

Abstract
Small CAB-like proteins (SCPs) are single-helix light-harvesting-like proteins found in all organisms performing oxygenic photosynthesis. We investigated the effect of growth in moderate salt stress on these stress-induced proteins in the cyanobacterium Synechocystis sp. PCC 6803 depleted of Photosystem I (PSI), which expresses SCPs constitutively, and compared these cells with a PSI-less/ScpABCDE− mutant. SCPs, by stabilizing chlorophyll-binding proteins and Photosystem II (PSII) assembly, protect PSII from photoinhibitory damages, and in their absence electrons accumulate and will lead to ROS formation. The presence of 0.2 M NaCl in the growth medium increased the respiratory activity and other PSII electron sinks in the PSI-less/ScpABCDE− strain. We postulate that this salt-induced effect consumes the excess of PSII-generated electrons, reduces the pressure of the electron transport chain, and thereby prevents 1O2 production.
Electronic supplementary material
The online version of this article (doi:10.1007/s11120-017-0426-3) contains supplementary material, which is available to authorized users.
Keywords: Small CAB-like proteins (SCPs), Photosystem II photoinhibition, Singlet oxygen, Salt stress, Terminal oxidases
Introduction
In oxygenic photosynthesis, light energy is used to drive two specialized protein complexes called Photosystem I (PSI) and Photosystem II (PSII), which are embedded in a special membrane system, the thylakoid membrane. Pigmented antenna proteins collect and transfer the sunlight to the photosystem reaction centers, where charge separation takes place. Concomitant with the electron transfer via the photosystems, protons are pumped through membrane-inserted complexes and create a chemo-osmotic gradient, required for the synthesis of ATP. ATP and NADPH are finally used in the biochemical reduction of carbon, nitrogen, and sulfur. In cyanobacteria, thylakoid membranes are not just the site of photosynthesis, but also of respiration (Vermaas 2001). Respiration and photosynthesis share many components including plastoquinones (PQ), the cytochrome b 6 f (Cyt b 6 f), and the two soluble redox carriers, plastocyanin (PC) and Cyt c 6. In the model cyanobacterium Synechocystis sp. PCC 6803 (hereafter Synechocystis), PQ can be reduced either by PSII or by the NADP(H) dehydrogenase-like complex type I (NDH-1), the succinate dehydrogenase (SDH), and different NDH-2s (Cooley and Vermaas 2001). Electrons are transferred directly from plastoquinol (PQH2) to the bd quinol oxidases (Cyd; Berry et al. 2002) or via Cyt b 6 f to PSI or an aa3-type cytochrome c oxidase (Cox) complex (Howitt and Vermaas 1998; Ermakova et al. 2016). In addition, there is the alternative respiratory terminal oxidase (ARTO). The NDH-1, SDH, and NDH-2 complexes and Cyd are ubiquitous in the thylakoid and the cytoplasmic membranes, while Cox and ARTO have been located only in the thylakoid or cytoplasmic membrane, respectively.
Although light is essential for photosynthesis, an excess of light energy can damage the photosynthetic apparatus in a process called photoinhibition, where PSII is the major site of damage (Aro et al. 1993). Excess of energy that cannot be used to drive photosynthesis enhances the production of reactive oxygen species (ROS) and induces photooxidative damages. A ROS marker for PSII damage is singlet oxygen (1O2) that is produced by the reaction of excited chlorophyll in its triplet state (3Chl*) with O2. When absorbed light cannot be fully utilized for electron transfer reactions, the probability of 1O2 formation increases. To dissipate excess excitation, photosynthetic organisms have developed different mechanisms. The fastest mechanisms are generally known as nonphotochemical quenching (NPQ; Niyogi 1999; Kirilovsky and Kerfeld 2012). They occur at the antenna level and they function by converting the excess light energy into heat through carotenoid molecules. In Synechocystis, an orange carotenoid protein (OCP) has been found to be the main component responsible for NPQ. OCP binds a single xanthophyll, 3′-hydroxyechinenone, whose absorption of blue-green light induces a conformational change of the protein that converts from an inactive form (OCPO) to an active red form (OCPr). OCPr then binds to the phycobilisomes, the antenna of most species of cyanobacteria, quenching excess energy (for review see Kirilovsky and Kerfeld 2012).
An alternative to photoprotection via NPQ is to increase the electron sink capacity, i.e., enhance the capacity to consume photochemically generated electrons through the reduction of CO2 and O2. Several alternative electron pathways have been identified in cyanobacteria, higher plants, and algae involved in the dissipation of excess of light (McDonald et al. 2011; Chaux et al. 2015; Peltier et al. 2010; Roach and Krieger-Liszkay 2014).
Synechocystis contains five small CAB-like proteins (SCPs, named ScpA–E; Funk and Vermaas 1999, or also called high-light-induced proteins, HLIPs; Dolganov et al. 1995), which are induced during general stress, including light stress (He et al. 2001). SCPs consist of a single membrane-spanning helix, which has high homology to the first and third transmembrane regions of the higher plant light-harvesting complex. ScpA is the C-terminal extension of the ferrochelatase HemH, an enzyme involved in heme biosynthesis (Funk and Vermaas 1999; Sobotka et al. 2008, 2011; Storm et al. 2013); ScpB–E are proteins of around 6 kDa and have been found to be associated with PSII (Promnares et al. 2006; Yao et al. 2007; Kufryk et al. 2008; Shi et al. 2012). SCPs have been proposed to function in exciton dissipation (Havaux et al. 2003), to act as chlorophyll (Chl) carriers during assembly/repair of PSII (Knoppová et al. 2014; Hernandez-Prieto et al. 2011; Yao et al. 2012), or to regulate Chl biosynthesis (Xu et al. 2002, 2004).
While in wild-type Synechocystis SCPs only are expressed during stress, in a PSI-less background strain (Shen et al. 1993) they are expressed constitutively (Funk and Vermaas 1999). The PSI-less/ScpABCDE− mutant appears chlorotic compared to the control PSI-less strain due to decreased PSII content, and glycogen accumulates in the cell (Hernandez-Prieto et al. 2011; Tibiletti et al. 2016). It has been proposed that the absence of SCPs decreases the stability of the Chl-binding proteins within PSII and leads to the formation of ROS (Hernandez-Prieto et al. 2011; Sinha et al. 2012).
Comparing the PSI-less/ScpABCDE− mutant with the SCP-expressing PSI-less control strain, here we show that in the presence of 0.2 M sodium chloride in the growth medium the control phenotype is restored in the PSI-less/ScpABCDE− mutant. We propose that SCPs protect PSII from photoinhibitory damages by decreasing the 1O2 production.
Materials and methods
Growth conditions, cell counting, and measurement of cell size
For each experiment, Synechocystis PSI-less (Shen et al. 1993) and PSI-less/ScpABCDE− (Xu et al. 2004) mutants were first plated freshly from frozen stock cultures and then cultivated in flasks with BG-11 medium (Rippka et al. 1979) shaken at 100 rpm at 28 °C and low light intensity (4–5 μmol photons m−2 s−1). Different frozen stocks were used for the biological replicates to avoid effects of secondary mutations. The liquid growth medium was supplemented with 10 mM glucose and 10 mM TES–NaOH, pH 8.0. For growing on plates, solid BG-11 medium was supplemented with 10 mM glucose, 10 mM TES–NaOH, pH 8.2, and 20 mM Na-thiosulfate.
Cell precultures were inoculated in liquid BG-11 medium supplemented with 10 mM glucose in the presence or absence of 0.2 M NaCl and allowed to acclimate for 3 days (corresponding roughly to four generations). In exponential logarithmic growth, an inoculum of these precultures was then used to inoculate fresh medium (in the presence or absence of 0.2 M NaCl) at an optical density at 730nm (OD730) of 0.2 for cultures used in the experiments. The concentration of 0.2 M sodium chloride in the medium to induce salt stress was chosen based on the study by Howitt et al. (2001) determining the salt tolerance of different Synechocystis mutants. The influence of pH on the mutants was tested by adding 20 mM TES–NaOH to obtain pH 8, 20 mM of CAPS (pH 9), or 20 mM of MES–NaOH to obtain pH 6.5 to the medium. A final concentration 0.5 mM NaHCO3 was added when indicated in a buffered (pH 8) BG-11 medium supplemented with glucose. In this case, cells precultured in liquid BG-11 medium supplemented with 10 mM glucose were inoculated in media with different pH or in the presence of NaHCO3. Each experiment was performed with three biological replicates. To ensure that the culture was in exponential growth phase at the beginning of the experiment, the cells were counted using a Neubauer improved chamber. Growth of the cultures was also monitored by measuring the OD730 using a T90+ spectrophotometer (PG Instruments), OD730 0.5–0.8 corresponding to a logarithmic growth phase. Cell size was measured using the Multisizer™ 4 Coulter CounterR (Beckman Coulter).
