

Abstract
Statins have become of interest in research due to their anticancer effects. However, the exact mechanism of their anticancer properties remains unclear. The authors previously reported that statins decrease intracellular cholesterol levels in androgen-independent prostate cancer cells. In de novo androgen synthesis, cholesterol is the primary material and certain enzymes have important roles. The present study aimed to determine whether simvastatin alters the expression of androgen synthesis-associated enzymes in androgen-independent prostate cancer cells. A novel combination therapy of statins and other drugs that inhibit the overexpression of enzymes involved in androgen synthesis was explored. The cytotoxicity of simvastatin and meclofenamic acid was assessed in prostate cancer cells using MTS and migration assays. Testosterone and dihydrotestosterone concentrations in the culture medium were measured using liquid chromatography-tandem mass spectrometry. RAC-α-serine/threonine-protein kinase (Akt) phosphorylation was detected by western blot analysis. Following treatment with simvastatin, aldo-keto reductase family 1 member C3 (AKR1C3) expression increased in PC-3 (>60-fold) and LNCaP-LA cells, however not in 22Rv1 cells. Small interfering (si)RNA was used to clarify the effects of AKR1C3 expression. The reduction in AKR1C3 expression in PC-3 cells following siRNA transfection was not associated with basal cell proliferation and migration; however, treatment with simvastatin decreased cell proliferation and migration. The combination of simvastatin and meclofenamic acid, an AKR1C3 inhibitor, further enhanced the inhibition of cell proliferation and migration compared with treatment with either drug alone. Furthermore, treatment with simvastatin attenuated insulin-like growth factor 1-induced Akt activation; however, the combination of simvastatin and meclofenamic acid further inhibited Akt activation. These results suggest that the combination of simvastatin and meclofenamic acid may be an effective strategy for the treatment of castration-resistant prostate cancer.
Keywords: prostate cancer, statins, meclofenamic acid, AKR1C3
Introduction
A statin is a drug used to treat hyperlipidemia and functions by inhibiting 3-hydroxy-3-metylglutaryl coenzyme A reductase. Statins have gained much recent attention due to their anticancer effects. Previous studies have shown that statins can prolong survival, while others have reported no benefits in cancer patients (1). Concerning prostate cancer, the anticancer effect of statins is controversial (2,3). We previously reported that statins inhibit prostate cancer progression via suppressing the expression of insulin-like growth factor 1 receptor (IGF1R) and increasing ANXA10 (4,5). However, the exact mechanism of their anticancer properties remains unclear.
There has been recent interest and concerns regarding intratumoral de novo androgen synthesis in castration-resistant prostate cancer (CRPC). Now we are treating CRPC patients with enzalutamide and abiraterone, which attenuate the effects of intratumoral de novo androgens. In de novo androgen synthesis, cholesterol is the primary material, and various enzymes play important roles. We previously reported that intracellular cholesterol levels are decreased in androgen-independent prostate cancer cells after treatment with simvastatin (6); however, alterations in androgen synthesis-related enzymes are not clear.
In this study, we determined whether simvastatin alters the expression of enzymes involved in androgen synthesis in CRPC cells. We also explored a new combination therapy of statins and other drugs that inhibit the overexpression of androgen synthesis-related enzymes.
Materials and methods
Cells and chemicals
Human prostate cancer cell lines PC-3, LNCaP, and 22RV1 were purchased from DS Pharma Biomedical (Osaka, Japan) and cultured in RPMI 1640 (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) supplemented with 10% FBS (Moregate BioTech, Bulimba, Australia). PC-3 is an androgen receptor-negative human prostate cancer cell line (7). LNCaP-LA cells, which were generated from LNCaP cells, were cultured in medium containing 10% charcoal-stripped fetal bovine serum (FBS) for more than 3 months.
Measurement of testosterone and dihydrotestosterone (DHT) in culture medium
Cells were cultured on a 6-well plate and incubated overnight in medium containing 10% FBS. Cells were then incubated with or without simvastatin (5 µM). After 48 h, androstenedione (100 µM) was added to the medium. After 24 h, culture medium was collected, and testosterone and DHT concentrations were measured using liquid chromatography-tandem mass spectrometry (LC-MS/MS) (ASKA Pharmaceutical Medical Co., Ltd., Kawasaki, Japan). RIPA buffer was added to wells and protein concentration was measured by the DC Protein Assay (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Testosterone and DHT levels were calculated by dividing the results of the protein assay by the total protein concentration.
