

Summary
Theobroma cacao—The Food of the Gods, provides the raw material for the multibillion dollar chocolate industry and is also the main source of income for about 6 million smallholders around the world. Additionally, cocoa beans have a number of other nonfood uses in the pharmaceutical and cosmetic industries. Specifically, the potential health benefits of cocoa have received increasing attention as it is rich in polyphenols, particularly flavonoids. At present, the demand for cocoa and cocoa‐based products in Asia is growing particularly rapidly and chocolate manufacturers are increasing investment in this region. However, in many Asian countries, cocoa production is hampered due to many reasons including technological, political and socio‐economic issues. This review provides an overview of the present status of global cocoa production and recent advances in biotechnological applications for cacao improvement, with special emphasis on genetics/genomics, in vitro embryogenesis and genetic transformation. In addition, in order to obtain an insight into the latest innovations in the commercial sector, a survey was conducted on granted patents relating to T. cacao biotechnology.
Keywords: Theobroma cacao, chocolate, somatic embryogenesis, genetics, genomics, breeding, transformation
Introduction
The diploid tropical fruit crop species (2n = 2x = 20), Theobroma cacao (cacao) (Figure 1), is an economically important agricultural commodity for millions of people worldwide. It is grown by about 6 million farmers globally, and livelihoods of more than 40 million people depend on cocoa (Beg et al., 2017; World Cocoa Foundation, 2012). The majority of world cocoa production (approximately 80%–90%) comes from smallholder farmers (World Cocoa Foundation, 2014). This crop originated from the Amazonian basin (Motamayor et al., 2002; Wood and Lass, 1985), and today, it is cultivated in many regions of the humid tropics.
Figure 1.

Cacao tree with multiple pods.
The cocoa beans are the primary source of raw material for the multibillion dollar industry that produces chocolate and associated confectionery products, with Switzerland being the country with the highest consumption (Figure 2), although much of this is due to purchases by tourists to that country. The economic significance of the chocolate industry has been recently reviewed (Squicciarini and Swinnen, 2016), with the global market for chocolate rising 13% from 2010 to reach US$101 billion in 2015.
Figure 2.

Annual consumption of chocolate per year (modified from Statista.org).
This crop belongs to the Malvaceae family, and more than 20 species are known within the Theobroma genus (Wood and Lass, 1985). Among these, T. cacao is the only species that is cultivated extensively (Wood and Lass, 1985). This species has three genetic groups based on morphological and anatomical characteristics—Criollo (T. cacao Spp. Criollo), Forastero (T. cacao Spp. Sphaerocarpum) and Trinitario (Pridmore et al., 2000). Of these, the Criollo type is well known for its superior flavour and provides the raw material from which fine flavour chocolates are produced; these represent 5%–10% of world chocolate production (Rusconi and Conti, 2010). However, increased susceptibility to pest and diseases, low vigour and yield has made this variety less popular among cacao growers. Today, most of the world's chocolate production (approximately 80%) comes from the Forastero type of cacao; this variety is favoured over the Criollo for its disease‐resistant and high‐yielding nature, and beans from this variety are relatively cheaper than those from the Criollo type (Rusconi and Conti, 2010). The third genetic group, Trinitario, is a hybrid produced from crosses between Criollo and Forastero varieties. This variety was initially developed in Trinidad, and today, it is cultivated in many parts of South and Central America, Africa, South‐East Asia and Oceania for its aroma, productivity and disease‐resistant character.
World cocoa production
The International Cocoa Organization (ICCO) estimated that more than 4.0 million metric tons of cocoa beans were produced worldwide in 2015/16 (Pipitone, 2016). Of this total, it is also estimated that Africa contributed approximately 74% (2.92 million tonnes) in the 2015/16 season. This is 5000 tonnes less than the estimated production for 2014/15. Among the cocoa‐producing regions, Côte d'Ivoire, Ghana and Cameroon contributed 1.57, 0.8 and 0.23 million tonnes, respectively, to global production in 2015/16 (Pipitone, 2016). It is important to note that there is a discrepancy between the cocoa bean production data published by the Food and Agriculture Organization of the United Nations (FAO) and the production estimates by the ICCO. This is mainly due to the use of different sources to estimate the production data. Figures 3 and 4 in the present review are based on the data published by the FAO. Figure 3 shows cocoa bean production (tonnes) from 1993 to 2013 in the leading production regions in Africa, namely Côte d'Ivoire, Ghana, Nigeria and Cameroon. With the exception of Cameroon, a slight decline in production from 2012 to 2013 is noted in the other African countries considered.
Figure 3.
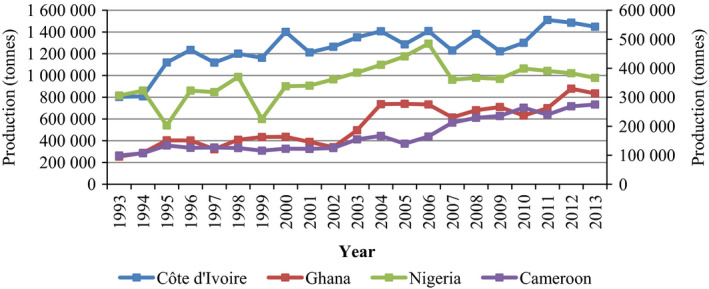
Cocoa bean production in Africa—Côte d'Ivoire, Ghana, Nigeria and Cameroon from 1993 to 2013. Primary axis left: Côte d'Ivoire and Ghana; secondary axis right: Nigeria and Cameroon. Source: FAOSTAT (http://faostat3.fao.org/browse/Q/*/E, retrieved on 7 February 2017).
Figure 4.
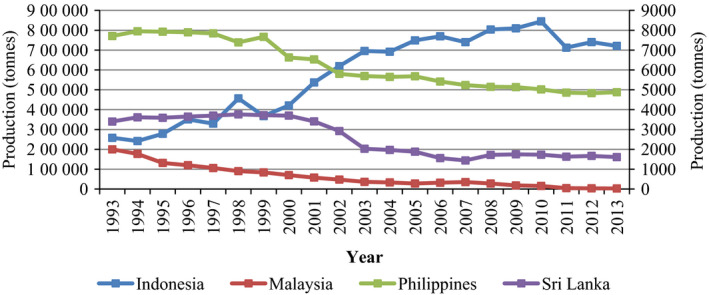
Cocoa bean production in Asia—Indonesia, Malaysia, Philippines and Sri Lanka from 1993 to 2013. Primary axis left: Indonesia; secondary axis right: Malaysia, Philippines and Sri Lanka. Source: FAOSTAT (http://faostat3.fao.org/browse/Q/*/E, retrieved on 7 February 2017).
In addition, the Americas region (16%, 0.64 million tonnes), and Asia and Oceania (10%, 0.4 million tonnes) are ranked as the second and third largest producers of cocoa beans worldwide. At present, Indonesia is the third largest producer after Côte d'Ivoire and Ghana, with an estimated production of 0.33 million tonnes in 2015/16 (Pipitone, 2016). However, production is still relatively low in many Asian countries such as Malaysia, The Philippines and Sri Lanka, which all have a tremendous potential to grow cacao (Figure 4). In addition to the fact that cocoa bean production contributes significantly to the economy of the growing regions, it also serves as a main source of income for millions of smallholder farmers (Darkwah and Verter, 2014).
The demand for cocoa is increasing considerably (approximately 3% per year) (World Cocoa Foundation, 2014). At present, global cocoa production is considered to be at risk, and it has been reported that the world may experience a cocoa shortage by 2020 (Earth Security Group, 2015; Jégourel, 2016). However, the surplus seen in the 2016/17 crop season (up to May 2017) in some cacao growing areas such as Cote d'Ivoire and Ghana is favourable for the future cocoa sector; an 18% increase in the world production is expected for the current crop season as compared to that of the previous season (International Cocoa Organization, 2017).
Uses of cocoa
This perennial shade grown tree crop provides biodiversity benefits. It is cultivated either as monocultures or in association with other crops such as fruit crops (Guiltinan et al., 2008). Cocoa beans are the key raw materials in the production of chocolate and other cocoa‐based products. However, the freshly harvested cocoa beans do not contain the determinants of chocolate aroma or flavour, and hence, postharvesting processing of raw beans (fermentation, drying and roasting) is essential for optimum flavour formation (De Vuyst and Weckx, 2016; Kadow et al., 2015; Kongor et al., 2016; Loureiro et al., 2017). The process of cocoa bean fermentation is trigged by action of micro‐organisms (e.g. yeast, acetic acid bacteria and lactic acid bacteria) (Illeghems et al., 2015; de Melo Pereira et al., 2016), and the flavour precursors such as organic acids, reducing sugars and free amino acids are produced at the end of the process. In addition, the process of fermentation involves significant reduction in polyphenols (epicatechin and catechin) and alkaloids (methylxanthines caffeine, theobromine) found in raw cocoa beans that give rise to bitterness and unpleasant astringency (Kadow et al., 2015; Lee et al., 2016).
Most production of cocoa takes place in the tropics, and the beans produced in this region used to be principally processed elsewhere into cocoa powder and cocoa butter (Wood and Lass, 1985). Now, although most of the cocoa grindings (38%) are carried out in the Europe and Russia region (principally the Netherlands), the remainder is processed close to production areas in the Americas (22%), Asia and Oceania (21%) and Africa (19%) (Pipitone, 2016). In addition to its use in confectionery, cocoa products are also considered to have other functional properties (Konar et al., 2016; Wilson and Hurst, 2015) and are used in a range of pharmaceutical and cosmetic products. Cacao seeds are a rich source of polyphenolic antioxidants, and consequently, it has been reported that cocoa‐based products contribute a greater proportion of the dietary intake of phenolic antioxidants than do green tea, wine, soya beans and blueberries, which are known antioxidant‐rich food products and beverages (Lee et al., 2003). The antioxidant properties of cocoa, particularly the high flavonoid content, are now of great interest due to its profound effects on human health. Specifically, the claim that cocoa polyphenols could prevent cancer or delay/slow down the progression of cancer (chemo‐preventive agents) has received increased attention (Martin et al., 2013). Furthermore, flavonoids extracted from cocoa have been shown to play a pivotal role in mediating innate and acquired immunity (Ramiro‐Puig and Castell, 2009), and also have been shown to have an effect on diet induced obesity and insulin resistance (Dorenkott et al., 2014). Emerging data support the suggestion that cocoa flavanols may serve as cardioprotective agents. These compounds have been reported to modulate mediators of inflammation (Keen et al., 2005). Cocoa flavanols and procyanidins (Bowser et al., 2017; Liu et al., 2015) have been shown to have beneficial effects including decreased platelet aggregation through increasing concentration of epicatechin and catechin in the plasma (Keen et al., 2005; Murphy et al., 2003). Furthermore, cacao shell is a rich source of theobromine and vitamin D. The pods contain a high level of potash that is used in soap production (Bart‐Plange and Baryeh, 2003).
Cocoa bean quality
It is also important to maintain or enhance bean quality. Recently, several bean quality attributes, both physical and chemical, that are required by the cocoa manufacturers/buyers, have been documented in detail to encourage the cacao community towards the production of better quality cocoa (CAOBISCO/ECA/FCC, 2015). These quality characteristics include flavour, purity or wholesomeness (e.g. free from bacteria, infestation, allergens, mycotoxins, heavy metals and pesticide residues), physical characteristics (e.g. consistency, yield of edible material bean, bean size and uniformity, shell content, fat content and moisture content) and cocoa butter characteristics (e.g. free fatty acid content). Some of these bean quality attributes, such as total fat content, acidity, total phenols, organic acids, heavy metals, amino acids, caffeine, theobromine, pH, sugars, macro‐ and micronutrient content, have been considered in the proposed Cocoa Quality Index (CQI) for Forastero‐type beans (Araujo et al., 2014). Such an indexing system may represent a useful tool in research programmes designed to improve bean quality for sustainable cocoa production. One recommended source of information on cocoa quality is the Cocoa Atlas (Rohsius et al., 2010; http://www.cocoa-atlas.org/). This DVD is funded by the German Cocoa and Chocolate Foundation and produced by the Cocoa Research Group of the Biocenter Klein‐Flottbek, University of Hamburg, Germany. It includes, in addition to global data, valuable information from 32 individual countries, with information from each country divided into 12 sections with the following titles: background notes, cocoa growing areas, cocoa production, cocoa trade, foreign trade, aroma description, bean weight/count, cut test, fat composition, free amino acids, further compounds and pictures of samples. Most recently, a Working Group on the Development of International Standards for the assessment of cocoa quality has been established by the Cocoa of Excellence (CoEx) programme and an initial draft document on this theme is now available (Cocoa of Excellence, 2017).
