

Abstract
Just like all matter, proteins can also switch between gas, liquid and solid phases. Protein phase transition has claimed the spotlight in recent years as a novel way of how cells compartmentalize and regulate biochemical reactions. Moreover, this discovery has provided a new framework for the study of membrane-less organelle biogenesis and protein aggregation in neurodegenerative disorders. We now argue that this framework could be useful in the study of cell cycle regulation and cancer. Based on our work on phase transitions of arginine-rich proteins in neurodegeneration, via combining mass spectroscopy with bioinformatics analyses, we found that also numerous proteins involved in the regulation of the cell cycle can undergo protein phase separation. Indeed, several proteins whose function affects the cell cycle or are associated with cancer, have been recently found to phase separate from the test tube to cells. Investigating the role of this process for cell cycle proteins and understanding its molecular underpinnings will provide pivotal insights into the biology of cell cycle progression and cancer.
Keywords: Protein phase separation, Oncogenic fusion, Protein aggregation, Cancer, Stress granules, Nucleolus, Centrosome
Background
Compartmentalization is a key feature of life. The cell membrane defines the context of what is living and what is not. Yet, this is only the first stage of the spatial organization of living matter. Cells, and in particularly eukaryotes, are further divided in subcompartments termed organelles, each of them carrying out specific biochemical reactions. These organelles can be roughly divided in two classes: membrane-bound and membrane-less. Whereas the processes behind the formation of membrane-bound vesicles have been relatively well-studied, the biogenesis and properties of their membrane-less counterparts remained elusive.
Membrane-less organelles (e.g. the nucleolus, stress granules, …) often consist of protein and RNA. Yet how does a cell concentrate these biomolecules without a membrane barrier? In recent years the phenomenon of liquid–liquid phase separation was found to underlie the biogenesis of these compartments [1–9]. Multivalent interactions predominantly mediated by intrinsically disordered and low complexity domains drive the spontaneous demixing of the RNA binding proteins involved [1–10]. This demixing results in the formation of liquid-like protein droplets or protein hydrogels, and depends on specific in vitro conditions, such as concentration, salt and temperature [1–9]. Interestingly, such test-tube phases are highly reminiscent of cellular RNA granules, suggesting that protein phase transition might indeed be the physical basis of the biogenesis of membrane-less organelles [1, 7, 8, 11]. However, these dynamic test tube assemblies seem to spontaneously mature to a more solid-like state [7, 8, 12–14], suggesting that they could act as stepping stones towards protein aggregation. Indeed, membrane-less organelles have been suspected to serve this role in some neurodegenerative disorders [15, 16].
For example, amyotrophic lateral sclerosis (ALS) is an adult-onset neurodegenerative disorder characterized by the aggregation of RNA binding proteins (RBPs) in the central nervous system [17]. Based on similarities in protein content between RBP aggregates and stress granules, these membrane-less organelles have been suggested as seeds for pathological aggregation of RBPs in patients [15, 16]. However, why these proteins undergo this liquid-to-solid switch during aging is unknown. Rare disease mutations found in some of these proteins make them more aggregation-prone [7, 8, 13], yet these cases do not explain why the wildtype proteins also aggregate in the majority of ALS cases. Hexanucleotide repeat expansions in the C9orf72 gene are the most common genetic cause of ALS [18, 19], and recent evidence points at unconventional dipeptide repeat (DPR) peptides derived from the expanded repeat RNA as a major pathogenic species in the disease [20–25]. While five different DPRs are formed, two arginine-rich ones (i.e. glycine–arginine and proline–arginine, or GR and PR), are highly toxic in disease models [20–25]. We and others have recently found that these arginine-rich DPRs can phase separate in the presence of RNA [25]. Additionally, these toxic peptides promote a liquid-to-solid switch of stress granules in cells [25, 26]. Hence, providing an explanation why the involved RBPs start to aggregate in the most common genetic form of the disease.
Main text
Proteins involved in cell cycle phase separate in vitro
We have extensively characterized our PR-RNA granule system and argue its usefulness as a test tube model for protein phase separation [26]. To identify cellular proteins prone to arginine-mediated phase separation, we performed mass spectrometry (MS) [27] (see Fig. 1). We incubated soluble HeLa cell lysate, cleared from the insoluble fraction, with PR peptide. This resulted instantaneously in phase separation of PR with cellular proteins, observed as a clouding of the sample. Through mild centrifugation we collected these phase separated proteins into a pellet, which we showed was dependent on both weak liquid-like and more stable solid-like interactions [27]. Such a stable core/liquid shell topology is also observed for membrane-less organelles in living cells [11]. We identified 874 proteins in our sample, which were enriched for RBPs and proteins involved in stress granule metabolism, hereby confirming our observations from cells where we found that PR targets and perturbs stress granules [27].
Fig. 1.