RT-qPCR
Transcript levels were quantified using the Applied Biosystems 7300 Real-Time PCR system (Applied Biosystems; Thermo Fisher Scientific, Inc., Waltham, MA, USA) according to the manufacturer's protocol. Total RNA was extracted, cDNA was synthesized (8), and polymerase chain reaction (PCR) amplification was performed, using 2 µl cDNA and the StAR, CYP11A1, CYP17A1, aldo-keto reductase family 1 member C3 (AKR1C3), HSD3B1, HSD3B2, SRD5A1, SRD5A2, and AKR1C2 primers (No. Hs00986559_g1, Hs00167984_m1, Hs01124136_m1, Hs00366267_m1, Hs00426435_m1, Hs00605123_m1, Hs00602694_mH, Hs00165843_m1, and Hs00912742_m1, respectively; Applied Biosystems). Next, PCR was performed for one cycle of 10 min at 95°C followed by 40 cycles of 15 sec at 95°C and 60 sec at 60°C. b-Actin (No. 4326315E, Applied Biosystems) transcript levels were used as the internal control. mRNA fold changes were quantified using ΔΔCq.
MTS assay
Cells were plated onto a 96-well plate in 100 µl culture medium containing 10% FBS. After 24 h, cells were incubated with medium containing simvastatin (5 µM) and/or meclofenamic acid (50 µM). After incubation for 48 h, the number of living cells was measured using the MTS assay.
Migration assay
Cells were plated onto a 12-well plate and grown to confluence. A 1,000-µl tip was used to make a denuded area. Cells were washed twice with PBS and incubated with medium containing various concentrations of simvastatin for 48 h. Mitomycin C (0.5 µM) was added to the medium to inhibit cell proliferation. Photographs were taken at 0 and 48 h, and the distance of cell migration was determined by subtracting the values obtained at 0 h from those obtained at 48 h. Migration distance is expressed as fold change over the control.
siRNA transfection
Cells were transfected with ON-TARGETplus Non-targeting Pool (no. D-001810-10-05; Dharmacon, Waltham, MA, USA) or ON-TARGETplus AKR1C3 siRNA (No. L-008116-00-0005, Dharmacon) using DharmaFect 2 (Dharmacon). Cells were incubated for 48 h after transfection.
Western blot analysis
Cell lysates were prepared in RIPA buffer containing 1 mM sodium orthovanadate (Sigma-Aldrich; Merck KGaA) and Halt Protease Inhibitor Cocktail (Pierce; Thermo Fisher Scientific, Inc.). Samples were boiled for 5 min; an equal amount of protein (30 µg/lane) was subjected to 4–12% SDS-PAGE and transferred onto nitrocellulose membranes. Each membrane was incubated with the following primary polyclonal antibodies: rabbit anti-Akt (1:1,000), rabbit anti-phospho-Akt (Ser473) (1:1,000) (Cell Signaling Technology, Inc., Beverly, MA, USA). Blots were developed using a 1:2,000 dilution of the HRP-conjugated secondary antibody (Cell Signaling Technology, Inc.). Proteins were visualized using Immobilon (Merck Millipore, Darmstadt, Germany).
Statistical analysis
Data are expressed as the mean ± standard deviation. Differences between values were evaluated by one-way ANOVA using Tukey's post hoc analysis and Student's t-test. P<0.05 was considered to indicate a statistically significant difference.
Results
Simvastatin altered the expression of genes encoding steroidogenic enzymes in androgen-independent prostate cancer cells
We examined PC-3, LNCaP-LA and 22Rv1 cells to determine whether simvastatin alters genes that encode steroidogenic enzymes in androgen-independent prostate cancer cells. After treatment with simvastatin, the expression of AKR1C3 was increased in PC-3 and LNCaP-LA cells (Figs. 1A and 2A) but not in 22Rv1 cells (data not shown). Moreover, the fold change was more than 60 times in PC-3 cells. Conversely, the expression of hydroxy-delta-5-steroid dehydrogenase, 3 beta- and steroid delta-isomerase 1 (HSD3B1) was decreased in PC-3 and LNCaP-LA cells (Figs. 1A and 2A) but not in 22Rv1 cells (data not shown). Moreover, simvastatin increased steroid 5 alpha-reductase 1 (SRD5A1) expression in PC-3 (Fig. 1A) but not in LNCaP-LA or 22Rv1 cells (data not shown). The expression of steroidogenic acute regulatory protein (StAR), cytochrome P450 family 11 subfamily A member 1 (CYP11A1), cytochrome P450 family 17 subfamily A member 1 (CYP17A1), hydroxy-delta-5-steroid dehydrogenase, 3 beta- and steroid delta-isomerase 2 (HSD3B2), steroid 5 alpha-reductase 2 (SRD5A2), and aldo-keto reductase family 1 member C2 (AKR1C2) did not change following treatment with simvastatin (data not shown).