The flavour profile of beans is a key quality measure in cocoa. For instance, the clone CCN 51, which is planted extensively in Ecuador, exhibits many attractive agronomic traits like disease resistance, high butter content and high productivity; however, it is less popular among fine flavour chocolate manufacturers, especially due to the lack of fine flavour trait (Boza et al., 2014). In addition to the cacao genotype, several other factors such as location where the trees are grown (i.e. soil condition), the age of trees and postharvest treatments (fermentation, drying and roasting) also affect cocoa bean flavour (Kongor et al., 2016). A comprehensive overview of factors affecting cocoa flavour attributes has been published elsewhere (Afoakwa et al., 2008; Kongor et al., 2016). Furthermore, a sensory study conducted on raw cacao seeds and fruit pulp using a gas chromatography‐mass spectrometry method has identified monoterpenes, methylketones, and secondary alcohols and their respective esters as the main volatile aroma components in fine flavour clones such as SCA6 and EET62 (Kadow et al., 2013). Analytical methods such as MS fingerprinting (Qin et al., 2017; Tran et al., 2015) and the near‐infrared spectroscopy (NIRS) method (Krähmer et al., 2015) have been successfully employed in evaluation of cocoa biochemical quality parameters related to flavour attributes and quality of fermentation, as efficient and routinely applicable approaches. These studies have provided a foundation for understanding the molecular basis of fine aroma components in cocoa, and thereby for the development of molecular markers linked to fine aroma quality in this species.
Good preharvest and postharvest practices are key to maintaining many of the above mentioned bean quality descriptors (CAOBISCO/ECA/FCC, 2015). For instance, selection of suitable planting materials or the desired genetic background for cultivation is necessary to maintain the required flavour, yield, bean size and colour, and cocoa butter content (CAOBISCO/ECA/FCC, 2015; Loureiro et al., 2017). Furthermore, the quality of soil in which the cacao plants are grown is also a concern today as there is some evidence for the presence of heavy metals, especially cadmium, in cocoa beans produced in some parts of the producing countries (Arévalo‐Gardini et al., 2017; CAOBISCO/ECA/FCC, 2015; Loureiro et al., 2017).
Cocoa bean quality is also influenced by postharvest practices, especially the fermentation and drying processes. For example, controlled drying of the fermented cocoa beans is a crucial step to avoid development of off‐flavours that may affect quality of beans. High‐throughput molecular analysis tools could be used for rapid and efficient identification of microbial population diversity during cocoa fermentation and drying, and for development of microbial markers associated with the process (Hamdouche et al., 2015). For instance, the powerful biotyping tool, matrix‐assisted laser desorption ionization time‐of‐flight mass spectrometry (MALDI‐TOF MS) method, has recently been used for molecular identification of micro‐organisms involved in cocoa bean fermentation (Miguel et al., 2017; Schwenninger et al., 2016). Recently, fermentation‐like incubation systems or laboratory‐scale fermentation methods have proven to be applicable for the fermentation process of fresh cacao seeds (Evina et al., 2016; Kadow et al., 2015). This system, which does not depend on micro‐organisms, may provide a better alternative to the natural fermentation process that is usually difficult to control. Furthermore, the experimental model described in Lee et al. (2016) for cocoa fermentation that mimics on‐farm cocoa fermentation process may speed‐up fermentation studies at a laboratory level in the future.
Another important factor that influences the quality of cocoa beans is the specific environmental condition in which cacao plants are cultivated. The increasing atmospheric temperature and evapotranspiration caused by global warming are likely to have a profound impact on global cacao cultivation (Oyekale et al., 2009). Additionally, the climate in cacao growing regions has a considerable impact on cocoa fermentation and drying processes. Läderach et al. (2013) have projected that by 2050, the present cacao farming areas or cacao‐favoured growing areas in Côte d'Ivoire and Ghana may shift to areas with higher altitudes due to progressive increase in temperatures. A more recent detailed study of this topic is that by Schroth et al. (2016). If the predicted climate and weather variability continues, this may have an impact on the economic status of cocoa farmers and major cocoa‐producing countries; as result, global chocolate and confectionery industry is likely to be affected due to a cocoa shortage. Breeding for climate‐smart cacao varieties is vitally important to long‐term sustainability of cocoa production (World Cocoa Foundation, 2016). This subject of climate‐smart agriculture (CSA) is the basis of the ongoing project—“Mainstreaming CSA practices in cocoa production in Ghana,” which aims to implement CSA practices with cacao farmers (http://www.sustainablefoodlab.org/initiatives/climate-smart-agriculture/).
Cacao genetics and breeding
Cacao is a diploid fruit crop species with a relatively small genome, organized into ten chromosomes (da Silva et al., 2017); that is, the genome size is approximately double that of Arabidopsis thaliana, the model dicot. Recently, the published genome of the most cultivated type of cacao, T. cacao Matina 1‐6 clone reports a genome size of 445 Mbp (Motamayor et al., 2013), which is considerably larger than the previously published genome of a Criollo genotype (430 Mbp) (Argout et al., 2011). According to the genome statistics reported by Argout et al. (2011), 28 798 protein coding genes from more than 682 gene families are present in the cacao genome. These include many genes related to disease resistance, lipid biosynthesis (Zhang et al., 2015), flavonoid biosynthetic pathway and terpenoid synthesis (Argout et al., 2011). The updated version of this Criollo sequence, with 99% of genes anchored to the 10 chromosomes, was released in January 2017 and is accessible at the Cocoa Genome Hub (http://cocoa-genome-hub.southgreen.fr/) (Argout et al., 2017). Availability of whole‐genome sequences for several cacao varieties (Argout et al., 2011; Motamayor et al., 2013) has allowed identification and characterization of novel genes of interest to breeders and also development of molecular markers for marker‐assisted selection (MAS) (Lopes et al., 2011). The release of cacao genome sequences has also provided the way for rapid identification, functional and structural characterization of many gene families in cacao, through in silico computational studies and expression analysis. For example, recently three legumain proteins, TcLEG3, TcLEG6 and TcLEG9, which play diverse roles in programmed cell death, seed germination and seed development have been identified and characterized through in silico analyses, three‐dimensional modelling and expression analyses (Santana et al., 2016). Also, comprehensive genomewide analysis of pathogenesis‐related (PR) gene family in the two published T. cacao genomes has identified a set of candidate genes that are likely to be involved in mediating defence responses against major pathogens such as Phytophthora palmivora (Maora et al., 2017) and Colletotrichum theobromicola (Fister et al., 2016a). Arrays constructed from subtractive libraries have also been used in an investigation of molecular responses to cocoa black pod infection (Legavre et al., 2015). Such findings may contribute to a better understanding of the genetics and genomics of T. cacao.
In terms of breeding targets, these can be divided into two main categories. The first is associated with resistance to biotic stress, as unfortunately, outbreaks of diseases (Bailey and Meinhardt, 2016) in major cacao growing areas have significantly affected production in South America and Africa. For instance, Witches’ broom disease (WBD) (Almeida et al., 2017; Teixeira et al., 2015) caused by the fungal pathogen Crinipellis perniciosa has reduced cacao yields in many cultivation areas in South America including Ecuador and Brazil (Brown et al., 2005). In this context, a cacao osmotin‐like protein and various synthetic peptides (Falcao et al., 2016), and a phylloplanin (Freire et al., 2017) have been shown to have be involved in the response to WBD. Also with reference to pathogens, MALDI‐TOF MS methods have been applied recently for the rapid identification of M. perniciosa, Phytophthora palmivora, P. capsici, P. citrophthora, P. heveae, Ceratocystis cacaofunesta, C. paradoxa and C. fimbriata (dos Santos et al., 2017).
Another major disease problem in cacao is Cacao swollen shoot virus (CSSV) (Muller, 2016), which is transmitted largely by mealy bugs (Wetten et al., 2016). Although efforts have been made to eradicate the problem by removing infected trees, this has proved unsuccessful (Ameyaw et al., 2015) and it is now hoped that a greater understanding of the genetic variation in both the virus (Abrokwah et al., 2016; Chingandu et al., 2017a,2017b) and its vector (Herrbach et al., 2016), together with studies in more amenable model species (Friscina et al., 2017), will lead to progress in understanding this important disease and related badnaviruses (Andres et al., 2017; Bhat et al., 2016).
The second main breeding objective relates to physiological traits, as in addition to major pest and disease outbreaks, cacao cultivation is also affected by several other factors, which include altered short‐term climatic variation (e.g. El Niño), longer term global warming, high labour costs, depletion of soil fertility, poor plant productivity, lack of breeding strategies to develop and distribute improved varieties, and outdated farming practices (Zhang and Motilal, 2016). Specific breeding objectives reported in one recent study include dwarfism or semi‐dwarfism, which might enable smaller trees to be planted at higher density, and photosynthetic efficiency, an important determinant of yield (Pereira et al., 2017). As an alternative approach to breeding efforts to increase yields in the major production areas, some cocoa producers are now considering new regions that might allow an extension in the area under cultivation.
Importantly in terms of breeding strategies, cacao has a relatively longer juvenile period, namely 3–5 years. This makes selection of fruit‐specific traits in breeding programmes more time‐consuming and expensive, as the trees must be maintained for a longer period of at least three years to visually observe such characters in pods. Moreover, this crop is primarily outbreeding (i.e. SCA 6 and EET 75 cacao clones), and therefore, many populations are mostly heterozygous. This makes generation of inbred lines from crosses more labour‐intensive, and doubled haploid lines (Dunwell, 2010) are not easily generated. Moreover, the self‐incompatibility that exists in some of the cultivated cacao clones means that breeding populations are often highly heterogeneous with a wide range of yields (Royaert et al., 2011). However, it should also be noted that genetic variability does exist in cacao populations, and there are several self‐compatible cacao clones, such as CCN 51 and ICS 6 (Cervantes‐Martinez et al., 2006). Cacao trees also require a large area of land and high input of resources, including labour, for their maintenance under field conditions. These characteristics have made this crop less attractive as a model system, although like Arabidopsis, it has a relatively small genome.
Because of the recalcitrant (do not survive drying) nature of its seeds, the germplasm of this allogamous tree crop must be conserved in field genebanks as a living collection (Motilal et al., 2013) or by cryopreservation (Adu‐Gyamfi and Wetten, 2012; Adu‐Gyamfi et al., 2016). The largest collections of cacao germplasm are those at the International Cocoa Genebank, Trinidad (ICG, T) (https://sta.uwi.edu/cru/index.asp) (2400 accessions) and at the Centro Agronomico Tropical de Investigación y Enseñanza (CATIE) in Costa Rica (https://www.catie.ac.cr/en/) (1146 accessions), with another collection (c. 400 accessions), the International Cocoa Quarantine Centre, housed at the University of Reading, UK (http://www.icgd.reading.ac.uk/icqc/). However, maintaining genetic resources as living collections in situ or ex situ is practically difficult and is also an expensive process. A significant number of mislabelled accessions have been reported in these field genebanks (Motilal and Butler, 2003; Motilal et al., 2012). Thus, an efficient strategy to eliminate these mislabelled and/or duplicated accessions in large cacao germplasm collections is required for efficient and accurate management of genetic resources. In this context, DNA fingerprinting as a screening tool has been extensively used in rapid and accurate identification of cacao accessions. Restriction fragment length polymorphisms (RFLPs), random amplified polymorphic DNA (RAPD), amplified fragment length polymorphisms (AFLPs), microsatellites and single nucleotide polymorphisms (SNPs) are some of the molecular markers commonly used in cacao molecular studies (Kuhn et al., 2012; Lanaud et al., 1999; Laurent et al., 1994; Motilal and Butler, 2003; Santos et al., 2012a; Turnbull et al., 2004). Livingstone et al. (2012) report an optimized 5′‐nuclease (TaqMan)‐based SNP assay for efficient genotyping of cacao trees under field conditions. This simple, cost‐effective method would be a useful technique for cacao breeders.
Development of high‐density molecular‐linkage maps and characterization of molecular markers linked to major quantitative trait loci (QTL) have greatly accelerated breeding programmes in cacao by facilitating examination of particular fruit‐specific characters at a genotype level. The QTL mapping studies of this species have been performed using different mapping populations, that is F1 or F2 mapping populations (Lanaud et al., 2009; Motamayor et al., 2013; Royaert et al., 2011; Schnell et al., 2007) and association mapping or linkage disequilibrium mapping systems (Marcano et al., 2007; Stack et al., 2015). The first genomic map of cacao with a total of 193 loci covering 759 cM in 10 linkage groups was published by Lanaud et al. (1995). RFLP and RAPD markers were mainly used to construct this genetic map, which was subsequently used to produce high‐density molecular‐linkage maps in several subsequent studies. For example, Risterucci et al. (2000) published a high‐resolution molecular‐linkage map comprising 424 markers covering 885.4 cM over ten linkage groups; AFLP and simple sequence repeat (SSR) markers were employed to construct this high‐density map, which was considered as a good reference map for research activities in cacao (Clément et al., 2001). Development and mapping of codominant microsatellite markers to the cacao genome has accelerated genetic studies and breeding experiments. These PCR based codominant SSR markers are highly polymorphic, and easily transferrable between/across populations and/or laboratories; they can thus be used in MAS (Pugh et al., 2004). The high‐density linkage map described in this study has 465 markers with 268 SSR markers. A small number of such SSR markers have also been used in a study of genetic diversity in historical cacao plantations in Brazil (Santos et al., 2015) and germplasm assessments in Indonesia (Dinarti et al., 2015) and Cuba (Martínez et al., 2017). The collection of T. cacao expressed sequence tags (ESTs) generated from a range of organs, genotypes and environmental conditions is a valuable resource for discovery of important candidate genes and molecular markers for cacao genetic improvements (Argout et al., 2008). For instance, Fouet et al. (2011) discovered 174 EST‐based SSRs markers by screening a cacao EST data set and developed a high‐density linkage map with 582 codominant markers including 384 SSR markers. Most recently, da Silva et al. (2017) used a set of 20 EST‐SSRs to examine the evolutionary relationship of species within the Theobroma genus. Also, within the last few years, the use of SNP‐based codominant markers in genetics has increased significantly, due to advances in high‐throughput sequencing systems. A SNP‐based linkage map for cacao was initially developed by Allegre et al. (2012). This genetic linkage map contains a set of 1262 markers spanning in a length of 734 cM, and of these markers, 681 are EST‐based SNPs.