Pigment determination
Chl was extracted from total cells using 100% methanol and its concentration was determined from the absorbance at 665 nm using the extinction coefficient for Chl a in methanol of 71.43 mM−1 cm−1 taken by Porra (2002). The carotenoid/Chl (Car/Chl a) ratio was calculated from the pigment absorbance at 664 nm (for Chl a) and 474 nm (for carotenoids).
Low-temperature fluorescence
Low-temperature fluorescence spectra were recorded using a FluoroMax-2 fluorometer (Spex spectrofluorometer system, Jobin Yvon, Longjumeau, France). Cells after growth in the presence or absence of NaCl were concentrated to 1 mg Chl a/mL and were frozen without glycerol to avoid functional uncoupling of the phycobilisomes from thylakoid components. The obtained spectra were normalized to their maximum values (Shen and Vermaas 1994).
SDS-PAGE and immunoblot analysis
An equal number of cells were pelleted. After Chl extraction, the proteins were resuspended in SDS-loading buffer (245 mM Tris–HCl, pH 8.2, 0.5 mM ethylenediaminetetraacetic acid (EDTA), 2% lithium dodecyl sulfate (LDS), 10% glycerol, and 50 mM dithiothreitol (DTT)) and heated at 65 °C for 15 min. After the removal of unsolubilized proteins by centrifugation (20,000 g for 10 min), the sample was loaded onto a sodium dodecyl sulfate (SDS)-acrylamide gel with 6 M urea. Immunoblotting using antibodies against the PSII proteins D1, CP47, and PsbH, as well as against ScpC/ScpD and ScpE, was performed as described in Hernandez-Prieto et al. (2011).
Oxygen evolution and respiration measurements
In cultures with an OD730 of 0.6, the electron transport activity of PSII and the dark respiration were measured at 30 °C with a Clark-type oxygen electrode (Hansatech Instrument). The oxygen-evolving activity of PSII was measured in BG-11 medium supplemented with 10 mM glucose in the presence of 600 μM 2,6-dichloro-p-benzoquinone (DCBQ) and 1.2 mM potassium ferricyanide (FeCN) under saturating light intensity (2000 μmol photons m−2 s−1).
For respiration measurements, an early logarithmic culture was harvested and resuspended in fresh BG-11 containing 10 mM glucose. The cell density was adjusted to a chlorophyll concentration of 10 μM. 1 mL of this concentrated culture was assayed for the capacity to consume O2 in the dark. 15 mM potassium cyanide (KCN) was added to the culture to inhibit the respiratory activity and to obtain the baseline, following Wang et al. (2012).
Histidine-mediated oxygen uptake measurement
For quantification of singlet oxygen production, histidine-mediated oxygen uptake measurements were applied, which are based on the oxidation of histidine by 1O2. The subsequent removal of dissolved oxygen in aqueous media was detected by a standard oxygen electrode (Rehman et al. 2013). The rate of singlet oxygen-induced oxygen uptake was measured in the presence of 5 mM His at 2300 µmol photons m−2 s−1 light intensity using a Hansatech DW2 O2 electrode at a Chl concentration of 2 µg mL−1 in the absence of artificial electron acceptors as described earlier (Rehman et al. 2013). Before the O2 uptake measurements, Synechocystis cells were centrifuged and resuspended in fresh BG-11 medium.
Photoinhibition measurements
Photoinhibition measurements were performed as described in Hernandez-Prieto et al. (2011) and Rehman et al. (2013). High-light illumination experiments were performed in open, square glass containers, in which the cell suspension formed a 14-mm-high layer, with continuous stirring at 25 °C. An array of 50 W halogen lamps with adjustable light intensities provided the homogenous white light illumination of 200 µmol photons m−2 s−1. PSII activity was assayed by the initial amplitude of the flash-induced Chl fluorescence signal.
Attenuated total reflection Fourier transform infrared (ATR FT-IR) spectroscopy
A volume of 25 mL of cells was harvested by centrifugation at 4000 rpm and washed twice with water. The cell pellet was frozen in liquid nitrogen, freeze-dried, and stored until analysis. FT-IR measurements were carried out with a Tensor 27 Spectrophotometer (Bruker). A tip of a spatula of freeze-dried cells was resuspended in water and applied directly on a crystal surface. After drying under air flux, 500 scans over 7 min were recorded for each sample, within the range of 3500–800 cm−1 at a resolution of 8 cm−1. A background spectrum was recorded before each sample measurement and automatically subtracted from the sample spectrum. Spectra were cut to 1850–800 cm−1, baseline-corrected, and normalized to the total peak area. Spectra were analyzed using OPUS v. 5.5 software.
Statistics
Data are reported as the mean ± standard deviation of three or more biological replicates. Statistical significance of differences among the means was determined by an unpaired two-tailed t test when only two groups were compared, or by one-way analysis of variance (ANOVA) and Tukey’s post hoc test when more than two groups were compared. GraphPad Prism 4.03 software (GraphPad Software, San Diego, CA, USA) was used, with the level of significance set at 95%. P values were classified as follows: <0.001 extremely significant***, 0.001–0.01 very significant**, and 0.01–0.05 significant*.
Results
NaCl induces pigment biosynthesis and PSII assembly in the PSI-less/ScpABCDE−strain
The SCP-expressing PSI-less mutant and the PSI-less/ScpABCDE− strain were cultured in BG-11 medium supplemented with 10 mM glucose in the presence or absence of 0.2 M NaCl. Under control growth conditions (4 µmol photons m−2 s−2 in continuous light at 30 °C, absence of NaCl), the PSI-less/ScpABCDE− strain appeared chlorotic compared to the PSI-less control strain (Fig. 1). In the presence of 0.2 M NaCl, however, the pigmentation was affected in both strains with higher impact on the PSI-less/ScpABCDE− mutant, as could be observed by eye (Fig. 1) and by measuring the pigment absorption (Table 1). On cell basis, the amount of Chl a increased by 4.6 times in the PSI-less/ScpABCDE− strain (p < 0.001), while the amount of Chl a in the PSI-less control doubled (p < 0.05, Table 1). The cell division time of the PSI-less/ScpABCDE− strain was not significantly altered in the presence of NaCl (p > 0.05, Table 1), while doubling of the PSI-less control strain decreased 1.6 times (p < 0.05, Table 1).
Fig. 1.

Color appearance of the PSI-less and PSI-less/ScpABCDE− mutants grown in the presence or absence of 0.2 M NaCl. Cell cultures of PSI-less and PSI-less/ScpABCDE− strains were in BG-11 supplemented with glucose in the presence or absence of 0.2 M NaCl as described in Material and Methods. The photo was taken after dilution at OD730 0.4–0.5. Representative cultures of three biological replicates are shown
Table 1.