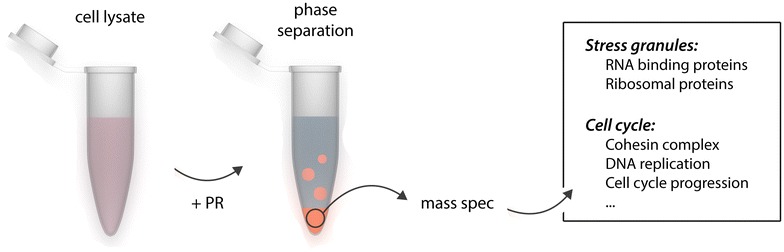
Identification of the phase separating proteome. Cleared cell lysate was incubated with poly-PR peptide to induce phase separation of cellular proteins. Phase separated proteins were precipitated by mild centrifugation and subjected to mass spectrometry. Identified proteins included stress granule factors and other membrane-less organelle components, but surprisingly as well proteins annotated as implicated in the regulation of the cell cycle
Interestingly though, further analysis in follow-up work showed that our PR dataset is also highly enriched for proteins involved in the regulation of the cell cycle (GO:0000278; fold enrichment = 3.83, p = 2.93E−23, Fisher Exact Bonferroni). In Table 1 we provide an overview of some cell cycle proteins (KEGG pathway: hsa04110) that we identified in our MS experiment. This finding suggests that phase separation could also play a role in the regulation of this process. A common feature of proteins that undergo phase separation is structural disorder [10]. Indeed, proteins involved in cell cycle regulation are on average more disordered (IUPred score; median fold change = 1.69, p < 1.00E−4, Mann–Whitney) compared to the proteome. Besides being mostly intrinsically disordered, phase separating proteins also often show low sequence complexity, as exemplified by prion-like domains [28] (rich in uncharged polar amino acids and glycine) and arginine-rich domains [29]. Again, cell cycle regulatory proteins are enriched for both prion-like domains (fold enrichment = 1.39, p = 4.90E−02, binomial test) and arginine-rich domains (≥ 6 R-motifs/protein; fold enrichment = 2.44, p = 9.33E−15, binomial test) compared to the proteome. Besides these typical protein characteristics, several proteins that are known to phase separate also affect the regulation of the cell cycle (Table 2). Additionally, proteins regulating or affecting (even indirectly) the cell cycle are enriched in several known membrane-less organelles (Fig. 2a). Interestingly, some of these organelles also have been shown to dynamically change over the course of the cell cycle, as exemplified by the nucleolus, purinosome and centrosome [30–32]. Given that we found numerous cell cycle proteins in our MS dataset, and based on their physical characteristics and targeting to membrane-less compartments, we argue that there could be a previously unappreciated role for protein phase transition in the regulation and execution of the cell cycle.
Table 1.
Cell cycle proteins identified in the mass spec experiment
| Gene name | Protein name | Role in cell cycle |
|---|---|---|
| CDC5L | Cell division cycle 5-like | Cell cycle progression |
| CDC37 | Cell division cycle 37 | Cell cycle progression |
| CDC42 | Cell division cycle 42 homolog | Kinetochore complex |
| CDC73 | Cell division cycle protein 73 homolog | Cell cycle progression |
| CDK11A | Cyclin-dependent kinase 11A | Cell cycle progression |
| CDK11B | Cyclin-dependent kinase 11B | Cell cycle progression |
| CUL4A | Cullin-4A | Cell cycle progression |
| CUL4B | Cullin-4B | Cell cycle progression |
| HDAC1 | Histone deacetylase 1 | Cell cycle progression |
| MAD1L1 | Mitotic spindle assembly checkpoint protein MAD1 | Spindle-assembly checkpoint |
| MCM2 | DNA replication licensing factor MCM2 | DNA replication initiation |
| MCM3 | DNA replication licensing factor MCM3 | DNA replication initiation |
| MCM4 | DNA replication licensing factor MCM4 | DNA replication initiation |
| MCM5 | DNA replication licensing factor MCM5 | DNA replication initiation |
| MCM6 | DNA replication licensing factor MCM6 | DNA replication initiation |
| MCM7 | DNA replication licensing factor MCM7 | DNA replication initiation |
| PCNA | Proliferating cell nuclear antigen | Control of DNA replication |
| PRKDC | DNA-dependent protein kinase catalytic subunit | Sensor for DNA damage |
| RAD21 | Double-strand-break repair protein rad21 homolog | Cohesin complex |
| SMC1A | Structural maintenance of chromosomes protein 1A | Cohesin complex |
| SMC2 | Structural maintenance of chromosomes protein 2 | Cohesin complex |
| SMC3 | Structural maintenance of chromosomes protein 3 | Cohesin complex |
| SMC4 | Structural maintenance of chromosomes protein 4 | Cohesin complex |
| YWHAB | 14-3-3 protein beta/alpha | Cell cycle progression |
| YWHAE | 14-3-3 protein epsilon | Cell cycle progression |
| YWHAG | 14-3-3 protein gamma | Cell cycle progression |
| YWHAH | 14-3-3 protein eta | Cell cycle progression |
| YWHAZ | 14-3-3 protein zeta/delta | Cell cycle progression |
Table 2.
