

Abstract
Borna disease virus (BDV) is a neurotropic and non-cytolytic virus, which causes behavioral disorders in a wide range of warm-blooded species. It is well established that BDV induces neurodegeneration by impairing neurogenesis and interfering with neuronal functioning in the limbic system. In the present study, the potential role of BDV infection in SH-SY5Y cells was identified, and comparisons of two original BDV strains (the human Hu-H1 and the laboratory Strain V) were performed to further elucidate the phenotypes of BDV pathogenesis with strain differences. Cell Counting Kit-8 and flow cytometric analyses revealed that the two BDV strain-infected groups exhibited marked anti-proliferation and cell cycle arrest compared with the control group, and the Hu-H1 strain caused more evident effects. However, the Hu-H1 strain did not exert effects on the apoptosis of SH-SH5Y cells, while Strain V led to a marked increase in apoptosis upon initial infection. Western blot analysis confirmed the upregulation of apoptosis regulator BAX protein and the downregulation of apoptosis regulator Bcl-2 protein caused by the two BDV strains. The results of the present study provided evidence that infection with BDV suppressed SH-SY5Y cellular functioning and exhibited divergent antiproliferative and apoptotic roles in cells between the two strains. The present study provided an insight for future investigation of strain differences and underlying pathomechanisms.
Keywords: apoptosis, Borna disease virus, cell cycle, proliferation, SH-SY5Y
Introduction
It is known that viruses trigger typically strong immunological responses to stop viral dissemination and resolve infection. However, certain viruses may cause persistent infections in the central nervous system (CNS) without an immunological reaction or symptoms of acute inflammation, including lymphocytic choriomeningitis virus and rabies (1,2). The mechanisms underlying persistent viruses, and the latent associations between brain function impairment and neurological or neuropsychiatric diseases, remain to be completely elucidated.
Borna disease virus (BDV), a single-stranded, non-cytolytic RNA virus with a non-segmented genome of ~8.9 kb with six open reading frames (3,4), is a highly neurotropic virus and causes persistent infections in the CNS of the infected individuals for their entire life. BDV was first reported as the causative agent of Borna disease, causing nonpurulent encephalomyelitis in horses in Germany at the end of the 19th century (5), and natural BDV has been observed to infect a series of warm-blooded species ranging from birds to non-human primates (6,7). In neonatal rats, BDV infection leads to severe neurodegeneration in the cortex and hippocampus, and generates neurodevelopmental abnormalities and complex behavioral changes. BDV infection has been demonstrated to be associated with human psychiatric diseases, including schizophrenia, mental disorders including learning difficulties, affective disorders and autism (8–10), and certain BDV-associated antibodies, antigens and RNAs have been identified in patients with neurological diseases, including Parkinson's disease, chronic fatigue syndrome, Guillain-Barre syndrome, viral encephalitis and multiple sclerosis (11). Increasing evidence has indicated that BDV infection may induce neuronal loss, gliocyte impairment and dysfunction in the development of neural stem/progenitor cells (12–14). However, it may be noted that differences in the genetic background of the host, viral strains and even viral proteins may exert different effects on apoptotic activity (15,16). Epidemiological studies have been performed to address the latent association between BDV infection and human neurological diseases (17,18), although the controversy of the possible association remains under debate and certain studies have reported an absence of these associations (19).
Hu-H1 and Strain V are two different BDV strains. Hu-H1 is a natural strain which was recovered in 1994 from the blood cells of a female patient with bipolar I disorder in Germany (4). The lab-derived Strain V is a non-natural strain, which was originated from the brain of a horse with fatal Borna disease in 1927 (20). Compared with Strain V, Hu-H1 exhibits few meaningful point mutations at the molecular level and a differing pathogenicity at the host level, including the induction behavioral changes in rabbits without fatal disease (21).
In previous studies, it was observed that BDV strains Hu-H1 and Strain V infected oligodendrocyte (OL) cells and rat neurons, leading to different alterations in cell proliferation, apoptosis and metabonomics (21,22). The present study sought to further investigate the pathomechanisms involved in SH-SY5Y cell dysfunction induced by various viral strains. The present study revealed that two BDV strains may serve a divergent antiproliferative and apoptotic role in SH-SY5Y cell and provided an insight for future examination of the strain differences and underlying pathomechanisms.