Comparison of the PSI-less/ScpABCDE− mutant and the PSI-less control strain grown in the presence or absence of 0.2 M NaCl
| Division time (hours) | Cell diameter (μm) | nmol Chl a mL−1 OD730 | fmol Chl a cell−1 | Car/Chl a | O2 evolution fmol O2 h−1 cell−1 | Respiration fmol O2 consumption h−1 cell−1 | |
|---|---|---|---|---|---|---|---|
| PSI-less | 27.38 (± 1.33) |
1.67 (± 0.03) |
0.63 (± 0.01) |
3.92 (± 0.05) |
2.11 (± 0.01) |
13.99 (± 1.57) |
0.82 (± 0.06) |
| PSI-less+NaCl | 43.4* (± 4.1) |
1.7 (± 0.01) |
0.87*** (± 0.02) |
8.22* (± 0.27) |
1.51*** (± 0.02) |
33.52** (± 7.28) |
1.08 (± 0.15) |
| PSI-less/ScpABCDE− | 37.59 (± 3.74) |
2.13*** (± 0.03) |
0.23*** (± 0.01) |
3.68 (± 0.03) |
3.36*** (± 0.16) |
8.81 (± 2.03) |
1.63*** (± 0.10) |
| PSI-less/ScpABCDE−+NaCl | 44.6 (± 11.67) |
2.23** (± 0.04) |
0.67*** (± 0.04) |
16.85*** (± 3.61) |
1.78*** (± 0.1) |
47.72*** (± 4.46) |
3.04*** (± 0.18) |
Both strains were cultivated in BG-11 standard medium supplemented with 10 mM glucose. Standard deviations with n ≥ 3 are given in brackets
P values are classified as: *0.01 to 0.05 significant; **0.001 to 0.01 very significant; ***< 0.001 extremely significant
Both strains, the PSI-less/ScpABCDE− and the PSI-less control, contained more Chl when grown in the presence of NaCl. Free Chl is potentially damaging the cell and therefore scavengers like a carotenoid molecule have to be in its proximity. To analyze if the NaCl-induced Chl molecules were used to assemble additional PSII complexes, oxygen evolution measurements were performed using a Clark electrode (Table 1). Indeed, cells grown in BG-11 supplemented with NaCl evolved more oxygen; while more than five times O2 h−1 was produced per cell by the PSI-less/ScpABCDE− strain grown in the presence of NaCl (p < 0.001, Table 1), oxygen evolution of the PSI-less culture only slightly increased. Immunoblotting experiments were performed to confirm the biosynthesis of more PSII complexes per cell (Fig. 2). Proteins were extracted from the same number of cells of the PSI-less/ScpABCDE− and the PSI-less control strain, which had been grown in the presence or absence of NaCl for 4 days. After SDS-PAGE, several proteins were immunostained with specific antibodies: the PSII reaction center protein D1, the PSII core antenna protein CP47, and the low-molecular weight PSII protein PsbH. Additionally, antibodies recognizing ScpC/ScpD or ScpE were used. In untreated cells, the PSI-less/ScpABCDE− strain contained very few PSII centers (judged by the presence of D1, CP47, and PsbH) per cell (Fig. 2), confirming earlier results (Hernandez-Prieto et al. 2011). In the PSI-less control, more PSII proteins were detected, while the SCPs were strongly expressed (Funk and Vermaas 1999). Presence of NaCl further stimulated SCP expression in the control strain with ScpE accumulating stronger per cell than ScpC/D. Interestingly, while the presence of NaCl induced an increasing amount of PSII proteins in both strains, its impact in the PSI-less/ScpABCDE− mutant per cell was much stronger compared to the PSI-less control.
Fig. 2.

Immunodetection of the PSII proteins D1, CP47, and PsbH, as well as of ScpC/D and ScpE, in the PSI-less and the PSI-less/ScpABCDE− strains, when grown in the presence or absence of 0.2 M NaCl. Proteins extracted from the same number of cells were loaded in each lane. A representative immunoblot of three biological replicates is shown
Low-temperature (77 K) fluorescence emission spectra offer good resolution of the fluorescence produced by PSII components. Additionally, selection of the emission wavelength allows relative quantification of the ratio between antenna and photosystem pigments. Upon excitation at 435 nm, which excites mainly Chl a, two peaks with maximum at 685 and 695 nm were observed. The peak at 685 nm reflects all Chls associated with PSII (including Chl from CP43, D2, D1, and CP47), except for one low-energy Chl a 627 that appears to be associated with His114 of CP47. This red low-energy Chl is the main contributor to the 695 nm emission maximum (Vermaas et al. 1986; Shen and Vermaas 1994). In the PSI-less control strain, fluorescence spectra were similar, independent of growth in the presence or absence of NaCl (Fig. 3a). However, when Chl (435 nm) was excited in the PSI-less/ScpABCDE− strain grown in the presence of 0.2 M NaCl, the fluorescence maximum at 695 nm increased compared to the control cells grown in the absence of NaCl (Fig. 3b).
Fig. 3.
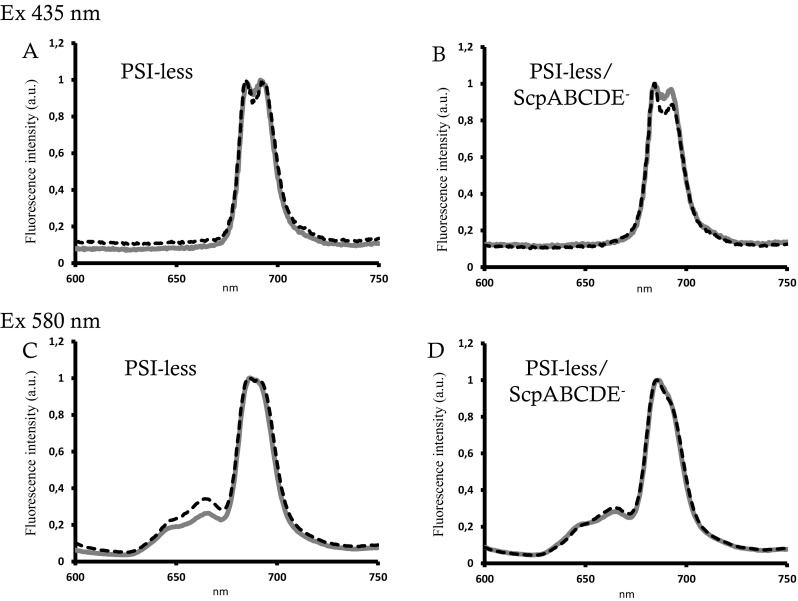
77 K fluorescence spectra of the PSI-less (left) and the PSI-less/ScpABCDE− mutant (right). Spectra were recorded from 600 to 750 nm after growth in the absence (black line) or presence (gray line) of 0.2 M NaCl and normalized to the 685 nm peak. a Excitation of chlorophyll at 435 nm. b Excitation of phycobilisomes at 580 nm
Excitation of the phycobilisomes at 580 nm showed a decrease of phycobilisome fluorescence in the PSI-less control after growth in the presence of salt indicating an improved energy transfer between phycobilisomes and PSII or increased OCP-mediated NPQ (Fig. 3c). On the contrary, no difference in fluorescence upon growth in the presence of NaCl was observed in the PSI-less/ScpABCDE− strain (Fig. 3d). The higher 695 nm emission after 435 nm excitation observed in the PSI-less/ScpABCDE− strain upon growth in the presence of salt could therefore originate from a disturbed energy transfer between CP43 and CP47 and/or from a structural rearrangement around the PSII antenna CP47 (D’Haene et al. 2015).
Enhanced cell pigmentation is independent of pH/osmolarity
To understand the effect of NaCl on the phenotype of the PSI-less/ScpABCDE− mutant and to distinguish the salt effect from a possible pH/osmolarity effect, the mutant strain and its PSI-less control were grown in BG-11 medium buffered to pH 8 (standard), pH 6.5, or pH 9, or NaHCO3 was added at a final concentration of 0.5 mM (Supplementary Fig. 1 and Supplementary Table 1). While at pH 9 the amount of Chl a per cell increased in both strains (accompanied with an increased doubling time) (Supplementary Table 1), at pH 6.5 no significant differences to the standard pH of 8 were observed in the PSI-less strain. Notably, at pH 6.5 the PSI-less/ScpABCDE− strain was not viable, and it died within 4 days after inoculation. The growth rate in the presence of 0.5 mM HCO3 − was diminished in both strains (by a factor of 1.5 for the PSI-less and 2 for the PSI-less/ScpABCDE− strain) compared to standard conditions (p < 0.001, Supplementary Fig. 1, Supplementary Table 1), and the amount of chlorophyll per cell was slightly, but not significantly, lower in both strains when grown in the presence of HCO3 − compared to growth at pH 8 (p > 0.05, Supplementary Table 1). HCO3 − is transported into the cyanobacterial cell via a symporter system powered by Na+; increased HCO3 − concentration in the medium could therefore induce effects similar to increased NaCl concentration. Based on these data, we conclude that NaCl directly affects the cell growth, and a secondary effect based on pH osmolarity can be excluded.
