

Abstract
Twin-arginine translocation (Tat) systems transport folded proteins that harbor a conserved arginine pair in their signal peptides. They assemble from hexahelical TatC-type and single-spanning TatA-type proteins. Many Tat systems comprise two functionally diverse, TatA-type proteins, denominated TatA and TatB. Some bacteria in addition express TatE, which thus far has been characterized as a functional surrogate of TatA. For the Tat system of Escherichia coli we demonstrate here that different from TatA but rather like TatB, TatE contacts a Tat signal peptide independently of the proton-motive force and restricts the premature processing of a Tat signal peptide. Furthermore, TatE embarks at the transmembrane helix five of TatC where it becomes so closely spaced to TatB that both proteins can be covalently linked by a zero-space cross-linker. Our results suggest that in addition to TatB and TatC, TatE is a further component of the Tat substrate receptor complex. Consistent with TatE being an autonomous TatAB-type protein, a bioinformatics analysis revealed a relatively broad distribution of the tatE gene in bacterial phyla and highlighted unique protein sequence features of TatE orthologs.
Introduction
Bacteria, archaea, and plant chloroplasts have the capability to transport precursor proteins in a folded state across membranes. Precursor proteins that qualify for this mode of transport are primarily distinguished by an SRRxFLK sequence motif in the N-terminal part of their signal sequences. This consensus motif, of which the double arginine is largely invariant, is recognized by so-called Tat translocases (Tat stands for twin-arginine translocation). A second determinant for the specific recognition of signal peptides by Tat translocases is the hydrophobicity of their core region1,2.
Tat translocases do not pre-exist in the membrane as stable protein complexes but rather assemble on demand3,4 from TatA-type and TatC-type membrane proteins. The TatC subunits of Tat translocases possess six transmembrane helices5,6. They associate with a varying number of homologous TatA-type proteins, whose N-terminal structure is characterized by a single transmembrane helix followed by an amphipathic domain7–10. Our model organism, the Gram-negative bacterium Escherichia coli, expresses three TatA-type proteins, TatA, TatB and TatE. As per degree of homology, TatA and TatB evolved early from a common ancestor whilst TatE emerged from TatA by a late gene duplication event.
TatC functions as the primary docking site for the Tat signal peptide at the Tat translocase11,12 directly recognizing the RR-consensus motif13. TatC provides binding sites for TatA and TatB12,14–16. In concert with TatB, TatC forms a dome-shaped intramembrane binding cavity allowing the hairpin-like insertion of the Tat signal peptide14,17.
Despite their structural homology, TatA and TatB have distinct, non- interchangeable functions during Tat-dependent translocation. By interacting with two different sites on TatC, TatB links neighboring TatC monomers thereby forming circular, hetero-multimeric substrate receptor complexes14,15. TatB interacts with the Tat signal peptide downstream of the RR-consensus motif in a proton-motive force independent manner11,14,18. In addition, it functions as a major binding platform for the folded mature domain of Tat substrates19.
TatA is more abundant than all the other Tat subunits with the actual stoichiometry, however, being at issue20–22. Depending on the proton-motive force, TatA associates with the TatBC complex23 and the signal peptide14,24. It facilitates through its homo-oligomerization25,26 the transmembrane translocation of the substrate in a still elusive manner. TatA competes with TatB for binding to TatC14–16.
The function of the smallest TatA-type protein of E. coli, TatE, remains to be elucidated. In vivo studies demonstrated that TatE can maintain Tat transport in the absence of a functional TatA3,27,28 suggesting TatE be a functional paralog of TatA. This, however, would be insufficient to explain why TatE has persisted during evolution as an individual isoform. Previously we demonstrated that TatE is a regular constituent of the Tat translocase in E. coli28. We found TatE to interact with TatA, TatB, and TatC and to localize to active Tat translocases in vivo. Here we show that TatE exhibits distinct properties rendering it a functional hybrid between TatA and TatB. Using a bioinformatic approach we demonstrate that tatE genes are more abundant among the bacterial kingdom than anticipated, further emphasizing an individual relevance of this Tat subunit.
Results and Discussion
TatE and TatB of E. coli share functional properties
We previously demonstrated that TatE displays the properties of a constitutive component of the E. coli Tat translocase, as it localizes to functional Tat translocases in living cells and interacts individually with TatA, TatB, and TatC28. However, whereas TatA11,24,29, TatB14,18,30, and TatC11,13,18 have all been shown to come into contact with the RR-signal sequence of Tat substrate proteins, direct interactions of TatE with substrate have not yet been demonstrated. We therefore equipped the model Tat substrate TorA-MalE335 with the photo cross-linker p-benzoyl-phenylalanine (Bpa). In TorA-MalE335, the RR-signal peptide of the natural E. coli Tat substrate TorA (trimethylamine oxide reductase) is fused to a C-terminally truncated version of the periplasmic maltose-binding protein MalE12,17. Figure 1A highlights the four positions within the TorA signal peptide that were individually replaced by Bpa during cell-free transcription/translation, using an amber stop codon-based approach. The in vitro synthesized Bpa variants of TorA-MalE335 (pTMal) were incubated with inside-out inner membrane vesicles, which had been prepared from a derivative of E. coli strain DADE overexpressing various combinations of the plasmid-encoded TatABCE proteins in a ΔtatABCE background. When cross-linking was initiated by irradiation with UV-light, the previously described11,14,18,31 site-specific interactions with TatC and TatB were obtained. Thus, Bpa located at position F14 within the RR-consensus motif of the TorA signal peptide cross-links to TatC (Fig. 1B, lane 4, blue star), whereas Bpa incorporated into the hydrophobic core and the c-region of the TorA signal sequence (cf. Fig. 1A) yields adducts to TatB and TatA (Fig. 1B, lanes 10, 16, 22, green and red stars, respectively). When in this experimental setup membrane vesicles containing TatEBC instead of TatABC were employed (lanes 6, 12, 18, 24), cross-linking of the TorA signal sequence to TatB and TatC persisted (blue and green stars), whereas the TatA cross-links (red stars) were no longer obtained. Instead, lower molecular mass adducts (orange stars) appeared that by size correspond to a cross-link between TorA-MalE335 and TatE. This was confirmed by immuno-precipitation using antibodies directed towards TatE (Fig. 1C, lane 7). When vesicles contained both TatE and TatA, the signal sequence of TorA-MalE335 was found cross-linked to both Tat subunits (Fig. 1D, lane 6). These findings indicate that in the assembled Tat translocase, TatE locates close to the hydrophobic core and c-region of an RR-signal peptide exactly like TatA and TatB do. No competition between TatE and TatA for interacting with the Tat substrate was observed under these experimental conditions.