Materials and methods
Viral strains and cell culture
The virus strains and human OL cell lines were provided by Professor Hanns Ludwig and Professor Liv Bod of the Free University of Berlin (Berlin, Germany), and the viral solution was obtained by freezing and thawing the persistently BDV-infected human OL cells (OL cell lysates). The SH-SY5Y cell line (cat. no. SCSP-5014; Stem Cell Bank, Chinese Academy of Sciences, Shanghai, China) was cultured in Dulbecco's modified Eagle's medium (Hyclone; GE Healthcare Life Sciences, Logan, UT, USA) containing 5% fetal bovine serum (cat. no. 10099158; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 1% penicillin and 1% streptomycin, and was in a 5% CO2 incubator at 37°C.
Immunofluorescent staining
A total of 1×105 cells/well were seeded in six-well plates, and 200 µl volumes of the BDV strains were separately added to each well to form the infected cells following adherence overnight. A total of 2 h subsequent to infection, the medium was replaced with 3 ml new culture medium and culture was continued for 3 days. Following removal of the medium, the cells were fixed with 4% paraformaldehyde solution for 30 min at room temperature. As previously described (21), immunofluorescence staining was performed for BDV nucleoprotein detection. The primary antibody was a gift from Prof. Xie Peng of Chongqing University. The primary antibody rabbit anti-p40 (1:100) was incubated at 4°C overnight. Following rinsing three times with phosphate buffer, cells were incubated with fluorescein isothiocyanate (FITC)-conjugated anti-rabbit immunoglobulin G (1:200; cat. no. A0562, Beyotime Institute of Biotechnology, Haimen, China) in dark for 2 h at 37°C. Standard immunofluorescence staining and analysis was performed as described previously (21).
Cell proliferation and apoptosis analysis
A Cell Counting Kit-8 (CCK8) assay kit (Beyotime Institute of Biotechnology) was used for cell proliferation analysis. Cells were cultured in 96-well plates overnight at a density of 1,000 cells/well and were infected with BDV strains (multiplicity of infection, 4) as previously described (21). At 0–7 days post-infection, 10 µl CCK8 solution was added to each well for 2 h and the optical density was measured in triplicate using an ELISA reader (Bio-Rad Laboratories, Inc., Hercules, CA, USA) at λ=450 nm.
For the apoptosis analysis, cells were harvested following infection for 1, 3 and 5 days by trypsinization without EDTA, washed twice with cold phosphate buffer and stained with an annexin-FITC/propidium iodide (PI) apoptosis detection kit (Nanjing KeyGen Biotech Co., Ltd., Nanjing, China), according to the manufacturer's protocol. Apoptotic cells were assessed by flow cytometry and the results were recorded by matched Cell Quest software (version 5.1; BD Biosciences, San Jose, CA, USA). The experiment was repeated three times independently.
Cell cycle analysis
At 2 h post-infection with the BDV strains, cells were replaced with new medium without serum for 24 h to induce a G1 arrest, and were harvested at 0, 6, 15 and 24 h following the release of G1 arrest by adding 5% fetal bovine serum to the medium. All cells were fixed overnight using 70% ethanol at 4°C, followed by incubation with 100 U/ml RNase A and 50 µg/ml PI for 30 min at 37°C subsequent to washing twice with cold phosphate buffer. Analyses were performed on a flow cytometer by measurement of the percentage of cells in different phases of the cell cycle.