Recently, a large number of SNPs have been detected by aligning RNA sequence (RNAseq) data of 16 cacao cultivars to the assembled Matina 1–6 transcriptome (Livingstone et al., 2015). In this study, a saturated genetic linkage map with 2589 SNPs was constructed. More importantly, this study led to the development of an Illumina Infinium SNP array for cacao—Cacao6kSNP array that consisted of 6000 high‐quality SNPs. The newly developed array and the SNP data reported by Livingstone et al. (2015) provide a valuable genomic resource for cacao breeding. The latest genetic linkage map of cacao includes SNP data obtained from a large mapping population (459 trees) of a cross between WBD resistant, TSH 1,188 and WBD tolerant (moderately resistant to WBD) CCN (Royaert et al., 2016). It contains 3526 SNP markers and has a length of 852.8 cM. In addition to genetic linkage mapping studies, several recent studies have highlighted the importance of SNP‐based DNA fingerprinting in assessing cacao bean authentication (Fang et al., 2014), cacao variety development (Padi et al., 2015, 2017) and cacao genetic diversity (Cosme et al., 2016).
This rapid discovery of molecular markers also permits the efficient identification and study, in cacao, of the genetic basis of QTL for many agronomic traits such as bean traits and the number of ovules per ovary (Clement et al., 2003b), butter content and its hardness in cocoa beans (Araújo et al., 2009), diseases resistance for Ceratocystis wilt (Santos et al., 2012b), resistance for Phytophthora species (Akaza et al., 2016; Clement et al., 2003a; Efombagn et al., 2016; Flament et al., 2001; Lanaud et al., 2004; Legavre et al., 2015; Motilal et al., 2016; Risterucci et al., 2003), resistance for WBD (Brown et al., 2005; Faleiro et al., 2006; Motilal et al., 2016; Queiroz et al., 2003; Royaert et al., 2016), number of filled seeds (Motilal et al., 2016), yield (Clement et al., 2003a; Crouzillat et al., 2000) and self‐compatibility/incompatibility (Royaert et al., 2011; da Silva et al., 2016). Furthermore, meta‐analysis on QTL related to disease resistance in cacao has been performed by Lanaud et al. (2009). Such information would be of great use in MAS, which, to date, is employed in many crop breeding programs for development of improved cultivars including cacao. Recently, a semi‐automated genotyping platform for MAS known as amplicon sequencing (AmpSeq) has been successfully applied for grapevine breeding programme (Yang et al., 2016). This study showed the applicability of this strategy in heterozygous crop breeding by generation of AmpSeq markers for several traits with high breeding value including disease resistance in grapevine. It is possible that such a high‐throughput, cost‐effective, flexible and rapid breeding strategy could be implemented with some modifications to assist MAS in cacao in the future.
In addition to markers based on nuclear genomic DNA, several loci of chloroplast DNA (cpDNA) such as matK, rbcL and trnH‐psbA can be used as markers in DNA barcoding (Bieniek et al., 2015). Recently, sequence variation of the trnH‐psbA intergenic spacer has been analysed in 28 cacao accession obtained from different farms in southern Mexico (Gutiérrez‐López et al., 2016). It was found that the indels located in this region could be considered as potential markers for development of a DNA barcoding system in cacao. These markers are useful in identification of accessions in situations when other marker systems can only discriminate between accessions on the basis of a very small number of SSR markers (Gutiérrez‐López et al., 2016).
Propagation methods and in vitro embryogenesis
Genetic improvement of cacao for improved traits has been hindered due to its narrow genetic base and long life cycle (Li et al., 1998). It is estimated that approximately 30% of world cacao production is lost due to pest and diseases, annually (Guiltinan et al., 2008). Therefore, an efficient propagation method for cacao is essential to accelerate breeding programmes and to avoid production shortages in the future. The cacao crop is grown with an approximate planting density of 1100 trees per hectare, and it has been estimated that with a replanting rate of 10%, there is an annual requirement for one billion units. This requirement is not being met at present, and some of the alternative propagation options (Laliberte and End, 2015) are considered below.
Cacao is normally propagated by means of seeds. Additionally, to maintain a genetically stable population, it is also propagated through a number of vegetative/asexual methods, of which a variety of grafting methods (Miguelez‐Sierra et al., 2017) are the most commonly practiced; these methods have been reviewed in detail by Sena Gomes et al. (2015). However, these propagation systems are not widely practiced in developing countries (Maximova et al., 2002). This could be due to the low rate of propagation and undesirable morphological features observed in some propagules, which often lack normal dimorphic nature and display bush‐like growth with a fibrous root system. Therefore, maintenance of such material is a more labour‐intensive process and requires skilled workers (Traore et al., 2003).
In cacao, in vitro embryogenesis or somatic embryogenesis (SE) is an alternative to traditional propagation methods and allows rapid clonal propagation of true‐to‐type plants with normal dimorphic architecture and taproot formation. Importantly, this system has shown to be an effective method in propagation of CSSV disease‐free plantlets (Quainoo et al., 2008). Moreover, a study conducted by López and co‐workers in 2010 found that de novo genetic mutations and epigenetic modifications do not accumulate with ageing of in vitro induced cacao calli (López et al., 2010b); this conclusion is also relevant to a recent study of long‐term SE (Quinga et al., 2017). Additionally, the SE system has been utilized in cacao germplasm conservation through cryopreservation (Adu‐Gyamfi and Wetten, 2012; Adu‐Gyamfi et al., 2016) and in genetic transformation (Maximova et al., 2002; da Silva et al., 2008). Induction of cacao somatic embryos has been observed from a range of its tissues, that is zygotic embryos (Pence et al., 1980), floral parts—petals and staminodes (Alemanno et al., 1996, 1997; Boutchouang et al., 2016; Li et al., 1998; Tan and Furtek, 2003), and nucellar tissues (Figueira and Janick, 1993). The SE method developed by Li et al. (1998) was applicable to many different cacao genotypes, and later, Maximova et al. (2002) improved this system to produce secondary somatic embryos from primary somatic embryos. Somatic embryo‐derived plants have been successfully grown under field conditions (Maximova and Guiltinan, 2015; Maximova et al., 2008). However, the efficiency of SE in this species is strongly influenced by genotype, particularly in respect of the conversion rate of mature somatic embryos into complete plants (Maximova et al., 2002). In addition, the type of explant used and its position, that is flower bud position (Boutchouang et al., 2016; Traore and Guiltinan, 2006), tissue culture media composition (Traore and Guiltinan, 2006) and phenological parameters such as the periodicity of new leaf development (Issali et al., 2008) are also have an impact on SE efficiency in cacao. Several researchers have focused on optimizing in vitro culture media composition to improve somatic embryo differentiation in this species (Minyaka et al., 2008; Niemenak et al., 2008, 2012; Traore and Guiltinan, 2006). Furthermore, establishment of a temporary immersion bioreactor system for mass production of cacao somatic embryos was a breakthrough process in cacao biotechnology research (Niemenak et al., 2008). Consequently, studies were designed to further optimize somatic embryo induction in liquid suspension cultures (Niemenak et al., 2012). More recently, an alternative method to induce cacao primary SE at a high efficiency has been achieved by supplementing DKW medium with different ratios of kinetin to 2,4‐dichlorophenoxyacetic acid (1.0: 3.9 callus induction medium; 1.0: 7.8 secondary callus growth medium) (Ajijah et al., 2016). Furthermore, this methodology yielded a 65% plantlet conversion rate and a relatively low percentage of somaclonal variation (López et al., 2010a). Other beneficial modifications to SE media have been reported recently by Kouassi et al. (2017) and Modeste et al. (2017).
Further details and modified SE protocols developed by the various commercial chocolate companies are also given in the various patent documents mentioned in the section below. Despite continuous progress, the overall low efficiency and reproducibility of the methods developed as well as the genotype dependent nature of the many steps involved in the SE process still present a significant challenge for mass propagation of many elite cacao genotypes at the commercial scale required for many parts of the cacao growing regions (da Silva et al., 2008). In an ambitious scheme, Nestlé's Cocoa Plan aims to produce and distribute at least 12 million plants of elite varieties that are disease free, high yielding and high quality in terms of beans and taste by the year 2022 (Fair Labor Association, 2012; Guillou et al., 2014). They aim to improve farmers’ income and living conditions and avoid deforestation through sustainable production of cocoa, technology transfer and distribution of quality planting material for propagation in Côte d'Ivoire.
Understanding the molecular mechanism of SE would allow us to improve this process in economically important crops, including cacao, and thereby provide an efficient system to speed‐up commercial plant production of many crops in the coming years. Recent advances in high‐throughput sequencing systems and ‘omics’ resources have facilitated generation of high‐resolution transcriptome data for plant embryogenesis, both in vivo and in vitro and thereby, to provide novel insights into the molecular basis of embryogenesis (Wickramasuriya and Dunwell, 2015; Xu et al., 2012). In a relevant recent example in cacao, gene expression profiles of zygotic embryogenesis (ZE) and SE have been generated using whole‐genome microarray (Maximova et al., 2014). This study reported that a large number of genes including those encoding for transcription factors, genes related to flavonoid and lipid biosynthesis were differentially expressed between the two embryo developmental processes. Such results thus provide an insight into cacao SE at a molecular level, and the information provided could be used to develop and characterize novel molecular markers for SE. In addition, proteome profiles of cacao SE and their equivalent ZE at various developmental stages have been generated and analysed through 2D PAGE and nano‐LC‐MC (Niemenak et al., 2015; Noah et al., 2013). Availability of the genome sequence together with such recent proteomic and transcriptomic information for cacao embryogenesis provides a good starting point for functional studies of many genes and their encoded proteins essential for embryo development in this species.
In addition, several key regulators of plant embryogenesis have also been isolated and characterized from cacao. Of these, the members of the Leafy cotyledon (LEC) gene family—LEC1, LEC2 and FUSCA3 (FUS3)—serve as master regulators of embryo development, and they have been well characterized in Arabidopsis (Lotan et al., 1998; Stone et al., 2001). Recently, a functional ortholog of Arabidopsis LEC2 has been isolated and characterized in T. cacao (TcLEC2) by Zhang et al. (2014). This gene was found to be expressed at a significant level in endosperm and cotyledons but not in flower and leaf tissues (Zhang et al., 2014). Furthermore, a 20‐fold higher level of TcLEC2 transcript accumulation was observed in embryogenic calli than in nonembryogenic calli (Zhang et al., 2014). Moreover, overexpression of TcLEC2 has led to increased expression of several seed‐specific genes in leaves of cacao, that is TcAGL15 (>129‐fold), TcABI3 (>9‐fold) and WRINKLED1 (WRI1) (>10‐fold). This also increased the embryogenic competency of cotyledon explants and regeneration capacity of somatic embryos, supporting the fact that, as in many other plants, TcLEC2 is a key regulator of cacao embryogenesis. Furthermore, a functional homologue of the LEC1‐like gene has also been reported from the cacao genome (Alemanno et al., 2008); increased expression of this gene has been detected in early stages of cacao zygotic and somatic embryogenesis.
Another well‐studied regulator of embryogenesis is the AP2/ERF family member BABY BOOM (BBM), which was first identified from Brassica napus microspore‐derived in vitro embryos (Boutilier et al., 2002). Being significantly expressed in developing embryos and seeds, BBM is considered as one of the key marker genes in embryogenesis (Boutilier et al., 2002; Ikeda et al., 2006; Karami et al., 2009). A functional gene with a high degree of similarity to BBM in A. thaliana has been isolated and characterized from cacao (TcBBM) (Florez et al., 2015). TcBBM expression has been detected throughout embryogenesis in cacao; a higher level of expression has been detected in SE than in ZE. In a manner similar to that in species including Arabidopsis (Boutilier et al., 2002) and cereals (Lowe et al., 2016), overexpression of TcBBM in cacao has been found spontaneously to induce somatic embryos in hormone‐free media (Florez et al., 2015); thus, TcBBM could be used to enhance the efficiency of SE in cacao. The most recent advance in this area is the report of an inducible SE system by exploiting a dexamethasone activatable embryogenic transcription factor to promote somatic embryo formation from juvenile leaves (Shires et al., 2017).