NaCl increases the respiration rate in the PSI-less/ScpABCDE−mutant
Growth in the presence of NaCl has been shown to increase the respiratory activity in Synechocystis wild-type cells (Jeanjean et al. 1990, 1993). To analyze if a similar effect could be observed in the two PSI-less mutants used in this study, the respiration rates of the PSI-less control and the PSI-less/ScpABCDE− strain were measured in the presence or absence of NaCl (Table 1). In standard BG-11 medium, the respiration rate of the PSI-less/ScpABCDE− mutant was twice as high as in the PSI-less control. Growth in the presence of 0.2 M NaCl led to an additional doubling of its respiration rate (p < 0.001, Table 1), while respiration of the PSI-less control only slightly increased. NaCl therefore strongly affects the respiration rate of the PSI-less/ScpABCDE− mutant and concomitantly induces a higher flux of electrons through the respiration chain.
NaCl prevents the production of singlet oxygen in the PSI-less/ScpABCDE−mutant
1O2 is highly damaging in cells, and the SCPs have been suggested to prevent ROS damage (Sinha et al. 2012). Based on our result that the biosynthesis of chlorophyll and the assembly of PSII complexes are enhanced in the presence of NaCl in the PSI-less/ScpABCDE− strain, the effect of NaCl on 1O2 was measured in intact mutant cells using chemical trapping of histidine (Rehman et al. 2013). As expected, an increased rate of 1O2 production was observed in the PSI-less/ScpABCDE− mutant, 1.4 times higher than in the PSI-less strain, as calculated from the rate of O2 uptake due to oxidation of histidine by 1O2 (p < 0.001, Fig. 4). In the presence of 0.2 M NaCl, the production rate of 1O2 slightly (non-significantly) decreased in the PSI-less control (p > 0.05, Fig. 4). However, upon growth in the presence of 0.2 M NaCl the rate of 1O2 production of the PSI-less/ScpABCDE− strain was decreased by 2.6 times relative to the value obtained in standard BG-11 (p < 0.001, Fig. 4). In fact, in the presence of NaCl the 1O2 production rate of the PSI-less/ScpABCDE− mutant was only half of the one in the PSI-less control.
Fig. 4.

Measurement of 1O2 production expressed as the inverse rate of His-mediated oxygen uptake in the PSI-less and the PSI-less/ScpABCDE− strains grown in the presence or absence of NaCl. The measurements were performed in the presence of 5 mM His at 2300 μmol photon m−2 s−1 light intensity. The results are a mean (± SD) of three independent experiments. P values are classified as: *0.01 to 0.05 significant; **0.001 to 0.01 very significant; ***< 0.001 extremely significant
NaCl leads to decreased carbon/nitrogen (C/N) ratio in the PSI-less/ScpABCDE−strain
Attenuated total reflectance Fourier Transform Infrared (ATR FT-IR) spectroscopy is a rapid and inexpensive method to ensure the cell fitness by measuring the C/N ratio. Using this method, we had previously observed a strong increase of the C/N ratio in the PSI-less/ScpABCDE− strain compared to the PSI-less control, being a symptom of oxidative damage and ultimately reduced fitness of the cells (Tibiletti et al. 2016). These findings could be confirmed by measuring ATR FT-IR on both mutants upon growth in BG-11 medium supplemented with 10 mM glucose (Table 2). However, after growth in BG-11 supplemented with glucose and 0.2 M NaCl, the C/N ratio in the PSI-less/ScpABCDE− strain significantly decreased (p < 0.001) by more than half, becoming comparable to the PSI-less strain cultivated in standard conditions (Table 2), while NaCl in the PSI-less control strain only induced a decrease of 30%. Growth in the presence of NaCl therefore seems to influence not only the electron transport chain, but also the entire cell metabolism.
Table 2.
C/N ratios of the PSI-less control and the PSI-less/ScpABCDE− mutant in the presence or absence of NaCl calculated after ATR FT-IR
| C/N | |
|---|---|
| PSI-less | 0.63 (± 0.12) |
| PSI-less+NaCl | 0.44* (± 0.1) |
| PSI-less/ScpABCDE− | 1.38*** (± 0.06) |
| PSI-less/ScpABCDE−+NaCl | 0.75*** (± 0.05) |
Standard deviations with n ≥ 3 are given in brackets
P values are classified as: *0.01 to 0.05 significant; **0.001 to 0.01 very significant; ***< 0.001 extremely significant
PSII photoinhibition
We monitored PSII photoinhibition in the PSI-less and PSI-less/ScpABCDE− strains grown in standard BG-11 or in BG-11 supplemented with NaCl (Fig. 5). In the PSI-less control strain, PSII activity decreased to 70% after high light stress (90 min at 200 μmol photons m−2 s−1) and recovered to about 85% after 90 min in weak light, independent of the presence or absence of NaCl (Fig. 5a, b). Addition of gabaculine, an inhibitor of chlorophyll biosynthesis (Hill et al. 1985), and phycocyanin (dotted lines) led to further decrease of PSII activity during light stress (increased photoinhibition) and diminished recovery in weak light (to less than 80%; Fig. 5a, b, dotted lines). We conclude that the enhanced loss of PSII in the presence of gabaculine is related to a partial impairment of PSII repair, since the elimination of protein synthesis by the inhibitor lincomycin led to the same PSII activity loss (rate and extent) independent of the presence or absence of gabaculine (Fig. 5a, b, down triangle symbol, solid line and circle symbol, dotted lines).
Fig. 5.
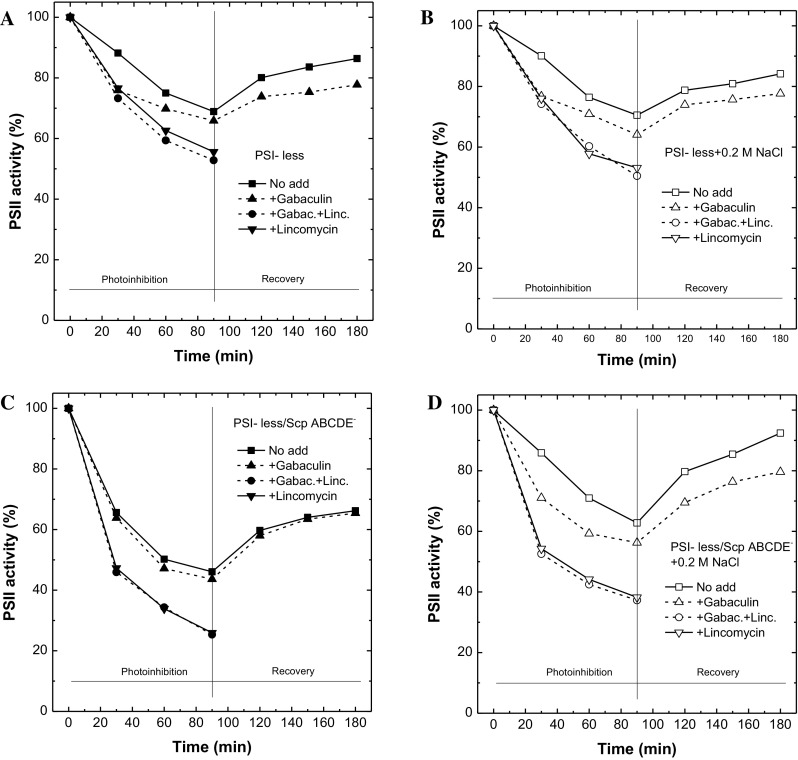
Measurements of photodamage and PSII repair of the PSI-less (upper panels) and PSI-less/ScpABCDE− (lower panels) mutants grown in the presence (right, open symbols) or absence (left, closed symbols) of 0.2 M NaCl. Photoinhibitory treatment was performed in the presence (down triangle symbol, solid line and circle symbol, dotted lines) or absence (square symbol, solid line and upper triangle symbol, dotted lines) of 300 μg/mL lincomycin, as well as in the presence (dotted lines) or absence (solid lines) of 5 μM gabaculine
The PSI-less/ScpABCDE− mutant showed stronger photoinhibition compared to the PSI-less strain when grown in standard BG-11 supplemented with glucose (Fig. 5c), as observed earlier (Hernandez-Prieto et al. 2011). More than 50% of the PSII activity was lost, and weak light allowed recovery to 60% activity. In the presence of 0.2 M NaCl, however, only 30–40% of PSII became photoinhibited in the PSI-less/ScpABCDE− mutant, and recovery was up to 90% (Fig. 5d). Interestingly, in the PSI-less/ScpABCDE− strain grown in the absence of NaCl, addition of gabaculine had no effect, neither on its photoinhibition, nor on recovery (Fig. 5c, dotted line) (see also Hernandez-Prieto et al. 2011), while after growth in the presence of 0.2 M NaCl gabaculine induced a stronger photoinhibition (40% loss of PSII activity) and prevented full recovery (only up to around 80%). Even in this mutant addition of lincomycin induced the same rates independent of the presence or absence of gabaculine (Fig. 5c, d, open down triangle symbol, solid line and open circle symbol, dotted lines).
