

Abstract
The role of long non-coding RNAs (lncRNAs) in the carcinogenesis and progression of tumors has been receiving increasing attention. Colon cancer-associated transcript 2 (CCAT2), a type of oncogenic lncRNA, is regarded as a novel biomarker of poor prognosis and metastasis in various types of cancer. However, the molecular contributions of CCAT2 to gastric cancer (GC) progression remain largely unclear. The aim of the present study was to demonstrate the effect of silencing CCAT2 on the biological behavior of GC BGC-823 cells and illustrate the potential underlying molecular mechanisms. A short hairpin RNA interference plasmid pRNAT-U6.1-CCAT2 targeting CCAT2 was successfully constructed. At 48 h after transfection with the interference plasmid, the survival rate of BGC-823 cells was significantly decreased, as determined by the MTT assay. In addition, RT-qPCR results revealed that CCAT2 gene expression was effectively suppressed by the transfection, while POU domain class 5 transcription factor 1B (POU5F1B) gene expression was significantly decreased. Terminal deoxynucleotidyl transferase dUTP nick end labeling assay further revealed that the apoptotic index was significantly higher in the interference group. Western blot analysis also demonstrated that the expression of beclin-1 protein was significantly increased, whereas the expression levels of phosphoinositide 3-kinase (PI3K) and mammalian target of rapamycin (mTOR) proteins were downregulated in the interference group. In conclusion, CCAT2 was able to positively regulate the expression of POU5F1B gene. Furthermore, silencing of CCAT2 gene inhibited the proliferation of BGC-823 cells, as well as induced apoptosis and autophagy in BGC-823 cells, by suppression of the PI3K/mTOR signaling pathways.
Keywords: gastric cancer, long non-coding RNA, colon cancer-associated transcript 2, POU domain class 5 transcription factor 1B, apoptosis, autophagy
Introduction
Gastric cancer (GC) is currently the fourth most common cancer and the second most common cause of cancer-associated mortality worldwide (1). In spite of the continuous development of novel treatment options for GC, the prognosis of this disease remains generally poor. The carcinogenesis and progression of GC are complicated processes, among which the detailed molecular mechanisms have not been elaborated thus far. The role of long non-coding RNAs (lncRNAs) in the carcinogenesis and progression of tumors has been receiving increasing attention. lncRNAs, a type of non-coding RNA of >200 nucleotides, lack an open reading frame and are unable to encode proteins; however, they are involved in the regulation of cellular development, proliferation, differentiation and apoptosis (2). The dysregulation of lncRNAs has been confirmed to be associated with tumor initiation, progression, invasion and metastasis in various types of cancer (3). Therefore, comprehensive investigation of the lncRNA function will provide a novel insight into the diagnosis, prognosis and targeted therapy for cancer.
Colon cancer-associated transcript 2 (CCAT2), a novel lncRNA first identified by Ling et al (4), encompasses the rs6983267 single nucleotide polymorphism (SNP) and is highly overexpressed in microsatellite-stable colorectal cancer. A meta-analysis demonstrated that high expression of CCAT2 may serve as a novel biomarker of poor prognosis and metastasis in various cancer types (5). Two previous experimental studies examining the association of CCAT2 and GC confirmed that a high expression of CCAT2 was closely associated with a higher incidence of lymph node and distant metastases, as well as shorter overall and progression-free survival rates (6,7). However, the molecular contributions of CCAT2 to GC progression remain largely unclear.
In the present study, it was attempted to illustrate the function of CCAT2 by the construction of an interference short hairpin RNA (shRNA) plasmid targeting CCAT2. In addition, the effect of this plasmid on the proliferation, apoptosis and autophagy of GC cells, as well as the potential underlying molecular mechanisms, were investigated.