Similarly, kinases play an important role in plant embryogenesis. For example, somatic embryogenesis receptor kinases (SERKs) are a subgroup of protein kinase genes that are expressed in early stages of somatic and zygotic embryo development. These genes were initially isolated from in vitro embryogenic cultures of carrot by Schmidt et al. (1997) and have subsequently SERKs been identified and characterized in many species, that is A. thaliana (Hecht et al., 2001), Solanum tuberosum (Sharma et al., 2008), Cocos nucifera (Perez‐Nunez et al., 2009), Zea mays (Baudino et al., 2001), Momordica charantia (Talapatra et al., 2014) and cacao (de Oliveira Santos et al., 2005). This latter gene (TcSERK) was highly expressed in embryogenic calli, and also in mature zygotic and somatic embryos at a moderate level, suggesting that the functional copy of SERK found in cacao plays a key role during the process of embryo development.
Genetic transformation
Completion of whole‐genome sequencing for many economically important crops has significantly contributed to their respective genetic improvement. In addition, identification and functional characterization of novel genes and transfer of genes that regulate agronomically valuable traits such as disease resistance have been achieved in many crops. The earliest attempt of cacao transformation was recorded in 1994 by Sain et al. (1994) using the Agrobacterium‐mediated gene transfer method. Although transformed callus cells derived from leaf tissues were obtained, no plant regeneration was recorded from those transformed cells. This was due to the lack of an efficient protocol to recover plants from cacao leaf tissue‐derived calli at that time (Sain et al., 1994).
Subsequently, a more efficient method for stable genetic transformation and recovery of transformed plants from transformed cacao cells was established by Maximova et al. (2003). This study employed SE as a regeneration system together with Agrobacterium tumefaciens cocultivation to obtain transgenic plants. This system has been successfully used to produce transgenic cacao plants overexpressing cacao class I Chitinase gene (TcChil1) (Maximova et al., 2006). These transgenic plants showed enhanced fungal pathogen resistance against Colletotrichum gloeosporioides. Although this transformation system was proven to be reproducible (Maximova et al., 2003, 2006), the reliable production of a large number of transgenic embryos remains a challenge. Subsequently, several studies have been conducted in an attempt to improve the efficiency of the transformation method described in Maximova et al. (2003) (Silva et al., 2009). Most recently, an optimized method for transient transformation of cacao for several genotypes through Agrobacterium infiltration has been published by Fister et al. (2016b). As a tool, this will allow more efficient in vivo functional analysis of cacao genes, subcellular localization of proteins and promoter analysis. It is also possible that the use of rapidly flowering transgenic lines, as used in other perennial species (Callahan et al., 2016), may be applicable to cacao. Most recently, an inducible SE system in cacao has been developed by use of a transgenic plant expressing the LEC2 embryogenic transcription factor (Shires et al., 2017).
Although much effort has been devoted to improve cacao varieties through genetic transformation, genetically modified (GM) cacao material has not been released commercially so far, and the studies have been limited to laboratories and greenhouses. Although genetic transformation serves as a valuable tool in crop improvements, the future of GM cacao is not clear as the consumer acceptance of food from genetically modified organisms is still a controversial issue in some countries (Dunwell, 2014; Guiltinan et al., 2008).
It is now possible to effect gene‐specific mutagenesis by genome editing, (e.g. CRISPR/Cas9) for functional characterization of genes; this also serves as a promising approach for genetic manipulation of disease resistance, and other genes in cacao. Whether or not such methods will be considered as a form of GM is still uncertain, at least in Europe.
Resources available for cacao functional studies
Open‐source bioinformatic tools and web databases have greatly contributed to the rapid developments in omics‐based researches and thereby crop improvements. A list of tools/databases freely accessible for cacao researchers and breeders is summarized in Table 1. The latest genome sequence of cacao is available on the Cacao Genome Database (CGD, http://www.cacaogenomedb.org/), which was developed in collaboration with MARS, USDA/ARS, IBM, Clemson University Genomics Institute, PIPRA, HudsonAlpha Institute for Biotechnology, National Center for Genome Resources, Indiana University and Washington State University—Main Lab Bioinformatics. In addition, tools such as BLAST and GBrowse can be accessed through this web database.
Table 1.
A list of web based resources available for cacao
| Resource | URL |
|---|---|
| CacaoNet—Global Network for Cacao Genetic Resources | https://sites.google.com/a/cgxchange.org/cacaonet/home/partners-of-cacaonet |
| International Cocoa Germplasm Database (ICGD) | http://www.icgd.reading.ac.uk/index.php |
| Cacao Genome Database | http://www.cacaogenomedb.org/ |
| Cocoa Genome Hub | http://cocoa-genome-hub.southgreen.fr/ |
| CocoaGenDB | http://cocoagendb.cirad.fr/ |
| CEMID—Cocoa EST Marker Information Database | http://riju.byethost31.com/cocoa/?ckattempt=1 |
| Ensembl Plants | http://plants.ensembl.org/Theobroma_cacao/Info/Index |
| Dicots PLAZA 3.0 | http://bioinformatics.psb.ugent.be/plaza/versions/plaza_v3_dicots/organism/view/Theobroma+cacao |
| Phytozome v11.0 | https://phytozome.jgi.doe.gov/pz/portal.html#!info?alias=Org_Tcacao |
| GenBank NCBI | http://www.ncbi.nlm.nih.gov/genome/?term=cocoa |
| Witches’ Broom Disease Transcriptome Atlas | http://bioinfo08.ibi.unicamp.br/wbdatlas/ |
Survey of patents relating to cacao
Often, very useful information about advances in scientific research can be obtained from a study of patent databases, as information is often published here before appearing in more usual scientific publications (Dunwell, 2012). In addition to academic institutes, private sector industries play a key role in cacao research and development. Hence, a patent search analysis was performed using the Lens patent database (https://www.lens.org/lens/) to provide an overview of public and private sector involvement, and application of biotechnology techniques in cacao research; structured search was carried out on granted patents using the terms ‘Theobroma ‘ or ‘cacao’ or ‘cocoa’ in the abstract or claims. A total of 2732 granted patents were recorded prior to 13 July 2017. Thus, because it is not feasible to summarize all these granted patents in this review only a selected summary is provided here (Tables 2). In brief, granted patents related to plant breeding and biotechnology applications were filtered based on the International Patent Classification (IPC) codes; a total of 10 granted patents with following IPC codes were identified and listed in Table 2: A01G 17/00, A01H 4/00, A01H 5/08, C12N 5/00, C12N 5/02, C12N 5/04, C12N 15/82, C12N 15/84 and C12N 15/87 (definition of these IPC codes can be obtained through the World Intellectual Property Organization (WIPO) page http://www.wipo.int/classifications/ipc/en/). We identified several granted patents relating to the production of cacao somatic embryos through optimized tissue culture techniques. Not surprisingly, several studies are funded either fully or partially by the leading chocolate manufacturers. Notable publications of particular relevance to this review include those describing methods for SE with granted patents from Hershey Foods (US 5312801) in 1994, Penn State Research Foundation (US6150587) in 2000, Nestle S.A. (US 8921087) in 2014, and most recently a granted patent from Mars Inc. (AU 2014/353082 B2) in April 2017. This patent on the production of cacao plants claims micropropagation via direct SE and is available at https://www.lens.org/images/patent/AU/2014353082/B2/20170420/AU_2014_353082_B2.pdf. This method uses explants such as staminodes and petal base tissues for induction of primary embryos in a medium supplemented with 6‐benzylaminopurine (BAP). Subsequently, the epicotyl segments removed from primary embryos are placed in an induction medium containing BAP to induce direct secondary embryos, followed by further embryo development in a medium containing gibberellic acid, if needed. All the cultures are maintained in the light (photoperiod: 16 : 8 (light: dark) at a temperature of 23–29 °C for a sufficient period of time to obtain embryos. Although several attempts have been made in the past to develop a direct SE system for cacao micropropagation, there has been only limited success (Pence et al., 1980). Therefore, the detailed information provided in this patent related to a direct SE method may provide additional valuable information for cacao researchers working on this subject. However, there is no patent information yet on the use of novel breeding techniques such as Zinc Finger Nuclease (ZFN) or CRISPR/Cas9 technology, cisgenesis and RNA‐dependent DNA methylation (RdDM) in cacao plants.
Table 2.
A list of granted patents, in chronological order, on T. cacao biotechnology, prior to July 2017
| Publication Number | Publication Year | Title | Applicant(s) |
|---|---|---|---|
| US 4301619 A | 1981 | Plant tissue produced by non‐agricultural proliferation of Cacao embryos | Purdue Research Foundation [owner] |
| US 4291498 A | 1981 | Method for production of mature asexual Cacao embryos, and product thereof | Purdue Research Foundation |
| US 4306022 A | 1981 | Cocoa bean cell culture | Cornell Research Foundation Inc |
| US 4545147 A | 1985 | Asexual embryogenesis of callus from Theobroma Cacao L. | Purdue Research Foundation [owner] |
| US 5312801 A | 1994 | Somatic embryogenesis and plant regeneration of Cacao | DNA Plant Technology Corporation |
| Hershey Foods Corporation [owners] | |||
| US 6150587 A | 2000 | Method and tissue culture media for inducing somatic embryogenesis, Agrobacterium‐mediated transformation and efficient regeneration of Cacao plants | Penn State Research Foundation [owner] |
| US 8921087 B2 | 2014 | Cocoa somatic embryogenesis | Florin Bruno Jean‐Marie |
| Masseret Bernard | |||
| Vachet Caroline Denise Monique | |||
| Nestec SA[owner] | |||
| US 8969655 B1 | 2015 | Modulation of flavonoid content in Cacao plants | Daniel Preston |
| Randall B. Murphy | |||
| Cacao Biotechnologies LLC [owner] | |||
| US 9428759 B2 | 2016 | Methods for increasing the production of phenolic compounds from Theobroma Cacao | Rengifo Raul Cuero |
| Casa Luker SA [owner; International Park of Creativity] | |||
| AU 2014/353082 B2 | 2017 | Production of plants using somatic embryogenesis | Mars Inc [owner] |
Conclusion
There is a great demand for high‐quality cocoa beans. Thus, to ensure long‐term sustainability of cocoa production, future research should focus on the development of improved cacao varieties that can both tolerate changing climates, but also meet the stringent quality criteria demanded by the chocolate industry. Implementation of modern molecular tools in cacao biotechnology research will undoubtedly be an integral part of this process.
Conflict of Interest
The authors declare no conflict of interest.