Figure 1.

Effects of simvastatin on the expression of genes encoding steroidogenic enzymes in androgen-independent prostate cancer cells (A) PC-3 and (B) LNCaP-LA. Cells were incubated in medium containing various concentrations of simvastatin. After 48 h, we harvested the cells, and the mRNA expression levels of genes encoding steroidogenic enzymes were evaluated by qPCR. The temporal changes of gene expressions were evaluated following simvastatin treatment. Values are expressed as the mean ± SD (n=4). *P<0.01 vs. 0 µM, **P<0.05 vs. 0 µM.
Figure 2.
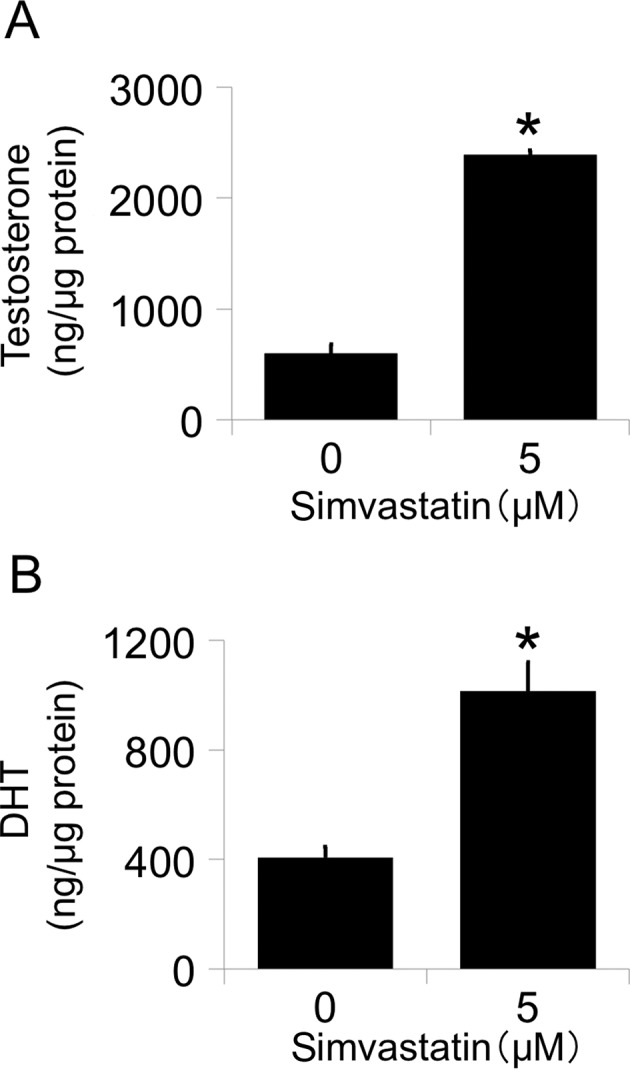
Effects of simvastatin on testosterone and dihydrotestosterone (DHT) levels in PC-3 cell culture medium. (A) Testosterone and (B) DHT concentrations were measured by LC-MS/MS. Values are expressed as the mean ± SD (n=3). *P<0.01 vs. 0 µM.
Effects of AKR1C3 expression on testosterone and DHT levels in PC-3 cell culture medium
To determine whether increased levels of AKR1C3 affect the de novo synthesis of intracellular androgen, we measured the testosterone and DHT levels in culture medium following treatment with simvastatin by LC-MS/MS. Simvastatin significantly increased both testosterone (Fig. 2A) and DHT (Fig. 2B) levels after the addition of androstenedione. These data show that the up-regulation of AKR1C3 is functional.