Materials and methods
Construction of shRNA plasmid targeting CCAT2
An interference sequence was designed according to the sequence of CCAT2 in GenBank (NC_000008.11), and Basic Local Alignment Search Tool Analysis (blast.ncbi.nlm.nih.gov/Blast.cgi) indicated no homology with other genes. shRNA was synthesized by two complementary oligonucleotide strands, as follows: 5′-CGCGGGATCCCGGTGCAACTCTGCAATTTAATTTTCAAGAGAAATTAAATTGCAGAGTTGCACTTTTTTCCAAAAGCTTAA-3′ (sense) and 5′-AACTTAAGCTTTTGGAAAAAAGTGCAACTCTGCAATTTAATTTCTCTTGAAAATTAAATTGCAGAGTTGCACCGGGATCCC-3′ (antisense). Double-stranded oligonucleotides (ds-oligo) of shRNA were obtained subsequent to annealing at 95°C for 5 min. Plasmid pRNAT-U6.1 (GenScript, Jiangsu, China) and shRNA ds-oligo were digested using the enzymes BamHI and HindIII (Takara Biotechnology Co., Ltd., Dalian, China) simultaneously in a water bath of 37°C for 30 min. The reaction system of dual enzyme digestion included 1 µl BamHI, 1 µl HindIII, 2 µl 10X K buffer, 5 µl pRNAT-U6.1 plasmid or shRNA ds-oligo, and 11 µl ddH2O. DNA was purified and recovered according to the instructions of the DNA gel extraction kit (Tiangen Biotech Co., Ltd., Beijing, China). Next, T4 DNA ligase (NEB, Beijing, China) was used to clone the shRNA ds-oligo into the linear plasmid pRNAT-U6.1, and the reaction system of DNA directional ligation was built following the manufacturer's protocol. The products were transformed into Trans5α competent cells (Tiangen Biotech Co., Ltd.), and then monoclonal clones were identified with polymerase chain reaction (PCR) and enzyme digestion. Briefly, Luria-Bertani bacterial liquid medium was added to the 96-well plate (200 µl/well). Monoclonal transformants were randomly selected in the bacterial culture plate and added to the wells of 96-well plate with sterile toothpicks. Following incubation in a bacterial incubator for 1 h at 37°C, the bacterial liquid was used for PCR identification. Reaction system of PCR included 10 µl 2X Taq Master Mix (Lifefeng Biotechnology Co., Ltd., Shanghai, China), 2 µl 10 µM primer mixture, 1 µl bacteria liquid, and 7 µl ddH2O. The following primers were used: The following primers for PCR identification were used: forward, 5′-GGATCCCGGTGCAACTCTG-3′ and reverse, 5′-AAGGCACAGTCGAGGCTGAT-3′ (Chongqing Western Biological Medicine Science and Technology Co., Ltd., Chongqing, China). PCR reaction conditions were as follows: 95°C for 5 min; 94°C for 30 sec, 60°C for 30 sec, 72°C for 10 min, 30 cycles. The PCR products were analyzed using 1% agarose gel electrophoresis. Positive clones were considered when there were DNA bands. The steps of enzyme digestion were the same as those methods mentioned above, and the products were added to 1% agarose gel electrophoresis. Positive clones were considered when there were DNA fragments 69 bp long. The positive clone was further sequenced to identify the successfully constructed recombinant pRNAT-U6.1-CCAT2.
Cell culture and transfection
The human BGC-823 cell line was purchased from the Type Culture Collection of the Chinese Academy of Sciences (Beijing, China). The cells were cultured in RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.), 2 mM glutamine, 100 U/ml penicillin and 100 µg/ml streptomycin, and maintained at 37°C in an atmosphere of 95% O2 and 5% CO2. The experimental groups were as follows: i) Blank control (BC) group, untransfected BGC-823 cells; ii) negative control (NC) group, BGC-823 cells transfected with the empty pRNAT-U6.1 plasmid; and iii) interference group, BGC-823 cells transfected with the pRNAT-U6.1-CCAT2 plasmid. Prior to transfection, BGC-823 cells (1×105/ml) were grown in 24-well plates until 70–80% confluence was reached. Cell transfection was performed using Lipofectamine® 2000 (Invitrogen; Thermo Fisher Scientific, Inc.) according to the manufacturer's instructions. Briefly, 2 µl/well Lipofectamine® 2000 or 0.8 µg vector DNA was diluted to a final volume of 50 µl using Opti-MEM medium (Invitrogen; Thermo Fisher Scientific, Inc.), respectively. The diluent of vector DNA was added to Lipofectamine® 2000, and then the mixture were incubated for 20 min at room temperature. The culture medium was removed from the plate, and then the transfection complex was added. Following incubation for 4–6 h at 37°C, fresh medium was used for further cell culturing.
Cell proliferation assays
BGC-823 cells in the logarithmic growth phase were counted to adjust the cell concentration to 1×105/ml. Next, the cells were seeded into 96-well plates with 100 µl/well, transfected with the plasmid based on the experimental grouping and then cultured for 12, 24 and 48 h. Triplicate wells were used for each group. The cell proliferation was assessed using an MTT Cell Proliferation Reagent kit (Beyotime Institute of Biotechnology, Jiangsu, China) following the manufacturer's instructions. Briefly, 10 µl MTT (5 g/l) was added to each well subsequent to culture for 12, 24 or 48 h, and the cells were maintained in the culture medium for 4 h. Next, 150 µl dimethyl sulfoxide per well was added following the removal of the culture medium. The 96-well plate was then placed in an oscillator for low speed oscillation for 10 min. Subsequently, the absorbance of each well at 490 nm was detected using a microplate reader, and the cell survival was calculated as follows: Cell survival rate (%) = (absorbance of experimental group)/(absorbance of untransfected blank control group) × 100%.
