calculated by subtracting the final micromolar concentration of glucose per gram of wet weight of the non-hydrolyzed tissue sample from the final micromolar concentration of glucose per gram of wet weight of the hydrolyzed sample.
Statistical analyses
Data are expressed as mean ± standard error (SEM) and were analyzed using Prism 5 (MDF Co., Ltd, Tokyo, Japan). Comparisons of two groups were performed using Student’s t test for unpaired data. Group comparisons were performed using a one-way ANOVA with Dunnett’s post hoc tests. Correlations were calculated using Pearson’s product-moment correlations. Statistical significance is P values < 0.05.
Electronic supplementary material
Acknowledgements
This work was supported in part by special funds of Education and Research of the Ministry of Education, Culture, Sports, Science and Technology (MEXT) granted to the “Human High Performance (HHP) Research Project”; a grant by the Japan Sports Agency for the “Sports Research Innovation Project (SRIP)”; the Japan Society for the Promotion of Science (JSPS) Grant-in-Aid for Challenging Exploratory Research (No. 23650384), Young Scientist A (16H05920), JSPS Fellows (16J05042), and Scientific Research on Innovative Areas (16H06405).
Author Contributions
M.S., T.M. and H.S. designed the study. M.S., T.S. and T.M. collected the data. M.S., T.M. and H.S. performed the analysis. M.S., T.M., S.J., N.O. and H.S. interpreted the data. M.S., T.M., S.J. and H.S. wrote the manuscript. All authors have approved to submit the final manuscript.
Competing Interests
The authors declare that they have no competing interests.
Footnotes
Electronic supplementary material
Supplementary information accompanies this paper at 10.1038/s41598-018-19445-4.
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Takashi Matsui, Email: matsui.takashi.ga@u.tsukuba.ac.jp.
Hideaki Soya, Email: soya.hideaki.gt@u.tsukuba.ac.jp.
References
- 1.Gollnick PDKP, Saltin B. Selective glycogen depletion pattern in human muscle fibers after exercise of varying intensity and at varying pedalling rates. J. Physiol. 1974;241:45–57. doi: 10.1113/jphysiol.1974.sp010639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Hermansen L, Hultman E, Saltin B. Muscle Glycogen during Prolonged Severe Exercise. Acta Physiol. Scand. 1967;71:129–139. doi: 10.1111/j.1748-1716.1967.tb03719.x. [DOI] [PubMed] [Google Scholar]
- 3.Ahlborg B, Bergstrom J, Ekelund L, Hultman E. Muscle glycogen and msucle electrolytes during prolonged phyiscal exercise. Acta Physiol. Scand. 1967;70:129–142. doi: 10.1111/j.1748-1716.1967.tb03608.x. [DOI] [Google Scholar]
- 4.Bergström J, Hermansen L, Hultman E, Saltin B. Diet, Muscle Glycogen and Physical Performance. Acta Physiol. Scand. 1967;71:140–150. doi: 10.1111/j.1748-1716.1967.tb03720.x. [DOI] [PubMed] [Google Scholar]
- 5.Costill DL. Carbohydrates for Exercise: Dietary Demands for Optimal Performance. Int. J. Sports Med. 1988;9:1–18. doi: 10.1055/s-2007-1024971. [DOI] [PubMed] [Google Scholar]
- 6.Pitsiladis YP, Maughan RJ. The effects of exercise and diet manipulation on the capacity to perform prolonged exercise in the heat and in the cold in trained humans. J. Physiol. 1999;517:919–930. doi: 10.1111/j.1469-7793.1999.0919s.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sherman W, Costill D, Fink W, Miller J. Effect of Exercise-Diet Manipulation on Muscle Glycogen and Its Subsequent Utilization During Performance. Int. J. Sports Med. 1981;2:114–118. doi: 10.1055/s-2008-1034594. [DOI] [PubMed] [Google Scholar]
- 8.Shinohara A, Takakura J, Yamane A, Suzuki M. Effect of the classic 1-week glycogen-loading regimen on fat-loading in rats and humans. J. Nutr. Sci. Vitaminol. (Tokyo). 2010;56:299–304. doi: 10.3177/jnsv.56.299. [DOI] [PubMed] [Google Scholar]
- 9.Bergström J, Hultman E. Muscle glycogen synthesis after exercise: an enhancing factor localized to the muscle cells in man. Nature. 1966;210:309–310. doi: 10.1038/210309a0. [DOI] [PubMed] [Google Scholar]
- 10.Berardi JM, Price TB, Noreen EE, Lemon PWR. Postexercise muscle glycogen recovery enhanced with a carbohydrate-protein supplement. Med. Sci. Sports Exerc. 2006;38:1106–1113. doi: 10.1249/01.mss.0000222826.49358.f3. [DOI] [PubMed] [Google Scholar]
- 11.Ivy JL, et al. Early postexercise muscle glycogen recovery is enhanced with a carbohydrate-protein supplement. J. Appl. Physiol. 2002;93:1337–1344. doi: 10.1152/japplphysiol.00394.2002. [DOI] [PubMed] [Google Scholar]
- 12.Åstrand, P. Nutrition and Physical Activity. Nutr. Phys. Act. (1967).