AKR1C3 inhibition increased the simvastatin-induced inhibition of cell proliferation and migration
PC-3 is an AR-negative human prostate cancer cell line. Therefore, there is a possibility that an increase in testosterone and DHT levels does not affect cell viability. In contrast, the overexpression of AKR1C3 promotes angiogenesis and aggressiveness in PC-3 cells (9). To further determine whether increased AKR1C3 expression affects simvastatin-induced cell viability, AKR1C3 expression was reduced by transfection with siRNA against AKR1C3. siRNA treatment inhibited the expression of AKR1C3 mRNA in PC-3 cells (Fig. 3A). The reduction in AKR1C3 expression in PC-3 cells following siRNA transfection was not associated with basal cell proliferation and migration; however, siRNA transfection with simvastatin significantly decreased both cell proliferation (Fig. 3B) and cell migration (Fig. 3C and D) compared to simvastatin alone.
Figure 3.
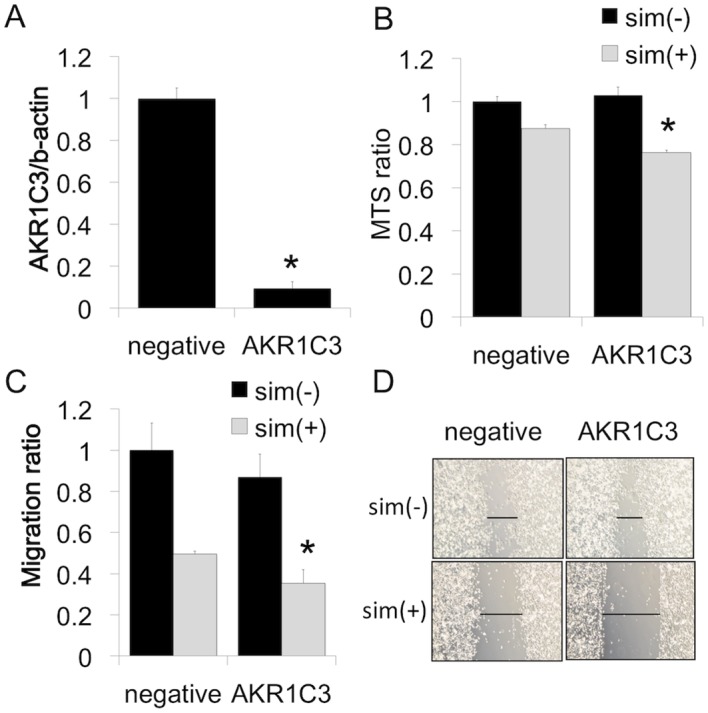
Effects of AKR1C3 siRNA on PC-3 cells. (A) Effects of siRNA on ABCA1 expression in PC-3 cells. Cells transfected with AKR1C3 siRNA (AKR1C3) or non-specific control siRNA (negative). Cells were incubated for 48 h before harvest for real-time PCR. Values are expressed as the mean ± SD (n=3). *P<0.01 vs. non-specific control siRNA. (B) After transfection of AKR1C3 or non-specific control siRNA, PC-3 cells were incubated with medium containing 10% FBS. After 48 h, PC-3 cells were cultured with or without simvastatin (5 µM) in medium containing 10% FBS. After 48 h, the number of viable cells was evaluated by the MTS assay. Values are expressed as the mean ± SD (n=4). *P<0.01 vs. negative/sim(+). (C and D) After transfection, PC-3 cells were incubated with medium containing 10% FBS. After 48 h, cells were wounded and cultured for 48 h in the presence or absence of simvastatin (5 µM). Cell migration into the wound was examined by phase-contrast microscopy. Values are expressed as the mean ± SD (n=3). *P<0.05 vs. negative/sim(+).
Meclofenamic acid increased the simvastatin-induced inhibition of cell proliferation and migration
Some drugs are reported to inhibit AKR1C3. Meclofenamic acid is an NSAID as well as one of the best inhibitors of AKRs, especially AKR1C3 (10). Therefore, we evaluated the combinatory effects of simvastatin and meclofenamic acid in PC-3 cells. Treatment with either simvastatin or meclofenamic acid alone inhibited cell proliferation (Fig. 4A) and migration (Fig. 4B and C). The combination of the two drugs further enhanced cell proliferation (Fig. 4A) and migration (Fig. 4B and C).
Figure 4.