Reverse transcription-quantitative PCR (RT-qPCR) assay
BGC-823 cells were harvested at 48 h after transfection. Total RNA was isolated from the cultured cells using TRIzol® reagent (Shinegene Molecular Biotech, Inc., Shanghai, China). RT of total RNA into first-strand cDNA and qPCR analysis were performed using RT kit (Chongqing Western Biological Medicine Science and Technology Co., Ltd., Chongqing, China) and Shinegene Real Time PCR Core kit (Shinegene Molecular Biotech, Inc.), respectively, according to the manufacturer's instructions.
The reaction system of RT assay included 10 µl 2X RT buffer, 1 µl 6N random primer (100 pmol/µl), 1 µl RT-Mix, 5 µl RNA and 3 µl RNase-free ddH2O. The RT reaction was conducted at 25°C for 10 min, 42°C for 50 min and then 85°C for 5 min. Subsequently, qPCR was performed in a 50 µl reaction system containing 25 µl 2X PCR buffer, 1 µl each primer (25 pmol/µl), 0.5 µl SYBR Green I (Shinegene Real Time PCR Core kit; Shinegene), 2 µl cDNA and 20.5 µl RNase-free ddH2O. The house keeping gene β-actin mRNA was used to normalize the level of cDNA. The following primers were used in the qPCR assay: CCAT2 forward, 5′-AAGAGGAAACCACCTTGGACTG-3′ and reverse, 5′-GCAATAAGAGCAGGAAAAGAAGC-3′; POU domain class 5 transcription factor 1B (POU5F1B) forward, 5′-GCGATCAAGCAGCGACTATG-3′ and reverse, 5′-CAGGGAAAGGGACTGAGGAG-3′; and β-actin forward, 5′-TGACGTGGACATCCGCAAAG-3′ and reverse, 5′-CTGGAAGGTGGACAGCGAGG-3′. The qPCR reaction conditions were as follows: 94°C for 4 min, followed by 35 cycles of amplification at 94°C for 20 sec, 60°C for 15 sec and 72°C for 30 sec. All samples were amplified in triplicates. The results were evaluated with the Bio-Rad CFX Manager software (version 3.1; Bio-Rad Laboratories, Inc., Hercules, CA, USA). The quantification cycle (Cq) value was read, and the relative expression of target gene was calculated according to the 2−ΔΔCq method (8).
Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay
BGC-823 cells were seeded in the 6-well plates containing sterile coverslips, and cultured 48 h after plasmid transfection. Subsequent to fixing using 4% paraformaldehyde for 30 min, apoptotic cells were detected with an In Situ Cell Apoptosis Detection kit (Boster Biological Technology, Pleasanton, CA, USA) according to the method described by Zhao et al (9). Apoptotic cells exhibited positive staining (pale brown) in cell nucleus. Five high power fields of each slide at a magnification of ×400 were randomly examined under a light microscope to determine the percentage of apoptotic cells (expressed as the apoptotic index) among at least 500 cells.
Western blot analysis
The culture medium of BGC-823 cells was discarded 48 h after transfection. Cells were then lysed and total proteins were extracted using a radioimmunoprecipitation assay protein extraction reagent (Beyotime Institute of Biotechnology). Next, the protein concentration was quantified using a bicinchoninic acid protein assay kit (Beyotime Institute of Biotechnology). Equal amounts (20 µg) of protein extracts were separated by 10% SDS-PAGE, and then transferred to polyvinylidene fluoride membranes (Bio-Rad Laboratories, Inc.). Subsequent to blocking with 5% skim-milk for 2 h at room temperature, the membranes were incubated with rabbit polyclonal antibodies against beclin-1 (1:500; cat. no. ab62557), mammalian target of rapamycin (mTOR; 1:50; cat. no. ab2732), phosphoinositide 3-kinase (PI3K; 1:500; cat. no. ab151549) and β-actin (1:1,000; cat. no. ab8227; all from Abcam, Cambridge, UK) at 4°C overnight. The membranes were then washed in Tris-buffered saline/Tween-20 and incubated with goat anti-rabbit immunoglobulin G (1:1,000; cat. no. A0545; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) for 2 h at 37°C. Protein bands were visualized using the BeyoECL Plus reagent (Beyotime Institute of Biotechnology), and the fluorescence intensity was quantified using the ImageJ v.2.1.4.7 software (National Institutes of Health, Bethesda, MD, USA).