- 13.Sharman IM. Glycogen loading: advantages but possible disadvantages. Br. J. Sports Med. 1981;15:64–67. doi: 10.1136/bjsm.15.1.64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kiens B. Diet and training in the week before competition. Can. J. Appl. Physiol. 2001;26(Suppl):S56–S63. doi: 10.1139/h2001-042. [DOI] [PubMed] [Google Scholar]
- 15.Burke LM, van Loon LJC, Hawley JA. Postexercise muscle glycogen resynthesis in humans. J. Appl. Physiol. 2017;122:1055–1067. doi: 10.1152/japplphysiol.00860.2016. [DOI] [PubMed] [Google Scholar]
- 16.Matsui T, et al. Brain glycogen supercompensation following exhaustive exercise. J. Physiol. 2012;590:607–616. doi: 10.1113/jphysiol.2011.217919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Matsui T, et al. Brain glycogen decreases during prolonged exercise. J. Physiol. 2011;589:3383–3393. doi: 10.1113/jphysiol.2011.203570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Benarroch EE. Glycogen metabolism: Metabolic coupling between astrocytes and neurons. Neurology. 2010;74:919–923. doi: 10.1212/WNL.0b013e3181d3e44b. [DOI] [PubMed] [Google Scholar]
- 19.Matsui T, et al. Astrocytic glycogen-derived lactate fuels the brain during exhaustive exercise to maintain endurance capacity. Proc. Natl. Acad. Sci. 2017;114:6358–6363. doi: 10.1073/pnas.1702739114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Yang J, et al. Lactate promotes plasticity gene expression by potentiating NMDA signaling in neurons. Proc. Natl. Acad. Sci. USA. 2014;111:12228–12233. doi: 10.1073/pnas.1322912111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Newman LA, Korol DL, Gold PE. Lactate Produced by Glycogenolysis in Astrocytes Regulates Memory Processing. PLoS One. 2011;6:e28427. doi: 10.1371/journal.pone.0028427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Suzuki A, et al. Astrocyte-neuron lactate transport is required for long-term memory formation. Cell. 2011;144:810–823. doi: 10.1016/j.cell.2011.02.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Shima T, et al. Moderate exercise ameliorates dysregulated hippocampal glycometabolism and memory function in a rat model of type 2 diabetes. Diabetologia. 2016;60:597–606. doi: 10.1007/s00125-016-4164-4. [DOI] [PubMed] [Google Scholar]
- 24.Duran J, Saez I, Gruart A, Guinovart JJ, Delgado-García JM. Impairment in long-term memory formation and learning-dependent synaptic plasticity in mice lacking glycogen synthase in the brain. J. Cereb. Blood Flow Metab. 2013;33:550–556. doi: 10.1038/jcbfm.2012.200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Suh SW, et al. Astrocyte Glycogen Sustains Neuronal Activity during Hypoglycemia: Studies with the Glycogen Phosphorylase Inhibitor CP-316,819 ([R-R*,S*]-5-Chloro-N-[2-hydroxy-3-(methoxymethylamino)-3-oxo-1-(phenylmethyl)propyl]-1H-indole-2-carboxamide) J. Pharmacol. Exp. Ther. 2007;321:45–50. doi: 10.1124/jpet.106.115550. [DOI] [PubMed] [Google Scholar]
- 26.Garriga J, Cussó R. Effect of starvation on glycogen and glucose metabolism in different areas of the rat brain. Brain Res. 1992;591:277–82. doi: 10.1016/0006-8993(92)91708-M. [DOI] [PubMed] [Google Scholar]