Effects of combination simvastatin with meclofenamic acid on PC-3 cells. (A) PC-3 cells were incubated with medium containing 10% FBS. After 48 h, the PC-3 cells were cultured with or without simvastatin (5 µM) and meclofenamic acid (50 µM) in medium containing 10% FBS. After 48 h, the number of viable cells was evaluated by the MTS assay. Values are expressed as the mean ± SD (n=4). *P<0.01. (B and C) PC-3 cells were incubated with medium containing 10% FBS. After 48 h, cells were wounded and cultured with or without simvastatin (5 µM) and meclofenamic acid (50 µM). After 48 h, cell migration into the wound was examined by phase-contrast microscopy. Values are expressed as the mean ± SD (n=3). *P<0.01. sim, simvastatin.
Combination of simvastatin and meclofenamic acid inhibited IGF1-induced Akt activation
AKR1C3 overexpression induces Akt activation in PC-3 cells (9). We previously showed that simvastatin without IGF1 decreases IGF1R expression strongly in PC-3 cells (4). IGF1-Akt activation is a well-known pathway in prostate cancer. We hypothesized that inhibiting simvastatin-stimulated AKR1C3 expression with an AKR1C3 inhibitor would have a synergistic effect on simvastatin-blocked IGF1-induced Akt activation. Therefore, the effects of the combination of simvastatin and meclofenamic acid on IGF1-induced Akt activation were evaluated in PC-3 cells. Treatment with either simvastatin or meclofenamic acid alone attenuated IGF1-induced Akt activation, whereas the combination of simvastatin and meclofenamic acid further inhibited Akt activation (Fig. 5).
Figure 5.
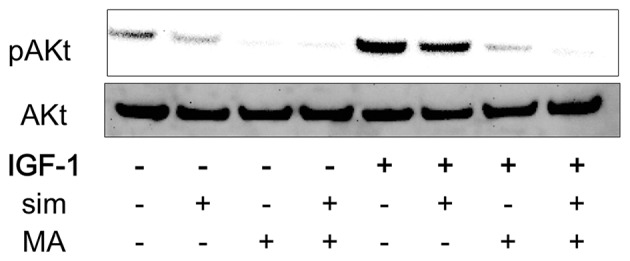
Effects of the combination of simvastatin with meclofenamic acid on IGF-1-induced Akt activation in PC-3 cells. PC-3 cells were incubated with medium containing 10% FBS. After 48 h, PC-3 cells were cultured with or without simvastatin (5 µM) and meclofenamic acid (50 µM) in medium containing 1% FBS. After 48 h, the cells were cultured with or without IGF-1 (40 ng/ml) for 120 min. All cells were harvested at the same time and cell lysates were prepared for western blotting. A representative experiment is shown, which was repeated three times with similar results. sim, simvastatin; MA, meclofenamic acid.
Discussion
The main finding of the present study was that simvastatin increased AKR1C3 expression in androgen-independent prostate cancer cells. Furthermore, the combination of simvastatin and meclofenamic acid, an AKR1C3 inhibitor, further suppressed PC-3 cell proliferation, migration, and Akt activation compared with simvastatin alone.
Statins have recently been studied for their pleiotrophic effects, which may make them relevant for cancer prevention or treatment. Clinical reports have shown that statin use is beneficial for overall survival and cancer-specific survival both before and after prostate cancer diagnosis (1,2). In contrast, Platz et al reported that the use of statin drugs was not associated with the overall risk of prostate cancer (3). In vitro, statins exert many biological activities that inhibit prostate cancer progression (e.g., lowering raft cholesterol content, inhibiting cyclin-dependent-kinase-2 activity, decreasing IGF1 receptor expression, and increasing ANXA10 expression) (4,5,11,12). These results indicate that statins have anticancer potential.
Statins inhibit 3-hydroxy-3-metylglutaryl coenzyme A reductase, one of the most important players in cholesterol biosynthesis. In androgen-dependent prostate cancer cells, statins do not lower intracellular cholesterol levels by up-regulating the low density lipoprotein receptor in the same manner as in normal cells (6). Conversely, statins decrease intracellular cholesterol levels in androgen-independent prostate cancer cells, which cannot regulate low density lipoprotein receptor expression (6). In this study, we focused on the decrease in intracellular cholesterol levels following treatment with statins in CRPC. Recent reports have shown that de novo androgen synthesis is a therapeutic objective in CRPC (13). In the androgen synthesis pathway, cholesterol is the primary material. Therefore, we examined the effects of statins on androgen synthesis-related enzymes in CRPC cells.