Statistical analysis
All data are presented as the mean ± standard deviation. Differences among groups were tested by one-way analysis of variance, followed by Student-Newman-Keuls test. Statistical analyses were performed using SPSS v.22.0 software (IBM Corp., Armonk, NY, USA). P<0.05 was considered to indicate a statistically significant difference. Graphs were constructed with GraphPad Prism 5.0 software (GraphPad Software, Inc., La Jolla, CA, USA).
Results
Construction of pRNAT-U6.1-CCAT2 interference plasmid
In the present study, a recombinant shRNA interference plasmid was constructed to investigate the function of CCAT2 gene in GC cells. The positive clones were sequenced and compared with target DNA sequencing, and the results demonstrated that the inserted DNA fragment was identical to the expected CCAT2 interference sequence, suggesting that the interference plasmid pRNAT-U6.1-CCAT2 was successfully constructed.
MTT assay for determination of the cell viability
The viability of the GC cells was determined at three time-points, namely 12, 24 and 48 h, using an MTT assay. As observed in Fig. 1, the survival rate of BGC-823 cells gradually declined with the increase in culture time following transfection with the pRNAT-U6.1-CCAT2 plasmid. The lowest cell viability was observed at 48 h, with a statistically significant difference observed at this time-point when compared with the BC and NC groups (P<0.05).
Figure 1.
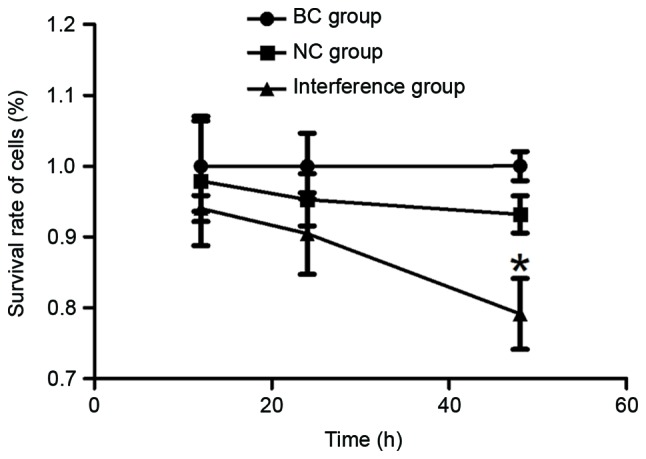
Survival rate of BGC-823 cells in each group at three time-points (12, 24 and 48 h). The survival rate was only 79.15% at 48 h in the interference group transfected with the pRNAT-U6.1-CCAT2 plasmid. *P<0.05 vs. the BC and NC groups. CCAT2, colon cancer-associated transcript 2; BC, blank control; NC, negative control.
RT-qPCR detection of CCAT2 and POU5F1B gene expression levels
To confirm the effect of pRNAT-U6.1-CCAT2 plasmid on the gene expression levels of CCAT2 and POU5F1B in BGC-823 cells, RT-qPCR detection was performed. As presented in Fig. 2, the RT-qPCR results demonstrated that the relative expression of CCAT2 in the interference group decreased significantly as compared with that of the BC and NC groups (P<0.05), whereas no significant difference in CCAT2 expression was observed between the BC and NC groups, indicating that the pRNAT-U6.1-CCAT2 plasmid was able to silence the function of the CCAT2 gene in BGC-823 cells. Along with the decrease in the expression of CCAT2 in the plasmid-transfected BGC-823 cells, the relative expression of POU5F1B gene was also downregulated in the interference group, with a significant difference detected compared with the BC and NC groups (P<0.05; Fig. 2).
Figure 2.