Figure 1.

TatE interacts with the signal sequence of a Tat substrate. (A) Primary structure of the TorA-signal sequence. The twin-arginine consensus motif, the hydrophobic core and the c-region are indicated. Amino acid residues substituted by the photo-crosslinker p-benzoyl-L-phenylalanine (Bpa) are labeled in red. (B–E) The model Tat substrate TorA-MalE335 (TMal) was in vitro synthesized and radioactively labeled. Bpa was incorporated into the signal sequence at the indicated positions (red). After incubation with inverted inner membrane vesicles containing the indicated Tat components, crosslinking was induced by UV-irradiation (UV). The samples were resolved by 10% SDS-PAGE and analyzed by phosphorimaging. Adducts between the substrate and TatC (blue star), TatB (green star), TatA (magenta stars) and TatE (orange stars) are indicated. (C) Adducts between TMal, TatE and TatB were confirmed by immuno-precipitation using antisera against TatE and TatB. The eight lanes are derived from a single gel after excising a piece between lanes 6 and 7. (D) In vesicles containing TatE alongside TatA, the signal peptide cross-links to both Tat proteins. (E) Dissipation of the proton-motive force by 0.1 mM CCCP abolishes contacts with TatA but not with TatE.
A characteristic feature of the cross-links between RR-signal peptides and TatA is their sensitivity towards dissipation of the H+-motive force11. This is illustrated in Fig. 1E, where the UV-dependent adduct between the TorA-MalE335 precursor and TatA (red star) disappears upon addition of the protonophore cyanide m-chlorophenyl-hydrazone (CCCP), whereas the TatB adducts (green stars) persist in the presence of CCCP (lanes 5 and 6). In contrast to TatA and exactly like TatB, TatE was found cross-linked to TorA-MalE335 regardless of whether CCCP was present or not (lanes 8 and 9, orange stars). The results presented in Fig. 1 therefore reveal a property of TatE that would not be expected, if TatE were a functional homologue solely of TatA. Interaction with a Tat signal sequence independently of the H+-motive force rather is a typical feature of TatB.
We previously reported that in the absence of TatA and TatB, TatC by itself enables RR-signal sequences of Tat substrates to insert into the cytoplasmic membrane of E. coli. Insertion was shown to proceed to the point that RR-signal sequences are recognized by signal peptidase and prematurely cleaved without the actual Tat substrates being translocated17. In this scenario, a typical feature of TatB, which is not shared by TatA, is to prevent this TatC-mediated premature cleavage of the signal peptide. This is addressed in Fig. 2A. When the precursor form of TorA-MalE335 (pTMal) is synthesized in vitro in the presence of membrane vesicles lacking all Tat components (ΔTat), it is completely sensitive towards digestion by proteinase K (compare lanes 1 and 2). In the presence of TatABC-containing vesicles, however, pTMal becomes processed by the signal peptidase of the vesicles to the mature form (lane 3, mTMal), which due to transport into the vesicle lumen is now resistant towards proteinase K (lane 4). Only a minor fraction of uncleaved precursor (pTMal) is translocated under these conditions as indicated by protease resistance (lane 4). Note that the slightly smaller size of the protease-treated pTMal (compare lanes 3 and 4) is the result of proteinase K removing of a few N-terminal amino acids from the membrane-embedded signal peptide of translocated yet non-processed pTMal17.
Figure 2.
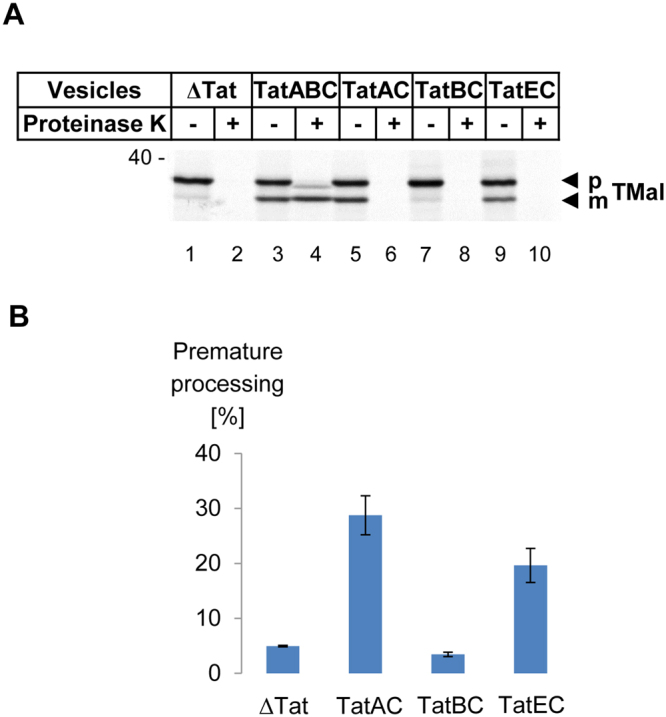
TatE partly prevents premature cleavage of the TorA signal peptide. (A) The Tat substrate TorA-MalE335 (TMal) was synthesized and radioactively labeled in vitro and incubated with inverted inner membrane vesicles obtained from E. coli strain BL21(DE3)Δtat expressing the plasmid-encoded Tat components indicated. The samples were resolved by 10% SDS-PAGE and analyzed by phosphorimaging. In the presence of TatABC-containing vesicles, cleavage of the precursor (pTMal) to the mature protein (mTMal) is accompanied by the acquirement of proteinase K resistance (compare lanes 2 and 4) indicating transport into the lumen of the vesicles. Vesicles lacking TatB cleave pTMal (lane 5) but do not allow transport to occur (lane 6, no protease resistance). TatB (lane 7) and to a lesser extent TatE (lane 9) prevent this premature cleavage. (B) Quantification of premature processing of pTMal to mTMal by the indicated membrane vesicles was obtained from nine independent experiments using vesicles from two different preparations each, except for ΔTat vesicles (three experiments, one preparation).