Simvastatin increased the expression of AKR1C3 in PC-3 cells. AKR1C3 exhibits 3α-, 17β- and 20α-hydroxysteroid dehydrogenase activities (14). The expression of AKR1C3 is increased in several human cancers, including kidney (15) and breast (16). Concerning prostate cancer, localized, metastatic and recurrent prostate cancer has high levels of AKR1C3 (17–19). In addition, elevated AKR1C3 expression promotes the aggressiveness of PC-3 cells, which lack AR (7). These data indicate that increased levels of AKR1C3 induce prostate cancer progression not only by synthesizing intracellular androgen but also by androgen-independent mechanisms.
Medical agents such as non-steroidal anti-inflammatory drugs (NSAIDs), steroids, flavonoids, cyclopentane derivatives, and benzodiazepines inhibit AKR1C3 (20). One example of an NSAID is meclofenamic acid, which inhibits AKR1C3, cyclooxygenase-1, and cyclooxygenase-2 more strongly than other NSAIDs (21,22). Meclofenamic acid inhibits androgen-independent prostate cancer progression both in vitro and in vivo (23). In this study, AKR1C3 siRNA did not affect PC-3 cell proliferation and migration, whereas meclofenamic acid inhibited these processes, suggesting that meclofenamic acid also has anticancer effects without the AKR1C3 mechanism. The combination of simvastatin and meclofenamic acid inhibited PC-3 cell proliferation, migration, and Akt activation to a greater extent than simvastatin or meclofenamic acid alone. Previous reports have described combination therapy using statins and NSAIDs for the treatment of prostate cancer, which works by inhibiting NF-κB (24) or IL-6 (25). These results reveal that the inhibition of AKR1C3 is an underlying mechanism of the combination therapy of simvastatin and meclofenamic acid.
The present study had several limitations. First, three androgen-independent prostate cancer cell lines responded to simvastatin with different levels of AKR1C3 expression. Prostate cancer is very heterogenic. In particular, androgen-independent prostate cancer cells have a different genetic background (26). Therefore, our results may not be applicable to all CRPCs. In addition, we evaluated the effects of simvastatin and meclofenamic acid only in PC-3 cells, and studied the changes of gene expressions following simvastatin treatment only in one single time rather than performing a time-course experiment. Concerning Akt activation by IGF-1, we also checked only in one single time and IGF1R phosphorylation status was not evaluated. Moreover, in vivo models are required to show whether the combination of simvastatin and meclofenamic acid may have a curative influence on CRPC.
In summary, simvastatin increased AKR1C3 expression in androgen-independent prostate cancer cells, and the combination of simvastatin and meclofenamic acid further inhibited PC-3 cell proliferation, migration and Akt activation compared with simvastatin alone. These results suggest that the combination of statin and NSAIDs may be an effective strategy for the treatment of prostate cancer.
Acknowledgements
We thank Ms. Naomi Takase, Ms. Atsuko Oyama, and Ms. Hayumi Oyama for their technical assistance. This work was supported by JSPS KAKENHI Grant no. 25861410.