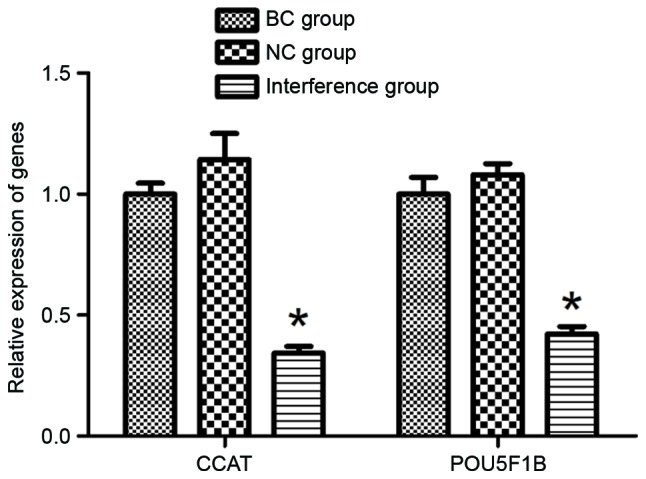
Relative expression levels of the CCAT2 and POU5F1B genes in BGC-823 cells at 48 h in each group, as determined using reverse transcription-quantitative polymerase chain reaction. The expression levels of the CCAT2 and POU5F1B genes in the interference group that was transfected with the pRNAT-U6.1-CCAT2 plasmid were downregulated by 0.34- and 0.42-fold, respectively, compared with the BC group. *P<0.05 vs. BC and NC groups. CCAT2, colon cancer-associated transcript 2; POU5F1B, POU domain class 5 transcription factor 1B; BC, blank control; NC, negative control.
TUNEL detection of cell apoptosis
TUNEL staining identified that typical apoptotic features were observed in the BGC-823 cells, with numerous brown granules products in the nucleus of apoptotic cells (Fig. 3A-C). The TUNEL results revealed that the apoptotic index of BGC-823 cells in the interference group (33.98±5.22%) was significantly higher as compared with that in the BC and NC groups (10.23±3.34 and 14.12±5.23%, respectively; P<0.05; Fig. 3D).
Figure 3.

Detection of BGC-823 cell apoptosis at 48 h in each group using TUNEL assay. TUNEL staining of the BGC-823 apoptotic cells are presented for the (A) BC, (B) NC and (C) interference groups (magnification, ×400). (D) Histogram results demonstrated the difference of apoptotic index between three groups. *P<0.05 vs. BC and NC groups. TUNEL, terminal deoxynucleotidyl transferase dUTP nick end labeling; BC, blank control; NC, negative control.
Western blot detection of protein expression
The autophagy-associated protein beclin-1 and the PI3K/mTOR signaling pathway proteins in BGC-823 cells at 48 h after transfection with the plasmids were detected using western blot assay. The results demonstrated that BGC-823 cells in the interference group exhibited a significantly increased expression of beclin-1 protein (1.99- and 1.81-fold increase vs. the BC and NC groups, respectively; P<0.05; Fig. 4). Furthermore, the expression levels of PI3K and mTOR proteins in the interference group were downregulated by 0.78- and 0.69-fold compared with those in the BC and NC groups (P<0.05).
Figure 4.
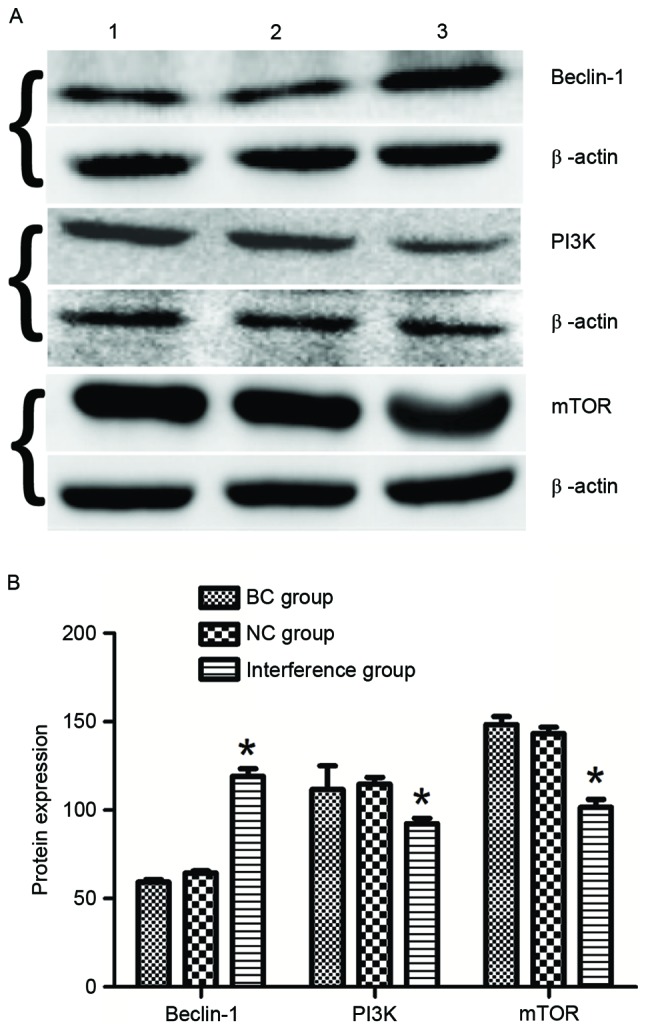
Detection of protein expression levels of Beclin-1, PI3K and mTOR of BGC-823 cells at 48 h in each group using western blot analysis. (A) Protein bands are demonstrated for the BC, NC and interference groups. β-actin was used as an internal reference. (B) Quantified results demonstrate the difference in the relative protein expression levels among the three groups. *P<0.05 vs. the BC and NC groups. PI3K, phosphoinositide 3-kinase; mTOR, mammalian target of rapamycin; BC, blank control; NC, negative control.