Western blot analysis
Western blotting was performed as previously described (19). Cell lysates were prepared from the cell sample using radioimmunoprecipitation assay lysis buffer (Beyotime Institute of Biotechnology) combined with a protease inhibitor. Protein concentrations were determined using a bicinchoninic acid protein assay kit (Thermo Fisher Scientific, Inc.). Proteins from each sample (40 µg) were separated by 10% SDS-PAGE, and transferred to polyvinylidene fluoride membranes (Merck KGaA, Darmstadt, Germany). Following blocking by 5% fetal bovine serum for 12 h at 4°C, the membranes were probed with the primary antibodies rabbit monoclonal anti-apoptosis regulator BAX (Bax; Sigma-Aldrich; Merck KGaA; cat. no. SAB5500012) and anti-apoptosis regulator Bcl-2 (Bcl-2; 1:1,000; Santa Cruz Biotechnology, Inc., Dallas, TX, USA), followed by incubation with horseradish peroxidase-conjugated goat anti-rabbit secondary antibody for 1 h at 25°C (1:1,000; cat. no. AB501-01A; NovoProtein Biotech Co., Ltd., Shanghai, China). A chemiluminescence detection kit (Beyotime Institute of Biotechnology) was employed to detect the labeled bands. β-actin (1:1,000; Sangon Biotech Co., Ltd., Shanghai, China) served as an internal reference and the relative protein expression of target proteins was determined. Image Pro Plus 6.0 software was used for densitometric analysis (Media Cybernetics, Inc., Rockville, MD, USA).
Statistical analysis
GraphPad Prism version 5.0 (GraphPad Software, Inc., La Jolla, CA, USA) was used for statistical analysis. All results are presented as the mean ± standard deviation of three independent experiments. The statistical analysis was performed using one-way analysis of variance with Tukey's post-hoc test. P<0.05 was considered to indicate a statistically significant difference.
Results
P40 protein assay in BDV-Infected SH-SY5Y
As a specific biomarker of BDV infection, BDV nucleoprotein P40 was testified in the Strain V and Hu-H1 cells by immunofluorescent staining to evaluate the permissivity of SH-SY5Y to BDV infection. On day 3 post-infection, the two BDV strain-infected groups exhibited fluorescent green focal points, stained positive for P40, while the observation of the control cells revealed no fluorescent expression (Fig. 1). The results suggested that Strain V and Hu-H1 successfully infected SH-SY5Y cells directly, as P40 is a specific marker of BDV infection.
Figure 1.

SH-SY5Y cells are permissive to BDV infection. Immunofluorescence staining is presented of SH-SY5Y with BDV nucleoprotein P40 (green) at day 3 post-infection. Scale bar, 100 µm. BDV, Borna disease virus.
BDV strain infection decreases SH-SY5Y cell viability
The CCK8 assay revealed a gradual increase in SH-SY5Y cell proliferation between days 1 and 7 in the three groups (Table I; Fig. 2). The control group exhibited significantly increased cell viability compared with the BDV strains groups (P<0.05), and the Hu-H1-infected cells exhibited a significantly decreased viability compared with the Strain V-infected cells (P<0.01).
Table I.
Cell viability (%) of control, Strain V and Hu-H1 cells (mean ± standard deviation; n=3).
| Group | P-value | |||||
|---|---|---|---|---|---|---|
| Day | Control | Strain V | Hu-H1 | Control vs. Strain V | Control vs. Hu-H1 | Strain V vs. Hu-H1 |
| 0 | 100.00±1.12 | 100.63±2.42 | 99.48±1.15 | ns | ns | ns |
| 1 | 137.92±0.96 | 130.68±2.75 | 122.63±0.45 | P<0.01 | P<0.001 | P<0.01 |
| 2 | 196.42±4.59 | 185.22±3.93 | 178.51±2.32 | P<0.05 | P<0.001 | P<0.05 |
| 3 | 237.75±3.97 | 225.40±4.42 | 212.97±1.57 | P<0.05 | P<0.001 | P<0.05 |
| 4 | 403.40±19.02 | 346.78±23.64 | 334.97±12.76 | P<0.05 | P<0.01 | ns |
| 5 | 429.43±9.41 | 399.82±12.82 | 383.18±9.49 | P<0.05 | P<0.01 | ns |
| 6 | 431.93±15.22 | 418.43±9.45 | 418.78±2.70 | ns | ns | ns |
| 7 | 438.91±13.56 | 430.32±11.31 | 426.57±3.05 | ns | ns | ns |
The cell viability of control cells at 0 days post-infection was designated as 100%. ns, not significant.
Figure 2.