If in these experimental conditions membrane vesicles are used that contain only TatA and TatC (lane 5), roughly 30% of pTMal are still converted to mTMal (Fig. 2B) similar to what is seen with TatABC vesicles (Fig. 2A, compare lanes 3 and 5). As previously shown, the cleavage of pTMal by TatAC vesicles requires an intact RR-motif, i.e. recognition by TatC, as well as a functional signal peptidase cleavage site17. Although caused by signal peptidase, cleavage of pTMal is premature, because the TatAC vesicles do not allow translocation, as demonstrated by the accessibility of the cleaved mTMal to proteinase K (lane 6). In contrast, TatBC vesicles lacking TatA do not allow for the conversion of pTMal to mTMal (Fig. 2A, lane 7; Fig. 2B). A minimal cleavage of pTMal by TatBC vesicles to a product slightly bigger in size than mTMal (lane 7) was also observed by vesicles entirely lacking the Tat translocase (lane 1) and is therefore caused by an unknown protease. Prevention of premature processing by TatBC vesicles is consistent with TatB and TatC concertedly forming an intramembrane binding cavity for the RR-signal peptide. Because TatA is not a primary constituent of this binding cavity, it is not able to prevent the signal peptide from crossing the membrane and being prematurely processed by signal peptidase14,17. Membrane vesicles harboring only TatE and TatC, however, showed a significantly reduced premature processing of pTMal, although they were not as inhibitory as TatBC vesicles (Fig. 2A, lane 9; Fig. 2B). Thus like TatB and different from TatA, TatE is also able to counteract the TatC-mediated premature cleavage of the Tat substrate, yet with less efficiency than TatB. The data presented in Figs. 1 and 2 collectively suggest that TatE might play a role as part of the TatBC receptor complex for Tat substrates.
TatE paralogs show distinct sequence motifs and occur also outside of Enterobacteria
A TatB-like function of TatE would also be consistent with the about 50-fold lower expression level of tatE compared to tatA20. Nevertheless, TatE of E. coli shows a much higher sequence identity with TatA than with TatB and was shown to partially compensate the phenotype of a tatA deletion mutant under certain experimental conditions3,15,27,28. Given such a seeming bifunctionality and the fact that TatE has been characterized as a constitutive member of the E. coli Tat translocase28, TatE of E. coli seems to be a distinct member of the TatA family rather than a dormant surrogate for TatA or TatB. Consistent with this assumption, the N-terminal amino acid sequences of TatA, TatB, and TatE from E. coli reveal individual differences as manifested by a disparate distribution of charged amino acids (Fig. 3A). TatE and TatB each possess two charged amino acid residues in position 3 (N-tail) and position 8 (transmembrane helix), although the latter one is of opposite polarity (Lys in TatE, Glu, however, in TatB). In contrast, TatA of E. coli is uncharged in its N-terminus.
Figure 3.

Similar to TatB, TatE has conserved N-terminal charges. (A) Sequence alignment of the amino acid sequences of E. coli TatE (accession number: P0A843), TatB (P69425) and TatA (P69428). N-terminal, charged amino acid residues of TatE and TatB are highlighted in red, other charged residues are in bold. Secondary structural elements are assigned according to the NMR structure of TatAd from B. subtilis9 based on the multiple sequence alignment to its E. coli homologues (see Fig. 5B). (B) Consensus sequence of TatE obtained from the alignment of 111 distinct TatE protein sequences encoded by members of the Family of Enterobacterales. The N-terminal charged amino acids are boxed and their degree of conservation is indicated.
In order to investigate the significance of the two N-terminal charged residues of E. coli TatE, we searched the NCBI Reference Sequence protein database for homologous sequences. 3120 sequences were obtained, of which 121 were annotated as TatE, 99 as SecE, 2041 as TatA, and 859 were annotated differently. The sequences annotated as SecE clearly share characteristics with TatE orthologues, whereas SecE is a structurally totally diverse subunit of the functionally unrelated Sec translocase32. It is therefore likely that these 99 TatE homologues have erroneously been annotated as SecE in the data base used. For reasons of unambiguity, we, however, excluded them from our analysis.
Among the 121 TatE sequences, 111 are from the Order of Enterobacterales that contains the Family of Enterobacteriaceae, for which almost exclusively TatE paralogs had hitherto been described. The 10 non-enterobacterial TatE were recovered from the Deltaproteobacterium Bacteriovorax sp. DB6 and the Gammaproteobacteria Halotalea alkalilenta and Gilliamella apicola. A consensus sequence model of the complete 111 enterobacterial TatE sequences is shown in Fig. 3B. Lysine at position 8 was found to be conserved to 99%. The majority (72%) of those TatE sequences display glutamate at position 3 with a preceding Gly. In the remaining 28% of sequences, Glu is found at position 2 and Gly at position 3 resulting in either a Gly-Glu or a Glu-Gly pair at this place. Thus in all enterobacterial TatE sequences, the N-termini are distinguished by an E3xxxxK8 or an E2xxxxxK8 motif.
In line with similar functional properties of TatE and TatB, an N-proximal negatively charged residue, as found here to be conserved among the enterobacterial TatE orthologs, was previously shown to be associated with TatB-like functions. The Freudl group isolated mutants in E. coli tatA that phenotypically suppressed the deletion of TatB. Most of the suppressors had gained an aspartate at the otherwise uncharged N-terminus of TatA and thereby the ability to functionally replace TatB33.
Out of the 3120 sequences obtained from our database search for homologues of the E. coli TatE, 2041 entries were annotated as TatA (Table 1), which reflects the high sequence identity between both Tat proteins15. These TatE homologues are almost entirely of proteobacterial origin (Table 1). When screened for the occurrence of charged N-terminal amino acids, 108 of those 2041 TatA proteins were found to harbor the E3xxxxK8 or E2xxxxxK8 motif, 44 merely a Lys at position 8, whereas 1889 do not carry similar charge patterns in their N-termini. Interestingly, of those bacterial species encoding TatA homologues with the aforementioned charge patterns, 50 possess an additional TatA paralog, of which the N-terminus is not charged. This is exemplified in Fig. 4 for selected species from the Order of Vibrionales, of which both types of TatA paralogs were separately aligned. The one with the higher homology to E. coli TatE and the shorter length (63–78 amino acids) is labeled TatA_1. Seven of them in fact display the E3xxxxK8 motif (boxed), whilst all of them possess the Lys at position 8. The second group of TatA paralogs (denominated TatA_2) lacks any N-terminal charges and consistently contains the Gln8 of E. coli TatA. These findings demonstrate that the occurrence of TatE-type orthologs harboring distinct N-terminal charged amino acids is not limited to enterobacteria but also encompasses other Gamma-proteobacteria. Furthermore, their co-existence in the genomes with TatA-type paralogs, which do not carry TatE-specific N-terminal charge patterns, would support the idea that TatE-type proteins when co-expressed might serve a unique functional purpose.