Discussion
During salt stress, it is important for cyanobacteria like Synechocystis sp. PCC 6803 (Jeanjean et al. 1990, 1993; Peschek et al. 1994) or Synechococcus sp. PCC 6311 (Fry et al. 1986) to provide a proton gradient for Na+/H+ antiporters localized in the thylakoid or cytoplasmic membrane (Hagemann 2011). The ATP to drive these Na+/H+ antiporters is provided by oxidase activity and the respiratory electron transport (Hagemann 2011). Enhanced respiration in the presence of NaCl is also known as salt respiration (Paschinger 1977; Pils and Schmetterer 2001). In cyanobacteria, the respiratory electron transport shares common components with the photosynthetic electron transport (Scherer 1990; Schultze et al. 2009; Ermakova et al. 2016), and therefore respiration can function as a sink for PSII-generated electrons when needed (e.g., during excess of light). The efficiency of oxidases to function as electron sinks in Synechocystis (Vermaas et al. 1994; Berry et al. 2002; Howitt and Vermaas 1998; Nomura et al. 2006; Ermakova et al. 2016; Lea-Smith et al. 2013) and other photosynthetic organisms (Houille-Vernes et al. 2011; Laureau et al. 2013) has been shown earlier.
In Synechocystis, most Chl is bound to PSI, and mutants deficient of PSI contain only 20% of the wild-type chlorophyll amount (Shen et al. 1993). In PSI-less mutants, PSII is very vulnerable to photooxidation as PSII-generated electrons are not efficiently consumed by PSI (Sandström et al. 2002). As a consequence, PSI-less mutants are not able to grow at normal or high irradiances and express the stress-induced SCPs constitutively. Additional deletion of the five SCPs leads to further reduction in chlorophyll (Xu et al. 2004; Tibiletti et al. 2016). In this work, we show that the presence of 0.2 M NaCl restores the bleached phenotype of the PSI-less/ScpABCDE− mutant strain. We propose that the presence of 0.2 M NaCl induces increased respiratory activity (Table 1) that consumes the excess of PSII-generated electrons, thereby reducing the pressure of the electron transport chain, lowering incidences of charge recombination, and preventing 1O2 production (Fig. 4). Lower ROS in turn reduces the risk for PSII photoinhibition (Fig. 5) and allows the accumulation of Chl a assembled into PSII (Table 1).
In the PSI-less/ScpABCDE− mutant, the rate of oxygen evolution is higher than the respiration rate (Table 1); besides oxidases, other cell components have therefore to be involved in the consumption of PSII-generated electrons. Alternative electron sinks, such as the Flv2/Flv4 complex, which accepts electrons directly from PSII (Zhang et al. 2012), the hydrogenase complex, nitrate reductase, or carbon fixation might fulfill this function (Gutthann et al. 2007). The lower C/N ratio observed in PSI-less/ScpABCDE− cells exposed to NaCl indicates the nitrate reductase and CO2 (possibly through a PQH2-oxidizing pathway, Wang et al. 2012) to serve as terminal acceptors. As respiration and photosynthesis are intertwined in cyanobacteria, we cannot exclude in PSI-less mutants the O2 uptake to vary during the light phase. In the absence of PSI, O2 uptake in light might be stimulated; in the cyanobacterium Synechococcus elongatus, growth in high light stress induced O2 uptake (Hoch et al. 1963).
Na+/H+ antiporters not only extrude Na+, but also are important to maintain the intracellular pH by transporting H+. However, recovery of the bleached phenotype in the PSI-less/ScpABCDE− was pH independent (Supplementary Fig. 1 and Supplementary Table 1). Presence of HCO3 −, which is mainly transported into the cyanobacterial cell via a symporter system powered by Na+ (Shibata et al. 2002), did not recover the phenotype either. These data suggest Na+ (or Cl−) ions and not H+ to be mainly responsible for the recovery of the PSI-less/ScpABCDE− mutant. Interestingly, the PSI-less/ScpABCDE− mutant died when grown at low pH. Already in our previous microarray data comparing the PSI-less/ScpABCDE− to PSI-less control cells (Tibiletti et al. 2016), we observed a differential expression of several genes involved in ion homeostasis and inorganic carbon metabolism, indicating a low capacity to regulate pH/osmolarity of the mutant. For instance, aqpZ encoding an aquaporin, sbta encoding the Na+-dependent HCO3 − transporter, nhaS3 encoding the only thylakoid localized Na+/H+ antiporter, and pxcA encoding an ATP-dependent proton extrusion system were found to be up-regulated in the PSI-less/ScpABCDE− mutant compared to the PSI-less control. High H+ influx into the cell at low pH might block or saturate the light-induced proton extrusion system that is essential for growth, CO2 transport, and nitrate uptake at pH 6.5 (Sonoda et al. 1998; Katoh et al. 1996). In mixotrophic growth, inorganic carbon (Ci) transportation through the carbon concentration mechanisms (CCM) could be important to regulate the internal pH, pumping H+ out of the cell. This mechanism is most important at low pH when CO2 is the most abundant Ci species dissolved in water (Kirk 2011). The high C/N ratio observed in the PSI-less/ScpABCDE− could block the full induction of CCM, leading to accumulation of H+ in the cell and finally to death. Unfortunately, we failed to measure the proton extrusion induced by NaCl and the pH difference across the membranes using the dye acridine yellow as described by Teuber and coworkers (Teuber et al. 2001).
At 77 K, the ratio of fluorescence at 695 and 685 nm (F695/F685) in the PSI-less/ScpABCDE− mutant upon growth in the presence of NaCl was restored to approximately 1, similar to the PSI-less mutant, while during growth under standard conditions the 695 nm fluorescence peak was enlarged compared to the one at 685 nm (Fig. 3a, b). This might indicate a disturbed energy transfer between CP43 and CP47 and/or a structural rearrangement around the PSII antenna CP47 in PSI-less/ScpABCDE−. We propose that the lack of SCPs affects the structural organization of PSII in stressed conditions, possibly under the environment of the red-emitting Chl 627 of CP47 as suggested for a mutant deficient in PsbH (D’Haene et al. 2015). Association of SCPs to CP47 seems to be PsbH dependent and in the absence of PsbH and SCPs the environment of the Chl 627 of CP47 is modified. Thus, SCPs might effectively stabilize PSII complexes under stress conditions.
With the reduction of 1O2, a strong recovery from photoinhibition was observed in the PSI-less/ScpABCDE− upon growth in NaCl (Fig. 5). The percentage of PSII recovering after photoinhibition in the PSI-less/ScpABCDE− mutant upon growth with NaCl was similar to the PSI-less strain, both in the presence and absence of gabaculine. Gabaculine is an inhibitor of Chl biosynthesis that affects the synthesis of the 5-aminolevulinic acid and inhibits phycocyanin (Hill et al. 1985; Beale 1999). Hence, the cells grown in the presence of gabaculine can only repair or assemble new PSII with Chl molecules recycled from damaged PSII. Comparing the relative loss of PSII activity in the presence or absence of gabaculine, the shortage of Chl molecules affects strongly the photoinhibitory period (mostly during the first 20 min) than the recovery. Addition of gabaculine had no additional effect on photoinhibition and recovery indicating that the novo assembly and repair of PSII in the PSI-less/ScpABCDE− mutant grown in the absence of NaCl relies only on Chl recycling. Based on the data presented here and in Hernandez-Prieto et al. (2011), we calculated 5–15% of PSII repair to depend on newly synthesized Chl molecules during photoinhibition and 5% during recovery in PSI-less and PSI-less/ScpABCDE− upon growth with NaCl. This calculation was performed considering the PSII activity in the presence or absence of gabaculine at 90 and 180 min.