Phase separated proteins whose function affects the cell cycle
| Protein name | Effect on cell cycle | Phase separation | |
|---|---|---|---|
| Nucleolus | |||
| FIBL | Knockdown reduces cell growth [57] | [12] | |
| NPM-1a | Role in tumorigenesis [58] | [12] | |
| Stress granule | |||
| eIF4G2 | Knockdown induces apoptosis and impairs proliferation [59] | [9] | |
| FUSa | Knockdown impairs cell proliferation [60] | [6, 8, 9, 51] | |
| EWSa | Knockdown induces apoptosis and impairs proliferation [61] | [2] | |
| hnRNPA1 | Knockdown induces cell cycle arrest [62] | [7, 9] | |
| TDP-43 | Overexpression induces cell cycle arrest [63] | [64] | |
| TIA-1 | Knockdown promotes cell proliferation [65] | [9] | |
| Centrosome | |||
| PCM-1 | Involved in cell cycle progression [66] | [67] | |
| Purinosome | |||
| PPAT | Regulates growth rate via de novo purine biosynthesis [68] | [69] | |
| Nuclear pore | |||
| Nup98a | Regulates expression cell cycle genes [70] | [71] | |
| T-cell receptor | |||
| LAT | Regulates T-cell activation and proliferation [72] | [40] | |
Fig. 2.
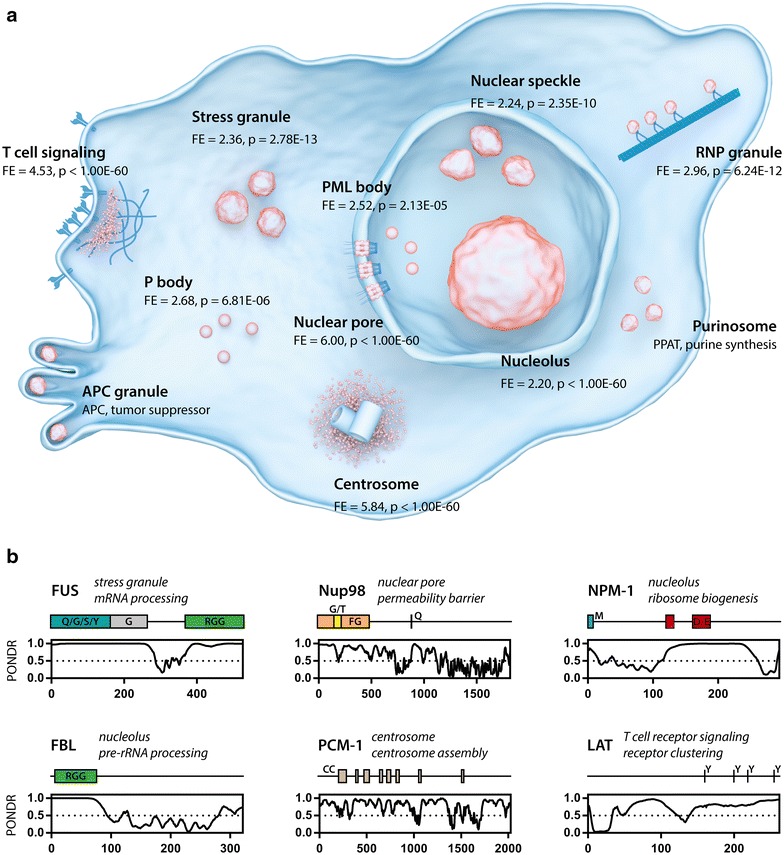
Proteins regulating or affecting the cell cycle are involved in cellular phase separations. a Overview of different membraneless organelles (orange). The fold enrichment of cell cycle proteins (GO:0000278) is shown for each organelle for which the protein content was available. T cell signaling (GO:0050852) [40], stress granule [11], nuclear speckle (GO:0016607), RNP granule (GO:0035770), PML body [73], P body (GO:0000932), nuclear pore (GO:0005643) [74], nucleolus [75], centrosome (GO:0005813) [76]. APC granules and purinosomes were positive for cell cycle proteins APC [77] and PPAT [78] respectively. b Examples of cell cycle proteins found in membrane-less organelles which can undergo phase separation (see Table 1). PONDR disorder prediction plots are shown, indicating prevalence of disordered regions in these proteins (score > 0.5). Coiled coil (CC) and low complexity domains (letters indicate overrepresented amino acids) are also indicated. Phosphotyrosine residues necessary for receptor clustering are indicated for LAT
How could membrane-less organelles be involved in the cell cycle and cancer?
We have shown that proteins involved in the regulation of the cell cycle are enriched for features commonly associated with phase separation, and many of these proteins are components of membrane-less organelles. This begs the question how membrane-less organelles could be functionally implicated in the regulation of the cell cycle? What purpose do they serve, and are they altered in cancer?