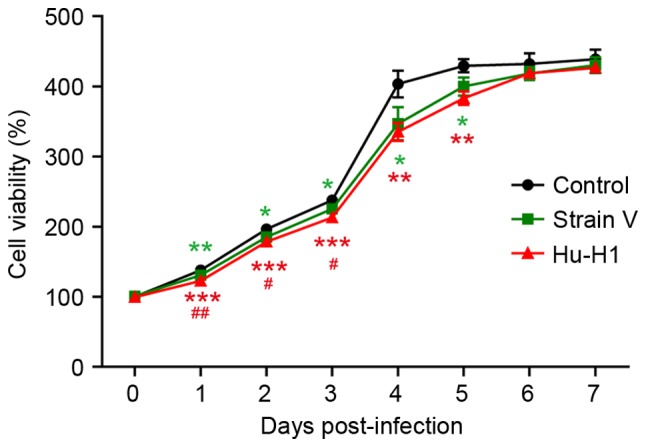
Infection with Borna disease virus strains decreases SH-SY5Y cell viability. Cell viability was determined by CCK8 assays at days 0–7 post-infection. Each data point is expressed as the mean ± standard deviation from independent experiments performed in triplicate. *P<0.05, **P<0.01, ***P<0.001 vs. control; #P<0.05, ##P<0.01 vs. Strain V. CCK8, Cell Counting Kit-8.
Strain V, although not the Hu-H1 strain, promotes apoptosis in SH-SY5Y cells
According to the protocol of annexin V-FITC/PI labeling, cellular apoptosis was determined by flow cytometry on days 1, 3 and 5 post-infection (Fig. 3A). There was no significant difference between the control and Hu-H1group (P>0.05). The Strain V group exhibited an increased apoptotic percentage compared with the control and Hu-H1groups on days 1 and 3 post-infection (P<0.05). However, on day 5 post-infection there was no statistically significant difference among the three groups (P>0.05). Therefore, only Strain V exerted an apoptotic effect at the initial stage of the assay (Fig. 3B-D). It may be suggested that Strain V induced cellular apoptosis upon initial infection, while the Hu-H1 strain exerted no effect on cellular apoptosis.
Figure 3.

Effect of Borna disease virus strain infection on SH-SY5Y cellular apoptosis. (A) An annexin V/PI apoptosis detection kit was used for staining the apoptotic cells and flow cytometry was performed on days (B) 1, (C) 3 and (D) 5 post-infection. The percentages of early, later and total apoptotic cells are displayed as histograms. The data are presented as the mean ± standard deviation from three independent experiments. *P<0.05, **P<0.01 vs. control. FITC, fluorescein isothiocyanate; PI, propidium iodide; ns, not significant.
BDV strain infection induces SH-SY5Y cell cycle arrest to different degrees
Following fixing with 70% ethanol overnight, the harvested cells were stained with PI and analyzed by flow cytometry to assay cell cycle distribution. It was observed that there was a larger percentage of cells in the G0/G1 phases and fewer cells in the S or G2/M phases when the cell cycle was arrested (Fig. 4A). At 0 h post-serum release, there was no significant difference among the three groups (P>0.05; Fig. 4B). Between 6 and 24 h post-serum release, the two BDV strains exhibited distinct cell cycle arrest compared with the control group (Fig. 4C-E). In addition, the Hu-H1 strain exerted a more significantly inhibitory effect compared with Strain V.
Figure 4.

Borna disease virus strains induce cell cycle arrest. (A) The harvested cells were stained with propidium iodide and assessed by flow cytometry at (B) 0, (C) 6, (D) 15 and (E) 24 h post-serum release. The data are expressed as the mean ± standard deviation from three independent experiments. *P<0.05, **P<0.01 vs. control. ns, not significant.
Different effects of BDV strain infection on apoptotic pathway protein expression
In order to investigate whether proteins in the apoptotic pathways were differently regulated, the relative expression of the apoptosis-associated proteins Bax and Bcl-2 were measured via western blotting (Fig. 5A). The western blot analysis results demonstrated that the Bax protein in BDV strain cells was upregulated compared with the control (P<0.01) on days 1–5 post-infection, particularly in Strain V cells (Fig. 5B). By contrast, Bcl-2, the anti-apoptotic protein, exhibited decreased levels in Strain V cells compared with corresponding controls (P<0.01), and there was no significant difference between Hu-H1 strains and controls until 5 days post-infection (P>0.05) (Fig. 5C).
Figure 5.