Table 1.
Listed are the 2041 TatE homologues annotated as TatA.
| Class | Order | Total Hits | N-Charged |
|---|---|---|---|
| Alphaproteobacteria | Rhizobiales | 158 | |
| Sphingomonadales | 25 | ||
| Rhodobacterales | 14 | ||
| others | 2 | ||
| Betaproteobacteria | Burkholderiales | 339 | |
| Neisseriales | 38 | ||
| Rhodocyclales | 26 | ||
| Methylophilales | 17 | ||
| others | 26 | ||
| Gammaproteobacteia | Enterobacterales | 443 | 96 |
| Pseudomonadales | 269 | 8 | |
| Alteromonadales | 153 | ||
| Vibrionales | 138 | 42 | |
| Oceanospirillales | 107 | 1 | |
| Pasteurellales | 41 | ||
| Legionellales | 39 | ||
| Thiotrichales | 27 | ||
| Cellvibrionales | 24 | ||
| Xanthomonadales | 22 | ||
| Aeromonadales | 21 | 2 | |
| Chromatiales | 19 | ||
| Methylococcales | 16 | 3 | |
| others | 33 | ||
| Epsilonproteobacteria | Campylobacterales | 9 | |
| Sulfurovum | 4 | ||
| Deltaproteobacteria | 5 | ||
| All others | 26 | ||
| 2041 | 152 |
The sequences are sorted by the Class of the source strains, the number of sequences found in each Order, and the occurrence of N-terminal charges (including 108 entries with E3xxxxK8 or E2xxxxxK8 motifs and 44 entries with K8).
Figure 4.

Two distinct TatA orthologs of Vibrionales. TatA_1 paralogs (A) feature N-terminal charged residues, which are similar to E. coli TatE (Glu3 and Lys8, boxed), or a single lysine at the 8th position. TatA_2 paralogs (B) carry no charged residues at the corresponding section. The sole polar residue of TatA_2 is Gln8, which is conserved in TatA of E. coli. Listed are selected strains of the Order of Vibrionales with the following accession numbers of their TatA_1 and TatA _2 paralogs, respectively: Photobacterium angustum (WP_005363978, WP_005372432); Photobacterium aquimaris (WP_060996792, WP_060998634); Photobacterium kishitanii (WP_045045764, WP_058120333); Photobacterium leiognathi (WP_008988792, WP_008986002); Photobacterium phosphoreum (WP_045028541, WP_045032465); Photobacterium sp. SKA34 (WP_006644312, WP_006643468); Photobacterium (WP_036791541, WP_036788465); Vibrio cholerae (WP_000508976, WP_000508970/WP_000508978, WP_000508968/WP_000508979, WP_000508969/WP_000508980, WP_000508971).
A TatE paralog had also been described for the Gram-positive organism Corynebacterium glutamicum34. In contrast, the inclusion threshold for TatE homologues that we defined for our database search did not yield any result, neither from C. glutamicum nor from other Gram-positive organisms. We therefore performed a sequence alignment of the TatABE proteins form C. glutamicum and E. coli and found that the annotated TatE of C. glutamicum actually shares a higher sequence similarity with TatA than with TatE from E. coli (Fig. 5A). Moreover, the annotated TatE of C. glutamicum lacks any charged amino acids within its N-terminal region but shares the Gln8 with E. coli TatA (Fig. 5A). However, without further functional analyses, a clear assignment of the two TatA paralogs of C. glutamicum to the TatA and TatE families is difficult to undertake.
Figure 5.

TatE paralogs in Gram-positive bacteria. The degree of sequence similarity, indicated by the percent of identity and the expect e-value (which decreases with the lower chance of random match) between Tat proteins from Escherichia coli and Corynebacterium glutamicum (A) and Bacillus subtilis (B), respectively, were calculated using NCBI BLAST (upper panels). (A) The closest homologue of C. glutamicum TatE is TatA from E. coli, both featuring non-charged N-termini as revealed in the multiple sequence alignment (lower panel). (B) In B. subtilis, TatAc is most similar to TatE of E coli, as shown by the common ExxxxK motif. Sequence source: C. glutamicum TatA (Q8NQE4), TatE (Q2MGW8), TatB (Q8NRD0); B. subtilis TatAy (O05522), TatAc (O31804), TatAd (O31467).
On the other hand, we found the ExxxxK motif characteristic for E. coli TatE also contained in TatAc, which is one of the three annotated TatA paralogs of the Gram-positive bacterium Bacillus subtilis (Fig. 5B). Intriguingly, TatAc was recently shown to functionally compensate for mutation-borne defects of TatAy, which is another TatA paralog of B. subtilis35. TatAc was, however, not able to complement the complete absence of TatAy suggesting that TatAc and TatAy might functionally interact35. Thus the minimal Tat translocases of B. subtilis consisting of single TatC and TatA components36,37 might in fact associate with an auxiliary TatA component. In Fig. 5B we aligned the three TatA paralogs TatAc, TatAy, TatAd of B. subtilis with the protein sequences of E. coli TatA, TatE, and TatB. Strikingly, the sequence alignment of B. subtilis TatAc and E. coli TatE, both exhibiting the ExxxxK motif, gave 45% identical amino acid residues within the aligned parts indicating that TatAc might more likely be a functional paralog of E. coli TatE than of E. coli TatA. Hence, all these sequence compilations demonstrate a wide distribution of TatA-family members that possess a Lys at a position equivalent to the 8th position of enterobacaterial TatE (Fig. 3B). This suggests that unique TatE-type paralogs of TatA might operate in a much larger number of bacterial Tat translocases than previously appreciated.