Compartmentalization has been known to serve different functions: (1) Catalyzing biochemical reactions by concentrating reaction compounds, (2) shielding components from each other by localization in different compartments, (3) storage of biomolecules for later use, and (4) signal amplification. First, the laws of chemistry dictate that at higher concentrations of the reaction components, the reaction efficiency will increase. It seems that exactly this is the function of the pericentriolar material. This membrane-less organelle concentrates tubulin monomers, which are subsequently efficiently nucleated and grown into microtubules [33]. Secondly, chromatin architecture is known to be linked to the cell cycle [34]. Recently, two groups reported that heterochromatin domains form also by a process of phase separation [35, 36], showing that this process is key in regulating chromatin architecture. Additionally, during mitosis a specific disordered protein associates with the compacted DNA and acts as a biological surfactant to prevent the condensed sister chromatids from sticking together [37]. Upon nuclear envelope formation however, another helical protein exactly does the opposite, by crosslinking the condensed DNA to ensure the formation of one nucleus [38]. These findings show that phase separation is key in the organization of DNA over the cell cycle. Thirdly, upon quiescence due to nutrient-limiting conditions the cell’s proteasomes will assemble in cytoplasmic proteasome storage granules. Upon reentry into the cell cycle, these granules disassemble and the proteasome complexes translocate back to the nucleus to carry out their function [39]. Lastly, protein phase separation can also promote signal amplification. One of the best examples illustrating this process is the phase separation of T-cell receptors upon stimulation. Their phase separated intracellular domains concentrate signaling molecules to generate a robust signaling response activating cellular differentiation programs [40].
Besides the regulatory functions of membrane-less organelles and protein phase separation in processes associated with the cell cycle, they are sometimes also altered in cancer. For example, different cancers have a reported increase in stress granules and paraspeckles. As the increased number of these membrane-less organelles has been linked to a poor prognosis for survival [41–43], it makes them an interesting therapeutic target. Additionally, aggregation of different tumor suppressor proteins, including p53, results in their loss of function and is a major mechanism in cancer [44]. Compounds preventing its aggregation have been successful in preclinical animal models [45], indicating that indeed protein phase transitions could be viable therapeutic options.
Phase separation is a novel mechanism of oncogenic fusion proteins
Interestingly, several of the disordered proteins prone to phase separation are also known to be involved in cancer-related fusion events (see Table 2). The most relevant examples are FUS and EWS, which are also components of stress granules and aggregate in ALS [17]. For example, FUS is involved in the chimera FUS-CHOP in liposarcomas [46]. EWS on the other hand is found as an EWS–FLI1 fusion causal for sarcomas and leukemias [47], and as an EWS–ATF1 fusion in melanomas [48]. Another notable example is nucleophosmin (NPM-1), a key component of the nucleolus, which is also part of the NPM-ALK fusion product in non-Hodgkin’s lymphoma [49]. Lastly the nuclear pore protein NUP98 is found in many oncogenic fusions involved in leukemias [50].
Several observations confirm that the correlation between phase separation and fusion proteins is more than coincidental. There are three essential features of proteins that prevail in both classes, which suggests a causative link between the two cellular processes. First, phase separating proteins have a high level of structural disorder [5, 51], which is also the case with oncogenic fusion proteins [52]. Second, phase separation relies on transient and multivalent protein–protein interactions [5], and the same principle holds true for oncogenic fusions. For example, activation of the oncogenic NPM-ALK chimera requires transient oligomerization mediated by the NPM segment [49], and coiled-coil interaction motifs are central to the autoactivation of many other oncogenic fusions [52]. Exactly, such protein–protein interaction domains are enriched in proteins undergoing phase separation (see Fig. 2b). Third, a recurrent feature of cellular phase separation is the presence of RNA and RNA-binding proteins [6, 16]. Again, oncogenic fusion proteins are also significantly enriched in RNA- and DNA-binding domains [52, 53].
It is believed that these disordered domains act as transcription activation domains, and hence drive gene expression where the fusion protein interacts with the DNA [51, 52]. Recent evidence has shown that the transcriptional activation potential of these disordered domains is directly correlated to their ability to phase separate. Kwon et al. [54] generated synthetic fusions of the FUS low complexity domain and fused it to a GAL4 DNA binding domain. By making point mutations which interfere with the hydrophobic interactions required for phase separation, the researchers not only perturbed hydrogel formation in the test tube, but also with transcriptional activity of the fusion protein in cells [54]. Compellingly, a new study found exactly the same mechanism of action in relevant EWS–FLI fusions observed in patients [55]. Also in this study there was a direct correlation between the potential to phase separate and to activate transcription by these disordered domains [55]. Indeed, several groups have shown that the C-terminal domain of RNA polymerase II has a strong affinity for phase separated disordered domains [51, 54, 56], illustrating how local phase separation can recruit the transcriptional machinery to distinct genomic regions and drive the oncogenic transformation of cells.
Conclusions
In the last few years, the concept of protein phase separation has taken the field of cell biology by storm. This physical phenomenon provides a clear framework for the understanding of membrane-less organelle biogenesis. Moreover, this insight has given us a new view on protein aggregation in the context of human disease, and more specifically in neurodegenerative disorders. In our previous experiments on the role of protein phase transition in the pathogenesis of ALS, we developed a test tube model for the study of this process. Using MS, this simple model allowed us to perform a proteome-wide search for proteins which could undergo phase separation. Unexpectedly, we identified numerous proteins which were directly involved in the cell cycle or its regulation. This finding strongly suggests that protein phase separation could be at play in cell cycle regulation and associated diseases such as cancer.