Relative expression of apoptotic proteins in Borna disease virus-infected cells. (A) Western blot analysis of Bax and Bcl-2 in Strain V and Hu-H1 infected SH-SY5Y cells was performed at day 1, 3 and 5 post-infection. Densitometric analysis was performed using Image Pro Plus 6.0 software and the protein expression levels of (B) Bax and (C) Bcl-2 were normalized to β-actin. The data are expressed as the mean ± standard deviation from three independent experiments, *P<0.05, **P<0.01, ***P<0.001 vs. control. Bax, apoptosis regulator BAX; Bcl-2, apoptosis regulator Bcl-2; ns, not significant.
Discussion
As a neurotropic virus, which causes CNS dysfunction in a wide range of warm-blooded species, BDV may lead to persistent infection and biomolecular alterations in neurological cells (17,23). Based on the cellular phenotype of SH-SY5Y cells, the present study revealed different proliferative and apoptotic effects that distinguished among the non-infected control, BDV Strain V- and Hu-H1-infected cells for the first time, to the best of our knowledge. The results of the present study suggested that the biological functions of natural BDV Hu-H1 and non-natural BDV Strain V were different due to properties of the hosts, and provided evidence for biological strain differences. The present study may provide a basis for biological characterization and an insight into taxonomical issues at the species level.
The results of the present study indicated that BDV strains were able to inhibit SH-SY5Y cellular proliferation and obstruct the cell cycle, and that human Hu-H1 exerted stronger inhibitory effects compared with Strain V. A previous study demonstrated that Strain V was able to promote OL cell proliferation while Hu-H1 inhibited OL cell proliferation (21), and Planz et al (24) reported that BDV nucleoprotein He/80 interacted with the cyclin-dependent kinase 1-cyclin B1 complex and prolonged the G2 cell cycle phase. These previous results implied that the virus may directly or indirectly regulate certain cell cycle regulatory proteins, cytokines or growth factors during the proliferation and cell cycle of SH-SY5Y cells to execute its inhibitory effects. The Hu-H1 strain has been demonstrated to undergo a number of species crossings in animals and tissue culture cells (25), thus may be exhibiting divergent influences on different human cells. In addition, although the molecules binding to the viral proteins may be the same in different cell lines, virus strains may affect different signaling or metabolic pathways, as revealed in a metabonomic study of the two BDV strains in rat cortical neurons (21); elucidating the differences in specific mechanisms requires further investigation.
The BDV non-natural Strain V exerted significant, although temporary, effects on the apoptosis of SH-SY5Y cells, while the Hu-H1 strain exerted almost no effect. In order to examine the underlying mechanisms of apoptosis, the relative expression of Bax and Bcl-2 protein was detected by western blot analysis. The results of the present study demonstrated that Bax and Bcl-2 proteins were involved in the SH-SY5Y cellular apoptosis induced by BDV initial infection, and the Hu-H1 strain also promoted apoptosis in the infected OL cells, while laboratory strain V was demonstrated to inhibit cell apoptosis. In addition, Wu et al (16) reported that granule cell neurons of the dentate gyrus were not affected in Sprague-Dawley rats with BDV infection. Therefore, different strains may cause unpredictable apoptotic effects in different hosts. The different inherent properties of BDV strains may explain why they may cause acute nonpurulent encephalomyelitis in animals and chronic asymptomatic infection in humans.
Based on the genomic and protein sequences (26), the taxonomy within the family of the Bornaviridae is intuitive and comprehensible. However, even with high conservation of BDV sequence, strains may cause different effects between hosts. Therefore, the host phenotype alone is inadequate to explain the mechanisms of action of BDV strains. Studies into the acetylation and metabonomics of BDV infection (20,27) have provided a novel perspective for understanding the pathogenic mechanisms of BDV infection. However, further analysis of biological functions and systematic interpretations are required to better define BDV strains in future studies.