TatE-specific interactions with the Tat proteins
Following an analysis of co-evolutionarily predicted residue contacts between TatC and TatA-family proteins as well as the results of molecular dynamics simulations, it was recently proposed that Glu8, which is conserved among almost all TatB orthologs, is part of a polar cluster that ligates TatB to TatC in a functional manner15. Similarly, Gln or His that populate residue 8 in the vast majority of TatA paralogs would allow TatA to interact with TatC via the same polar cluster15. In contrast, the Lys8 residue, which obviously is a hallmark of TatE-type orthologs, is likely to mediate different contacts with TatC. This is strongly suggested by the phenotype of an E. coli TatB variant, which due to a Glu8 to Lys8 substitution associates with TatC in a manner that allows proficient recognition of otherwise transport-defective signal peptides38–40. By inference, TatA-family members naturally displaying a positively charged residue at position 8, such as most TatE paralogs, are also likely to interact with TatC in a manner different from the mostly negatively charged TatB orthologs. Association of TatE paralogs with TatC could even represent an advanced step in the assembly of a functional Tat translocase following the initial TatBC interaction.
In order to obtain more information on how TatE might associate with the other Tat proteins, we explored N,N’-dicyclohexylcarbodiimide (DCCD) as a cross-linking agent. DCCD is known to form amide bonds between carboxyl groups located in hydrophobic environments and primary amines41. We recently realized that TatB and TatC present in inner membrane vesicles of E. coli can be cross-linked in this way (unpublished results). As shown in Fig. 6A, dependent on the addition of DCCD to membrane vesicles containing TatABC a ~45 kDa product appears (green diamond), which is recognized by anti-TatB and anti-TatC antibodies (αTatB, lane 2; αTatC, lane 8). If the vesicles contain TatE instead of TatA (TatEBC), DCCD treatment results in an additional cross-link, ~37 kDa in size, which is recognized by anti-TatB and anti-TatE antibodies (lane 4, orange triangle, αTatB, αTatE). We therefore conclude that in these vesicles, TatE is so closely spaced to TatB that both proteins can be cross-linked by DCCD. This is consistent with previously identified contact sites between TatE and TatB, which photo cross-linking revealed in the transmembrane and amphipathic helices of TatE28. An immediate proximity between TatE and TatB within the substrate receptor complex is further reflected by the findings of Fig. 1 demonstrating that in the absence of the H+-motive force, the same sites of the TorA signal peptide contact both TatB and TatE.
Figure 6.

TatE-specific interactions with TatA, TatB and TatC. Western blot analysis of crosslinking experiments with inverted inner membrane vesicles containing the indicated Tat subunits (TatABC, EBC, EABC). After crosslinking, the samples were resolved using 9% Tricine-SDS-PAGE and detected by immunoblotting using antibodies against TatB (αTatB), TatC (αTatC), TatE (αTatE) and TatA (αTatA). (A) Crosslinking using N,N′-dicyclohexylcarbodiimide (DCCD). After incubation with DCCD higher molecular bands representing TatB-TatC adducts (green diamond), TatE-TatB adducts (orange triangle), and TatE-TatA adducts (orange dots) were detected. Indicated are the positions corresponding in size to TatC dimers (blue star), TatE dimers (orange star) and TatA oligomers (magenta stars). Lanes are derived from four individual gels. The gaps mark the excision of single unrelated lanes. (B) Crosslinking using p-benzoyl-L-phenylalanine (Bpa). During vesicle preparation the crosslinker was incorporated into TatC at position 205 (Met205Bpa). Bpa-crosslinking was induced by exposing the vesicles to UV irradiation. UV-dependent higher molecular bands were identified via their immuno-reactivities as TatC oligomers (blue star), TatB-TatC adducts (blue diamond), TatC-TatA adducts (magenta square) and TatC-TatE adducts (orange square).
We next asked where on TatC such a TatB-TatE heterodimer could be located. As to TatB, several recent studies have identified the transmembrane helix 5 of TatC as a docking site for the transmembrane helix of TatB5,14,15,42. One of the residues located in transmembrane helix 5 of TatC that repeatedly was shown to mediate contacts with TatB is methionine 205. We therefore incorporated Bpa into E. coli TatC at position 205 using an in vivo amber stop codon approach and prepared membrane vesicles that carried the TatC205Bpa mutant in the presence of TatAB and TatEB (Fig. 6B). In the context of the TatAB proteins, TatC205Bpa when activated by UV light yielded four prominent adducts (lane 2, αTatC). Adducts running at ~65 and ~40 kDa (blue stars) were recognized only by antibodies directed against TatC and therefore represent dimers and trimers of TatC14,42. The ~50 kDa cross-linking product (blue diamond) was also recognized by anti-TatB antibodies (αTatB, lane 6) confirming this known contact site of TatC for TatB. The ~37 kDa adduct of TatC205Bpa (red square) was also detected by anti-TatA antibodies (αTatA, lane 6) consistent with the previously established overlap of the TatA and TatB cross-linking sites on transmembrane helix 5 of TatC14,15,43. In vesicles containing TatC205Bpa together with TatEB instead of TatAB, the UV-dependent cross-links were weaker owing to a lower TatC content of those vesicles (αTatC, compare lanes 2 and 4). As expected, the TatA adduct (red square) was no longer visible. Instead a new ~30 kDa cross-link of TatC205Bpa to TatE appeared (orange square in αTatC, αTatE, lane 4), indicating that TatE, exactly as TatB and TatA, contacts TatC via its fifth transmembrane helix. Previously it was found that Bpa when incorporated into TatE at position 9 of its transmembrane helix cross-links to TatC28. It is therefore reasonable to assume that the contact of TatE with residue 205 in helix five of TatC reflects the alignment of both transmembrane helices as demonstrated before for TatB and TatA5,24,42. However, as discussed above the oppositely charged transmembrane helices of TatE and TatB are not likely to occupy exactly the same binding sites on TatC.