References
- Abrokwah, F. , Dzahini‐Obiatey, H. , Galyuon, I. , Osae‐Awuku, F. and Muller, E. (2016) Geographical distribution of the Cacao swollen shoot virus molecular variability in Ghana. Plant Dis. 100, 2011–2017. [DOI] [PubMed] [Google Scholar]
- Adu‐Gyamfi, R. and Wetten, A. (2012) Cryopreservation of cocoa (Theobroma cacao L.) somatic embryos by vitrification. Cryo Letters, 33, 494–505. [PubMed] [Google Scholar]
- Adu‐Gyamfi, R. , Wetten, A. and López, C.M.R. (2016) Effect of cryopreservation and post‐cryopreservation somatic embryogenesis on the epigenetic fidelity of cocoa (Theobroma cacao L.). PLoS ONE, 11, e0158857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Afoakwa, E.O. , Paterson, A. , Fowler, M. and Ryan, A. (2008) Flavor formation and character in cocoa and chocolate: a critical review. Crit. Rev. Food Sci. Nutr. 48, 840–857. [DOI] [PubMed] [Google Scholar]
- Ajijah, N. , Hartati, R.S. , Rubiyo, R. , Sukma, D. and Sudarsono, S. (2016) Effective cacao somatic embryo regeneration on kinetin supplemented DKW medium and somaclonal variation assessment using SSRs markers. AGRIVITA 38, 80–92. [Google Scholar]
- Akaza, J.M. , Kouassi, A.B. , Akaffou, D.S. , Fouet, O. , N'Guetta, A.S.‐P. and Lanaud, C. (2016) Mapping QTLs for Black pod (Phytophthora palmivora) resistance in three hybrid progenies of cocoa (Theobroma cacao L.) using SSR markers. Int. J. Sci. Res. Public. 6, 298–311. [Google Scholar]
- Alemanno, L. , Berthouly, M. and Michaux‐Ferriere, N. (1996) Histology of somatic embryogenesis from floral tissues cocoa. Plant Cell Tiss. Organ Cult. 46, 187–194. [Google Scholar]
- Alemanno, L. , Berthouly, M. and Michaux‐Ferriere, N. (1997) A comparison between Theobroma cacao L. zygotic embryogenesis and somatic embryogenesis from floral explants. In Vitro Cell. Dev. Biol.‐Plant, 33, 163–172. [Google Scholar]
- Alemanno, L. , Devic, M. , Niemenak, N. , Sanier, C. , Guilleminot, J. , Rio, M. , Verdeil, J.‐L. et al. (2008) Characterization of leafy cotyledon1‐like during embryogenesis in Theobroma cacao L. Planta, 227, 853–866. [DOI] [PubMed] [Google Scholar]
- Allegre, M. , Argout, X. , Boccara, M. , Fouet, O. , Roguet, Y. , Berard, A. , Thevenin, J.M. et al. (2012) Discovery and mapping of a new expressed sequence tag‐single nucleotide polymorphism and simple sequence repeat panel for large‐scale genetic studies and breeding of Theobroma cacao L. DNA Res. 19, 23–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Almeida, D.S.M. , Gramacho, K.P. , Cardoso, T.H.S. , Micheli, F. , Alvim, F.C. and Pirovani, C.P. (2017) Cacao Phylloplane: the first battlefield against Moniliophthora perniciosa, which causes witches’ broom disease. Phytopathology, 107, 864–871. [DOI] [PubMed] [Google Scholar]
- Ameyaw, G.A. , Dzahini‐Obiatey, H.K. , Domfeh, O. , Oppong, F.K. and Abaka‐Ewusie, K. (2015) History and data analyses of ‘cutting out’ method for Cocoa Swollen Shoot Virus Disease (CSSVD) control in Ghana. J. Plant Dis. Prot. 122, 200–206. [Google Scholar]
- Andres, C. , Gattinger, A. , Dzahini‐Obiatey, H.K. , Blaser, W.J. , Offei, S.K. and Six, J. (2017) Combatting Cocoa swollen shoot virus disease: what do we know? Crop Protect. 98, 76–84. [Google Scholar]
- Araujo, Q.R. , Fernandes, C.A. , Ribeiro, D.O. , Efraim, P. , Steinmacher, D. , Lieberei, R. , Bastide, P. et al. (2014) Cocoa quality index – a proposal. Food Control, 46, 49–54. [Google Scholar]
- Araújo, I.S. , de Souza Filho, G.A. , Pereira, M.G. , Faleiro, F.G. , de Queiroz, V.T. , Guimarães, C.T. , Moreira, M.A. et al. (2009) Mapping of quantitative trait loci for butter content and hardness in cocoa beans (Theobroma cacao L.). Plant Mol. Biol. Rep. 27, 177–183. [Google Scholar]
- Arévalo‐Gardini, E. , Arévalo‐Hernández, C.O. , Baligar, V.C. and He, Z.L. (2017) Heavy metal accumulation in leaves and beans of cacao (Theobroma cacao L.) in major cacao growing regions in Peru. Sci. Total Environ. 605–606, 792–800. [DOI] [PubMed] [Google Scholar]
- Argout, X. , Fouet, O. , Wincker, P. , Gramacho, K. , Legavre, T. , Sabau, X. , Risterucci, A.M. et al. (2008) Towards the understanding of the cocoa transcriptome: production and analysis of an exhaustive dataset of ESTs of Theobroma cacao L. generated from various tissues and under various conditions. BMC Genom. 9, 512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Argout, X. , Salse, J. , Aury, J.M. , Guiltinan, M.J. , Droc, G. , Gouzy, J. , Allegre, M. et al. (2011) The genome of Theobroma cacao . Nat. Genet. 43, 101–108. [DOI] [PubMed] [Google Scholar]
- Argout, X. , Martin, G. , Droc, G. , Fouet, O. , Labadie, K. , Rivals, E. , Aury, J.M. et al. (2017) The cacao Criollo genome v2.0: an improved version of the genome for genetic and functional genomic studies. BMC Genom. 18, 730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bailey, B.A. and Meinhardt, L.W. (2016) Cacao Diseases: A History of Old Enemies and New Encounters. Switzerland: Springer International Publishing. [Google Scholar]
- Bart‐Plange, A. and Baryeh, E.A. (2003) The physical properties of Category B cocoa beans. J. Food Eng. 60, 219–227. [Google Scholar]
- Baudino, S. , Hansen, S. , Brettschneider, R. , Hecht, V.F. , Dresselhaus, T. , Lörz, H. , Dumas, C. et al. (2001) Molecular characterisation of two novel maize LRR receptor‐like kinases, which belong to the SERK gene family. Planta, 213, 1–10. [DOI] [PubMed] [Google Scholar]
- Beg, M.S. , Ahmad, S. , Jan, K. and Bashir, K. (2017) Status, supply chain and processing of Cocoa ‐ a review. Trends Food Sci. Technol. 66, 108–116. [Google Scholar]
- Bhat, A.I. , Hohn, T. and Selvarajan, R. (2016) Badnaviruses: the current global scenario. Viruses, 8, 177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bieniek, W. , Mizianty, M. and Szklarczyk, M. (2015) Sequence variation at the three chloroplast loci (matK, rbcL, trnH‐psbA) in the Triticeae tribe (Poaceae): comments on the relationships and utility in DNA barcoding of selected species. Plant Syst. Evol. 301, 1275–1286. [Google Scholar]
- Boutchouang, R.P. , Akitio, O.F.Z. , Tchouatcheu, A.G.N. and Niemenak, N. (2016) Influence of the position of flowers buds on the tree on somatic embryogenesis of cocoa (Theobroma cacao L.). Int. J. Plant Physiol. Biochem. 8, 7–16. [Google Scholar]
- Boutilier, K. , Offringa, R. , Sharma, V.K. , Kieft, H. , Ouellet, T. , Zhang, L. , Hattori, J. et al. (2002) Ectopic expression of BABY BOOM triggers a conversion from vegetative to embryonic growth. Plant Cell, 14, 1737–1749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowser, S.M. , Moore, W.T. , McMillan, R.P. , Dorenkott, M.R. , Goodrich, K.M. , Ye, L. , O'Keefe, S.F. et al. (2017) High‐molecular‐weight cocoa procyanidins possess enhanced insulin‐enhancing and insulin mimetic activities in human primary skeletal muscle cells compared to smaller procyanidins. J. Nutr. Biochem. 39, 48–58. [DOI] [PubMed] [Google Scholar]
- Boza, E.J. , Motamayor, J.C. , Amores, F.M. , Cedeno‐Amador, S. , Tondo, C.L. , Livingstone, D.S. , Schnell, R.J. et al. (2014) Genetic characterization of the cacao cultivar CCN 51: its impact and significance on global cacao improvement and production. J. Am. Soc. Hort. Sci. 139, 219–229. [Google Scholar]
- Brown, J.S. , Schnell, R. , Motamayor, J. , Lopes, U. , Kuhn, D.N. and Borrone, J.W. (2005) Resistance gene mapping for witches’ broom disease in Theobroma cacao L. in an F2 population using SSR markers and candidate genes. J. Am. Soc. Hort. Sci. 130, 366–373. [Google Scholar]
- Callahan, A.M. , Srinivasan, C. , Dardick, C. , Scorza, R. , Goldman, I.L. and Ortiz, R. (2016) Rapid cycle breeding: application of transgenic early flowering for perennial trees. In Plant Breeding Reviews ( Janick, J. , ed), pp. 299–334. USA: John Wiley & Sons Inc.. [Google Scholar]
- CAOBISCO/ECA/FCC (2015) Cocoa beans: chocolate and cocoa industry quality requirements: ECA‐Caobisco‐FCC Cocoa.
- Cervantes‐Martinez, C. , Brown, J.S. , Schnell, R.J. , Phillips‐Mora, W. , Takrama, J.F. and Motamayor, J.C. (2006) Combining ability for disease resistance, yield, and horticultural traits of cacao (Theobroma cacao L.) clones. J. Am. Soc. Hort. Sci. 131, 231–241. [Google Scholar]
- Chingandu, N. , Kouakou, K. , Aka, R. , Gutierrez, O. and Brown, J. (2017a) Unexpected genome variability at multiple loci suggests Cacao swollen shoot virus comprises multiple, divergent molecular variants. J. Emerg. Virol. Dis. 3, 10.16966/2473-1846.128. [DOI] [Google Scholar]
- Chingandu, N. , Sreenivasan, T.N. , Surujdeo‐Maharaj, S. , Umaharan, P. , Gutierrez, O.A. and Brown, J.K. (2017b) Molecular characterization of previously elusive badnaviruses associated with symptomatic cacao in the New World. Archiv. Virol. 162, 1363–1371. [DOI] [PubMed] [Google Scholar]
- Clement, D. , Risterucci, A.‐M. , Motamayor, J.C. , N'goran, J. and Lanaud, C. (2003a) Mapping QTL for yield components, vigor, and resistance to Phytophthora palmivora in Theobroma cacao L. Genome, 46, 204–212. [DOI] [PubMed] [Google Scholar]
- Clement, D. , Risterucci, A.‐M. , Motamayor, J.C. , N'Goran, J. and Lanaud, C. (2003b) Mapping quantitative trait loci for bean traits and ovule number in Theobroma cacao L. Genome, 46, 103–111. [DOI] [PubMed] [Google Scholar]
- Clément, D. , Risterucci, A.‐M. and Lanaud, C. (2001) Analysis of QTL studies related to yield and vigour traits carried out with different cocoa genotypes. In: International workshop on new technologies and cocoa breeding pp. 127–134. Malaysia: INGENIC. [Google Scholar]
- Cocoa of Excellence (2017) Elements of a harmonized international standard for cocoa quality and flavour assessment. https://static1.squarespace.com/static/56680247841abadb3a819e1c/t/59bd4e6190bade6be5c42243/1505578602122/CoEx-Quality-Flavour-Standards-ENGLISH-11Sept2017.pdf.
- Cosme, S. , Cuevas, H. , Zhang, D. , Oleksyk, T. and Irish, B. (2016) Genetic diversity of naturalized cacao (Theobroma cacao L.) in Puerto Rico. Tree Genet. Genomes, 12, 88. [Google Scholar]
- Crouzillat, D. , Ménard, B. , Mora, A. , Phillips, W. and Pétiard, V. (2000) Quantitative trait analysis in Theobroma cacao using molecular markers. Euphytica, 114, 13–23. [Google Scholar]
- Darkwah, S.A. and Verter, N. (2014) An empirical analysis of cocoa bean production in Ghana. Eur. Scient. J. 10, 295–306. [Google Scholar]
- De Vuyst, L. and Weckx, S. (2016) The cocoa bean fermentation process: from ecosystem analysis to starter culture development. J. Appl. Microbiol. 121, 5–17. [DOI] [PubMed] [Google Scholar]
- Dinarti, D. , Susilo, A.W. , Meinhardt, L.W. , Ji, K. , Motilal, L.A. , Mischke, S. and Zhang, D. (2015) Genetic diversity and parentage in farmer selections of cacao from Southern Sulawesi, Indonesia revealed by microsatellite markers. Breed. Sci. 65, 438–446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dorenkott, M.R. , Griffin, L.E. , Goodrich, K.M. , Thompson‐Witrick, K.A. , Fundaro, G. , Ye, L. , Stevens, J.R. et al. (2014) Oligomeric cocoa procyanidins possess enhanced bioactivity compared to monomeric and polymeric cocoa procyanidins for preventing the development of obesity, insulin resistance, and impaired glucose tolerance during high‐fat feeding. J. Agric. Food Chem. 62, 2216–2227. [DOI] [PubMed] [Google Scholar]
- Dunwell, J.M. (2010) Haploids in flowering plants: origins and exploitation. Plant Biotechnol. J. 8, 377–424. [DOI] [PubMed] [Google Scholar]
- Dunwell, J.M. (2012) Patents for plants: context and current status. Acta Hortic. 941, 125–138. [Google Scholar]
- Dunwell, J.M. (2014) Genetically modified (GM) crops: European and transatlantic divisions. Mol. Plant Pathol. 15, 119–121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Earth Security Group (2015) The Earth Security Index 2015. Managing Global Resource Risks and Resilience in the 21st Century, pp. 54. London: Earth Security Group. [Google Scholar]
- Efombagn, M.I.B. , Sounigo, O. , Courtois, B. , Fouet, O. , Jeanneau, M. , Lemainque, A. , Pavek, S. et al. (2016) Association mapping for Phytophthora pod rot resistance in a cacao (Theobroma cacao L.) population grown in farmers’ field. Int. J. Agr. Biosci. 5, 329–336. [Google Scholar]
- Evina, V.J.E. , De Taeye, C. , Niemenak, N. , Youmbi, E. and Collin, S. (2016) Influence of acetic and lactic acids on cocoa flavan‐3‐ol degradation through fermentation‐like incubations. LWT‐Food Sci. Technol. 68, 514–522. [Google Scholar]
- Fair Labor Association (2012) Sustainable management of Nestlé's cocoa supply chain in the Ivory Coast—focus on labor standards.