To test this hypothesis, we examined in this commentary the physical characteristics of proteins involved in cell cycle regulation, and combined this with new insights from recent studies in the phase transition field. Not only do proteins involved in cell cycle regulation have the right physical characteristics for phase separation, we do know they are actually enriched in cellular membrane-less organelles. Lastly, we suggest different mechanisms of how membrane-less organelle formation and related processes could be functionally involved in cell cycle regulation and misregulation in cancer. We postulate that there is increasing evidence for such a functional involvement which warrants further experiments to uncover its full extent. To conclude, we would like to argue that the framework of protein phase separation could be useful to the study of the cell cycle in health and disease, and may guide the development of novel therapeutic approaches.
Authors’ contributions
SB performed statistical analyses. SB, PT and LVDB wrote manuscript. All authors read and approved the final manuscript.
Acknowledgements
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
Mass spectrometry dataset was previously published and is the data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifier PRIDE: PXD005509.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Funding
Research was funded by the KU Leuven (C1), VIB, the European Research Council in the context of the European’s Seventh Framework Programme (FP7/2007-2013), the Research Foundation Flanders (FWO) G.0983.14N, the Interuniversity Attraction Poles Programme P7/16 initiated by the Belgian Science Policy Office, the Association Belge contre les Maladies Neuro-Musculaires (ABMM), the ALS Liga (Belgium) and the ‘Opening the Future’ Fund. S.B. received a Ph.D. fellowship from the Agency for Innovation by Science and Technology (IWT). P.T. was supported by the Odysseus Grant G.0029.12 from Research Foundation Flanders (FWO).
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- ALS
amyotrophic lateral sclerosis
- DPR
dipeptide repeats
- GR
glycine–arginine
- MS
mass spectrometry
- PR
proline–arginine
- RBP
RNA-binding protein
Contributor Information
Steven Boeynaems, Email: steven.boeynaems@vib-kuleuven.be.
Peter Tompa, Email: peter.tompa@vub.vib.be.
Ludo Van Den Bosch, Email: ludo.vandenbosch@kuleuven.vib.be.
References
- 1.Zhang HY, et al. RNA controls PolyQ protein phase transitions. Mol Cell. 2015;60:220–230. doi: 10.1016/j.molcel.2015.09.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Altmeyer M, et al. Liquid demixing of intrinsically disordered proteins is seeded by poly(ADP-ribose) Nat Commun. 2015;6:8088. doi: 10.1038/ncomms9088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Brangwynne CP, et al. Germline P granules are liquid droplets that localize by controlled dissolution/condensation. Science. 2009;324:1729–1732. doi: 10.1126/science.1172046. [DOI] [PubMed] [Google Scholar]
- 4.Brangwynne CP, Mitchison TJ, Hyman AA. Active liquid-like behavior of nucleoli determines their size and shape in Xenopus laevis oocytes. Proc Natl Acad Sci USA. 2011;108:4334–4339. doi: 10.1073/pnas.1017150108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Brangwynne CP, Tompa P, Pappu RV. Polymer physics of intracellular phase transitions. Nat Phys. 2015;11:899–904. doi: 10.1038/nphys3532. [DOI] [Google Scholar]
- 6.Kato M, et al. Cell-free formation of RNA granules: low complexity sequence domains form dynamic fibers within hydrogels. Cell. 2012;149:753–767. doi: 10.1016/j.cell.2012.04.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Molliex A, et al. Phase separation by low complexity domains promotes stress granule assembly and drives pathological fibrillization. Cell. 2015;163:123–133. doi: 10.1016/j.cell.2015.09.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Patel A, et al. A liquid-to-solid phase transition of the ALS protein FUS accelerated by disease mutation. Cell. 2015;162:1066–1077. doi: 10.1016/j.cell.2015.07.047. [DOI] [PubMed] [Google Scholar]
- 9.Lin Y, Protter DS, Rosen MK, Parker R. Formation and maturation of phase-separated liquid droplets by RNA-binding proteins. Mol Cell. 2015;60:208–219. doi: 10.1016/j.molcel.2015.08.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Uversky VN. Intrinsically disordered proteins in overcrowded milieu: membrane-less organelles, phase separation, and intrinsic disorder. Curr Opin Struct Biol. 2017;44:18–30. doi: 10.1016/j.sbi.2016.10.015. [DOI] [PubMed] [Google Scholar]