Interestingly, the DCCD-mediated contact between TatB and TatE (Fig. 6A, lane 4, orange triangle), was almost gone in the presence of TatA (lane 6). Instead membrane vesicles containing all four Tat components gave rise to new adducts that by size and immuno-reactivity represent TatE-TatA oligomers (orange dots, compare lanes 4 and 6, 10 and 12). If this reflects a true exchange of TatB for TatA as the binding partner of TatE following the recruitment of TatA to the substrate-bound TatBC receptor complex, is speculative at this point of time. The DCCD-mediated cross-links that we obtained between TatE and TatA, however, suggest that TatE could fulfil a role in the recruitment of TatA to the TatBC receptor complex through hetero-oligomerization with TatA. Because in vivo, TatE is present only in sub-stoichiometric amounts compared with TatA20, TatE could conceivably function as a nucleation point for a TatBC-dependent oligomerization event of TatA.
In conclusion, our results suggest that TatE paralogs have overlapping functions between TatA and TatB. Similar to TatB, TatE is involved in substrate binding yet possibly at a later step. Although it docks at transmembrane helix five of TatC much like TatA and TatB do, the individual interacting epitopes of TatC are likely to vary. We propose that TatE could play a role in the oligomerization of TatA.
Methods
Plasmids
The plasmids used in this study are listed in Table S1. To construct plasmid pEC, vector pEBC_LinkRBS was used as a template. Primers flanking tatB were designed (pECKI for and rev, Table S1) and phosphorylated. The tatB gene was deleted during vector amplification. The PCR product was purified and ligated prior to transformation. Plasmid pEBC_LinkRBS was also used to prepare the amber stop codon mutant at position Met205 in tatC in the context of tatEB using mutagenesis PCR19 and the primers Met205 for and rev listed in Table S1. Construction of the same TatC variant in the context of tatAB was previously described14.
In vitro reactions
The plasmid-encoded Tat substrate TorA-MalE335 and its Bpa variants were synthesized in a cell free transcription/translation system in 50 μl reactions44. Cell extracts were prepared using E. coli strain SL11945 or Top10 (Invitrogen) transformed with plasmid pSup-BpaRS-6TRN(D286R) for Bpa incorporation into TorA-MalE335. The vesicles were added 12 min after starting protein synthesis at 37 °C and incubated for 18 min at 37 °C. In order to disrupt the proton motive force, 0.1 mM CCCP was added at the onset of synthesis. After incubation with vesicles, Bpa crosslinking was induced by UV irradiation for 20 min on ice. To visualize transport of TorA-MalE335 into the vesicles, completed reactions were treated with 0.5 mg/ml Proteinase K.
For immuno-precipitation, the samples were denatured in 1% SDS for 10 min at 95 °C after crosslinking. Antisera against TatA, TatE and TatB were incubated with Protein A-Sepharose beads for 90 min at 4 °C. Denatured samples were cleared by brief centrifugation and applied to the antibody-loaded Protein A-Sepharose during 3 h at 4 °C on a spinning wheel. After 4 washing steps, antibody-bound material was released by incubating in SDS-PAGE loading buffer for 10 min at 37 °C and 1.400 rpm.
The proteins were resolved on SDS-PAGE using 10% polyacrylamide gels. Radioactive gels were developed by phosphorimaging using a Storm 845 instrument. Quantification was performed using ImageQuantTL.
Inverted inner membrane vesicles
Vesicles were prepared according to44 from E. coli strains DADE (MC4100∆tatABCDE)46 or Bl21(DE3)∆Tat (B. Ize and T. Palmer, personal communication) expressing the vectors pBADxTat (TatABC), pEBC_LinkRBS (TatEBC), pEABC_LinkRBS (TatEABC), p8737-tatAC (TatAC), pFAT75CH∆A (TatBC) or pECKI (TatEC), respectively. Vesicles containing TatABCMet205Bpa were prepared from BL21(DE3) E. coli cells transformed with pEVOL-pBpF and p8737, and vesicles containing TatEBCMet205Bpa from DADE cells containing pEVOL-pBpF and pEBC_LinkRBS. Expression of plasmid-encoded genes was induced by 0.1% arabinose or 1 mM IPTG (p8737-tatAC and pFAT75CH∆A). The concentration of the vesicles was adjusted to OD280 ≈ 100.
To detect crosslinks between the Tat subunits, 5 µl vesicles were diluted with 95 µl INV-buffer44. When indicated 1 µl DCCD (50 mM) was added. The samples were exposed to UV light for 20 min on ice, precipitated with trichloroacetic acid and resuspended in 100 µl Tricine sample buffer47. Proteins were then resolved using 9% Tricine SDS-PAGE and identified by Western blotting using antibodies against TatE (10 µl sample per lane), TatA (7 µl per lane), TatB and TatC (20 µl per lane).
Protein sequence analysis
All sequences used in this research were obtained from Uniprot or NCBI Reference Sequence (RefSeq) protein databases. The NCBI RefSeq protein database48 release 79 was queried with the TatE from Escherichia coli using BLASTP49 with cut-off e-value set to 1e-3. After removing incomplete entries, 3120 sequences were obtained. Among them, the 111 enterobacterial homologues were aligned with Clustal Omega50 and the consensus sequence logo was plotted using WebLogo351. The degree of sequence similarity of Tat proteins between E. coli (TatA, TatE, TatB) and C. glutamicum (TatA, TatE, TatB) or B. subtilis (TatAy, TatAc, TatAd) were investigated by pairwise-alignment using NCBI BLAST.
Data availability
All data generated or analysed during this study are included in this published article (and its Supplementary Information files).
Electronic supplementary material
Acknowledgements
This study was supported by SFB746 and the Excellence Initiative (GSC-4, Spemann Graduate School, EXC294 BIOSS to C.H.) of the German Research Foundation. We gratefully acknowledge MuDe Zou for excellent technical assistance.
Author Contributions
Acquisition and analysis of data: E.E., W.C.K., J.F., A.S.B. Conception and design: E.E., W.C.K., J.F., C.H., M.M. Drafting and revision of article: E.E., A.S.B., J.F., W.C.K., C.H., M.M. Final approval: J.F., M.M.
Competing Interests
The authors declare that they have no competing interests.
Footnotes
Electronic supplementary material
Supplementary information accompanies this paper at 10.1038/s41598-018-19640-3.