- Falcao, L.L. , Silva‐Werneck, J.O. , Ramos, A.D.R. , Martins, N.F. , Bresso, E. , Rodrigues, M.A. , Bemquerer, M.P. et al. (2016) Antimicrobial properties of two novel peptides derived from Theobroma cacao osmotin. Peptides, 79, 75–82. [DOI] [PubMed] [Google Scholar]
- Faleiro, F.G. , Queiroz, V.T. , Lopes, U.V. , Guimarães, C.T. , Pires, J.L. , Yamada, M.M. , Araújo, I.S. et al. (2006) Mapping QTLs for witches’ broom (Crinipellis perniciosa) resistance in cacao (Theobroma cacao L.). Euphytica, 149, 227–235. [Google Scholar]
- Fang, W. , Meinhardt, L.W. , Mischke, S. , Bellato, C.M. , Motilal, L. and Zhang, D. (2014) Accurate determination of genetic identity for a single cacao bean, using molecular markers with a nanofluidic system, ensures cocoa authentication. J. Agric. Food Chem. 62, 481–487. [DOI] [PubMed] [Google Scholar]
- Figueira, A. and Janick, J. (1993) Development of nucellar somatic embryos of Theobroma cacao . Acta Hortic. 336, 231–238. [Google Scholar]
- Fister, A.S. , Mejia, L.C. , Zhang, Y. , Herre, E.A. , Maximova, S.N. and Guiltinan, M.J. (2016a) Theobroma cacao L. pathogenesis‐related gene tandem array members show diverse expression dynamics in response to pathogen colonization. BMC Genom. 17, 363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fister, A.S. , Shi, Z. , Zhang, Y. , Helliwell, E.E. , Maximova, S.N. and Guiltinan, M.J. (2016b) Protocol: transient expression system for functional genomics in the tropical tree Theobroma cacao L. Plant Methods, 12, 19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flament, M.‐H. , Kebe, I. , Clément, D. , Pieretti, I. , Risterucci, A.‐M. , N'Goran, J.‐A. , Cilas, C. et al. (2001) Genetic mapping of resistance factors to Phytophthora palmivora in cocoa. Genome, 44, 79–85. [DOI] [PubMed] [Google Scholar]
- Florez, S.L. , Erwin, R.L. , Maximova, S.N. , Guiltinan, M.J. and Curtis, W.R. (2015) Enhanced somatic embryogenesis in Theobroma cacao using the homologous BABY BOOM transcription factor. BMC Plant Biol. 15, 1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fouet, O. , Allegre, M. , Argout, X. , Jeanneau, M. , Lemainque, A. , Pavek, S. , Boland, A. et al. (2011) Structural characterization and mapping of functional EST‐SSR markers in Theobroma cacao . Tree Genet. Genomes, 7, 799–817. [Google Scholar]
- Freire, L. , Santana, J.O. , de Sousa, A.O. , dos Santos, J.B. , de Oliveira, I.B. , Alvim, F.C. , Gramacho, K.P. et al. (2017) TcPHYLL, a cacao phylloplanin expressed in young tissues and glandular trichomes. Physiol. Mol. Plant Pathol.. 100, 126–135 (in press) 10.1016/j.pmpp.2017.1006.1002. [DOI] [Google Scholar]
- Friscina, A. , Chiappetta, L. , Jacquemond, M. and Tepfer, M. (2017) Infection of non‐host model plant species with the narrow‐host‐range Cacao swollen shoot virus. Mol. Plant Pathol. 18, 293–297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guillou, C. , Fillodeau, A. , Brulard, E. , Verdier, D. , Simon, M. , Landmann, A. , Fontanel, A. et al. (2014) Nestlé Cocoa plan: cocoa propagation by somatic embryogenesis. In: The third international conference of the IUFRO unit 2.09. 02: Somatic embryogenesis and other vegetative propagation technologies p. 75. Vitoria‐Gasteiz, Spain.
- Guiltinan, M.J. , Verica, J. , Zhang, D. and Figueira, A. (2008) Genomics of Theobroma cacao, “the Food of the Gods”. In Genomics of Tropical Crop Plants ( Moore, P.H. and Ming, R. , eds), pp. 145–170. Heidelberg: Springer. [Google Scholar]
- Gutiérrez‐López, N. , Ovando‐Medina, I. , Salvador‐Figueroa, M. , Molina‐Freaner, F. , Avendaño‐Arrazate, C.H. and Vázquez‐Ovando, A. (2016) Unique haplotypes of cacao trees as revealed by trnH‐psbA chloroplast DNA. PeerJ, 4, e1855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamdouche, Y. , Guehi, T. , Durand, N. , Kedjebo, K.B.D. , Montet, D. and Meile, J.C. (2015) Dynamics of microbial ecology during cocoa fermentation and drying: towards the identification of molecular markers. Food Control, 48, 117–122. [Google Scholar]
- Hecht, V. , Vielle‐Calzada, J.‐P. , Hartog, M.V. , Schmidt, E.D. , Boutilier, K. , Grossniklaus, U. and de Vries, S.C. (2001) The Arabidopsis SOMATIC EMBRYOGENESIS RECEPTOR KINASE 1 gene is expressed in developing ovules and embryos and enhances embryogenic competence in culture. Plant Physiol. 127, 803–816. [PMC free article] [PubMed] [Google Scholar]
- Herrbach, E. , Le Maguet, J. and Hommay, G. (2016) Virus transmission by mealybugs and soft scales (Hemiptera, Coccoidea). In Vector‐Mediated Transmission of Plant Pathogens ( Brown, J.K. , ed), pp. 147–161. USA: The American Phytopathological Society. [Google Scholar]
- Ikeda, M. , Umehara, M. and Kamada, H. (2006) Embryogenesis‐related genes; its expression and roles during somatic and zygotic embryogenesis in carrot and Arabidopsis . Plant Biotechnol. 23, 153–161. [Google Scholar]
- Illeghems, K. , Weckx, S. and De Vuyst, L. (2015) Applying meta‐pathway analyses through metagenomics to identify the functional properties of the major bacterial communities of a single spontaneous cocoa bean fermentation process sample. Food Microbiol. 50, 54–63. [DOI] [PubMed] [Google Scholar]
- International Cocoa Organization (2017) Cocoa Market Review. International Cocoa Organization: Côte d'Ivoire. [Google Scholar]
- Issali, A.E. , Traoré, A. , Ngoran, J.A.K. , Koffi, K. and Sangaré, A. (2008) Relationship between some phenological parameters and somatic embryogenesis in Theobroma cacao L. J. Crop Sci. Biotechnol. 11, 23–30. [Google Scholar]
- Jégourel, Y. (2016) A chequered African history of commodity markets. Part II: cocoa. In Policy Brief, pp. 5. Morocco: PB‐16/07.OCP Policy Center. [Google Scholar]
- Kadow, D. , Bohlmann, J. , Phillips, W. and Lieberei, R. (2013) Identification of main fine flavour components in two genotypes of the cocoa tree (Theobroma cacao L.). J. Appl. Bot. Food Qual. 86, 90–98. [Google Scholar]
- Kadow, D. , Niemenak, N. , Rohn, S. and Lieberei, R. (2015) Fermentation‐like incubation of cocoa seeds (Theobroma cacao L.) – reconstruction and guidance of the fermentation process. LWT‐Food Sci. Technol. 62, 357–361. [Google Scholar]
- Karami, O. , Aghavaisi, B. and Pour, A.M. (2009) Molecular aspects of somatic‐to‐embryogenic transition in plants. J. Chem. Biol. 2, 177–190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keen, C.L. , Holt, R.R. , Oteiza, P.I. , Fraga, C.G. and Schmitz, H.H. (2005) Cocoa antioxidants and cardiovascular health. Am. J. Clin. Nutr. 81, 298S–303S. [DOI] [PubMed] [Google Scholar]
- Konar, N. , Toker, O.S. , Oba, S. and Sagdic, O. (2016) Improving functionality of chocolate: a review on probiotic, prebiotic, and/or synbiotic characteristics. Trends Food Sci. Tech. 49, 35–44. [Google Scholar]
- Kongor, J.E. , Hinneh, M. , Van de Walle, D. , Afoakwa, E.O. , Boeckx, P. and Dewettinck, K. (2016) Factors influencing quality variation in cocoa (Theobroma cacao) bean flavour profile ‐ a review. Food Res. Int. 82, 44–52. [Google Scholar]
- Kouassi, M.K. , Kahia, J. , Kouame, C.N.G. , Tahi, M.G. and Koffi, E.K. (2017) Comparing the effect of plant growth regulators on callus and somatic embryogenesis induction in four elite Theobroma cacao L. genotypes. HortScience, 52, 142–145. [Google Scholar]
- Krähmer, A. , Engel, A. , Kadow, D. , Ali, N. , Umaharan, P. , Kroh, L.W. and Schulz, H. (2015) Fast and neat – determination of biochemical quality parameters in cocoa using near infrared spectroscopy. Food Chem. 181, 152–159. [DOI] [PubMed] [Google Scholar]
- Kuhn, D.N. , Livingstone, D. , Main, D. , Zheng, P. , Saski, C. , Feltus, F.A. , Mockaitis, K. et al. (2012) Identification and mapping of conserved ortholog set (COS) II sequences of cacao and their conversion to SNP markers for marker‐assisted selection in Theobroma cacao and comparative genomics studies. Tree Genet. Genomes, 8, 97–111. [Google Scholar]
- Läderach, P. , Martinez‐Valle, A. , Schroth, G. and Castro, N. (2013) Predicting the future climatic suitability for cocoa farming of the world's leading producer countries, Ghana and Côte d'Ivoire. Clim. Change. 119, 841–854. [Google Scholar]
- Laliberte, B. and End, M. (2015) Supplying New Cocoa Planting Material to Farmers: A Review of Propagation Methodologies. Rome, Italy: Bioversity International. [Google Scholar]
- Lanaud, C. , Risterucci, A.M. , N'goran, A.K. , Clement, D. , Flament, M.H. , Laurent, V. and Falque, M. (1995) A genetic linkage map of Theobroma cacao L. Theor. Appl. Genet. 91, 987–993. [DOI] [PubMed] [Google Scholar]
- Lanaud, C. , Risterucci, A.‐M. , Pieretti, I. , Falque, M. , Bouet, A. and Lagoda, P. (1999) Isolation and characterization of microsatellites in Theobroma cacao L. Mol. Ecol. 8, 2141–2143. [DOI] [PubMed] [Google Scholar]
- Lanaud, C. , Risterucci, A.M. , Pieretti, I. , N'goran, J.A. and Fargeas, D. (2004) Characterisation and genetic mapping of resistance and defence gene analogs in cocoa (Theobroma cacao L.). Mol. Breed. 13, 211–227. [Google Scholar]
- Lanaud, C. , Fouet, O. , Clément, D. , Boccara, M. , Risterucci, A.‐M. , Surujdeo‐Maharaj, S. , Legavre, T. et al. (2009) A meta–QTL analysis of disease resistance traits of Theobroma cacao L. Mol. Breed. 24, 361–374. [Google Scholar]
- Laurent, V. , Risterucci, A.‐M. and Lanaud, C. (1994) Genetic diversity in cocoa revealed by cDNA probes. Theoret. Appl. Genet. 88, 193–198. [DOI] [PubMed] [Google Scholar]
- Lee, K.W. , Kim, Y.J. , Lee, H.J. and Lee, C.Y. (2003) Cocoa has more phenolic phytochemicals and a higher antioxidant capacity than teas and red wine. J. Agric. Food Chem. 51, 7292–7295. [DOI] [PubMed] [Google Scholar]
- Lee, A.H. , Steger, D. , Neilson, A.P. , O'keefe, S.F. , Ogejo, J.A. and Stewart, A.C. (2016) Model fermentation of cocoa (Theobroma cacao) can produce similar microbial and chemical quality of cocoa compared to conventional on‐farm cocoa fermentation. FASEB J. 30, lb353. [Google Scholar]
- Legavre, T. , Ducamp, M. , Sabau, X. , Argout, X. , Fouet, O. , Dedieu, F. , Surujdeo‐Maharaj, S. et al. (2015) Identification of Theobroma cacao genes differentially expressed during Phytophthora megakarya infection. Physiol. Mol. Plant Pathol. 92, 1–13. [Google Scholar]
- Li, Z. , Traore, A. , Maximova, S. and Guiltinan, M.J. (1998) Somatic embryogenesis and plant regeneration from floral explants of cacao (Theobroma cacao L.) using thidiazuron. In Vitro Cell. Dev. Biol.‐Plant, 34, 293–299. [Google Scholar]
- Liu, Y. , Shi, Z. , Maximova, S.N. , Payne, M.J. and Guiltinan, M.J. (2015) Tc‐MYBPA is an Arabidopsis TT2‐like transcription factor and functions in the regulation of proanthocyanidin synthesis in Theobroma cacao . BMC Plant Biol. 15, 160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Livingstone, D.S. , Freeman, B. , Motamayor, J.C. , Schnell, R.J. , Royaert, S. , Takrama, J. , Meerow, A.W. et al. (2012) Optimization of a SNP assay for genotyping Theobroma cacao under field conditions. Mol. Breed. 30, 33–52. [Google Scholar]
- Livingstone, D. , Royaert, S. , Stack, C. , Mockaitis, K. , May, G. , Farmer, A. , Saski, C. et al. (2015) Making a chocolate chip: development and evaluation of a 6K SNP array for Theobroma cacao . DNA Res. 22, 279–291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopes, U.V. , Monteiro, W.R. , Pires, J.L. , Clement, D. , Yamada, M.M. and Gramacho, K.P. (2011) Cacao breeding in Bahia, Brazil: strategies and results. Crop Breed. Appl. Biotechnol. 11, 73–81. [Google Scholar]
- López, C.M.R. , Bravo, H.S. , Wetten, A.C. and Wilkinson, M.J. (2010a) Detection of somaclonal variation during cocoa somatic embryogenesis characterised using cleaved amplified polymorphic sequence and the new freeware Artbio. Mol. Breeding, 25, 501–516. [Google Scholar]