Discussion
lncRNAs function as oncogenes or tumor suppressors during the occurrence and development of GC. Several lncRNAs, including LINC00152, GHET1, HULC, HOTAIR, GACAT3, MALAT2 and H19, have been demonstrated to serve as oncogenes, whereas other lncRNAs, including LEIGC, GAS5 and FER1L4, have been considered to be tumor suppressors (10). Emerging evidence indicated that CCAT2 functions as a cancer-promoting molecule in several types of solid cancer, such as in non-small cell lung cancer (11), oral squamous cell carcinoma (12), ovarian cancer (13), and hepatocellular carcinoma (14). The present study investigated the effects of the lncRNA CCAT2 on the biological behavior of BGC-823 cells, and revealed that knockdown of CCAT2 expression effectively inhibited the proliferation, as well as induced the apoptosis and autophagy of GC cells in vitro. These effects may be derived from the downregulation of the expression of POU5F1B gene as a result of silencing CCAT2.
Recent studies confirmed that the lncRNA CCAT2, located at the 8q24 amplicon of the cancer risk-associated rs6983267 SNP, regulated the cancer metabolism in vitro and in vivo by directly interacting in an allele-specific manner with a protein complex (15,16). The chromosomal region 8q24 emerged as an important region for genetic susceptibility in various cancer types, thus DNA methylation or SNPs at this locus may contribute to cancer risk (17,18). The retrogene POU5F1B has been observed to be located adjacent to the MYC gene within this risk locus at chromosome 8q24 (19). POU5F1B has a preserved open reading frame encoding a homolog of the master embryonic stem cell transcription factor POU5F1, also known as octamer-binding transcription factor 4. A study revealed that the expression of POU5F1B was upregulated in GC cell lines and tissues, while POU5F1B also exhibited mitogenic, angiogenic and antiapoptotic effects in vivo (20). In the present study, it was observed that the expression of POU5F1B was downregulated along with the decreased expression of CCAT2 in BGC-823 cells transfected with the CCAT2-silencing plasmid, which suggested that CCAT2 positively regulated the expression of POU5F1B gene to a certain extent. Therefore, the POU5F1B gene may be one of the downstream target genes of CCAT2.
As the two main forms of programmed cell death, autophagy and apoptosis induce the degradation of proteins and organelles, or cell death upon cellular stress (21). Autophagy is associated with both the tumorigenic and protective effects in cancer; however, the role of autophagy in GC remains unclear (22). During the autophagic process, the autophagy-associated markers beclin-1 (which is the mammalian homologue of yeast Atg6) serves not only as a key autophagy regulator with its specific interactors, but also as a potential therapeutic target in cancer (23). A meta-analysis revealed that the expression of beclin-1 in GC tissues was significantly reduced when compared with that in non-GC tissues, and the upregulated expression of beclin-1 was associated with marked differentiation of tumor cells, no distant metastasis and a favorable overall survival of GC (24–26). Furthermore, there are close interactions between autophagy and apoptosis through shared signaling pathways, among which the PI3K/protein kinase B (AKT)/mTOR is a crucial intracellular signaling pathway in tumorigenesis (27). Liu et al (28) demonstrated that celecoxib may impact apoptosis and autophagy via the PI3K/AKT signaling pathway in the SGC-7901 GC cells. Therefore, the PI3K/AKT/mTOR signaling pathway serves a critical role in the regulation of apoptosis and autophagy. In the present study, the data revealed that silencing of the CCAT2 gene induced upregulation of the apoptosis and autophagy of GC cells by downregulating the expression levels of PI3K and mTOR proteins.
In conclusion, the present study constructed a CCAT2 interference plasmid, and further examined the effect of CCAT2 on the biological behavior of GC cells and its possible underlying molecular mechanism. However, the specific regulatory mechanisms require further investigation.
Acknowledgements
This study was supported by grants from the Guizhou Provincial Fund Project of Science and Technology [grant no. (2015)2095], the Science and Technology Fund Projects of Guizhou Provincial Health and Family Planning Commission (grant no. gzwjkj2015-1-019), and the Joint Fund Project of Guizhou Provincial Science and Technology Agency [grant no. (2015)7170].