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Huang, Q. & Palmer, T. Signal Peptide Hydrophobicity Modulates Interaction with the Twin-Arginine Translocase. MBio8 (2017). [DOI] [PMC free article] [PubMed]
- 2.Ulfig A, et al. The h-region of twin-arginine signal peptides supports productive binding of bacterial Tat precursor proteins to the TatBC receptor complex. J Biol Chem. 2017;292:10865–10882. doi: 10.1074/jbc.M117.788950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Alcock F, et al. Live cell imaging shows reversible assembly of the TatA component of the twin-arginine protein transport system. Proc Natl Acad Sci USA. 2013;110:E3650–9. doi: 10.1073/pnas.1306738110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Rose P, Fröbel J, Graumann PL, Müller M. Substrate-dependent assembly of the Tat translocase as observed in live Escherichia coli cells. PLoS One. 2013;8:e69488. doi: 10.1371/journal.pone.0069488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Rollauer SE, et al. Structure of the TatC core of the twin-arginine protein transport system. Nature. 2012;492:210–4. doi: 10.1038/nature11683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ramasamy S, Abrol R, Suloway CJ, Clemons WM., Jr. The glove-like structure of the conserved membrane protein TatC provides insight into signal sequence recognition in twin-arginine translocation. Structure. 2013;21:777–88. doi: 10.1016/j.str.2013.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hu Y, Zhao E, Li H, Xia B, Jin C. Solution NMR structure of the TatA component of the twin-arginine protein transport system from gram-positive bacterium Bacillus subtilis. J Am Chem Soc. 2010;132:15942–15944. doi: 10.1021/ja1053785. [DOI] [PubMed] [Google Scholar]
- 8.Rodriguez F, et al. Structural model for the protein-translocating element of the twin-arginine transport system. Proc Natl Acad Sci USA. 2013;110:E1092–101. doi: 10.1073/pnas.1219486110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Walther TH, Grage SL, Roth N, Ulrich AS. Membrane alignment of the pore-forming component TatA(d) of the twin-arginine translocase from Bacillus subtilis resolved by solid-state NMR spectroscopy. J Am Chem Soc. 2010;132:15945–15956. doi: 10.1021/ja106963s. [DOI] [PubMed] [Google Scholar]
- 10.Zhang Y, Wang L, Hu Y, Jin C. Solution structure of the TatB component of the twin-arginine translocation system. Biochim Biophys Acta. 2014;1838:1881–1888. doi: 10.1016/j.bbamem.2014.03.015. [DOI] [PubMed] [Google Scholar]
- 11.Alami M, et al. Differential interactions between a twin-arginine signal peptide and its translocase in Escherichia coli. Mol Cell. 2003;12:937–946. doi: 10.1016/S1097-2765(03)00398-8. [DOI] [PubMed] [Google Scholar]
- 12.Zoufaly S, et al. Mapping Precursor-binding Site on TatC Subunit of Twin Arginine-specific Protein Translocase by Site-specific Photo Cross-linking. J Biol Chem. 2012;287:13430–13441. doi: 10.1074/jbc.M112.343798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ma X, Cline K. Mapping the signal peptide binding and oligomer contact sites of the core subunit of the pea twin arginine protein translocase. Plant Cell. 2013;25:999–1015. doi: 10.1105/tpc.112.107409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Blümmel AS, Haag LA, Eimer E, Müller M, Fröbel J. Initial assembly steps of a translocase for folded proteins. Nat Commun. 2015;6:7234. doi: 10.1038/ncomms8234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Alcock, F. et al. Assembling the Tat protein translocase. Elife5 (2016). [DOI] [PMC free article] [PubMed]
- 16.Habersetzer, J. et al. Substrate-triggered position switching of TatA and TatB during Tat transport in Escherichia coli. Open Biol7 (2017). [DOI] [PMC free article] [PubMed]
- 17.Fröbel J, et al. Transmembrane insertion of twin-arginine signal peptides is driven by TatC and regulated by TatB. Nat Commun. 2012;3:1311. doi: 10.1038/ncomms2308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gerard F, Cline K. Efficient Twin Arginine Translocation (Tat) Pathway Transport of a Precursor Protein Covalently Anchored to Its Initial cpTatC Binding Site. J Biol Chem. 2006;281:6130–6135. doi: 10.1074/jbc.M512733200. [DOI] [PubMed] [Google Scholar]
- 19.Maurer C, Panahandeh S, Jungkamp AC, Moser M, Müller M. TatB functions as an oligomeric binding site for folded Tat precursor proteins. Mol Biol Cell. 2010;21:4151–61. doi: 10.1091/mbc.E10-07-0585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Jack RL, Sargent F, Berks BC, Sawers G, Palmer T. Constitutive expression of Escherichia coli tat genes indicates an important role for the twin-arginine translocase during aerobic and anaerobic growth. J Bacteriol. 2001;183:1801–1804. doi: 10.1128/JB.183.5.1801-1804.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Celedon JM, Cline K. Stoichiometry for binding and transport by the twin arginine translocation system. J Cell Biol. 2012;197:523–34. doi: 10.1083/jcb.201201096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hauer RS, et al. Enough is enough: TatA demand during Tat-dependent protein transport. Biochim Biophys Acta. 2013;1833:957–965. doi: 10.1016/j.bbamcr.2013.01.030. [DOI] [PubMed] [Google Scholar]
- 23.Mori H, Cline K. A twin arginine signal peptide and the pH gradient trigger reversible assembly of the thylakoid [Delta]pH/Tat translocase. J Cell Biol. 2002;157:205–10. doi: 10.1083/jcb.200202048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Fröbel J, Rose P, Müller M. Early contacts between substrate proteins and TatA translocase component in twin-arginine translocation. J Biol Chem. 2011;286:43679–43689. doi: 10.1074/jbc.M111.292565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Dabney-Smith C, Cline K. Clustering of C-terminal stromal domains of Tha4 homo-oligomers during translocation by the Tat protein transport system. Mol Biol Cell. 2009;20:2060–2069. doi: 10.1091/mbc.E08-12-1189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Dabney-Smith C, Mori H, Cline K. Oligomers of Tha4 organize at the thylakoid Tat translocase during protein transport. J Biol Chem. 2006;281:5476–5483. doi: 10.1074/jbc.M512453200. [DOI] [PubMed] [Google Scholar]