- 27.Kong J, et al. Brain glycogen decreases with increased periods of wakefulness: implications for homeostatic drive to sleep. J. Neurosci. 2002;22:5581–7. doi: 10.1523/JNEUROSCI.22-13-05581.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Saitoh S, Shimomura Y, Suzuki M. Effect of a high-carbohydrate diet intake on muscle glycogen repletion after exercise in rats previously fed a high-fat diet. Eur. J. Appl. Physiol. Occup. Physiol. 1993;66:127–133. doi: 10.1007/BF01427053. [DOI] [PubMed] [Google Scholar]
- 29.Ohiwa N, et al. Activation of A1 and A2 noradrenergic neurons in response to running in the rat. Neurosci. Lett. 2006;395:46–50. doi: 10.1016/j.neulet.2005.10.053. [DOI] [PubMed] [Google Scholar]
- 30.Pagliari R, Peyrin L. Norepinephrine release in the rat frontal cortex under treadmill exercise: a study with microdialysis. J. Appl. Physiol. 1995;78:2121–2130. doi: 10.1152/jappl.1995.78.6.2121. [DOI] [PubMed] [Google Scholar]
- 31.Wang J, Chen X, Zhang N, Ma Q. Effects of exercise on stress-induced changes of norepinephrine and serotonin in rat hippocampus. Chin. J. Physiol. 2013;56:245–252. doi: 10.4077/CJP.2013.BAB097. [DOI] [PubMed] [Google Scholar]
- 32.Kitaoka R, et al. Increased noradrenergic activity in the ventromedial hypothalamus during treadmill running in rats. J. Nutr. Sci. Vitaminol. (Tokyo). 2010;56:185–190. doi: 10.3177/jnsv.56.185. [DOI] [PubMed] [Google Scholar]
- 33.Allaman I, Pellerin L, Magistretti PJ. Protein targeting to glycogen mRNA expression is stimulated by noradrenaline in mouse cortical astrocytes. Glia. 2000;30:382–391. doi: 10.1002/(SICI)1098-1136(200006)30:4<382::AID-GLIA70>3.0.CO;2-0. [DOI] [PubMed] [Google Scholar]
- 34.Sorg O, Magistretti PJ. Vasoactive intestinal peptide and noradrenaline exert long-term control on glycogen levels in astrocytes: blockade by protein synthesis inhibition. J. Neurosci. 1992;12:4923–4931. doi: 10.1523/JNEUROSCI.12-12-04923.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Oe Y, Baba O, Ashida H, Nakamura KC, Hirase H. Glycogen distribution in the microwave-fixed mouse brain reveals heterogeneous astrocytic patterns. Glia. 2016;64:1532–1545. doi: 10.1002/glia.23020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Soya H, et al. Threshold-like pattern of neuronal activation in the hypothalamus during treadmill running: Establishment of a minimum running stress (MRS) rat model. Neurosci. Res. 2007;58:341–348. doi: 10.1016/j.neures.2007.04.004. [DOI] [PubMed] [Google Scholar]
- 37.Hasegawa H, et al. Influence of brain catecholamines on the development of fatigue in exercising rats in the heat. J. Physiol. 2008;586:141–149. doi: 10.1113/jphysiol.2007.142190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hirano M, et al. New protein extraction/solubilization protocol for gel-based proteomics of rat (female) whole brain and brain regions. Mol. Cells. 2006;22:119–125. [PubMed] [Google Scholar]
- 39.Normand J, Thibault L. Effect of Hypercaloric versus Isocaloric Lipid Diet Ration on Diurnal/Nocturnal Eating Pattern in Self-Selecting Rats. J. Clin. Biochem. Nutr. 1993;14:25–35. doi: 10.3164/jcbn.14.25. [DOI] [Google Scholar]
- 40.Luo T, et al. Consumption of Walnuts in Combination with Other Whole Foods Produces Physiologic, Metabolic, and Gene Expression Changes in Obese C57BL/6J High-Fat–Fed Male Mice 1–4. J. Nutr. 2016;146:1641–1650. doi: 10.3945/jn.116.234419. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.