References
- 1.Brenner H, Rothenbacher D, Arndt V. Epidemiology of stomach cancer. Methods Mol Biol. 2009;472:467–477. doi: 10.1007/978-1-60327-492-0_23. [DOI] [PubMed] [Google Scholar]
- 2.Zhang M, Du X. Noncoding RNAs in gastric cancer: Research progress and prospects. World J Gastroenterol. 2016;22:6610–6618. doi: 10.3748/wjg.v22.i29.6610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Zhang R, Xia LQ, Lu WW, Zhang J, Zhu JS. LncRNAs and cancer. Oncol Lett. 2016;12:1233–1239. doi: 10.3892/ol.2016.4770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ling H, Spizzo R, Atlasi Y, Nicoloso M, Shimizu M, Redis RS, Nishida N, Gafà R, Song J, Guo Z, et al. CCAT2, a novel noncoding RNA mapping to 8q24, underlies metastatic progression and chromosomal instability in colon cancer. Genome Res. 2013;23:1446–1461. doi: 10.1101/gr.152942.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Fan YH, Fang H, Ji CX, Xie H, Xiao B, Zhu XG. Long noncoding RNA CCAT2 can predict metastasis and poor prognosis: A meta-analysis. Clin Chim Acta. 2017;466:120–126. doi: 10.1016/j.cca.2017.01.016. [DOI] [PubMed] [Google Scholar]
- 6.Wang CY, Hua L, Yao KH, Chen JT, Zhang JJ, Hu JH. Long non-coding RNA CCAT2 is up-regulated in gastric cancer and associated with poor prognosis. Int J Clin Exp Pathol. 2015;8:779–785. [PMC free article] [PubMed] [Google Scholar]
- 7.Wang YJ, Liu JZ, Lv P, Dang Y, Gao JY, Wang Y. Long non-coding RNA CCAT2 promotes gastric cancer proliferation and invasion by regulating the E-cadherin and LATS2. Am J Cancer Res. 2016;6:2651–2660. [PMC free article] [PubMed] [Google Scholar]
- 8.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 9.Zhao X, Ma C, Cai X, Lei D, Liu D, Xu F, Jin T, Liu J, Pan X. RNA interference of caveolin-1 via lentiviral vector inhibits growth of hypopharyngeal squamous cell carcinoma FaDu cells in vitro and in vivo. Asian Pac J Cancer Prev. 2011;12:397–401. [PubMed] [Google Scholar]
- 10.Sun W, Yang Y, Xu C, Xie Y, Guo J. Roles of long noncoding RNAs in gastric cancer and their clinical applications. J Cancer Res ClinOncol. 2016;142:2231–2237. doi: 10.1007/s00432-016-2183-7. [DOI] [PubMed] [Google Scholar]
- 11.Chen S, Wu H, Lv N, Wang H, Wang Y, Tang Q, Shao H, Sun C. LncRNA CCAT2 predicts poor prognosis and regulates growth and metastasis in small cell lung cancer. Biomed Pharmacother. 2016;82:583–588. doi: 10.1016/j.biopha.2016.05.017. [DOI] [PubMed] [Google Scholar]
- 12.Ouyang S, Zhang P, Wang J, Huang Z, Liao L. Expression of long non-coding RNA colon cancer associated transcript 2 and its clinicopathologic significance in oral squamous cell carcinoma. Zhonghua Kou Qiang Yi XueZaZhi. 2016;51:286–291. doi: 10.3760/cma.j.issn.1002-0098.2016.05.007. (In Chinese) [DOI] [PubMed] [Google Scholar]
- 13.Huang S, Qing C, Huang Z, Zhu Y. The long non-coding RNA CCAT2 is up-regulated in ovarian cancer and associated with poor prognosis. Diagn Pathol. 2016;11:49. doi: 10.1186/s13000-016-0499-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Zhou N, Si Z, Li T, Chen G, Zhang Z, Qi H. Long non-coding RNA CCAT2 functions as an oncogene in hepatocellular carcinoma, regulating cellular proliferation, migration and apoptosis. Oncol Lett. 2016;12:132–138. doi: 10.3892/ol.2016.4580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Redis RS, Vela LE, Lu W, Ferreira de Oliveira J, Ivan C, Rodriguez-Aguayo C, Adamoski D, Pasculli B, Taguchi A, Chen Y, et al. Allele-specific reprogramming of cancer metabolism by the long non-coding RNA CCAT2. Mol Cell. 2016;61:520–534. doi: 10.1016/j.molcel.2016.02.