References
- 1.De la Torre JC, Mallory M, Brot M, Gold L, Koob G, Oldstone MB, Masliah E. Viral persistence in neurons alters synaptic plasticity and cognitive functions without destruction of brain cells. Virology. 1996;220:508–515. doi: 10.1006/viro.1996.0340. [DOI] [PubMed] [Google Scholar]
- 2.Dietzschold B, Li J, Faber M, Schnell M. Concepts in the pathogenesis of rabies. Future Virol. 2008;3:481–490. doi: 10.2217/17460794.3.5.481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Cubitt B, Oldstone C, de la Torre JC. Sequence and genome organization of Borna disease virus. J Virol. 1994;68:1382–1396. doi: 10.1128/jvi.68.3.1382-1396.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Briese T, Schneemann A, Lewis AJ, Park YS, Kim S, Ludwig H, Lipkin WI. Genomic organization of Borna disease virus. Proc Natl Acad Sci USA. 1994;91:4362–4366. doi: 10.1073/pnas.91.10.4362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Dürrwald R, Ludwig H. Borna disease virus (BDV), a (zoonotic?) worldwide pathogen. A review of the history of the disease and the virus infection with comprehensive bibliography. Zentralbl Veterinarmed B. 1997;44:147–184. doi: 10.1111/j.1439-0450.1997.tb00962.x. [DOI] [PubMed] [Google Scholar]
- 6.Bode L, Dürrwald R, Ludwig H. Borna virus infections in cattle associated with fatal neurological disease. Vet Rec. 1994;135:283–284. doi: 10.1136/vr.135.12.283. [DOI] [PubMed] [Google Scholar]
- 7.Hagiwara K, Tsuge Y, Asakawa M, Kabaya H, Okamoto M, Miyasho T, Taniyama H, Ishihara C, de la Torre JC, Ikuta K. Borna disease virus RNA detected in Japanese macaques (Macaca fuscata) Primates. 2008;49:57–64. doi: 10.1007/s10329-007-0068-8. [DOI] [PubMed] [Google Scholar]
- 8.Hornig M, Solbrig M, Horscroft N, Weissenböck H, Lipkin WI. Borna disease virus infection of adult and neonatal rats: Models for neuropsychiatric disease. Curr Top Microbiol Immunol. 2001;253:157–177. doi: 10.1007/978-3-662-10356-2_8. [DOI] [PubMed] [Google Scholar]
- 9.Pletnikov MV, Moran TH, Carbone KM. Borna disease virus infection of the neonatal rat: Developmental brain injury model of autism spectrum disorders. Front Biosci. 2002;7:d593–d607. doi: 10.2741/pletnik. [DOI] [PubMed] [Google Scholar]
- 10.Williams BL, Hornig M, Yaddanapudi K, Lipkin WI. Hippocampal poly(ADP-Ribose) polymerase 1 and caspase 3 activation in neonatal bornavirus infection. J Virol. 2008;82:1748–1758. doi: 10.1128/JVI.02014-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zhang L, Xu MM, Zeng L, Liu S, Liu X, Wang X, Li D, Huang RZ, Zhao LB, Zhan QL, et al. Evidence for Borna disease virus infection in neuropsychiatric patients in three western China provinces. Eur J Clin Microbiol Infect Dis. 2014;33:621–627. doi: 10.1007/s10096-013-1996-4. [DOI] [PubMed] [Google Scholar]
- 12.Das S, Basu A. Viral infection and neural stem/progenitor cell's fate: Implications in brain development and neurological disorders. Neurochem Int. 2011;59:357–366. doi: 10.1016/j.neuint.2011.02.020. [DOI] [PubMed] [Google Scholar]
- 13.Brnic D, Stevanovic V, Cochet M, Agier C, Richardson J, Montero-Menei CN, Milhavet O, Eloit M, Coulpier M. Borna disease virus infects human neural progenitor cells and impairs neurogenesis. J Virol. 2012;86:2512–2522. doi: 10.1128/JVI.05663-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Scordel C, Huttin A, Cochet-Bernoin M, Szelechowski M, Poulet A, Richardson J, Benchoua A, Gonzalez-Dunia D, Eloit M, Coulpier M. Borna disease virus phosphoprotein impairs the developmental program controlling neurogenesis and reduces human GABAergic neurogenesis. PLoS Pathog. 2015;11:e1004859. doi: 10.1371/journal.ppat.1004859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Poenisch M, Burger N, Staeheli P, Bauer G, Schneider U. Protein X of Borna disease virus inhibits apoptosis and promotes viral persistence in the central nervous systems of newborn-infected rats. J Virol. 