- 11.Jain S, et al. ATPase-modulated stress granules contain a diverse proteome and substructure. Cell. 2016;164:487–498. doi: 10.1016/j.cell.2015.12.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Feric M, et al. Coexisting liquid phases underlie nucleolar subcompartments. Cell. 2016;165:1686–1697. doi: 10.1016/j.cell.2016.04.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Murakami T, et al. ALS/FTD mutation-induced phase transition of FUS liquid droplets and reversible hydrogels into irreversible hydrogels impairs RNP granule function. Neuron. 2015;88:678–690. doi: 10.1016/j.neuron.2015.10.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Shin Y, et al. Spatiotemporal control of intracellular phase transitions using light-activated optoDroplets. Cell. 2017;168:159.e114–171.e114. doi: 10.1016/j.cell.2016.11.054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Li YR, King OD, Shorter J, Gitler AD. Stress granules as crucibles of ALS pathogenesis. J Cell Biol. 2013;201:361–372. doi: 10.1083/jcb.201302044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ramaswami M, Taylor JP, Parker R. Altered ribostasis: RNA-protein granules in degenerative disorders. Cell. 2013;154:727–736. doi: 10.1016/j.cell.2013.07.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Boeynaems S, Bogaert E, Van Damme P, Van Den Bosch L. Inside out: the role of nucleocytoplasmic transport in ALS and FTLD. Acta Neuropathol. 2016;132:159–173. doi: 10.1007/s00401-016-1586-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.DeJesus-Hernandez M, et al. Expanded GGGGCC hexanucleotide repeat in noncoding region of C9ORF72 causes chromosome 9p-linked FTD and ALS. Neuron. 2011;72:245–256. doi: 10.1016/j.neuron.2011.09.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Renton AE, et al. A hexanucleotide repeat expansion in C9ORF72 is the cause of chromosome 9p21-linked ALS-FTD. Neuron. 2011;72:257–268. doi: 10.1016/j.neuron.2011.09.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Boeynaems S, et al. Drosophila screen connects nuclear transport genes to DPR pathology in c9ALS/FTD. Sci Rep. 2016;6:20877. doi: 10.1038/srep20877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Jovicic A, et al. Modifiers of C9orf72 dipeptide repeat toxicity connect nucleocytoplasmic transport defects to FTD/ALS. Nat Neurosci. 2015;18:1226–1229. doi: 10.1038/nn.4085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kwon I, et al. Poly-dipeptides encoded by the C9orf72 repeats bind nucleoli, impede RNA biogenesis, and kill cells. Science. 2014;345:1139–1145. doi: 10.1126/science.1254917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Mizielinska S, et al. C9orf72 repeat expansions cause neurodegeneration in Drosophila through arginine-rich proteins. Science. 2014;345:1192–1194. doi: 10.1126/science.1256800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Tran H, et al. Differential toxicity of nuclear RNA foci versus dipeptide repeat proteins in a Drosophila Model of C9ORF72 FTD/ALS. Neuron. 2015;87:1207–1214. doi: 10.1016/j.neuron.2015.09.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wen XM, et al. Antisense proline–arginine RAN dipeptides linked to C9ORF72-ALS/FTD form toxic nuclear aggregates that initiate in vitro and in vivo neuronal death. Neuron. 2014;84:1213–1225. doi: 10.1016/j.neuron.2014.12.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Boeynaems S, De Decker M, Tompa P, Van Den Bosch L. Arginine-rich peptides can actively mediate liquid–liquid phase separation. Bio-protocol. 2017;7:e2525. doi: 10.21769/BioProtoc.2525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Boeynaems S, et al. Phase separation of C9orf72 dipeptide repeats perturbs stress granule dynamics. Mol Cell. 2017;65:1044.e1045–1055.e1045. doi: 10.1016/j.molcel.2017.02.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.King OD, Gitler AD, Shorter J. The tip of the iceberg: RNA-binding proteins with prion-like domains in neurodegenerative disease. Brain Res. 2012;1462:61–80. doi: 10.1016/j.brainres.2012.01.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Mitrea DM, et al. Nucleophosmin integrates within the nucleolus via multi-modal interactions with proteins displaying R-rich linear motifs and rRNA. Elife. 2016;5:e13571. doi: 10.7554/eLife.13571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hernandez-Verdun D. Assembly and disassembly of the nucleolus during the cell cycle. Nucleus. 2011;2:189–194. doi: 10.4161/nucl.2.3.16246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Chan CY, et al. Purinosome formation as a function of the cell cycle. Proc Natl Acad Sci USA. 2015;112:1368–1373. doi: 10.1073/pnas.1423009112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Conduit PT, Wainman A, Raff JW. Centrosome function and assembly in animal cells. Nat Rev Mol Cell Biol. 2015;16:611–624. doi: 10.1038/nrm4062. [DOI] [PubMed] [Google Scholar]
- 33.Woodruff JB, et al. The centrosome is a selective condensate that nucleates microtubules by concentrating tubulin. Cell. 2017;169:1066.e1010–1077.e1010. doi: 10.1016/j.cell.2017.05.028. [DOI] [PubMed] [Google Scholar]