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Redis RS, Calin GA. The interplay between lnRNAs, SNPs, and protein complexes-what does it mean for cancer metabolism? Mol Cell Oncol. 2016;3:e1166308. doi: 10.1080/23723556.2016.1166308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Barry KH, Moore LE, Sampson J, Yan L, Meyer A, Oler AJ, Chung CC, Wang Z, Yeager M, Amundadottir L, Berndt SI. DNA methylation levels at chromosome 8q24 in peripheral blood are associated with 8q24 cancer susceptibility loci. Cancer Prev Res (Phila) 2014;7:1282–1292. doi: 10.1158/1940-6207.CAPR-14-0132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Brisbin AG, Asmann YW, Song H, Tsai YY, Aakre JA, Yang P, Jenkins RB, Pharoah P, Schumacher F, Conti DV, et al. Meta-analysis of 8q24 for seven cancers reveals a locus between NOV and ENPP2 associated with cancer development. BMC Med Genet. 2011;12:156. doi: 10.1186/1471-2350-12-156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Breyer JP, Dorset DC, Clark TA, Bradley KM, Wahlfors TA, McReynolds KM, Maynard WH, Chang SS, Cookson MS, Smith JA, et al. An expressed retrogene of the master embryonic stem cell gene POU5F1 is associated with prostate cancer susceptibility. Am J Hum Genet. 2014;94:395–404. doi: 10.1016/j.ajhg.2014.01.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hayashi H, Arao T, Togashi Y, Kato H, Fujita Y, De Velasco MA, Kimura H, Matsumoto K, Tanaka K, Okamoto I, et al. The OCT4 pseudogene POU5F1B is amplified and promotes an aggressive phenotype in gastric cancer. Oncogene. 2015;34:199–208. doi: 10.1038/onc.2013.547. [DOI] [PubMed] [Google Scholar]
- 21.Liu G, Pei F, Yang F, Li L, Amin AD, Liu S, Buchan JR, Cho WC. Role of autophagy and apoptosis in non-small-cell lung cancer. Int J Mol Sci. 2017;18:E367. doi: 10.3390/ijms18020367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Zhou H, Yuan M, Yu Q, Zhou X, Min W, Gao D. Autophagy regulation and its role in gastric cancer and colorectal cancer. Cancer Biomark. 2016;17:1–10. doi: 10.3233/CBM-160613. [DOI] [PubMed] [Google Scholar]
- 23.Fu LL, Cheng Y, Liu B. Beclin-1: Autophagic regulator and therapeutic target in cancer. Int J Biochem Cell Biol. 2013;45:921–924. doi: 10.1016/j.biocel.2013.02.007. [DOI] [PubMed] [Google Scholar]
- 24.Yu ZY, Ding J, Yang XF, Zhang ZM. Significance of autophagy-related protein beclin-1 expression in gastric cancer: A Meta-analysis. Chin J Gen Surg. 2015;24:1389–1395. [Google Scholar]
- 25.He Y, Zhao X, Subahan NR, Fan L, Gao J, Chen H. The prognostic value of autophagy-related markers beclin-1 and microtubule-associated protein light chain 3B in cancers: A systematic review and meta-analysis. Tumour Biol. 2014;35:7317–7326. doi: 10.1007/s13277-014-2060-4. [DOI] [PubMed] [Google Scholar]
- 26.Cao QH, Liu F, Yang ZL, Fu XH, Yang ZH, Liu Q, Wang L, Wan XB, Fan XJ. Prognostic value of autophagy related proteins ULK1, Beclin 1, ATG3, ATG5, ATG7, ATG9, ATG10, ATG12, LC3B and p62/SQSTM1 in gastric cancer. Am J Transl Res. 2016;8:3831–3847. [PMC free article] [PubMed] [Google Scholar]
- 27.Polivka J, Jr, Janku F. Molecular targets for cancer therapy in the PI3K/AKT/mTOR pathway. Pharmacol Ther. 2014;142:164–175. doi: 10.1016/j.pharmthera.2013.12.004. [DOI] [PubMed] [Google Scholar]
- 28.Liu M, Li CM, Chen ZF, Ji R, Guo QH, Li Q, Zhang HL, Zhou YN. Celecoxib regulates apoptosis and autophagy via the PI3K/Akt signaling pathway in SGC-7901 gastric cancer cells. Int J Mol Med. 2014;33:1451–1458. doi: 10.3892/ijmm.2014.1713. [DOI] [PMC free article] [PubMed] [Google Scholar]