In the PSI-less/ScpABCDE− mutant, the photon flux density exceeds the photosynthetic capacity even at low light. In the absence of the SCPs, chlorophyll-binding proteins are destabilized (Xu et al. 2002, 2004; Hernandez-Prieto et al. 2011) leading to photoinhibition of PSII and 1O2 production (Fig. 6a). PSII-generated 1O2 induces the loss of PSII activity, bleaching, and decreased growth and fitness observed in the PSI-less/ScpABCDE− strain (Tibiletti et al. 2016). In the presence of NaCl, however, the electrons generated by PSII in the PSI-less/ScpABCDE− mutant are efficiently absorbed and consumed by Cox and Cyd, as well as by other electron acceptors (Fig. 6b). In the PSI-less/ScpABCDE− mutant, PSII electron sinks generate a shield against 1O2 and avoid cellular damages. A decreased amount of 1O2 in the presence of NaCl seems to enhance cell energization, the C catabolism as well as the N uptake, increased protein synthesis, and Chl biosynthesis leading to higher PSII assembly.
Fig. 6.
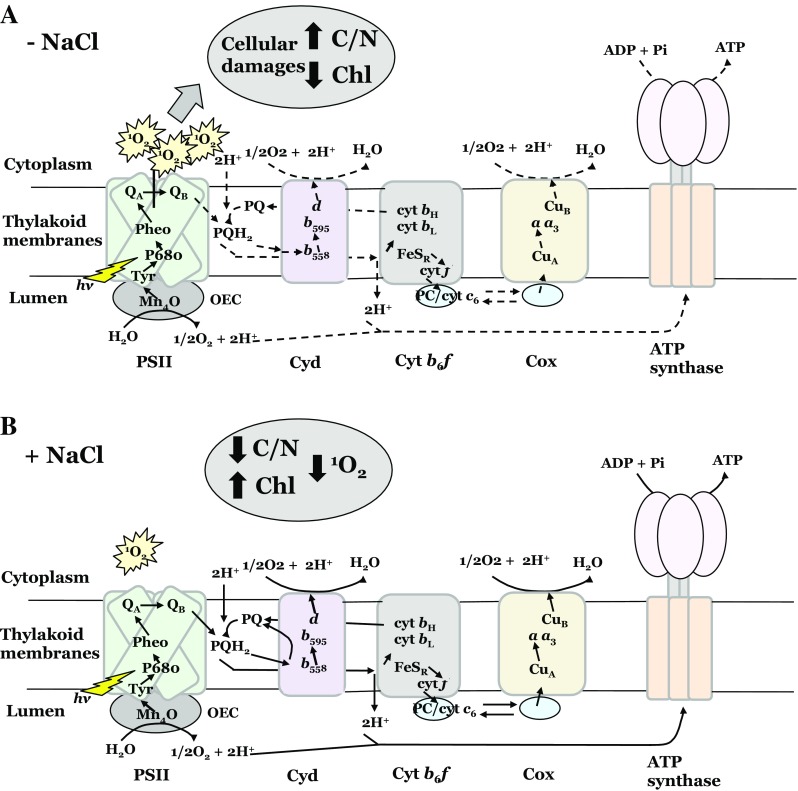
Schematic diagram of the thylakoid membrane-localized photosynthetic and respiratory electron transport chains in PSI-less/ScpABCDE− in the absence (a) or presence (b) of NaCl. Lines indicate the electron transport, filled lines indicate sustained electron transfer, and dotted lines indicate poor electron transfer. Cox, cytochrome c oxidase; Cyd, bd quinol oxidases; Cyt c 6, cytochrome c 6; PC, plastocyanin; PQ, plastoquinone; PQH2, plastoquinol; OEC, oxygen evolving complex; PSII, photosystem II
Conclusions
SCPs, by stabilizing chlorophyll-binding proteins and PSII assembly, protect PSII from photoinhibitory damages. In the absence of SCPs, electrons accumulate and will lead to ROS formation. Here we show that the presence of 0.2 M NaCl induces increased respiratory activity (salt respiration) in the PSI-less/ScpABCDE− that consumes the excess of PSII-generated electrons, thereby decreasing the electron flow through the electron transport chain, reducing the occurrence of charge recombination, and preventing 1O2 production.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Fig. 1 Color appearance of the PSI-less and PSI-less/ScpABCDE− mutants grown in BG-11 supplemented with 10 mM glucose at either pH 8 (standard conditions), pH 6.5 or pH 9 (upper panel) and at pH 8 in the absence or presence of 0.5 mM NaHCO3 (lower panel) (PDF 134 KB)
Acknowledgements
The work was supported by the Swedish Energy Agency (2012-005889 to C.F.) and Umeå University (to C.F.). I.V. and A.R. were supported by National Research, Development and Innovation office of Hungary (Grant No. K-116016). The authors thank Prof. Rainer Hienerwadel from Aix-Marseille University, France, for his help with the FT-IR measurements.
Footnotes
Electronic supplementary material
The online version of this article (doi:10.1007/s11120-017-0426-3) contains supplementary material, which is available to authorized users.
References
- Aro EM, Virgin I, Andersson B. Photoinhibition of photosystem II. Inactivation, protein damage and turnover. Biochim Biophys Acta. 1993;1143:113–134. doi: 10.1016/0005-2728(93)90134-2. [DOI] [PubMed] [Google Scholar]
- Beale SI. Enzymes of chlorophyll biosynthesis. Photosynth Res. 1999;60:43–73. doi: 10.1023/A:1006297731456. [DOI] [Google Scholar]
- Berry S, Schneider D, Vermaas WFJ, Rögner M. Electron transport routes in whole cells of Synechocystis sp. strain PCC 6803: the role of the cytochrome bd-type oxidase. BioChemistry. 2002;41:3422–3429. doi: 10.1021/bi011683d. [DOI] [PubMed] [Google Scholar]
- Chaux F, Peltier G, Johnson X. A security network in PSI photoprotection: regulation of photosynthetic control, NPQ and O2 photoreduction by cyclic electron flow. Front Plant Sci. 2015;6:875. doi: 10.3389/fpls.2015.00875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cooley JW, Vermaas WFJ. Succinate dehydrogenase and other respiratory pathways in thylakoid membranes of Synechocystis sp. Strain PCC 6803: capacity comparisons and physiological function. J Bacteriol. 2001;183:4251–4258. doi: 10.1128/JB.183.14.4251-4258.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- D’Haene SE, Sobotka R, Bučinská L, Dekker JP, Komenda J. Interaction of the PsbH subunit with a chlorophyll bound to histidine 114 of CP47 is responsible of the red 77 K fluorescence of Photosystem II. Biochim Biophys Acta. 2015;1847:1327–1334. doi: 10.1016/j.bbabio.2015.07.003. [DOI] [PubMed] [Google Scholar]
- Dolganov NA, Bhaya D, Grossman AR. Cyanobacterial protein with similarity to the chlorophyll a/b binding proteins of higher plants: evolution and regulation. Proc Natl Acad Sci USA. 1995;92:636–640. doi: 10.1073/pnas.92.2.636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ermakova M, Huokko T, Richaud P, Bersanini L, Howe CJ, Lea-Smith DJ, Peltier G, Allahverdiyeva Y. Distinguishing the role of thylakoid respiratory terminal oxidases in the cyanobacterium Synechocystis sp. PCC 6803. Plant Physiol. 2016;171:1307–1319. doi: 10.1104/pp.16.00479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fry IV, Huflejt M, Erber WW, Peschek GA, Packer L. The role of respiration during adaptation of the freshwater cyanobacterium Synechococcus 6311 to salinity. Arch Biochem Biophys. 1986;244:686–691. doi: 10.1016/0003-9861(86)90637-5. [DOI] [PubMed] [Google Scholar]
- Funk C, Vermaas WFJ. A cyanobacterial gene family coding for single-helix proteins resembling part of the light-harvesting proteins from higher plants. BioChemistry. 1999;38:9397–9404. doi: 10.1021/bi990545+. [DOI] [PubMed] [Google Scholar]
- Gutthann F, EgertM, Marques A, Appel J. Inhibition of respiration and nitrate assimilation enhances photohydrogen evolution under low oxygen concentrations in Synechocystis sp. PCC 6803. Biochim Biophys Acta. 2007;1767:161–169. doi: 10.1016/j.bbabio.2006.12.003. [DOI] [PubMed] [Google Scholar]
- Hagemann M. Molecular biology of cyanobacterial salt acclimation. FEMS Microbiol Rev. 2011;35:87–123. doi: 10.1111/j.1574-6976.2010.00234.x. [DOI] [PubMed] [Google Scholar]
- Havaux M, Guedeney G, He Q, Grossman AR. Elimination of high-light inducible polypeptides related to eukaryotic chlorophyll a/b-binding proteins results in aberrant photoacclimation in Synechocystis PCC6803. Biochim Biophys Acta. 2003;1557:21–33. doi: 10.1016/S0005-2728(02)00391-2. [DOI] [PubMed] [Google Scholar]
- He Q, Dolganov N, Bjorkman O, Grossman AR. The high light-inducible polypeptides in Synechocystis PCC6803. Expression and function in high light. J Biol Chem. 2001;276:306–314. doi: 10.1074/jbc.M008686200. [DOI] [PubMed] [Google Scholar]