- 34.Ma Y, Kanakousaki K, Buttitta L. How the cell cycle impacts chromatin architecture and influences cell fate. Front Genet. 2015;6:19. doi: 10.3389/fgene.2015.00019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Larson AG, et al. Liquid droplet formation by HP1alpha suggests a role for phase separation in heterochromatin. Nature. 2017;547:236–240. doi: 10.1038/nature22822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Strom AR, et al. Phase separation drives heterochromatin domain formation. Nature. 2017;547:241–245. doi: 10.1038/nature22989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Cuylen S, et al. Ki-67 acts as a biological surfactant to disperse mitotic chromosomes. Nature. 2016;535:308–312. doi: 10.1038/nature18610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Samwer M, et al. DNA cross-bridging shapes a single nucleus from a set of mitotic chromosomes. Cell. 2017;170:956.e923–972.e923. doi: 10.1016/j.cell.2017.07.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Yedidi RS, Fatehi AK, Enenkel C. Proteasome dynamics between proliferation and quiescence stages of Saccharomyces cerevisiae. Crit Rev Biochem Mol Biol. 2016;51:497–512. doi: 10.1080/10409238.2016.1230087. [DOI] [PubMed] [Google Scholar]
- 40.Su X, et al. Phase separation of signaling molecules promotes T cell receptor signal transduction. Science. 2016;352:595–599. doi: 10.1126/science.aad9964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Grabocka E, Bar-Sagi D. Mutant KRAS enhances tumor cell fitness by upregulating stress granules. Cell. 2016;167:1803.e1812–1813.e1812. doi: 10.1016/j.cell.2016.11.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Somasekharan SP, et al. YB-1 regulates stress granule formation and tumor progression by translationally activating G3BP1. J Cell Biol. 2015;208:913–929. doi: 10.1083/jcb.201411047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Adriaens C, et al. p53 induces formation of NEAT1 lncRNA-containing paraspeckles that modulate replication stress response and chemosensitivity. Nat Med. 2016;22:861–868. doi: 10.1038/nm.4135. [DOI] [PubMed] [Google Scholar]
- 44.Xu J, et al. Gain of function of mutant p53 by coaggregation with multiple tumor suppressors. Nat Chem Biol. 2011;7:285–295. doi: 10.1038/nchembio.546. [DOI] [PubMed] [Google Scholar]
- 45.Soragni A, et al. A designed inhibitor of p53 aggregation rescues p53 tumor suppression in ovarian carcinomas. Cancer Cell. 2016;29:90–103. doi: 10.1016/j.ccell.2015.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Rabbitts TH, Forster A, Larson R, Nathan P. Fusion of the dominant-negative transcription regulator CHOP with a novel gene FUS by translocation t(12; 16) in malignant liposarcoma. Nat Genet. 1993;4:175–180. doi: 10.1038/ng0693-175. [DOI] [PubMed] [Google Scholar]
- 47.Erkizan HV, et al. A small molecule blocking oncogenic protein EWS–FLI1 interaction with RNA helicase A inhibits growth of Ewing’s sarcoma. Nat Med. 2009;15:U750–U758. doi: 10.1038/nm.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Fujimura Y, et al. The EWS–ATF-1 gene involved in malignant melanoma of soft parts with t(12;22) chromosome translocation, encodes a constitutive transcriptional activator. Oncogene. 1996;12:159–167. [PubMed] [Google Scholar]
- 49.Bischof D, Pulford K, Mason DY, Morris SW. Role of the nucleophosmin (NPM) portion of the non-Hodgkin’s lymphoma-associated NPM-anaplastic lymphoma kinase fusion protein in oncogenesis. Mol Cell Biol. 1997;17:2312–2325. doi: 10.1128/MCB.17.4.2312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Gough SM, Slape CI, Aplan PD. NUP98 gene fusions and hematopoietic malignancies: common themes and new biologic insights. Blood. 2011;118:6247–6257. doi: 10.1182/blood-2011-07-328880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Burke KA, Janke AM, Rhine CL, Fawzi NL. Residue-by-residue view of in vitro FUS granules that bind the C-terminal domain of RNA polymerase II. Mol Cell. 2015;60:231–241. doi: 10.1016/j.molcel.2015.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Hegyi H, Buday L, Tompa P. Intrinsic structural disorder confers cellular viability on oncogenic fusion proteins. PLoS Comput Biol. 2009;5:e1000552. doi: 10.1371/journal.pcbi.1000552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Latysheva NS, et al. Molecular principles of gene fusion mediated rewiring of protein interaction networks in cancer. Mol Cell. 2016;63:579–592. doi: 10.1016/j.molcel.2016.07.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Kwon I, et al. Phosphorylation-regulated binding of RNA polymerase II to fibrous polymers of low-complexity domains. Cell. 2013;155:1049–1060. doi: 10.1016/j.cell.2013.10.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Boulay G, et al. Cancer-specific retargeting of BAF complexes by a Prion-like domain. Cell. 2017;171:163–178. doi: 10.1016/j.cell.2017.07.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Schwartz JC, Wang X, Podell ER, Cech TR. RNA seeds higher-order assembly of FUS protein. Cell Rep. 2013;5:918–925. doi: 10.1016/j.celrep.2013.11.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Amin MA, et al. Fibrillarin, a nucleolar protein, is required for normal nuclear morphology and cellular growth in HeLa cells. Biochem Biophys Res Commun. 2007;360:320–326. doi: 10.1016/j.bbrc.2007.06.092. [DOI] [PubMed] [Google Scholar]