2009;83:4297–4307. doi: 10.1128/JVI.02321-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wu YJ, Schulz H, Lin CC, Saar K, Patone G, Fischer H, Hübner N, Heimrich B, Schwemmle M. Borna disease virus-induced neuronal degeneration dependent on host genetic background and prevented by soluble factors. Proc Natl Acad Sci USA. 2013;110:1899–1904. doi: 10.1073/pnas.1214939110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Li Q, Wang Z, Zhu D, Xu M, Chen X, Peng D, Iwata Y, Xie P. Detection and analysis of Borna disease virus in Chinese patients with neurological disorders. Eur J Neurol. 2009;16:399–403. doi: 10.1111/j.1468-1331.2008.02516.x. [DOI] [PubMed] [Google Scholar]
- 18.Wang X, Zhang L, Lei Y, Liu X, Zhou X, Liu Y, Wang M, Yang L, Zhang L, Fan S, Xie P. Meta-analysis of infectious agents and depression. Sci Rep. 2014;4:4530. doi: 10.1038/srep04530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hornig M, Briese T, Licinio J, Khabbaz RF, Altshuler LL, Potkin SG, Schwemmle M, Siemetzki U, Mintz J, Honkavuori K, et al. Absence of evidence for bornavirus infection in schizophrenia, bipolar disorder and major depressive disorder. Mol Psychiatr. 2012;17:486–493. doi: 10.1038/mp.2011.179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zwick W. Neuere Untersuchungen über die seuchenhafte Gehirn-u. Rückenmarksentzündung (Borna'sche Krankheit) der Pferde. Dtsch. Z. Nervenheilk. 1929;110:316–322. doi: 10.1007/BF01652993. [DOI] [Google Scholar]
- 21.Liu S, Bode L, Zhang L, He P, Huang R, Sun L, Chen S, Zhang H, Guo Y, Zhou J, et al. GC-MS-Based metabonomic profiling displayed differing effects of Borna Disease Virus natural strain Hu-H1 and laboratory Strain V infection in rat cortical neurons. Int J Mol Sci. 2015;16:19347–19368. doi: 10.3390/ijms160819347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Li D, Lei Y, Deng J, Zhou C, Zhang Y, Li W, Huang H, Cheng S, Zhang H, Zhang L, et al. Human but not laboratory Borna Disease Virus inhibits proliferation and induces apoptosis in human oligodendrocytes in vitro. PLoS One. 2013;8:e66623. doi: 10.1371/journal.pone.0066623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Huang R, Gao H, Zhang L, Jia J, Liu X, Zheng P, Ma L, Li W, Deng J, Wang X, et al. Borna disease virus infection perturbs energy metabolites and amino acids in cultured human oligodendroglia cells. PLoS One. 2012;7:e44665. doi: 10.1371/journal.pone.0044665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Planz O, Pleschka S, Oesterle K, Berberich-Siebelt F, Ehrhardt C, Stitz L, Ludwig S. Borna disease virus nucleoprotein interacts with the CDC2-cyclin B1 complex. J Virol. 2003;77:11186–11192. doi: 10.1128/JVI.77.20.11186-11192.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Ludwig H, Furuya K, Bode L, Klein N, Dürrwald R, Lee DS. Biology and neurobiology of Borna disease viruses (BDV), defined by antibodies, neutralizability and their pathogenic potential. Arch Virol Suppl. 1993;7:111–133. doi: 10.1007/978-3-7091-9300-6_10. [DOI] [PubMed] [Google Scholar]
- 26.Kuhn JH, Dürrwald R, Bào Y, Briese T, Carbone K, Clawson AN, deRisi JL, Garten W, Jahrling PB, Kolodziejek J, et al. Taxonomic reorganization of the family Bornaviridae. Arch Virol. 2015;160:621–632. doi: 10.1007/s00705-014-2276-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Liu X, Liu S, Bode L, Liu C, Zhang L, Wang X, Li D, Lei Y, Peng X, Cheng Z, Xie P. Persistent human Borna disease virus infection modifies the acetylome of human oligodendroglia cells towards higher energy and transporter levels. Virology. 2015;485:58–78. doi: 10.1016/j.virol.2015.06.024. [DOI] [PubMed] [Google Scholar]