- López, C.M.R. , Wetten, A.C. and Wilkinson, M.J. (2010b) Progressive erosion of genetic and epigenetic variation in callus‐derived cocoa (Theobroma cacao) plants. New Phytol. 186, 856–868. [DOI] [PubMed] [Google Scholar]
- Lotan, T. , Ohto, M.‐A. , Yee, K.M. , West, M.A. , Lo, R. , Kwong, R.W. , Yamagishi, K. et al. (1998) Arabidopsis LEAFY COTYLEDON1 is sufficient to induce embryo development in vegetative cells. Cell, 93, 1195–1205. [DOI] [PubMed] [Google Scholar]
- Loureiro, G.A. , Araujo, Q.R. , Sodré, G.A. , Valle, R.R. , Souza, J.O. Jr , Ramos, E.M. , Comerford, N.B. et al. (2017) Cacao quality: highlighting selected attributes. Food Rev. Int. 33, 382–405. [Google Scholar]
- Lowe, K. , Wu, E. , Wang, N. , Hoerster, G. , Hastings, C. , Cho, M.‐J. , Scelonge, C. et al. (2016) Morphogenic regulators baby boom and wuschel improve monocot transformation. Plant Cell, 28, 1998–2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maora, J. , Liew, E. and Guest, D. (2017) Limited morphological, physiological and genetic diversity of Phytophthora palmivora from cocoa in Papua New Guinea. Plant. Pathol. 66, 124–130. [Google Scholar]
- Marcano, M. , Pugh, T. , Cros, E. , Morales, S. , Páez, E.A.P. , Courtois, B. , Glaszmann, J.C. et al. (2007) Adding value to cocoa (Theobroma cacao L.) germplasm information with domestication history and admixture mapping. Theor. Appl. Genet. 114, 877–884. [DOI] [PubMed] [Google Scholar]
- Martin, M.A. , Goya, L. and Ramos, S. (2013) Potential for preventive effects of cocoa and cocoa polyphenols in cancer. Food Chem. Toxicol. 56, 336–351. [DOI] [PubMed] [Google Scholar]
- Martínez, I.B. , de la Cruz, M.V. , Nelson, M.R. and Bertin, P. (2017) Establishment of a core collection of traditional Cuban Theobroma cacao plants for conservation and utilization purposes. Plant Mol. Biol. Rep. 35, 47–60. [Google Scholar]
- Maximova, S. and Guiltinan, M.J. (2015) Tissue Culture. In Supplying New Cocoa Planting Material to Farmers: A Review of Propagation Methodologies ( Laliberté, B. and End, M. , eds), pp. 67–87. Rome, Italy: Bioversity International. [Google Scholar]
- Maximova, S.N. , Alemanno, L. , Young, A. , Ferriere, N. , Traore, A. and Guiltinan, M.J. (2002) Efficiency, genotypic variability, and cellular origin of primary and secondary somatic embryogenesis of Theobroma cacao L. In Vitro Cell. Dev. Biol.‐Plant, 8, 252–259. [Google Scholar]
- Maximova, S. , Miller, C. , De Mayolo, G.A. , Pishak, S. , Young, A. and Guiltinan, M.J. (2003) Stable transformation of Theobroma cacao L. and influence of matrix attachment regions on GFP expression. Plant Cell Rep. 21, 872–883. [DOI] [PubMed] [Google Scholar]
- Maximova, S.N. , Marelli, J.P. , Young, A. , Pishak, S. , Verica, J.A. and Guiltinan, M.J. (2006) Over‐expression of a cacao class I chitinase gene in Theobroma cacao L. enhances resistance against the pathogen, Colletotrichum gloeosporioides . Planta, 224, 740–749. [DOI] [PubMed] [Google Scholar]
- Maximova, S.N. , Young, A. , Pishak, S. and Guiltinan, M.J. (2008) Field performance of Theobroma cacao L. plants propagated via somatic embryogenesis. In Vitro Cell. Dev. Biol.‐Plant, 44, 487–493. [Google Scholar]
- Maximova, S.N. , Florez, S. , Shen, X. , Niemenak, N. , Zhang, Y. , Curtis, W. and Guiltinan, M.J. (2014) Genome‐wide analysis reveals divergent patterns of gene expression during zygotic and somatic embryo maturation of Theobroma cacao L., the chocolate tree. BMC Plant Biol. 14, 185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Melo Pereira, G.V. , Soccol, V.T. and Soccol, C.R. (2016) Current state of research on cocoa and coffee fermentations. Curr. Opin. Food Sci. 7, 50–57. [Google Scholar]
- Miguel, M.G.C.P. , de Castro Reis, L.V. , Efraim, P. , Santos, C. , Lima, N. and Schwan, R.F. (2017) Cocoa fermentation: microbial identification by MALDI‐TOF MS, and sensory evaluation of produced chocolate. LWT Food Sci. Technol. 77, 362–369. [Google Scholar]
- Miguelez‐Sierra, Y. , Hernández‐Rodríguez, A. , Acebo‐Guerrero, Y. , Baucher, M. and El Jaziri, M. (2017) In vitro micrografting of apical and axillary buds of cacao. J. Hortic. Sci. Biotechnol. 92, 25–30. [Google Scholar]
- Minyaka, E. , Niemenak, N. , Sangare, A. and Omokolo, D.N. (2008) Effect of MgSO4 and K2SO4 on somatic embryo differentiation in Theobroma cacao L. Plant Cell Tiss. Organ Cult. 94, 149. [Google Scholar]
- Modeste, K.K. , Eliane, M.T. , Daouda, K. , Brahima, S.A. , Tchoa, K. , Kouablan, K.E. and Mongomaké, K. (2017) Effect of antioxidants on the callus induction and the development of somatic embryogenesis of cocoa [‘Theobroma cacao’(L.)]. Austral. J. Crop Sci. 11, 25–31. [Google Scholar]
- Motamayor, J.C. , Risterucci, A.M. , Lopez, P.A. , Ortiz, C.F. , Moreno, A. and Lanaud, C. (2002) Cacao domestication I: the origin of the cacao cultivated by the Mayas. Heredity, 89, 380–386. [DOI] [PubMed] [Google Scholar]
- Motamayor, J.C. , Mockaitis, K. , Schmutz, J. , Haiminen, N. , Livingstone, D. 3rd , Cornejo, O. , Findley, S.D. et al. (2013) The genome sequence of the most widely cultivated cacao type and its use to identify candidate genes regulating pod color. Genome Biol. 14, r53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Motilal, L. and Butler, D. (2003) Verification of identities in global cacao germplasm collections. Genet. Resour. Crop Evol. 50, 799–807. [Google Scholar]
- Motilal, L.A. , Zhang, D. , Umaharan, P. , Boccara, M. , Mischke, S. , Sankar, A. and Meinhardt, L.W. (2012) Elucidation of genetic identity and population structure of cacao germplasm within an international cacao genebank. Plant Genet. Resour. 10, 232–241. [Google Scholar]
- Motilal, L.A. , Zhang, D. , Mischke, S. , Meinhardt, L.W. and Umaharan, P. (2013) Microsatellite‐aided detection of genetic redundancy improves management of the International Cocoa Genebank, Trinidad. Tree Genet. Genomes, 9, 1395–1411. [Google Scholar]
- Motilal, L.A. , Zhang, D. , Mischke, S. , Meinhardt, L.W. , Boccara, M. , Fouet, O. , Lanaud, C. et al. (2016) Association mapping of seed and disease resistance traits in Theobroma cacao L. Planta, 244, 1265–1276. [DOI] [PubMed] [Google Scholar]
- Muller, E. (2016) Cacao Swollen Shoot Virus (CSSV): history, biology, and genome. In Cacao Diseases ( Bailey, B.A. and Meinhardt, L.W. , eds), pp. 337–358. Switzerland: Springer International Publishing. [Google Scholar]
- Murphy, K.J. , Chronopoulos, A.K. , Singh, I. , Francis, M.A. , Moriarty, H. , Pike, M.J. , Turner, A.H. et al. (2003) Dietary flavanols and procyanidin oligomers from cocoa (Theobroma cacao) inhibit platelet function. Am. J. Clin. Nutr. 77, 1466–1473. [DOI] [PubMed] [Google Scholar]
- Niemenak, N. , Saare‐Surminski, K. , Rohsius, C. , Ndoumou, D.O. and Lieberei, R. (2008) Regeneration of somatic embryos in Theobroma cacao L. in temporary immersion bioreactor and analyses of free amino acids in different tissues. Plant Cell Rep. 27, 667–676. [DOI] [PubMed] [Google Scholar]
- Niemenak, N. , Awah, T.M. and Lieberei, R. (2012) Establishment of suspension culture in Theobroma cacao and polyamines associated with cacao embryogenesis. Plant Growth Regul. 67, 1–8. [Google Scholar]
- Niemenak, N. , Kaiser, E. , Maximova, S.N. , Laremore, T. and Guiltinan, M.J. (2015) Proteome analysis during pod, zygotic and somatic embryo maturation of Theobroma cacao . J. Plant Physiol. 180, 49–60. [DOI] [PubMed] [Google Scholar]
- Noah, A.M. , Niemenak, N. , Sunderhaus, S. , Haase, C. , Omokolo, D.N. , Winkelmann, T. and Braun, H.P. (2013) Comparative proteomic analysis of early somatic and zygotic embryogenesis in Theobroma cacao L. J. Proteomics, 78, 123–133. [DOI] [PubMed] [Google Scholar]
- de Oliveira Santos, M. , Romano, E. , Yotoko, K.S.C. , Tinoco, M.L.P. , Dias, B.B.A. and Aragao, F.J.L. (2005) Characterisation of the cacao somatic embryogenesis receptor‐like kinase (SERK) gene expressed during somatic embryogenesis. Plant Sci. 168, 723–729. [Google Scholar]
- Oyekale, A. , Bolaji, M. and Olowa, O. (2009) The effects of climate change on cocoa production and vulnerability assessment in Nigeria. Agric. J. 4, 77–85. [Google Scholar]
- Padi, F.K. , Ofori, A. , Takrama, J. , Djan, E. , Opoku, S.Y. , Dadzie, A.M. , Bhattacharjee, R. et al. (2015) The impact of SNP fingerprinting and parentage analysis on the effectiveness of variety recommendations in cacao. Tree Genet. Genomes, 11, 1–14. [Google Scholar]
- Padi, F.K. , Ofori, A. and Arthur, A. (2017) Genetic variation and combining abilities for vigour and yield in a recurrent selection programme for cacao. J. Agr. Sci. 155, 444–464. [Google Scholar]
- Pence, V.C. , Hasegawa, P.M. and Janick, J. (1980) Initiation and development of asexual embryos of Theobroma cacao L. in vitro . Z. Pflanzenphysiol. 98, 1–14. [Google Scholar]
- Pereira, A.S. , De Almeida, A.‐A.F. , da Silva Branco, M.C. , Costa, M.G.C. and Ahnert, D. (2017) Combining ability, heritability and genotypic relations of different physiological traits in cacao hybrids. PLoS ONE, 12, e0178790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perez‐Nunez, M. , Souza, R. , Sáenz, L. , Chan, J. , Zuniga‐Aguilar, J. and Oropeza, C. (2009) Detection of a SERK‐like gene in coconut and analysis of its expression during the formation of embryogenic callus and somatic embryos. Plant Cell Rep. 28, 11–19. [DOI] [PubMed] [Google Scholar]
- Pipitone, L. (2016) Overview of cocoa supply and demand. In: ICCO Cocoa Market Outlook Conference September 2016. London: International Cocoa Organization (ICCO). https://www.icco.org/about-us/international-cocoa-agreements/cat_view/294-cocoa-market-outlook-conference-september-2016.html. [Google Scholar]
- Pridmore, R.D. , Crouzillat, D. , Walker, C. , Foley, S. , Zink, R. , Zwahlen, M.C. , Brussow, H. et al. (2000) Genomics, molecular genetics and the food industry. J. Biotechnol. 78, 251–258. [DOI] [PubMed] [Google Scholar]
- Pugh, T. , Fouet, O. , Risterucci, A.M. , Brottier, P. , Abouladze, M. , Deletrez, C. , Courtois, B. et al. (2004) A new cacao linkage map based on codominant markers: development and integration of 201 new microsatellite markers. Theor. Appl. Genet. 108, 1151–1161. [DOI] [PubMed] [Google Scholar]
- Qin, X.‐W. , Lai, J.‐X. , Tan, L.‐H. , Hao, C.‐Y. , Li, F.‐P. , He, S.‐Z. and Song, Y.‐H. (2017) Characterization of volatile compounds in Criollo, Forastero, and Trinitario cocoa seeds (Theobroma cacao L.) in China. Int. J. Food Propert. 20, 1–15. [Google Scholar]
- Quainoo, A. , Wetten, A. and Allainguillaume, J. (2008) The effectiveness of somatic embryogenesis in eliminating the cocoa swollen shoot virus from infected cocoa trees. J. Virol. Methods, 149, 91–96. [DOI] [PubMed] [Google Scholar]
- Queiroz, V. , Guimarães, C. , Anhert, D. , Schuster, I. , Daher, R. , Pereira, M. , Miranda, V. et al. (2003) Identification of a major QTL in cocoa (Theobroma cacao L.) associated with resistance to witches’ broom disease. Plant Breed. 122, 268–272. [Google Scholar]
- Quinga, L.A.P. , de Freitas Fraga, H.P. , Viera, L.N. and Guerra, M.P. (2017) Epigenetics of long‐term somatic embryogenesis in Theobroma cacao L.: DNA methylation and recovery of embryogenic potential. Plant Cell Tiss. Organ Cult. 131, 295–305. [Google Scholar]
- Ramiro‐Puig, E. and Castell, M. (2009) Cocoa: antioxidant and immunomodulator. Br. J. Nutr. 101, 931–940. [DOI] [PubMed] [Google Scholar]
- Risterucci, A. , Grivet, L. , N'goran, J. , Pieretti, I. , Flament, M. and Lanaud, C. (2000) A high‐density linkage map of Theobroma cacao L. Theor. Appl. Genet. 101, 948–955. [DOI] [PubMed] [Google Scholar]
- Risterucci, A.M. , Paulin, D. , Ducamp, M. , N'Goran, J.A. and Lanaud, C. (2003) Identification of QTLs related to cocoa resistance to three species of Phytophthora . Theor. Appl. Genet. 108, 168–174. [DOI] [PubMed] [Google Scholar]
- Rohsius, C. , Elwers, S. and Lieberei, R. (2010). Cocoa‐Atlas. German cocoa and chocolate foundation (2010 ed.)