- 58.Grisendi S, Mecucci C, Falini B, Pandolfi PP. Nucleophosmin and cancer. Nat Rev Cancer. 2006;6:493–505. doi: 10.1038/nrc1885. [DOI] [PubMed] [Google Scholar]
- 59.Contreras V, Richardson MA, Hao E, Keiper BD. Depletion of the cap-associated isoform of translation factor eIF4G induces germline apoptosis in C-elegans. Cell Death Differ. 2008;15:1232–1242. doi: 10.1038/cdd.2008.46. [DOI] [PubMed] [Google Scholar]
- 60.Ward CL, et al. A loss of FUS/TLS function leads to impaired cellular proliferation. Cell Death Dis. 2014;5:e1572. doi: 10.1038/cddis.2014.508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Duggimpudi S, Larsson E, Nabhani S, Borkhardt A, Hoell JI. The cell cycle regulator CCDC6 is a key target of RNA-binding protein EWS. PLoS ONE. 2015;10:e0119066. doi: 10.1371/journal.pone.0119066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Yu C, et al. Oral squamous cancer cell exploits hnRNP A1 to regulate cell cycle and proliferation. J Cell Physiol. 2015;230:2252–2261. doi: 10.1002/jcp.24956. [DOI] [PubMed] [Google Scholar]
- 63.Lee K, Suzuki H, Aiso S, Matsuoka M. Overexpression of TDP-43 causes partially p53-dependent G2/M arrest and p53-independent cell death in HeLa cells. Neurosci Lett. 2012;506:271–276. doi: 10.1016/j.neulet.2011.11.021. [DOI] [PubMed] [Google Scholar]
- 64.Conicella AE, Zerze GH, Mittal J, Fawzi NL. ALS mutations disrupt phase separation mediated by alpha-helical structure in the TDP-43 low-complexity C-terminal domain. Structure. 2016;24:1537–1549. doi: 10.1016/j.str.2016.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Reyes R, Alcalde J, Izquierdo JM. Depletion of T-cell intracellular antigen (TIA)-proteins promotes cell proliferation. Genome Biol. 2009;10:R87. doi: 10.1186/gb-2009-10-8-r87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Sluder G. Two-way traffic: centrosomes and the cell cycle. Nat Rev Mol Cell Bio. 2005;6:743–748. doi: 10.1038/nrm1712. [DOI] [PubMed] [Google Scholar]
- 67.Woodruff JB, et al. Regulated assembly of a supramolecular centrosome scaffold in vitro. Science. 2015;348:808–812. doi: 10.1126/science.aaa3923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Yamaoka T, et al. Amidophosphoribosyltransferase limits the rate of cell growth-linked de novo purine biosynthesis in the presence of constant capacity of salvage purine biosynthesis. J Biol Chem. 1997;272:17719–17725. doi: 10.1074/jbc.272.28.17719. [DOI] [PubMed] [Google Scholar]
- 69.Kyoung M, Russell SJ, Kohnhorst CL, Esemoto NN, An S. Dynamic architecture of the purinosome involved in human de novo purine biosynthesis. Biochemistry. 2015;54:870–880. doi: 10.1021/bi501480d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Kalverda B, Pickersgill H, Shloma VV, Fornerod M. Nucleoporins directly stimulate expression of developmental and cell-cycle genes inside the nucleoplasm. Cell. 2010;140:U360–U381. doi: 10.1016/j.cell.2010.01.011. [DOI] [PubMed] [Google Scholar]
- 71.Schmidt HB, Gorlich D. Nup98 FG domains from diverse species spontaneously phase-separate into particles with nuclear pore-like permselectivity. Elife. 2015;4:e04251. doi: 10.7554/eLife.04251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Vivar OI, et al. IFT20 controls LAT recruitment to the immune synapse and T-cell activation in vivo. Proc Natl Acad Sci USA. 2016;113:386–391. doi: 10.1073/pnas.1513601113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Liu J, et al. Functional proteomic analysis of promyelocytic leukaemia nuclear bodies in irradiation-induced MCF-7 cells. J Biochem. 2010;148:659–667. doi: 10.1093/jb/mvq105. [DOI] [PubMed] [Google Scholar]
- 74.Frey S, Richter RP, Goerlich D. FG-rich repeats of nuclear pore proteins form a three-dimensional meshwork with hydrogel-like properties. Science. 2006;314:815–817. doi: 10.1126/science.1132516. [DOI] [PubMed] [Google Scholar]
- 75.Andersen JS, et al. Nucleolar proteome dynamics. Nature. 2005;433:77–83. doi: 10.1038/nature03207. [DOI] [PubMed] [Google Scholar]
- 76.Zwicker D, Decker M, Jaensch S, Hyman AA, Julicher F. Centrosomes are autocatalytic droplets of pericentriolar material organized by centrioles. Proc Natl Acad Sci USA. 2014;111:E2636–E2645. doi: 10.1073/pnas.1404855111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Yasuda K, et al. The RNA-binding protein Fus directs translation of localized mRNAs in APC-RNP granules. J Cell Biol. 2013;203:737–746. doi: 10.1083/jcb.201306058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.An SG, Kumar R, Sheets ED, Benkovic SJ. Reversible compartmentalization of de novo purine biosynthetic complexes in living cells. Science. 2008;320:103–106. doi: 10.1126/science.1152241. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Mass spectrometry dataset was previously published and is the data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifier PRIDE: PXD005509.