Glossary
Abbreviations
- AKR1C3
aldo-keto reductase family 1 member C3
- IGF1R
insulin-like growth factor 1 receptor
- CRPC
castration-resistant prostate cancer
- DHT
dihydrotestosterone
- LC-MS/MS
liquid chromatography coupled with tandem mass spectrometry
- FBS
fetal bovine serum
- IGF
insulin-like growth factor
- NSAIDs
non-steroidal anti-inflammatory drugs
References
- 1.Zhong S, Zhang X, Chen L, Ma T, Tang J, Zhao J. Statin use and mortality in cancer patients: Systematic review and meta-analysis of observational studies. Cancer Treat Rev. 2015;41:554–567. doi: 10.1016/j.ctrv.2015.04.005. [DOI] [PubMed] [Google Scholar]
- 2.Bansal D, Undela K, D'Cruz S, Schifano F. Statin use and risk of prostate cancer: A meta-analysis of observational studies. PLoS One. 2012;7:e46691. doi: 10.1371/journal.pone.0046691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Platz EA, Tangen CM, Goodman PJ, Till C, Parnes HL, Figg WD, Albanes D, Neuhouser ML, Klein EA, Lucia MS, et al. Statin drug use is not associated with prostate cancer risk in men who are regularly screened. J Urol. 2014;192:379–384. doi: 10.1016/j.juro.2014.01.095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Sekine Y, Furuya Y, Nishii M, Koike H, Matsui H, Suzuki K. Simvastatin inhibits the proliferation of human prostate cancer PC-3 cells via down-regulation of the insulin-like growth factor 1 receptor. Biochem Biophys Res Commun. 2008;372:356–361. doi: 10.1016/j.bbrc.2008.05.043. [DOI] [PubMed] [Google Scholar]
- 5.Miyazawa Y, Sekine Y, Kato H, Furuya Y, Koike H, Suzuki K. Simvastatin Up-regulates annexin A10 that can inhibit the proliferation, migration, and invasion in androgen-independent human prostate cancer cells. Prostate. 2017;77:337–349. doi: 10.1002/pros.23273. [DOI] [PubMed] [Google Scholar]
- 6.Furuya Y, Sekine Y, Kato H, Miyazawa Y, Koike H, Suzuki K. Low-density lipoprotein receptors play an important role in the inhibition of prostate cancer cell proliferation by statins. Prostate Int. 2016;4:56–60. doi: 10.1016/j.prnil.2016.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sobel RE, Sadar MD. Cell lines used in prostate cancer research: A compendium of old and new lines-part 1. J Urol. 2005;173:342–359. doi: 10.1097/01.ju.0000149989.01263.dc. [DOI] [PubMed] [Google Scholar]
- 8.Suzuki K, Koike H, Matsui H, Ono Y, Hasumi M, Nakazato H, Okugi H, Sekine Y, Oki K, Ito K, et al. Genistein, a soy isoflavone, induces glutathione peroxidase in the human prostate cancer cell lines LNCaP and PC-3. Int J Cancer. 2002;99:846–852. doi: 10.1002/ijc.10428. [DOI] [PubMed] [Google Scholar]
- 9.Dozmorov MG, Azzarello JT, Wren JD, Fung KM, Yang Q, Davis JS, Hurst RE, Culkin DJ, Penning TM, Lin HK. Elevated AKR1C3 expression promotes prostate cancer cell survival and prostate cell-mediated endothelial cell tube formation: Implications for prostate cancer progression. BMC Cancer. 2010;10:672. doi: 10.1186/1471-2407-10-672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Flanagan JU, Yosaatmadja Y, Teague RM, Chai MZ, Turnbull AP, Squire CJ. Crystal structures of three classes of non-steroidal anti-inflammatory drugs in complex with aldo-keto reductase 1C3. PLoS One. 2012;8:e43965. doi: 10.1371/journal.pone.0043965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zhuang L, Kim J, Adam RM, Solomon KR, Freeman MR. Cholesterol targeting alters lipid raft composition and cell survival in prostate cancer cells and xenografts. J Clin Invest. 2005;115:959–968. doi: 10.1172/JCI200519935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sivaprasad U, Abbas Tand, Dutta V. Differential efficacy of 3-hydroxy-3-methlyglutaryl CoA reductase inhibitors on the cell cycle of prostate cancer cells. Mol Cancer Ther. 2006;5:2310–2316. doi: 10.1158/1535-7163.MCT-06-0175. [DOI] [PubMed] [Google Scholar]