- Royaert, S. , Phillips‐Mora, W. , Leal, A.M.A. , Cariaga, K. , Brown, J.S. , Kuhn, D.N. , Schnell, R.J. et al. (2011) Identification of marker‐trait associations for self‐compatibility in a segregating mapping population of Theobroma cacao L. Tree Genet. Genomes, 7, 1159–1168. [Google Scholar]
- Royaert, S. , Jansen, J. , da Silva, D.V. , de Jesus Branco, S.M. , Livingstone, D.S. 3rd , Mustiga, G. , Marelli, J.P. et al. (2016) Identification of candidate genes involved in Witches’ broom disease resistance in a segregating mapping population of Theobroma cacao L. in Brazil. BMC Genom. 17, 107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rusconi, M. and Conti, A. (2010) Theobroma cacao L., the Food of the Gods: a scientific approach beyond myths and claims. Pharmacol. Res. 61, 5–13. [DOI] [PubMed] [Google Scholar]
- Sain, S.L. , Oduro, K.K. and Furtek, D.B. (1994) Genetic transformation of cocoa leaf cells using Agrobacterium tumefaciens . Plant Cell Tiss. Organ Cult. 37, 243–251. [Google Scholar]
- Santana, J.O. , Freire, L. , de Sousa, A.O. , Fontes Soares, V.L. , Gramacho, K.P. and Pirovani, C.P. (2016) Characterization of the legumains encoded by the genome of Theobroma cacao L. Plant Physiol. Biochem. 98, 162–170. [DOI] [PubMed] [Google Scholar]
- Santos, E.S. , Cerqueira‐Silva, C.B. , Mori, G. , Ahnert, D. , Corrêa, R. and Souza, A. (2012a) New polymorphic microsatellite loci for Theobroma cacao: isolation and characterization of microsatellites from enriched genomic libraries. Biolog. Plant. 56, 789–792. [Google Scholar]
- Santos, R.M. , Lopes, U.V. , Silva, S.D. , Micheli, F. , Clement, D. and Gramacho, K.P. (2012b) Identification of quantitative trait loci linked to Ceratocystis wilt resistance in cacao. Mol. Breed. 30, 1563–1571. [Google Scholar]
- Santos, E.S. , Cerqueira‐Silva, C.B.M. , Mori, G.M. , Ahnert, D. , Mello, D.L. , Pires, J.L. , Corrêa, R.X. et al. (2015) Genetic structure and molecular diversity of cacao plants established as local varieties for more than two centuries: the genetic history of cacao plantations in Bahia, Brazil. PLoS ONE, 10, e0145276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- dos Santos, F.N. , Tata, A. , Belaz, K.R.A. , Magalhães, D.M.A. , Luz, E.D.M.N. and Eberlin, M.N. (2017) Major phytopathogens and strains from cocoa (Theobroma cacao L.) are differentiated by MALDI‐MS lipid and/or peptide/protein profiles. Anal. Bioanal. Chem. 409, 1765–1777. [DOI] [PubMed] [Google Scholar]
- Schmidt, E. , Guzzo, F. , Toonen, M. and De Vries, S. (1997) A leucine‐rich repeat containing receptor‐like kinase marks somatic plant cells competent to form embryos. Development, 124, 2049–2062. [DOI] [PubMed] [Google Scholar]
- Schnell, R.J. , Kuhn, D. , Brown, J. , Olano, C. , Phillips‐Mora, W. , Amores, F. and Motamayor, J. (2007) Development of a marker assisted selection program for cacao. Phytopathology, 97, 1664–1669. [DOI] [PubMed] [Google Scholar]
- Schroth, G. , Laderach, P. , Martinez‐Valle, A.I. , Bunn, C. and Jassogne, L. (2016) Vulnerability to climate change of cocoa in West Africa: patterns, opportunities and limits to adaptation. Sci. Total Environ. 556, 231–241. [DOI] [PubMed] [Google Scholar]
- Schwenninger, S.M. , Leischtfeld, S.F. and Gantenbein‐Demarchi, C. (2016) High‐throughput identification of the microbial biodiversity of cocoa bean fermentation by MALDI‐TOF MS. Lett. Appl. Microbiol. 63, 347–355. [DOI] [PubMed] [Google Scholar]
- Sena Gomes, A. , Andrade Sodré, G. , Guiltinan, M. , Lockwood, R. and Maximova, S. (2015) Supplying New Cocoa Planting Material to Farmers: A Review of Propagation Methodologies. Rome, Italy: Bioversity International. [Google Scholar]
- Sharma, S.K. , Millam, S. , Hein, I. and Bryan, G.J. (2008) Cloning and molecular characterisation of a potato SERK gene transcriptionally induced during initiation of somatic embryogenesis. Planta, 228, 319–330. [DOI] [PubMed] [Google Scholar]
- Shires, M.E. , Florez, S.L. , Lai, T.S. and Curtis, W.R. (2017) Inducible somatic embryogenesis in Theobroma cacao achieved using the DEX‐activatable transcription factor‐glucocorticoid receptor fusion. Biotechnol. Lett. 39, 1747–1755 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silva, T.E. , Cidade, L.C. , Alvim, F.C. , Cascardo, J.C. and Costa, M.G. (2009) Studies on genetic transformation of Theobroma cacao L.: evaluation of different polyamines and antibiotics on somatic embryogenesis and the efficiency of uidA gene transfer by Agrobacterium tumefaciens . Plant Cell Tiss. Organ Cult. 99, 287–298. [Google Scholar]
- da Silva, T.É.R. , Cidade, L.C. , Alvim, F.C. , Cascardo, J.C.D.M. and Costa, M.G.C. (2008) Somatic embryogenesis and plant regeneration in elite clones of Theobroma cacao . Pesq. Agropec. Bras. 43, 1433–1436. [Google Scholar]
- da Silva, M.R. , Clément, D. , Gramacho, K.P. , Monteiro, W.R. , Argout, X. , Lanaud, C. and Lopes, U. (2016) Genome‐wide association mapping of sexual incompatibility genes in cacao (Theobroma cacao L.). Tree Genet. Genomes, 12, 62. [Google Scholar]
- da Silva, R.A. , Souza, G. , Lemos, L.S.L. , Lopes, U.V. , Patrocínio, N.G.R.B. , Alves, R.M. , Marcellino, L.H. et al. (2017) Genome size, cytogenetic data and transferability of EST‐SSRs markers in wild and cultivated species of the genus Theobroma L. (Byttnerioideae, Malvaceae). PLoS ONE, 12, e0170799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Squicciarini, M.P. and Swinnen, J. (2016) The Economics of Chocolate. Oxford: Oxford University Press. [Google Scholar]
- Stack, J.C. , Royaert, S. , Gutiérrez, O. , Nagai, C. , Holanda, I.S.A. , Schnell, R. and Motamayor, J.‐C. (2015) Assessing microsatellite linkage disequilibrium in wild, cultivated, and mapping populations of Theobroma cacao L. and its impact on association mapping. Tree Genet. Genomes, 11, 19. [Google Scholar]
- Stone, S.L. , Kwong, L.W. , Yee, K.M. , Pelletier, J. , Lepiniec, L. , Fischer, R.L. , Goldberg, R.B. et al. (2001) LEAFY COTYLEDON2 encodes a B3 domain transcription factor that induces embryo development. Proc. Natl Acad. Sci. 98, 11806–11811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Talapatra, S. , Ghoshal, N. and Raychaudhuri, S.S. (2014) Molecular characterization, modeling and expression analysis of a somatic embryogenesis receptor kinase (SERK) gene in Momordica charantia L. during somatic embryogenesis. Plant Cell Tiss. Organ Cult. 116, 271–283. [Google Scholar]
- Tan, C. and Furtek, D. (2003) Development of an in vitro regeneration system for Theobroma cacao from mature tissues. Plant Sci. 164, 407–412. [Google Scholar]
- Teixeira, P.J.P.L. , Thomazella, D.P.T. and Pereira, G.A.G. (2015) Time for chocolate: current understanding and new perspectives on cacao Witches’ broom disease research. PLoS Pathog. 11, e1005130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tran, P.D. , Van de Walle, D. , De Clercq, N. , De Winne, A. , Kadow, D. , Lieberei, R. , Messens, K. et al. (2015) Assessing cocoa aroma quality by multiple analytical approaches. Food Res. Int. 77, 657–669. [Google Scholar]
- Traore, A. and Guiltinan, M.J. (2006) Effects of carbon source and explant type on somatic embryogenesis of four cacao genotypes. HortScience, 41, 753–758. [Google Scholar]
- Traore, A. , Maximova, S.N. and Guiltinan, M.J. (2003) Micropropagation of Theobroma cacao L. using somatic embryo‐derived plants. In Vitro Cell. Dev. Biol.‐Plant, 39, 332–337. [Google Scholar]
- Turnbull, C. , Butler, D. , Cryer, N. , Zhang, D. , Lanaud, C. , Daymond, A. , Ford, C. et al. (2004) Tackling mislabelling in cocoa germplasm collections. Ingenic Newsl. 9, 8–11. [Google Scholar]
- Wetten, A. , Campbell, C. and Allainguillaume, J. (2016) High‐resolution melt and morphological analyses of mealybugs (Hemiptera: Pseudococcidae) from cacao: tools for the control of Cacao swollen shoot virus spread. Pest Manag. Sci. 72, 527–533. [DOI] [PubMed] [Google Scholar]
- Wickramasuriya, A.M. and Dunwell, J.M. (2015) Global scale transcriptome analysis of Arabidopsis embryogenesis in vitro . BMC Genom. 16, 301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson, P.K. and Hurst, W.J. (2015) Chocolate and Health: Chemistry, Nutrition and Therapy. UK: Royal Society of Chemistry. [Google Scholar]
- Wood, G.A.R. and Lass, R.A. (1985) Cocoa. London: Longman. [Google Scholar]
- World Cocoa Foundation (2012) Cocoa market update. http://www.worldcocoafoundation.org/wp-content/uploads/Cocoa-Market-Update-as-of-3.20.2012.pdf.
- World Cocoa Foundation (2014) Cocoa market update. http://www.worldcocoafoundation.org/wp-content/uploads/Cocoa-Market-Update-as-of-4-1-2014.pdf.
- World Cocoa Foundation (2016) Leading cocoa and chocolate companies join together to help cocoa farmers adapt to weather and climate impacts. http://www.worldcocoafoundation.org/wp-content/uploads/FINAL-CSC-program-announcement-5-31.pdf.
- Xu, H. , Gao, Y. and Wang, J. (2012) Transcriptomic analysis of rice (Oryza sativa) developing embryos using the RNA‐Seq technique. PLoS ONE, 7, e30646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang, S. , Fresnedo‐Ramírez, J. , Wang, M. , Cote, L. , Schweitzer, P. , Barba, P. , Takacs, E.M. et al. (2016) A next‐generation marker genotyping platform (AmpSeq) in heterozygous crops: a case study for marker‐assisted selection in grapevine. Hortic. Res. 3, 16002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, D. and Motilal, L. (2016) Origin, dispersal, and current global distribution of cacao genetic diversity. In Cacao Diseases ( Bailey, B.A. and Meinhardt, L.W. , eds), pp. 3–31. Switzerland: Springer International Publishing. [Google Scholar]
- Zhang, Y. , Clemens, A. , Maximova, S.N. and Guiltinan, M.J. (2014) The Theobroma cacao B3 domain transcription factor TcLEC2 plays a duel role in control of embryo development and maturation. BMC Plant Biol. 14, 106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, Y. , Maximova, S.N. and Guiltinan, M.J. (2015) Characterization of a stearoyl‐acyl carrier protein desaturase gene family from chocolate tree, Theobroma cacao L. Front. Plant Sci. 6, 239. [DOI] [PMC free article] [PubMed] [Google Scholar]