- Hernández-Prieto MA, Tibiletti T, Abasova L, Kirilovsky D, Vass I, Funk C. The small CAB-like proteins of the cyanobacterium Synechocystis sp. PCC 6803: their involvement in chlorophyll biogenesis for Photosystem II. Biochim Biophys Acta. 2011;1807:1043–1151. doi: 10.1016/j.bbabio.2011.05.002. [DOI] [PubMed] [Google Scholar]
- Hill CM, Pearson SA, Smith AJ, Rogers LJ. Inhibition of chlorophyll synthesis in Hordeum vulgare by 3-amino 2,3-dihydrobenzoic acid (gabaculin) Biosci Rep. 1985;5:775–781. doi: 10.1007/BF01119876. [DOI] [PubMed] [Google Scholar]
- Hoch G, Owens OV, Kok B. Photosynthesis and respiration. Arch Biochem Byophys. 1963;101:171–180. doi: 10.1016/0003-9861(63)90547-2. [DOI] [PubMed] [Google Scholar]
- Houille-Vernes L, Rappaport F, Wollman FA, Alric J, Johnson X. Plastid terminal oxidase 2 (PTOX2) is the major oxidase involved in chlororespiration in Chlamydomonas. Proc Natl Acad Sci USA. 2011;108:20820–20825. doi: 10.1073/pnas.1110518109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howitt CA, Vermaas WFJ. Quinol and cytochrome oxidases in the cyanobacterium Synechocystis sp. PCC 6803. BioChemistry. 1998;37:17944–17951. doi: 10.1021/bi981486n. [DOI] [PubMed] [Google Scholar]
- Howitt CA, Cooley JW, Wiskich JT, Vermaas WFJ. A strain of Synechocystis sp. PCC 6803 without photosynthetic oxygen evolution and respiratory oxygen consumption: implications for the study of cyclic photosynthetic electron transport. Planta. 2001;214:46–56. doi: 10.1007/s004250100578. [DOI] [PubMed] [Google Scholar]
- Jeanjean R, Onana B, Peschek GA, Joset F. Mutants of the cyanobacterium Synechocystis PCC6803 impaired in respiration and unable to tolerate high salt concentrations. FEMS Microbiol Lett. 1990;68:125–129. doi: 10.1111/j.1574-6968.1990.tb04135.x. [DOI] [Google Scholar]
- Jeanjean R, Matthijs HCP, Onana B, Havaux M, Joset F. Exposure of the cyanobacterium Synechocystis PCC6803 to salt stress induces concerted changes in respiration and photosynthesis. Plant Cell Physiol. 1993;34:1073–1079. [Google Scholar]
- Katoh A, Sonoda M, Katoh H, Ogawa T. Absence of light-induced extrusion in a cotA-less mutant of Synechocystis sp. strain PCC6803. J Bacteriol. 1996;178:5452–5455. doi: 10.1128/jb.178.18.5452-5455.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirilovsky D, Kerfeld CA. The orange carotenoid protein in photoprotection of photosystem II in cyanobacteria. Biochim Biophys Acta. 2012;1817:158–166. doi: 10.1016/j.bbabio.2011.04.013. [DOI] [PubMed] [Google Scholar]
- Kirk JTO. Light and photosynthesis in aquatic ecosystem. New York: Cambridge University Press; 2011. [Google Scholar]
- Knoppová J, Sobotka R, Tichy M, Yu J, Konik P, Halada P, Nixon PJ, Komenda J. Discovery of a chlorophyll binding protein complex involved in the early steps of photosystem II assembly in Synechocystis. Plant Cell. 2014;26:1200–1212. doi: 10.1105/tpc.114.123919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kufryk G, Hernandez-Prieto MA, Kieselbach T, Miranda H, VermaasWFJ, Funk C. Association of small CAB-like proteins (SCPs) of Synechocystis sp. PCC 6803 with Photosystem II. Photosynth Res. 2008;95:135–145. doi: 10.1007/s11120-007-9244-3. [DOI] [PubMed] [Google Scholar]
- Laureau C, De Paepe R, Latouche G, Moreno-Chacón M, Finazzi G, Kuntz M, Cornic G, Streb P. Plastid terminal oxidase (PTOX) has the potential to act as a safety valve for excess excitation energy in the alpine plant species Ranunculus glacialis L. Plant Cell Environ. 2013;36:1296–1310. doi: 10.1111/pce.12059. [DOI] [PubMed] [Google Scholar]
- Lea-Smith DJ, Ross N, Zori M, Bendall DS, Dennis JS, Scott SA, Smith AG, Howe CJ. Thylakoid terminal oxidases are essential for the cyanobacterium Synechocystis sp. PCC 6803 to survive rapidly changing light intensities. Plant Physiol. 2013;162:484–495. doi: 10.1104/pp.112.210260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDonald AE, Ivanov AG, Bode R, Maxwell DP, Rodermel SR, Hüner NPA. Flexibility in photosynthetic electron transport: the physiological role of plastoquinol terminal oxidase (PTOX) Biochim Biophys Acta. 2011;1807:954–967. doi: 10.1016/j.bbabio.2010.10.024. [DOI] [PubMed] [Google Scholar]
- Niyogi KK. Photoprotection revisited: genetic and molecular approaches. Annu Rev Plant Physiol Plant Mol Biol. 1999;50:333–359. doi: 10.1146/annurev.arplant.50.1.333. [DOI] [PubMed] [Google Scholar]
- Nomura CT, Persson S, Shen G, Inoue-Sakamoto K, Bryant DA. Characterization of two cytochrome oxidase operons in the marine cyanobacterium Synechococcus sp. PCC 7002: inactivation of ctaDI affects the PS I:PS II ratio. Photosynth Res. 2006;87:215–228. doi: 10.1007/s11120-005-8533-y. [DOI] [PubMed] [Google Scholar]
- Paschinger H. DCCD induced sodium uptake by Anacystis nidulans. Arch Microbiol. 1977;113:285–291. doi: 10.1007/BF00492037. [DOI] [PubMed] [Google Scholar]
- Peltier G, Tolleter D, Billon E, Cournac L. Auxiliary electron transport pathways in chloroplast of microalgae. Photosynth Res. 2010;106:19–31. doi: 10.1007/s11120-010-9575-3. [DOI] [PubMed] [Google Scholar]
- Peschek GA, Obinger C, Fromwald S, Bergman B. Correlation between immunogold labels and activities of the cytochrome c oxidase (Aa (3) type) in membranes of salt-stressed cyanobacteria. FEMS Microbiol Lett. 1994;124:431–437. doi: 10.1111/j.1574-6968.1994.tb07320.x. [DOI] [Google Scholar]
- Pils D, Schmetterer G. Characterization of three bioenergetically active respiratory terminal oxidases in the cyanobacterium Synechocystis sp. Strain PCC6803. FEBS Microbiol Lett. 2001;203:217–222. doi: 10.1111/j.1574-6968.2001.tb10844.x. [DOI] [PubMed] [Google Scholar]
- Porra RJ. The chequered history of the development and use of simultaneous equations for the accurate determination of chlorophylls a and b. Photosynth Res. 2002;73:149–156. doi: 10.1023/A:1020470224740. [DOI] [PubMed] [Google Scholar]
- Promnares K, Komenda J, Bumba L, Nebesarova J, Vacha F, Tichy M. Cyanobacterial small chlorophyll-binding protein ScpD (HliB) is located on the periphery of photosystem II in the vicinity of PsbH and CP47 subunits. J Biol Chem. 2006;281:32705–32713. doi: 10.1074/jbc.M606360200. [DOI] [PubMed] [Google Scholar]
- Rehman AU, Cser K, Sass L, Vass I. Characterization of singlet oxygen production and its involvement in photodamage of Photosystem II in the cyanobacterium Synechocystis PCC 6803 by histidine-mediated chemical trapping. Biochim Biophys Acta. 2013;1827:689–698. doi: 10.1016/j.bbabio.2013.02.016. [DOI] [PubMed] [Google Scholar]
- Rippka R, Deruelles J, Waterbury JB, Herdman M, Stanier RY. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. J Gen Microbiol. 1979;111:1–61. [Google Scholar]