- 13.Cai C, Chen S, Ng P, Bubley GJ, Nelson PS, Mostaghel EA, Marck B, Matsumoto AM, Simon NI, Wang H, et al. Intratumoral de novo steroid synthesis activates androgen receptor in castration-resistant prostate cancer and is upregulated by treatment with CYP17A1 inhibitors. Cancer Res. 2011;71:6503–6513. doi: 10.1158/1538-7445.AM2011-LB-420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Matsuura K, Shiraishi H, Hara A, Sato K, Deyashiki Y, Ninomiya M, Sakai S. Identification of a principal mRNA species for human 3alpha-hydroxysteroid dehydrogenase isoform (AKR1C3) that exhibits high prostaglandin D2 11-ketoreductase activity. J Biochem. 1998;124:940–946. doi: 10.1093/oxfordjournals.jbchem.a022211. [DOI] [PubMed] [Google Scholar]
- 15.Azzarello JT, Lin HK, Gherezghiher A, Zakharov V, Yu Z, Kropp BP, Culkin DJ, Penning TM, Fung KM. Expression of AKR1C3 in renal cell carcinoma, papillary urothelial carcinoma, and Wilms' tumor. Int J Clin Exp Pathol. 2009;3:147–155. [PMC free article] [PubMed] [Google Scholar]
- 16.Byrns MC, Duan L, Lee SH, Blair IA, Penning TM. Aldo-keto reductase 1C3 expression in MCF-7 cells reveals roles in steroid hormone and prostaglandin metabolism that may explain its over-expression in breast cancer. J Steroid Biochem Mol Biol. 2010;118:177–187. doi: 10.1016/j.jsbmb.2009.12.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Nakamura Y, Suzuki T, Nakabayashi M, Endoh M, Sakamoto K, Mikami Y, Moriya T, Ito A, Takahashi S, Yamada S, et al. In situ androgen producing enzymes in human prostate cancer. Endocr Relat Cancer. 2005;12:101–107. doi: 10.1677/erc.1.00914. [DOI] [PubMed] [Google Scholar]
- 18.Stanbrough M, Bubley G, Ross K, Golub TR, Rubin MA, Penning TM, Febbo PG, Balk SP. Increased expression of genes converting adrenal androgens to testosterone in androgen-independent prostate cancer. Cancer Res. 2006;66:2815–2825. doi: 10.1158/0008-5472.CAN-05-4000. [DOI] [PubMed] [Google Scholar]
- 19.Wako K, Kawasaki T, Yamana K, Suzuki K, Jiang S, Umezu H, Nishiyama T, Takahashi K, Hamakubo T, Kodama T, Naito M. Expression of androgen receptor through androgen-converting enzymes is associated with biological aggressiveness in prostate cancer. J Clin Pathol. 2008;61:448–454. doi: 10.1136/jcp.2007.050906. [DOI] [PubMed] [Google Scholar]
- 20.Byrns MC, Jin Y, Penning TM. Inhibitors of type 5 17β-hydroxysteroid dehydrogenase (AKR1C3): Overview and structural insights. J Steroid Biochem Mol Biol. 2011;125:95–104. doi: 10.1016/j.jsbmb.2010.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kalgutkar AS, Crews BC, Rowlinson SW, Marnett AB, Kozak KR, Remmel RP, Marnett LJ. Biochemically based design of cyclooxygenase-2 (COX-2) inhibitors: Facile conversion of nonsteroidal antiinflammatory drugs to potent and highly selective COX-2 inhibitors; Proc Natl Acad Sci USA; 2000; pp. 925–930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ouellet M, Percival MD. Effect of inhibitor time-dependency on selectivity towards cyclooxygenase isoforms. Biochem J. 1995;306:247–251. doi: 10.1042/bj3060247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Soriano-Hernández AD, Galvan-Salazar HR, Montes-Galindo DA, Rodriguez-Hernandez A, Martinez-Martinez R, Guzman-Esquivel J, Valdez-Velazquez LL, Baltazar-Rodriguez LM, Espinoza-Gómez F, Rojas-Martinez A, et al. Antitumor effect of meclofenamic acid on human androgen-independent prostate cancer: A preclinical evaluation. Int Urol Nephrol. 2012;44:471–477. doi: 10.1007/s11255-011-0012-0. [DOI] [PubMed] [Google Scholar]
- 24.Zheng X, Cui XX, Avila GE, Huang MT, Liu Y, Patel J, Kong AN, Paulino R, Shih WJ, Lin Y, et al. Atorvastatin and celecoxib inhibit prostate PC-3 tumors in immunodeficient mice. Clin Cancer Res. 2007;13:5480–5487. doi: 10.1158/1078-0432.CCR-07-0242. [DOI] [PubMed] [Google Scholar]
- 25.Wang H, Cui XX, Goodin S, Ding N, Van Doren J, Du Z, Huang MT, Liu Y, Cheng X, Dipaola RS, et al. Inhibition of IL-6 expression in LNCaP prostate cancer cells by a combination of atorvastatin and celecoxib. Oncol Rep. 2014;31:835–841. doi: 10.3892/or.2013.2885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Fraser M, Berlin A, Bristow RG, van der Kwast T. Genomic, pathological, and clinical heterogeneity as drivers of personalized medicine in prostate cancer. Urol Oncol. 2015;33:85–94. doi: 10.1016/j.urolonc.2013.10.020. [DOI] [PubMed] [Google Scholar]