- 27.Sargent F, et al. Overlapping functions of components of a bacterial Sec-independent protein export pathway. EMBO J. 1998;17:3640–3650. doi: 10.1093/emboj/17.13.3640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Eimer E, Fröbel J, Blümmel AS, Müller M. TatE as a Regular Constituent of Bacterial Twin-arginine Protein Translocases. J Biol Chem. 2015;290:29281–29289. doi: 10.1074/jbc.M115.696005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Taubert J, et al. TatBC-independent TatA/Tat substrate interactions contribute to transport efficiency. PLoS One. 2015;10:e0119761. doi: 10.1371/journal.pone.0119761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Taubert J, Bruser T. Twin-arginine translocation-arresting protein regions contact TatA and TatB. Biol Chem. 2014;395:827–36. doi: 10.1515/hsz-2014-0170. [DOI] [PubMed] [Google Scholar]
- 31.Holzapfel E, et al. The entire N-terminal half of TatC is involved in twin-arginine precursor binding. Biochemistry. 2007;46:2892–2898. doi: 10.1021/bi062205b. [DOI] [PubMed] [Google Scholar]
- 32.Kudva R, et al. Protein translocation across the inner membrane of Gram-negative bacteria: the Sec and Tat dependent protein transport pathways. Res Microbiol. 2013;164:505–534. doi: 10.1016/j.resmic.2013.03.016. [DOI] [PubMed] [Google Scholar]
- 33.Blaudeck N, Kreutzenbeck P, Müller M, Sprenger GA, Freudl R. Isolation and characterization of bifunctional Escherichia coli TatA mutant proteins that allow efficient Tat-dependent protein translocation in the absence of TatB. J Biol Chem. 2005;280:3426–3432. doi: 10.1074/jbc.M411210200. [DOI] [PubMed] [Google Scholar]
- 34.Kikuchi Y, Date M, Itaya H, Matsui K, Wu LF. Functional analysis of the twin-arginine translocation pathway in Corynebacterium glutamicum ATCC 13869. Appl Environ Microbiol. 2006;72:7183–92. doi: 10.1128/AEM.01528-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Goosens VJ, De-San-Eustaquio-Campillo A, Carballido-Lopez R, van Dijl JM. A Tat menage a trois–The role of Bacillus subtilis TatAc in twin-arginine protein translocation. Biochim Biophys Acta. 2015;1853:2745–53. doi: 10.1016/j.bbamcr.2015.07.022. [DOI] [PubMed] [Google Scholar]
- 36.Goosens VJ, Monteferrante CG, van Dijl JM. The Tat system of Gram-positive bacteria. Biochim Biophys Acta. 2014;1843:1698–706. doi: 10.1016/j.bbamcr.2013.10.008. [DOI] [PubMed] [Google Scholar]
- 37.Monteferrante CG, Baglieri J, Robinson C, van Dijl JM. TatAc, the third TatA subunit of Bacillus subtilis, can form active twin-arginine translocases with the TatCd and TatCy subunits. Appl Environ Microbiol. 2012;78:4999–5001. doi: 10.1128/AEM.01108-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Kreutzenbeck P, et al. Escherichia coli twin arginine (Tat) mutant translocases possessing relaxed signal peptide recognition specificities. J Biol Chem. 2007;282:7903–7911. doi: 10.1074/jbc.M610126200. [DOI] [PubMed] [Google Scholar]
- 39.Lausberg F, et al. Genetic evidence for a tight cooperation of TatB and TatC during productive recognition of twin-arginine (Tat) signal peptides in Escherichia coli. PLoS One. 2012;7:e39867. doi: 10.1371/journal.pone.0039867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Huang Q, et al. A signal sequence suppressor mutant that stabilizes an assembled state of the twin arginine translocase. Proc Natl Acad Sci USA. 2017;114:E1958–E1967. doi: 10.1073/pnas.1615056114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Valeur E, Bradley M. Amide bond formation: beyond the myth of coupling reagents. Chem Soc Rev. 2009;38:606–31. doi: 10.1039/B701677H. [DOI] [PubMed] [Google Scholar]
- 42.Kneuper H, et al. Molecular dissection of TatC defines critical regions essential for protein transport and a TatB-TatC contact site. Mol Microbiol. 2012;85:945–961. doi: 10.1111/j.1365-2958.2012.08151.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Aldridge C, Ma X, Gerard F, Cline K. Substrate-gated docking of pore subunit Tha4 in the TatC cavity initiates Tat translocase assembly. J Cell Biol. 2014;205:51–65. doi: 10.1083/jcb.201311057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Moser M, Panahandeh S, Holzapfel E, Müller M. In vitro analysis of the bacterial twin-arginine-dependent protein export. Methods Mol Biol. 2007;390:63–80. doi: 10.1007/978-1-59745-466-7_5. [DOI] [PubMed] [Google Scholar]
- 45.Lesley SA, Brow MA, Burgess RR. Use of in vitro protein synthesis from polymerase chain reaction-generated templates to study interaction of Escherichia coli transcription factors with core RNA polymerase and for epitope mapping of monoclonal antibodies. J Biol Chem. 1991;266:2632–2638. [PubMed] [Google Scholar]
- 46.Wexler M, et al. TatD is a cytoplasmic protein with DNase activity. No requirement for TatD family proteins in Sec-independent protein export. J Biol Chem. 2000;275:16717–16722. doi: 10.1074/jbc.M000800200. [DOI] [PubMed] [Google Scholar]
- 47.Schagger H, von Jagow G. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal Biochem. 1987;166:368–79. doi: 10.1016/0003-2697(87)90587-2. [DOI] [PubMed] [Google Scholar]
- 48.Pruitt KD, Tatusova T, Maglott DR. NCBI reference sequences (RefSeq): a curated non-redundant sequence database of genomes, transcripts and proteins. Nucleic Acids Res. 2007;35:D61–5. doi: 10.1093/nar/gkl842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Altschul SF, et al. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 1997;25:3389–402. doi: 10.1093/nar/25.17.3389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Sievers F, et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol Syst Biol. 2011;7:539. doi: 10.1038/msb.2011.75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Crooks GE, Hon G, Chandonia JM, Brenner SE. WebLogo: a sequence logo generator. Genome Res. 2004;14:1188–90. doi: 10.1101/gr.849004. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
All data generated or analysed during this study are included in this published article (and its Supplementary Information files).