- Roach T, Krieger-Liszkay A. Regulation of photosynthetic electron transport and photoinhibition. Curr Prot Pept Sci. 2014;15:351–362. doi: 10.2174/1389203715666140327105143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sandström S, Ivanov AG, Park YI, Öquist G, Gustafsson P. Iron stress responses in the cyanobacterium Synechococcus sp. PCC7942. Physiol Plant. 2002;116:255–263. doi: 10.1034/j.1399-3054.2002.1160216.x. [DOI] [PubMed] [Google Scholar]
- Scherer S. Do photosynthetic and respiratory electron transport chains share redox proteins? Trends Biochem Sci. 1990;15:458–462. doi: 10.1016/0968-0004(90)90296-N. [DOI] [PubMed] [Google Scholar]
- Schultze M, Forberich B, Rexroth S, Dyczmon NG, Roegner M, Appel J. Localization of cytochrome b6f complexes implies an incomplete respiratory chain in cytoplasmic membranes of the cyanobacterium Synechocystis sp. PCC 6803. Biochim Biophys Acta. 2009;1787:1479–1485. doi: 10.1016/j.bbabio.2009.06.010. [DOI] [PubMed] [Google Scholar]
- Shen G, Vermaas WF. Chlorophyll in a Synechocystis sp. PCC 6803 without photosystem I and photosystem II core complexes. Evidences for peripheral antenna chlorophyll in cyanobacteria. J Biol Chem. 1994;269:13904–13910. [PubMed] [Google Scholar]
- Shen G, Boussiba S, Vermaas WFJ. Synechocystis sp PCC 6803 strains lacking photosystem I and phycobilisome function. Plant Cell. 1993;5:853–1863. doi: 10.1105/tpc.5.12.1853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shi LX, Hall M, Funk C, Schöder WP. Photosystem II a growing complex: updates on newly discovered components and low molecular mass proteins. Biochim Biophys Acta. 2012;1817:13–25. doi: 10.1016/j.bbabio.2011.08.008. [DOI] [PubMed] [Google Scholar]
- Shibata M, Katoh H, Sonoda M, Ohkawa H, Shimoyama M, Fukuzawa H, Kaplan A, Ogawa T. Genes essential to sodium-dependent bicarbonate transport in cyanobacteria. Function and phylogenetic analysis. J Biol Chem. 2002;277:18659–18664. doi: 10.1074/jbc.M112468200. [DOI] [PubMed] [Google Scholar]
- Sinha RK, Komenda J, Knoppová J, Sedlářová M, Pospíšil P. Small CAB like proteins prevent formation of singlet oxygen in the damaged photosystem II complex of the cyanobacterium Synechocystis sp. PCC 6803. Plant Cell Environ. 2012;35:806–818. doi: 10.1111/j.1365-3040.2011.02454.x. [DOI] [PubMed] [Google Scholar]
- Sobotka R, McLean S, Zuberova M, Hunter CN, Tichy M. The C-terminal extension of ferrochelatase is critical for enzyme activity and for functioning of the tetrapyrrole pathway in Synechocystis strain PCC 6803. J Bacteriol. 2008;190:2086–2095. doi: 10.1128/JB.01678-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sobotka R, Tichy M, Wilde A, Hunter CN. Functional assignment for the carboxyl-terminal domains of the ferrochelatase from Synechocystis PCC 6803: the CAB domain plays a regulatory role, and region II is essential for catalysis. Plant Physiol. 2011;155:1735–1747. doi: 10.1104/pp.110.167528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sonoda M, Katoh H, Vermass W, Schmetterer G, Ogawa T. Photosynthetic electron transport involved in PxcA-dependent proton extrusions in Synechocystis sp. strain PCC6803: effect of pxcA inactivation on CO2, HCO3–, and NO3– uptake. J Bacteriol. 1998;180:3799–3803. doi: 10.1128/jb.180.15.3799-3803.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Storm P, Tibiletti T, Hall M, Funk C. Refolding and enzyme kinetic studies on the ferrochelatase of the cyanobacterium Synechocystis sp. PCC 6803. PLoS One. 2013;8:e55569. doi: 10.1371/journal.pone.0055569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Teuber M, Rögner M, Berry S. Fluorescent probes for non-invasive bioenergetic studies of whole cyanobacterial cell. Biochim Biophys Acta. 2001;1506:31–46. doi: 10.1016/S0005-2728(01)00178-5. [DOI] [PubMed] [Google Scholar]
- Tibiletti T, Hernández-Prieto MA, Matthijs HC, Niyogi KK, Funk C. Deletion of the gene family of small chlorophyll-binding protein (ScpABCDE) offset C/N homeostasis in Synechocystis PCC 6803. Biochim Biophys Acta. 2016;1857:396–407. doi: 10.1016/j.bbabio.2015.11.011. [DOI] [PubMed] [Google Scholar]
- Vermaas WFJ. Photosynthesis and Respiration in Cyanobacteria: Encyclopedia of Life Sciences. Basingstoke: Macmillan Publishers Ltd.; 2001. [Google Scholar]
- Vermaas WF, Williams JG, Ruthrerford AW, Mathis P, Arntzen CJ. Genetically engineered mutant of the cyanobacterium Synechocystis 6803 lacks photosystem II chlorophyll-binding protein CP-47. Proc Natl Acad Sci USA. 1986;83:9474–9477. doi: 10.1073/pnas.83.24.9474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vermaas W, Shen G, Styring S. Electrons generated by photosystem II are utilized by an oxidase in the absence of photosystem I in the cyanobacterium Synechocystis sp. PCC 6803. FEBS Lett. 1994;337:103–108. doi: 10.1016/0014-5793(94)80638-1. [DOI] [PubMed] [Google Scholar]
- Wang QJ, Singh A, Li H, Nedbal L, Sherman LA, Govindjee, Whitmarsh J. Net light-induced oxygen evolution in photosystem I deletion mutants of the cyanobacterium Synechocystis sp. PCC 6803. Biochim Biophys Acta. 2012;1817:792–801. doi: 10.1016/j.bbabio.2012.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu H, Vavilin D, Funk C, Vermaas WFJ. Small Cab-like proteins regulating tetrapyrrole biosynthesis in the cyanobacterium Synechocystis sp. PCC 6803. Plant Mol Biol. 2002;49:149–160. doi: 10.1023/A:1014900806905. [DOI] [PubMed] [Google Scholar]
- Xu H, Vavilin D, Funk C, Vermaas WFJ. Multiple deletions of small Cab-like proteins in the cyanobacterium Synechocystis sp. PCC 6803: consequences for pigment biosynthesis and accumulation. J Biol Chem. 2004;279:27971–27979. doi: 10.1074/jbc.M403307200. [DOI] [PubMed] [Google Scholar]
- Yao DC, Kieselbach T, Komenda J, Promnares K, Hernandez-Prieto MA, Tichy M, Vermaas WFJ, Funk C. Localization of the small CAB-like proteins in photosystem II. J Biol Chem. 2007;282:267–276. doi: 10.1074/jbc.M605463200. [DOI] [PubMed] [Google Scholar]
- Yao DC, Brune DC, Vavilin D, Vermaas WF. Photosystem II component lifetimes in the cyanobacterium Synechocystis sp. strain PCC 6803: small Cab-like proteins stabilize biosynthesis intermediates and affect early steps in chlorophyll synthesis. J Biol Chem. 2012;287:682–692. doi: 10.1074/jbc.M111.320994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang P, Eisenhut M, Brandt AM, Carmel D, Silén HM, Vass I, Allahverdiyeva Y, Salminen TA, Aro EM. Operon flv4-flv2 provides cyanobacterial photosystem II with flexibility of electron transfer. Plant Cell. 2012;24:1952–1971. doi: 10.1105/tpc.111.094417. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Fig. 1 Color appearance of the PSI-less and PSI-less/ScpABCDE− mutants grown in BG-11 supplemented with 10 mM glucose at either pH 8 (standard conditions), pH 6.5 or pH 9 (upper panel) and at pH 8 in the absence or presence of 0.5 mM NaHCO3 (lower panel) (PDF 134 KB)